
Stochastic Processes Dominate Soil Microbial Community Assembly during the Restoration of Degraded Karst Forests
 and
and 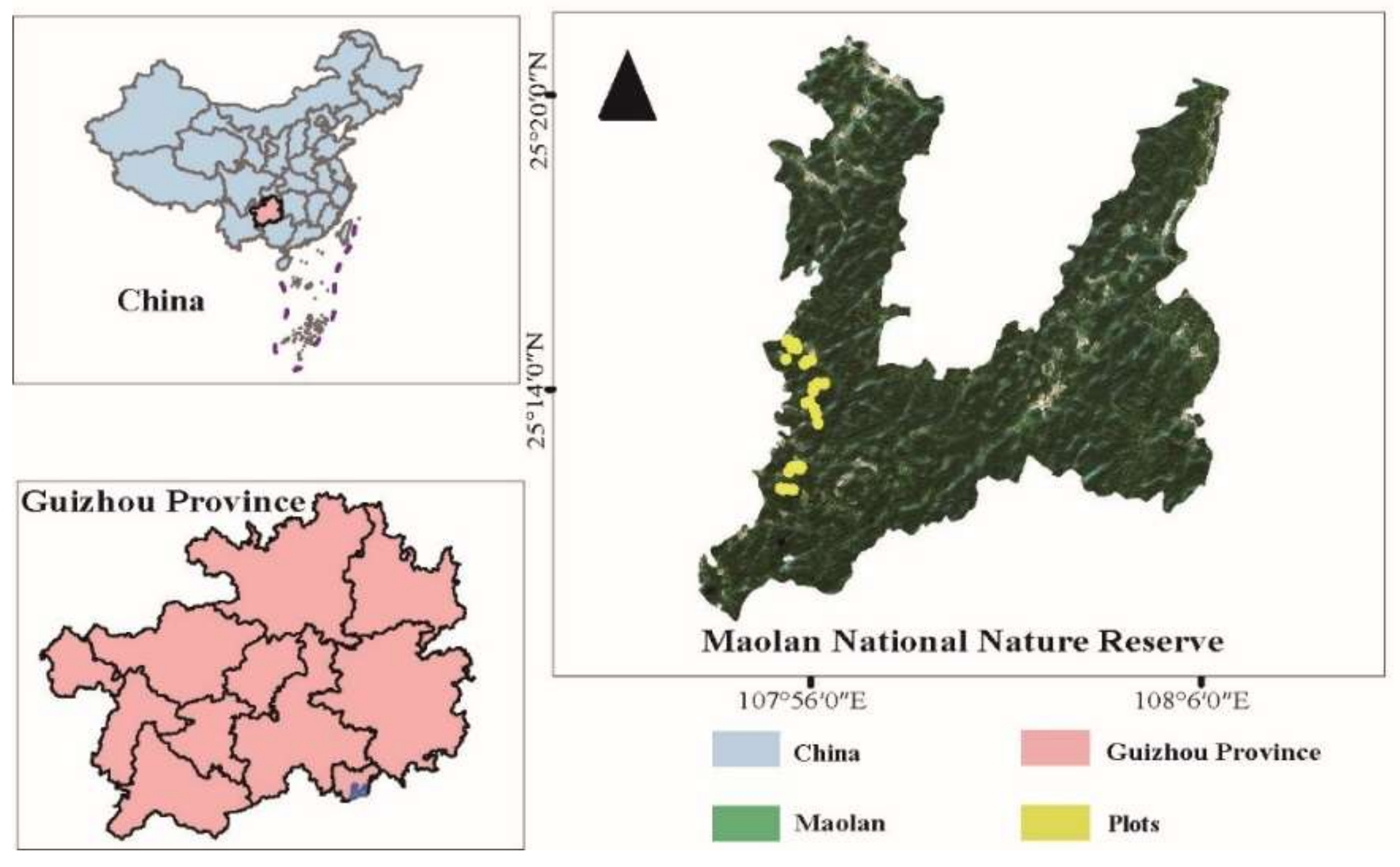{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Area
2.2. Data Acquisition
2.2.1. Demarcation of Plots and Sample Collection
2.2.2. DNA Extraction, Library Construction, and Metagenomic Sequencing Analysis
2.2.3. Sequence Quality Control and Genome Assembly
2.2.4. Gene Prediction, Taxonomy, and Species Annotation
2.2.5. Calculating Species Diversity
2.2.6. Determination of Soil Physical and Chemical Properties and Calculation of Soil Versatility
2.3. Statistical Analysis
3. Results
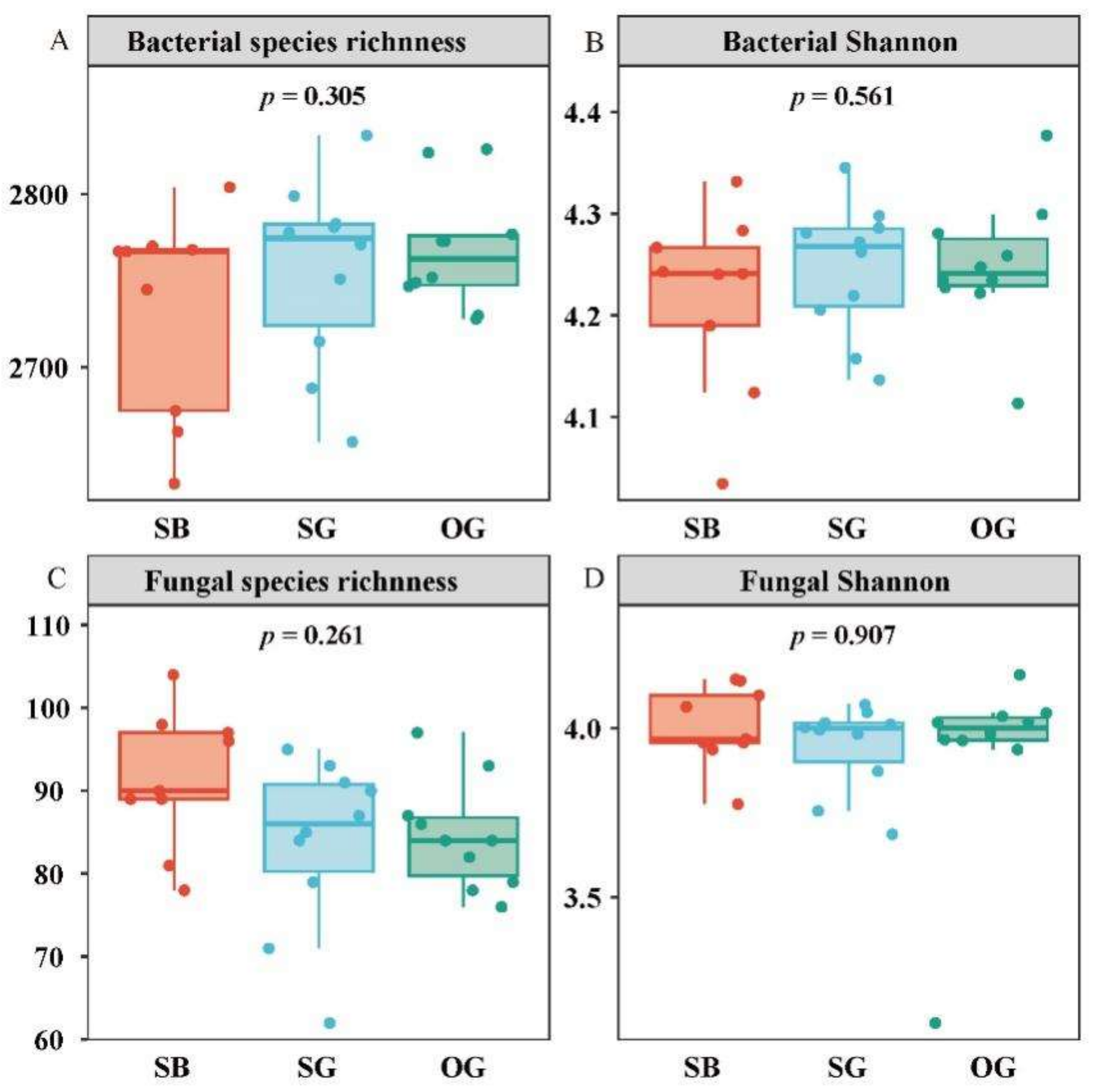
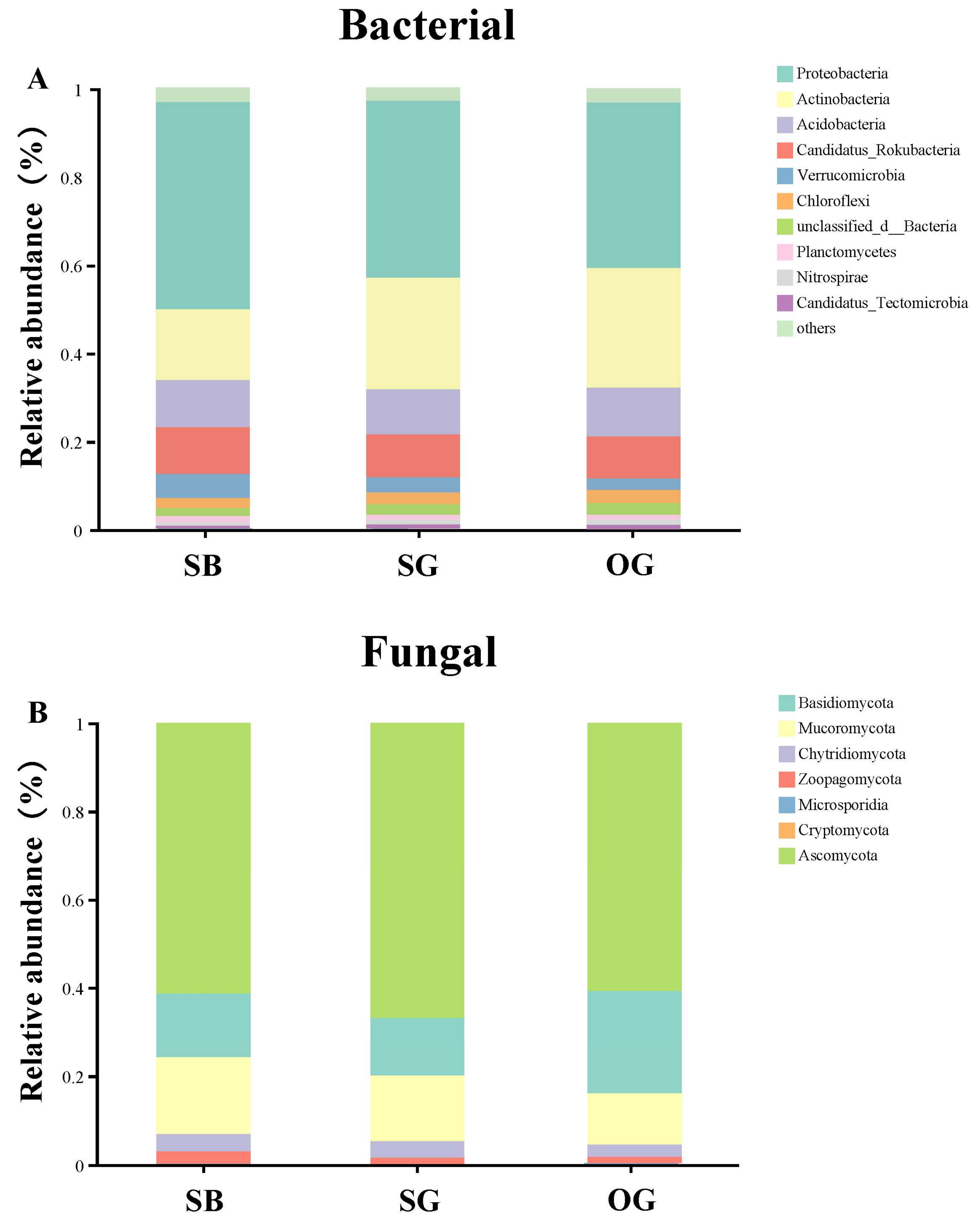
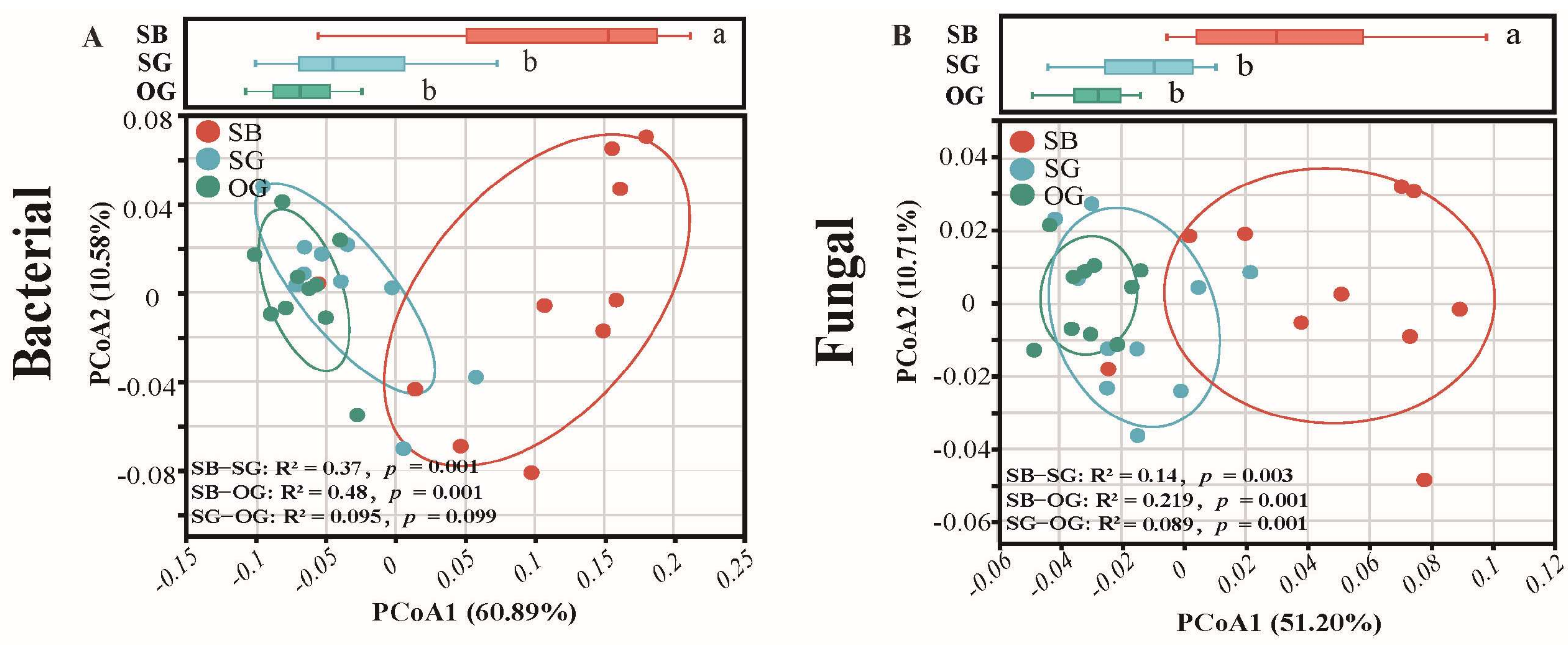
3.1. Trends in SOIL Microbial Species Diversity and Community Composition during Succession
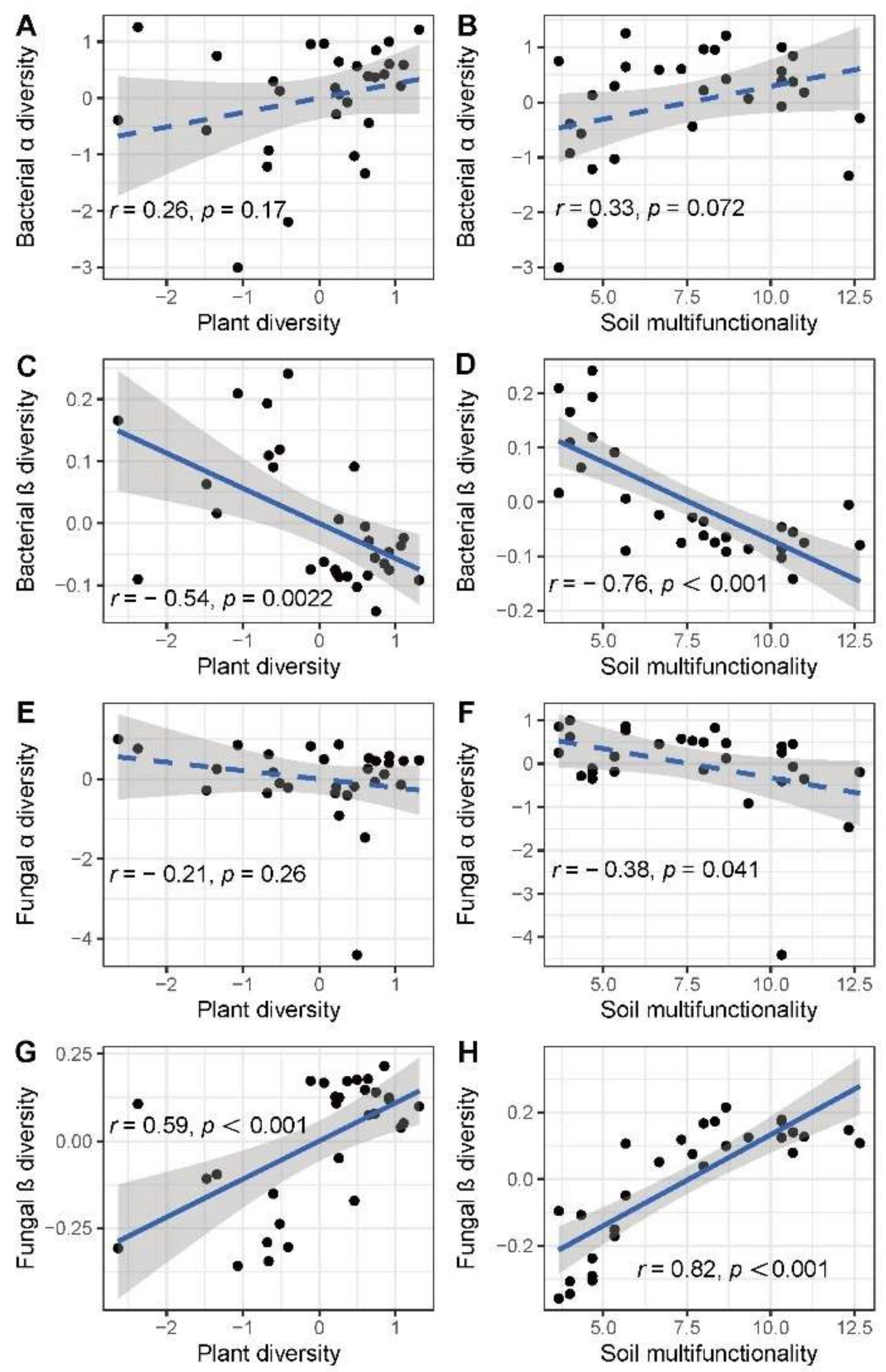
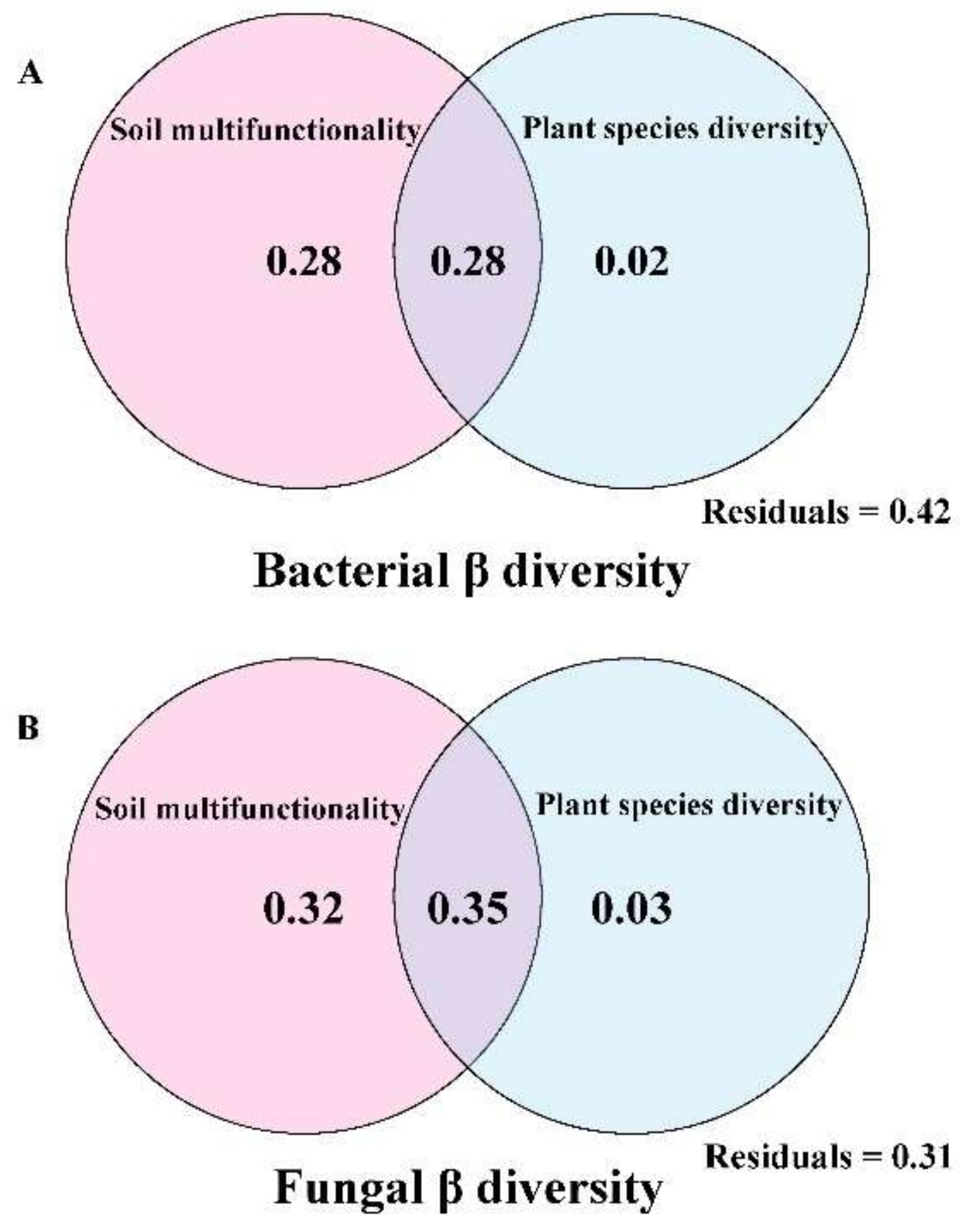
3.2. Plant Species Diversity and Soil Multifunctionality as Drivers of Soil Microbes
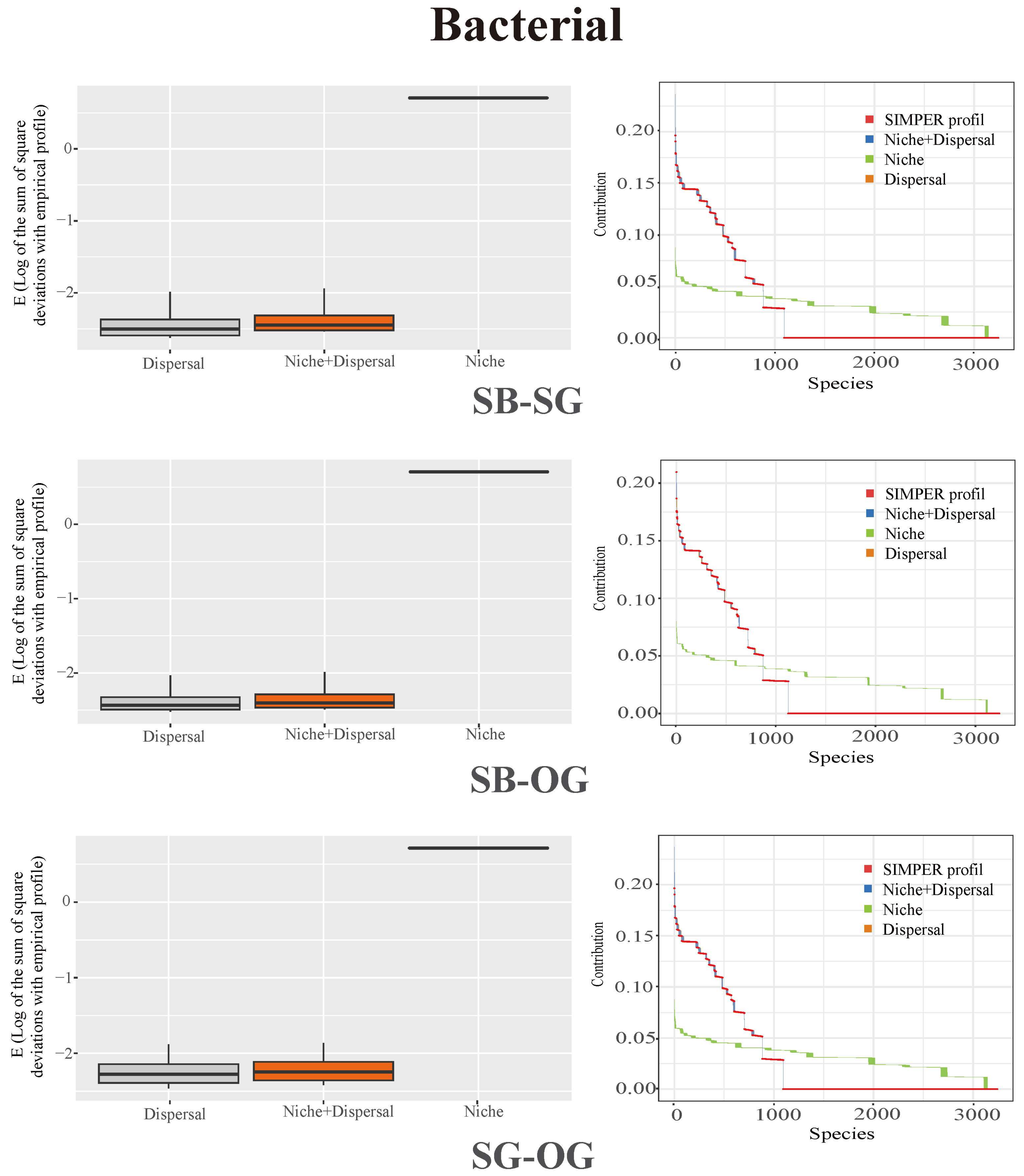
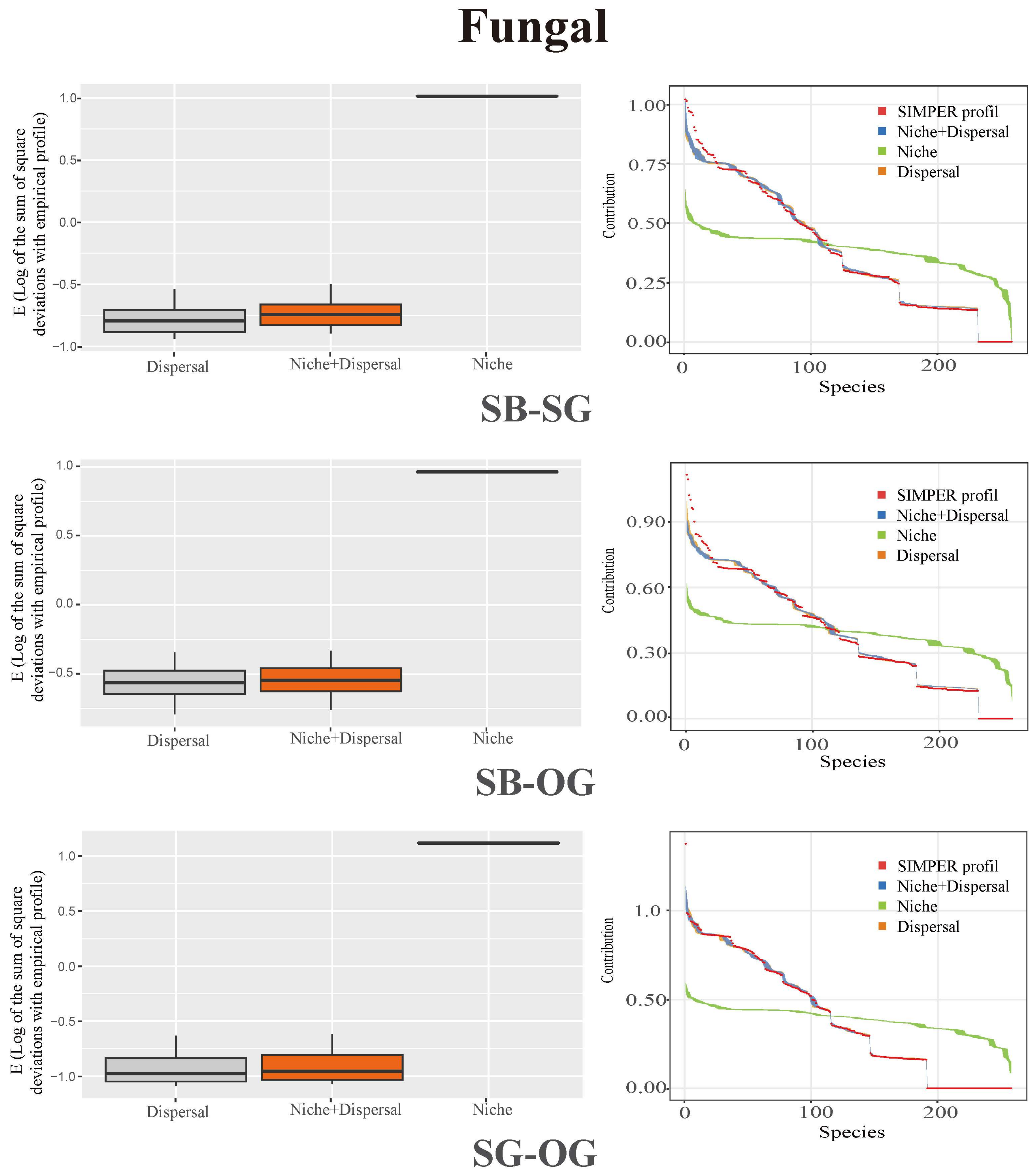
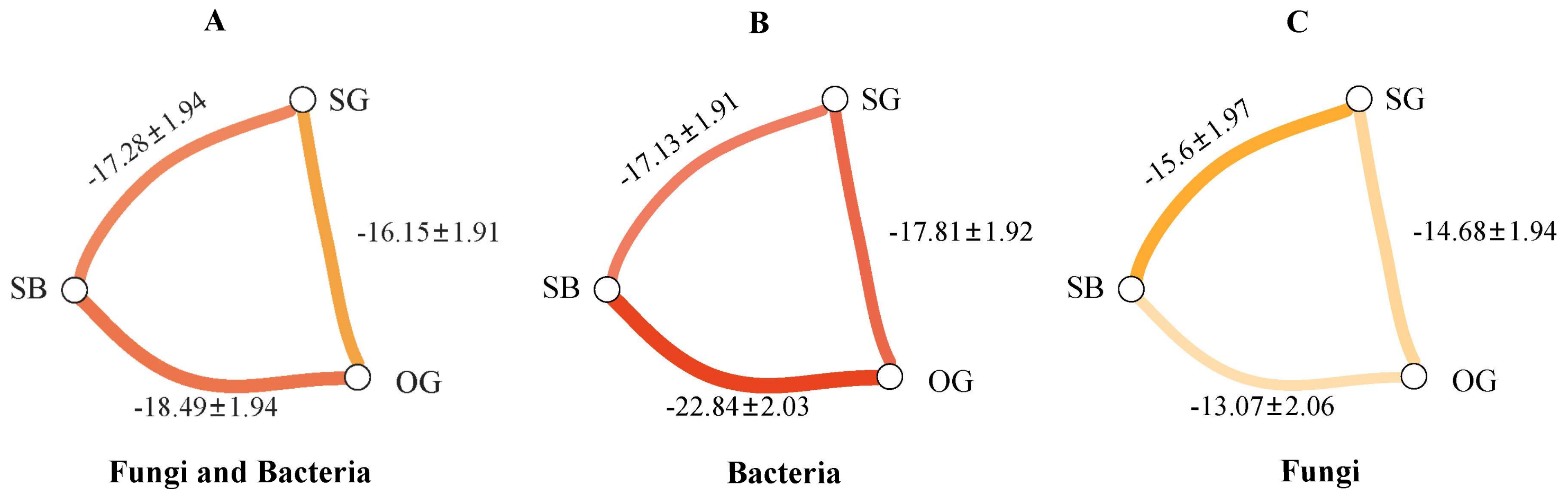
3.3. Soil Microbial Community Assembly Process
4. Discussion
4.1. Soil Microbial α- and β-Diversity Indices Show Different Trends during Restoration Processes
4.2. Soil Multifunctionality Drives the β-Diversity of Soil Microbial Communities
4.3. Stochastic Processes Dominate the Assembly of Soil Microbial Community during the Restoration of Degraded Karst Forests
5. Conclusions and Suggestions for Future Research
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hu, J.; Wei, Z.; Kowalchuk, G.A.; Xu, Y.; Shen, Q.; Jousset, A. Rhizosphere microbiome functional diversity and pathogen invasion resistance build up during plant development. Environ. Microbiol. 2020, 22, 5005–5018. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, W.; Kou, Y.; Liu, Y.; He, H.; Liu, Q. Secondary forest succession drives differential responses of bacterial communities and interactions rather than bacterial functional groups in the rhizosphere and bulk soils in a subalpine region. Plant Soil 2023, 484, 293–312. [Google Scholar] [CrossRef]
- Song, C.; Wu, J.; Lu, Y.; Shen, Q.; He, J.; Huang, Q.; Jia, Z. Advances of Soil Microbiology in the Last Decade in China. Adv. Earth Sci. 2013, 28, 1087–1105. [Google Scholar]
- Zhao, C.; Long, J.; Liao, H.; Zheng, C.; Li, J.; Liu, L.; Zhang, M. Dynamics of soil microbial communities following vegetation succession in a karst mountain ecosystem, Southwest China. Sci. Rep. 2019, 9, 2160. [Google Scholar] [CrossRef]
- van der Heijden, M.G.A.; Bardgett, R.D.; van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, S.; Martin, A.P.; Sousa-Pinto, I.; Arenas, F. Biodiversity effects on macroalgal productivity: Exploring the roles of richness, evenness and species traits. Mar. Ecol. Prog. Ser. 2016, 562, 79–91. [Google Scholar] [CrossRef]
- Mori, A.S. Environmental controls on the causes and functional consequences of tree species diversity. J. Ecol. 2018, 106, 113–125. [Google Scholar] [CrossRef]
- Shen, C.; He, J.; Ge, Y. Seasonal dynamics of soil microbial diversity and functions along elevations across the treeline. Sci. Total Environ. 2021, 794, 148644. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Wang, C.; Luo, Y. Meta-analysis of the impacts of global change factors on soil microbial diversity and functionality. Nat. Commun. 2020, 11, 3072. [Google Scholar] [CrossRef]
- Lozano, Y.M.; Hortal, S.; Armas, C.; Pugnaire, F.I. Interactions among soil, plants, and microorganisms drive secondary succession in a dry environment. Soil Biol. Biochem. 2014, 78, 298–306. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C.; Luo, Y. Effects of forest degradation on microbial communities and soil carbon cycling: A global meta-analysis. Glob. Ecol. Biogeogr. 2018, 27, 110–124. [Google Scholar] [CrossRef]
- Liu, J.; Jia, X.; Yan, W.; Zho, Y.; Shangguan, Z. Changes in soil microbial community structure during long-term secondary succession. Land Degrad. Dev. 2020, 31, 1151–1166. [Google Scholar] [CrossRef]
- Li, Y.; Liu, X.; Yin, Z.; Chen, H.; Cai, X.; Xie, Y.; Wang, S.; Lian, B. Changes in soil microbial communities from exposed rocks to arboreal rhizosphere during vegetation succession in a karst mountainous ecosystem. J. Plant Interact. 2021, 16, 550–563. [Google Scholar] [CrossRef]
- He, F.; Yang, B.; Wang, H.; Yan, Q.; Cao, Y.; He, X. Changes in composition and diversity of fungal communities along Quercus mongolica forests developments in Northeast China. Appl. Soil Ecol. 2016, 100, 162–171. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, J.; Chen, J.; Li, G.; He, H.; Huang, T.; Ding, J. Changes in diversity patterns and assembly processes of soil nematode communities during forest secondary succession on the Loess Plateau. For. Ecol. Manag. 2023, 529, 120734. [Google Scholar] [CrossRef]
- Kang, Y.; Wu, H.; Zhang, Y.; Wu, Q.; Guan, Q.; Lu, K.; Lin, Y. Differential distribution patterns and assembly processes of soil microbial communities under contrasting vegetation types at distinctive altitudes in the Changbai Mountain. Front. Microbiol. 2023, 14, 1152818. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Du, H.; Zeng, F.; Song, T.; Peng, W. Diminished rhizosphere and bulk soil microbial abundance and diversity across succession stages in Karst area, southwest China. Appl. Soil Ecol. 2021, 158, 103799. [Google Scholar] [CrossRef]
- Shang, R.; Li, S.; Huang, X.; Liu, W.; Lang, X.; Su, J. Effects of soil properties and plant diversity on soil microbial community composition and diversity during secondary succession. Forests 2021, 12, 805. [Google Scholar] [CrossRef]
- Martinez-Garcia, L.B.; Pugnaire, F.I. Arbuscular mycorrhizal fungi host preference and site effects in two plant species in a semiarid environment. Appl. Soil Ecol. 2011, 48, 313–317. [Google Scholar] [CrossRef]
- Chen, Y.; Xi, J.; Xiao, M.; Wang, S.; Chen, W.; Liu, F.; Shao, Y.; Yuan, Z. Soil fungal communities show more specificity than bacteria for plant species composition in a temperate forest in China. BMC Microbiol. 2022, 22, 208. [Google Scholar] [CrossRef]
- Yuan, Y.; Si, G.; Wang, J.; Luo, T.; Zhang, G. Bacterial community in alpine grasslands along an altitudinal gradient on the Tibetan Plateau. FEMS Microbiol. Ecol. 2014, 87, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, G.; Xue, S.; Wang, G. Soil bacterial community dynamics reflect changes in plant community and soil properties during the secondary succession of abandoned farmland in the Loess Plateau. Soil Biol. Biochem. 2016, 97, 40–49. [Google Scholar] [CrossRef]
- Zhou, J.; Deng, Y.; Zhang, P.; Xue, K.; Liang, Y.; Van Nostrand, J.D.; Yang, Y.; He, Z.; Wu, L.; Stahl, D.A.; et al. Stochasticity, succession, and environmental perturbations in a fluidic ecosystem. Proc. Natl. Acad. Sci. USA 2014, 111, E836–E845. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Ning, D. Stochastic community assembly: Does it matter in microbial ecology? Microbiol. Mol. Biol. Rev. 2017, 81, E00002-17. [Google Scholar] [CrossRef]
- Dini-Andreote, F.; Stegen, J.C.; van Elsas, J.D.; Salles, J.F. Disentangling mechanisms that mediate the balance between stochastic and deterministic processes in microbial succession. Proc. Natl. Acad. Sci. USA 2015, 112, E1326–E1332. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhu, K.; Krause, S.M.B.; Li, S.; Wang, X.; Zhang, Z.; Shen, M.; Yang, Q.; Lian, J.; Wang, X.; et al. Changes in assembly processes of soil microbial communities during secondary succession in two subtropical forests. Soil Biol. Biochem. 2021, 154, 108144. [Google Scholar] [CrossRef]
- Chase, J.M.; Myers, J.A. Disentangling the importance of ecological niches from stochastic processes across scales. Philos. Trans. R. Soc. B-Biol. Sci. 2011, 366, 2351–2363. [Google Scholar] [CrossRef]
- Stegen, J.C.; Lin, X.; Fredrickson, J.K.; Chen, X.; Kennedy, D.W.; Murray, C.J.; Rockhold, M.L.; Konopka, A. Quantifying community assembly processes and identifying features that impose them. ISME J. 2013, 7, 2069–2079. [Google Scholar] [CrossRef]
- Adler, P.B.; Hillerislambers, J.; Levine, J.M. A niche for neutrality. Ecol. Lett. 2007, 10, 95–104. [Google Scholar] [CrossRef]
- Gravel, D.; Canham, C.D.; Beaudet, M.; Messier, C. Reconciling niche and neutrality: The continuum hypothesis. Ecol. Lett. 2006, 9, 399–409. [Google Scholar] [CrossRef]
- Vilmi, A.; Gibert, C.; Escarguel, G.; Happonen, K.; Heino, J.; Jamoneau, A.; Passy, S.I.; Picazo, F.; Soininen, J.; Tison-Rosebery, J.; et al. Dispersal-niche continuum index: A new quantitative metric for assessing the relative importance of dispersal versus niche processes in community assembly. Ecography 2021, 44, 370–379. [Google Scholar] [CrossRef]
- Gibert, C.; Escarguel, G. PER-SIMPER-A new tool for inferring community assembly processes from taxon occurrences. Glob. Ecol. Biogeogr. 2019, 28, 374–385. [Google Scholar] [CrossRef]
- Gibert, C.; Shenbrot, G.I.; Stanko, M.; Khokhlova, I.S.; Krasnov, B.R. Dispersal-based versus niche-based processes as drivers of flea species composition on small mammalian hosts: Inferences from species occurrences at large and small scales. Oecologia 2021, 197, 471–484. [Google Scholar] [CrossRef]
- Graham, E.B.; Stegen, J.C. Dispersal-Based Microbial Community Assembly Decreases Biogeochemical Function. Processes 2017, 5, 65. [Google Scholar] [CrossRef]
- Bach, E.M.; Baer, S.G.; Meyer, C.K.; Six, J. Soil texture affects soil microbial and structural recovery during grassland restoration. Soil Biol. Biochem. 2010, 42, 2182–2191. [Google Scholar] [CrossRef]
- Condit, R.; Sukumar, R.; Hubbell, S.P.; Foster, R.B. Predicting population trends from size distributions: A direct test in a tropical tree community. Am. Nat. 1998, 152, 495–509. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, C.-M.; Luo, R.; Sadakane, K.; Lam, T.-W. MEGAHIT: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Delgado, E.O.; Winemiller, K.O.; Villa-Navarro, F.A. Local environmental factors influence beta-diversity patterns of tropical fish assemblages more than spatial factors. Ecology 2020, 101, e02940. [Google Scholar] [CrossRef]
- Weiher, E.; Freund, D.; Bunton, T.; Stefanski, A.; Lee, T.; Bentivenga, S. Advances, challenges and a developing synthesis of ecological community assembly theory. Philos. Trans. R. Soc. B-Biol. Sci. 2011, 366, 2403–2413. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Cai, X.; Liu, X.; Wang, J.; Cheng, S.; Zhang, X.; Li, D.; Li, M. Soil microbial population dynamics along a chronosequence of moist evergreen broad-leaved forest succession in southwestern China. J. Mt. Sci. 2010, 7, 327–338. [Google Scholar] [CrossRef]
- Wang, X.; Long, J.; Li, J.; Liu, L.; Liao, H.; Li, Y.; Yang, R. Diversity of soil Eukaryotic microbes in different succession stages in Maolan karst forest of Guizhou. Environ Sci. 2020, 41, 4314–4321. [Google Scholar] [CrossRef]
- Poosakkannu, A.; Nissinen, R.; Mannisto, M.; Kytoviita, M.-M. Microbial community composition but not diversity changes along succession in arctic sand dunes. Environ. Microbiol. 2017, 19, 698–709. [Google Scholar] [CrossRef]
- Yu, J.; Yin, Q.; Niu, J.; Yan, Z.; Wang, H.; Wang, Y.; Chen, D. Consistent effects of vegetation patch type on soil microbial communities across three successional stages in a desert ecosystem. Land Degrad. Dev. 2022, 33, 1552–1563. [Google Scholar] [CrossRef]
- Dini-Andreote, F.; Brossi, M.J.d.L.; van Elsas, J.D.; Salles, J.F. Reconstructing the genetic potential of the microbially-mediated nitrogen cycle in a salt marsh ecosystem. Front. Microbiol. 2016, 7, 902. [Google Scholar] [CrossRef]
- Bastida, F.; Eldridge, D.J.; Garcia, C.; Kenny Png, G.; Bardgett, R.D.; Delgado-Baquerizo, M. Soil microbial diversity-biomass relationships are driven by soil carbon content across global biomes. ISME J. 2021, 15, 2081–2091. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Chen, D.; Saleem, M.; Wang, B.; Hu, S.; Delgado-Baquerizo, M.; Bai, Y. Rare soil microbial taxa regulate the negative effects of land degradation drivers on soil organic matter decomposition. J. Appl. Ecol. 2021, 58, 1658–1669. [Google Scholar] [CrossRef]
- Qu, Q.; Zhang, Z.; Peijnenburg, W.J.G.M.; Liu, W.; Lu, T.; Hu, B.; Chen, J.; Chen, J.; Lin, Z.; Qian, H. Rhizosphere microbiome assembly and its impact on plant growth. J. Agric. Food Chem. 2020, 68, 5024–5038. [Google Scholar] [CrossRef]
- Zhalnina, K.; Louie, K.B.; Hao, Z.; Mansoori, N.; da Rocha, U.N.; Shi, S.; Cho, H.; Karaoz, U.; Loque, D.; Bowen, B.P.; et al. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nat. Microbiol. 2018, 3, 470–480. [Google Scholar] [CrossRef]
- Vives-Peris, V.; de Ollas, C.; Gomez-Cadenas, A.; Perez-Clemente, R.M. Root exudates: From plant to rhizosphere and beyond. Plant Cell Rep. 2020, 39, 3–17. [Google Scholar] [CrossRef]
- Yang, N.; Ji, L.; Salahuddin; Yang, Y.; Yang, L. The influence of tree species on soil properties and microbial communities following afforestation of abandoned land in northeast China. Eur. J. Soil Biol. 2018, 85, 73–78. [Google Scholar] [CrossRef]
- Wang, Z.; Bai, Y.; Hou, J.; Li, F.; Li, X.; Cao, R.; Deng, Y.; Wang, H.; Jiang, Y.; Yang, W. The changes in soil microbial communities across a subalpine forest successional series. Forests 2022, 13, 289. [Google Scholar] [CrossRef]
- Zhao, F.Z.; Bai, L.; Wang, J.Y.; Deng, J.; Ren, C.J.; Han, X.H.; Yang, G.H.; Wang, J. Change in soil bacterial community during secondary succession depend on plant and soil characteristics. Catena 2019, 173, 246–252. [Google Scholar] [CrossRef]
- Peng, X.; Tamura, K.; Asano, M.; Takano, A.; Kawagoe, M.; Kamijo, T. Changes in soil physical and chemical properties during vegetation succession on Miyake-jima island. Forests 2021, 12, 1435. [Google Scholar] [CrossRef]
- Zhou, J.; Jin, Z.; Yuan, W.; Chen, W.; Li, X.; Xiong, L.; Cheng, G. Microbial Communities and soil respiration during rice growth in paddy fields from karst and non-karst areas. Agronomy 2023, 13, 2001. [Google Scholar] [CrossRef]
- Torsvik, V.; Ovreas, L. Microbial diversity and function in soil: From genes to ecosystems. Curr. Opin. Microbiol. 2002, 5, 240–245. [Google Scholar] [CrossRef]
- Sielaff, A.C.; Upton, R.N.; Hofmockel, K.S.; Xu, X.; Polley, H.W.; Wilsey, B.J. Microbial community structure and functions differ between native and novel (exotic-dominated) grassland ecosystems in an 8-year experiment. Plant Soil 2018, 432, 359–372. [Google Scholar] [CrossRef]
- Daniel, G.F.; Nilsson, T.; Singh, A.P. Degradation of lignocellulosics by unique tunnel-forming bacteria. Can. J. Microbiol. 1987, 33, 943–948. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, N.; Tang, T.; Zhou, S.; Cai, Q. Small run-of-river dams affect taxonomic and functional beta-diversity, community assembly process of benthic diatoms. Front. Ecol. Evol. 2022, 10, 895328. [Google Scholar] [CrossRef]
- Yan, G.; Luo, X.; Huang, B.; Wang, H.; Sun, X.; Gao, H.; Zhou, M.; Xing, Y.; Wang, Q. Assembly processes, driving factors, and shifts in soil microbial communities across secondary forest succession. Land Degrad. Dev. 2023, 34, 3130–3143. [Google Scholar] [CrossRef]
- He, G.; Peng, T.; Guo, Y.; Wen, S.; Ji, L.; Luo, Z. Forest succession improves the complexity of soil microbial interaction and ecological stochasticity of community assembly: Evidence from Phoebe bournei-dominated forests in subtropical regions. Front. Microbiol. 2022, 13, 1021258. [Google Scholar] [CrossRef]
- Stegen, J.C.; Lin, X.; Fredrickson, J.K.; Konopka, A.E. Estimating and mapping ecological processes influencing microbial community assembly. Front. Microbiol. 2015, 6, 370. [Google Scholar] [CrossRef]
- Caruso, T.; Chan, Y.; Lacap, D.C.; Lau, M.C.Y.; McKay, C.P.; Pointing, S.B. Stochastic and deterministic processes interact in the assembly of desert microbial communities on a global scale. ISME J. 2011, 5, 1406–1413. [Google Scholar] [CrossRef]
- Martiny, A.C.; Jorgensen, T.M.; Albrechtsen, H.; Arvin, E.; Molin, S. Long-term succession of structure and diversity of a biofilm formed in a model drinking water distribution system. Appl. Environ. Microbiol. 2003, 69, 6899–6907. [Google Scholar] [CrossRef]
- Sigler, W.V.; Zeyer, J. Microbial diversity and activity along the forefields of two receding glaciers. Microb. Ecol. 2002, 43, 397–407. [Google Scholar] [CrossRef]
- Deiglmayr, K.; Philippot, L.; Tscherko, D.; Kandeler, E. Microbial succession of nitrate-reducing bacteria in the rhizosphere of Poa alpina across a glacier foreland in the Central Alps. Environ. Microbiol. 2006, 8, 1600–1612. [Google Scholar] [CrossRef]
- Badri, D.V.; Chaparro, J.M.; Zhang, R.; Shen, Q.; Vivanco, J.M. Application of natural blends of phytochemicals derived from the root exudates of Arabidopsis to the soil reveal that phenolic-related compounds predominantly modulate the soil microbiome. J. Biol. Chem. 2013, 288, 4502–4512. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Badri, D.V.; Bakker, M.G.; Sugiyama, A.; Manter, D.K.; Vivanco, J.M. Root exudation of phytochemicals in arabidopsis follows specific patterns that are developmentally programmed and correlate with soil microbial functions. PLoS ONE 2013, 8, e55731. [Google Scholar] [CrossRef]
- Chen, S.; Wang, W.; Xu, W.; Wang, Y.; Wan, H.; Chen, D.; Tang, Z.; Tang, X.; Zhou, G.; Xie, Z.; et al. Plant diversity enhances productivity and soil carbon storage. Proc. Natl. Acad. Sci. USA 2018, 115, 4027–4032. [Google Scholar] [CrossRef] [PubMed]
- Cline, L.C.; Zak, D.R. Soil microbial communities are shaped by plant-driven changes in resource availability during secondary succession. Ecology 2015, 96, 3374–3385. [Google Scholar] [CrossRef] [PubMed]
- Hooper, D.U.; Vitousek, P.M. Effects of plant composition and diversity on nutrient cycling. Ecol. Monogr. 1998, 68, 121–149. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Hillebrand, H.; Jonsson, P.R. Multiple functions increase the importance of biodiversity for overall ecosystem functioning. Ecology 2008, 89, 1223–1231. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zu, L.; Zhou, G.; Long, F.; Zang, L.; Chen, D.; Zhang, G.; Sui, M.; He, Y.; Liu, Q. Stochastic Processes Dominate Soil Microbial Community Assembly during the Restoration of Degraded Karst Forests. Forests 2024, 15, 594. https://doi.org/10.3390/f15040594
Zu L, Zhou G, Long F, Zang L, Chen D, Zhang G, Sui M, He Y, Liu Q. Stochastic Processes Dominate Soil Microbial Community Assembly during the Restoration of Degraded Karst Forests. Forests. 2024; 15(4):594. https://doi.org/10.3390/f15040594
Chicago/Turabian StyleZu, Lei, Guanghui Zhou, Fayu Long, Lipeng Zang, Danmei Chen, Guangqi Zhang, Mingzhen Sui, Yuejun He, and Qingfu Liu. 2024. "Stochastic Processes Dominate Soil Microbial Community Assembly during the Restoration of Degraded Karst Forests" Forests 15, no. 4: 594. https://doi.org/10.3390/f15040594