


The Dynamics of Stand Structure Development and Natural Regeneration of Common Beech (Fagus sylvatica L.) in Plitvice Lakes National Park

Abstract
:1. Introduction
2. Materials and Methods
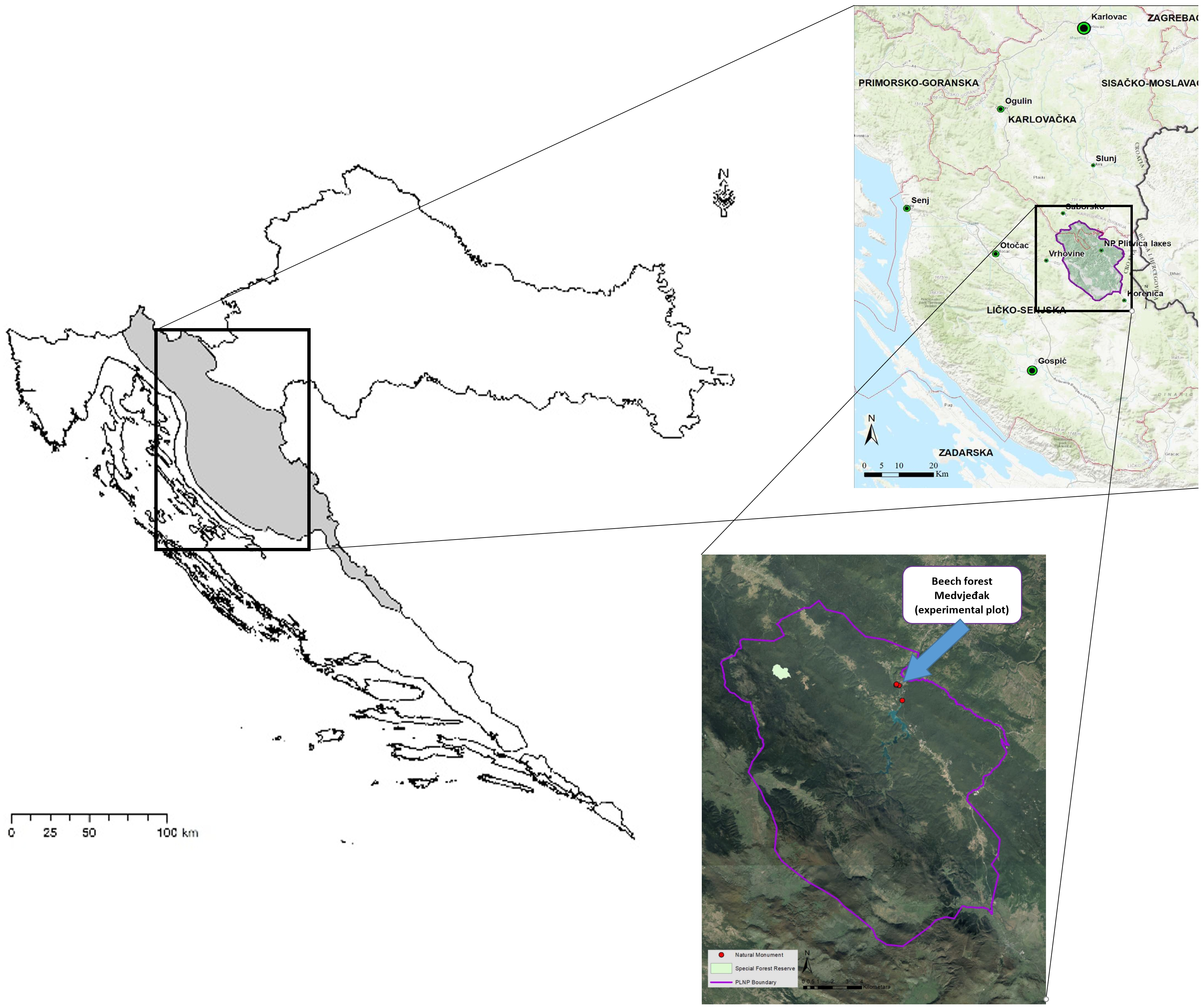
2.1. Study Area
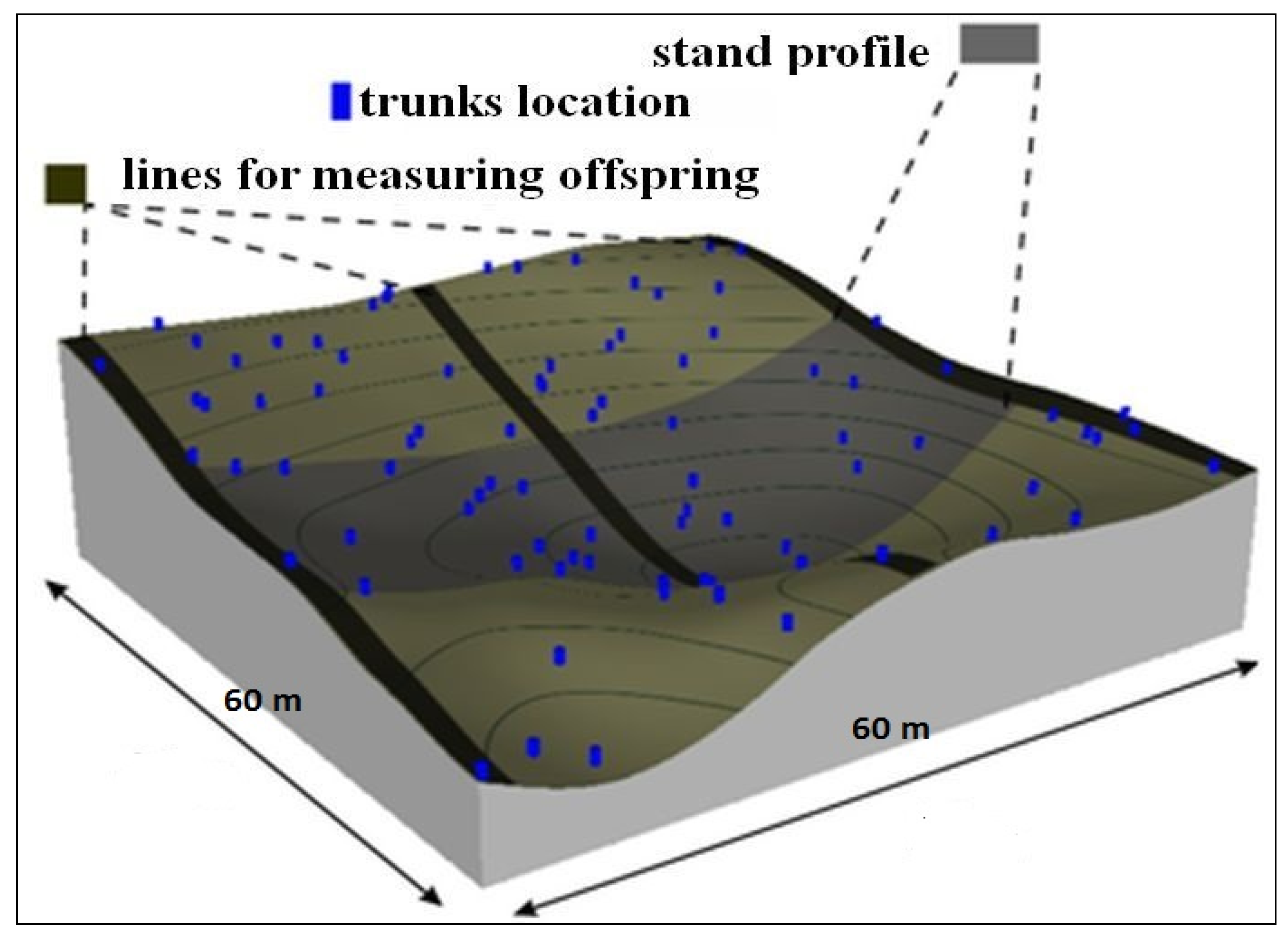
2.2. Experimental Design
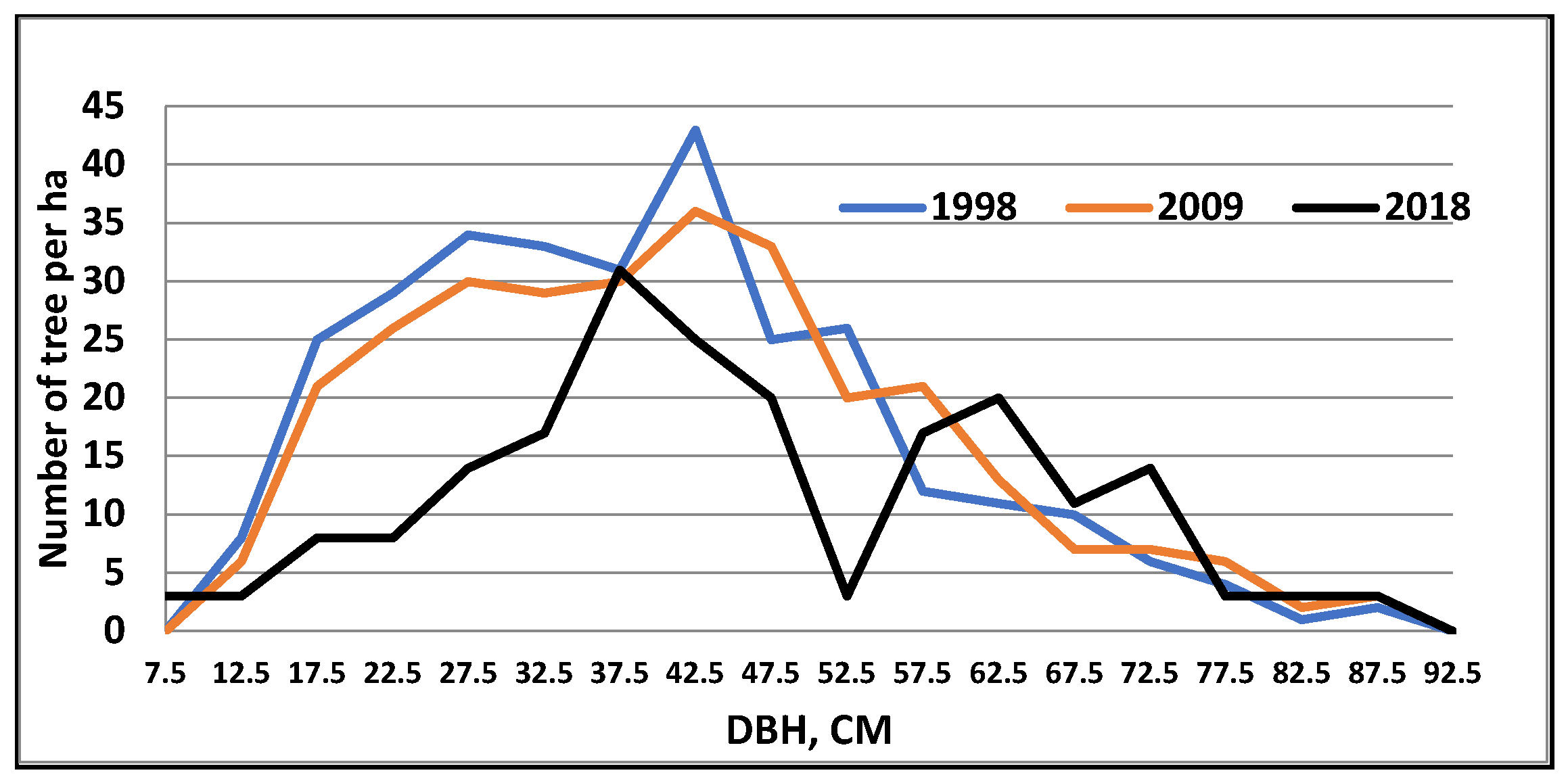
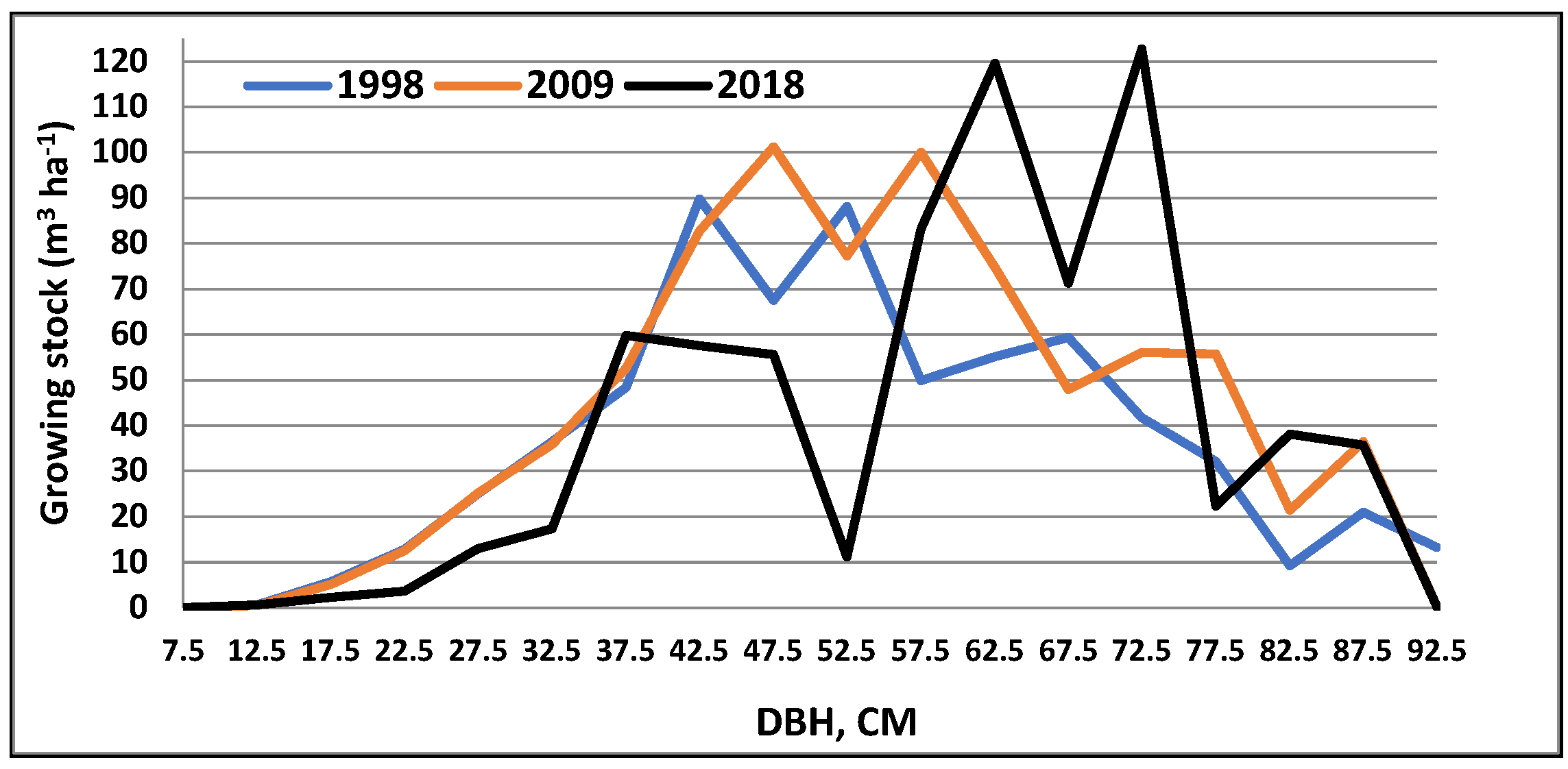
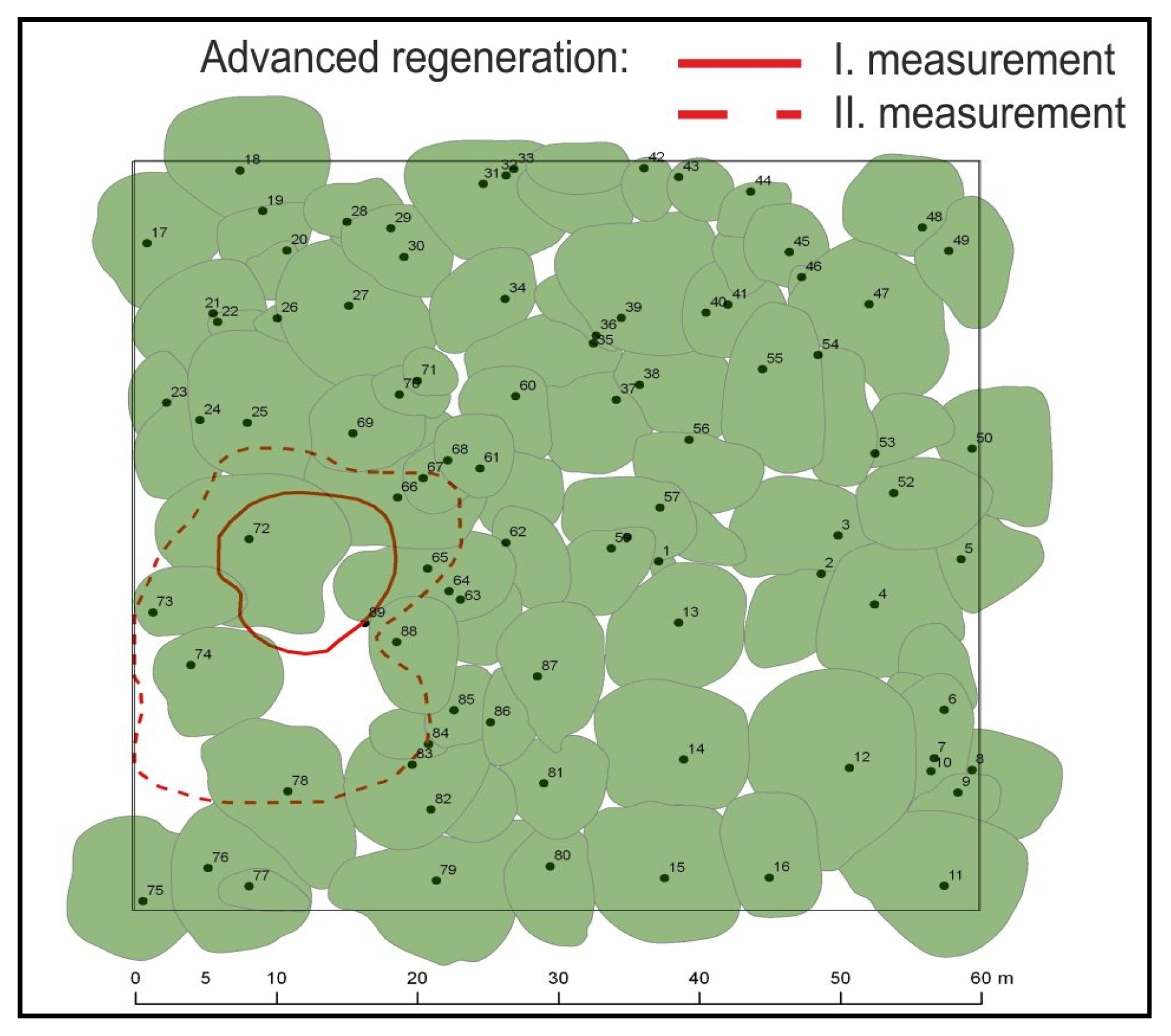
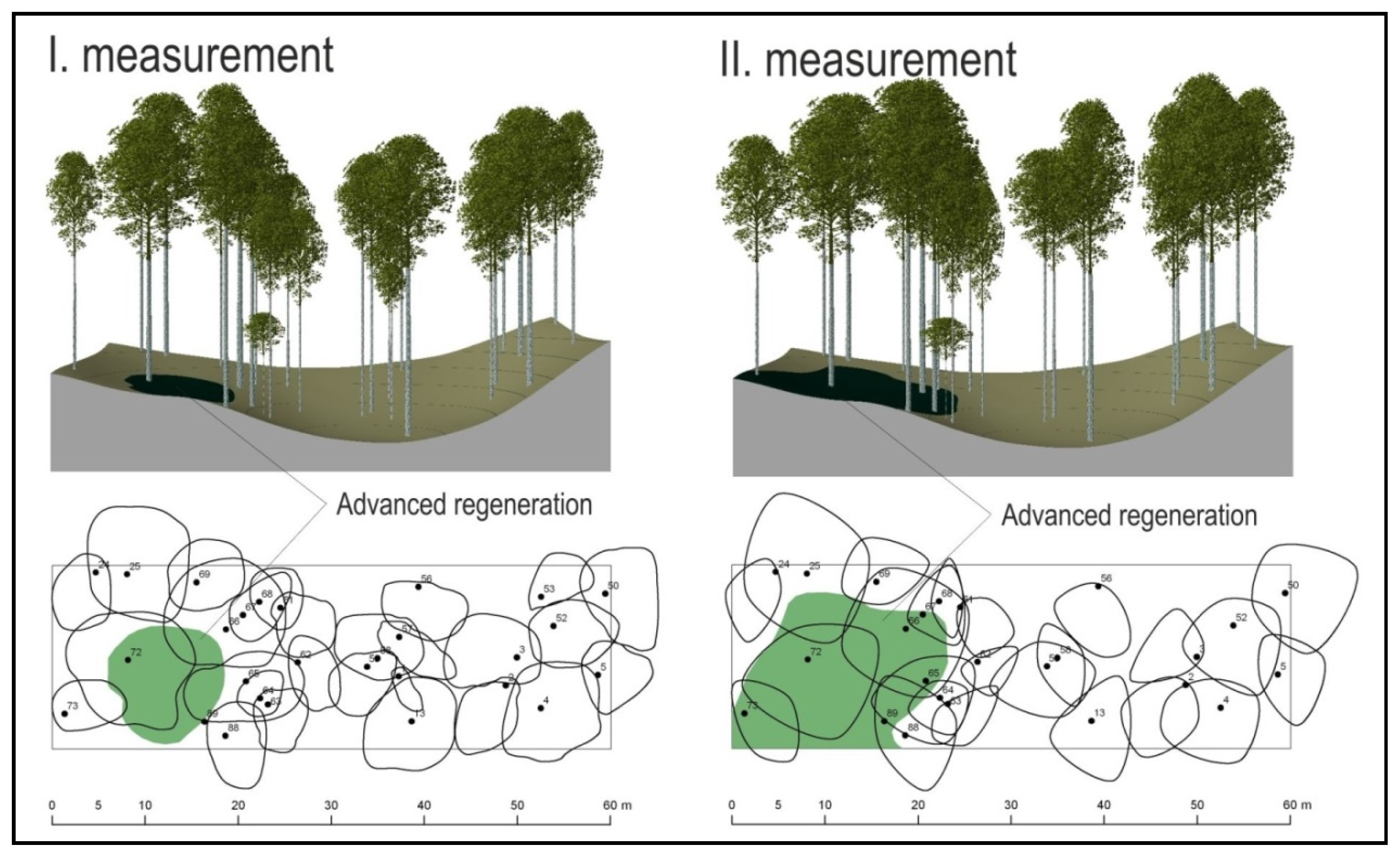
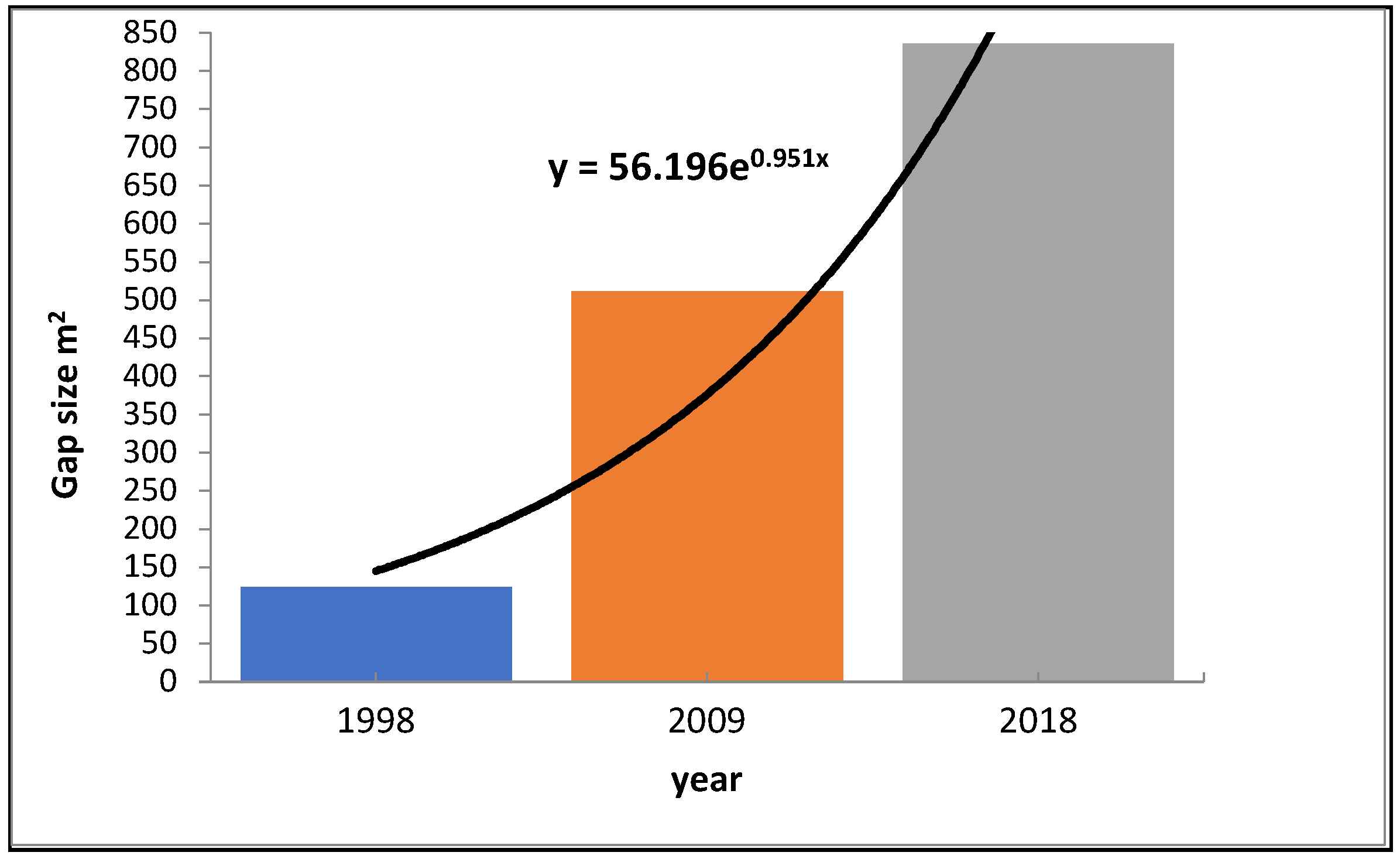
3. Results
4. Discussion
- (a)
- Strict protection zone, 80.73%;
- (b)
- Moderate protection zone, 17.12%;
- (c)
- Use zone, 2.15%.
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anić, I.; Mikac, S. Prirodno pomlađivanje sastojina obične bukve (Fagus sylvatica L.) oplodnim sječama na malim površinama. Croat. J. For. Eng. 2011, 32, 19–29, (In Croatian with English summary). [Google Scholar]
- Diaci, J.; Roženbergar, D.; Kolar, U.; Pisek, R.; Nagel, T.; Hladnik, D. Methodologies for monitoring forest development in strict forest reserves. In Gazdovanje Šumskim Ekosistemima Nacionalnih Parkova i drugih Zaštićenih Područja; Maunaga, Z., Ed.; Šumarski fakultet u Banjoj Luci, Bosna i Hercegovina: Banja Luka, Bosna i Hercegovina, 2006; pp. 25–33. [Google Scholar]
- FAO. Global Forest Resources Assessment; FAO: Rome, Italy, 2010; p. 343. [Google Scholar]
- Ellenberg, H. Vegetation Mitteleuropas mit den Alpen: In Ökologischer, Dynamischer und Historischer Sicht; Ulmer: Stuttgart, Germany, 1996; p. 1095. [Google Scholar]
- Ellenberg, H.; Leuschner, C. Vegetation Mitteleuropas mit den Alpen; Ulmer: Stuttgart, Germany, 2010; p. 1333. [Google Scholar]
- Leibundgut, H. Europäische Urwälder der Bergstufe; P. Haupt: Bern, Switzerland, 1982; p. 308. [Google Scholar]
- Pruša, E. Die Böhmischen und Mährischen Urwälder—Ihre Struktur und Ökologie; Verlag der Tschechoslowakischen Akademie der Wisenschaften: Prague, Czech Republic, 1985; p. 573. [Google Scholar]
- Korpel, Š. Die Urwalder der Westkarpaten; Gustav Fischer Erlag: Stuttgart/Jena, Germany; New York, NY, USA, 1995; p. 310. [Google Scholar]
- Korpel, Š. Razvoj i struktura bukovo-jelovih prašuma i njihova primjena kod gospodarenja prebornom šumom. Šum. List 1996, 3–4, 203–219. [Google Scholar]
- Emborg, J. Understorey light conditions and regeneration with respect to the structural dynamics of a near-natural temperate deciduous forest in Denmark. For. Ecol. Manag. 1998, 106, 83–95. [Google Scholar] [CrossRef]
- Christensen, M.; Emborg, J.; Busse Nielsen, A. The forest cycle of Suserup Skov-revisited and revised. Ecol. Bull. 2006, 52, 33–42. [Google Scholar]
- Rugani, T.; Diaci, J.; Hladnik, D. Gap Dynamics and Structure of Two Old-Growth Beech Forest Remnants in Slovenia. PLoS ONE 2013, 8, e52641. [Google Scholar] [CrossRef] [PubMed]
- Lertzman, K.P. Patterns of gap-phase replacement in a subalpine, old-growth forest. Ecology 1992, 73, 657–669. [Google Scholar] [CrossRef]
- Yamamoto, S.; Nishimura, N. Canopy gap formation and replacement pattern of major tree species among developmental stages of beech (Fagus crenata) stands, Japan. Plant Ecol. 1999, 140, 167–176. [Google Scholar] [CrossRef]
- Battles, J.J.; Fahey, T.J. Gap Dynamics Following Forest Decline: A Case Study of Red Spruce Forests. Ecol. Appl. 2000, 10, 760–774. [Google Scholar] [CrossRef]
- Nagel, T.A.; Svoboda, M. Gap disturbance regime in an old-growth Fagus-Abies forest in the Dinaric Mountains, Bosnia-Herzegovina. Can. J. For. Res. 2008, 38, 2728–2737. [Google Scholar] [CrossRef]
- Kucbel, S.; Jaloviar, P.; Saniga, M.; Vencurik, J.; Klimas, V. Canopy gaps in an old-growth fir-beech forest remnant of Western Carpathians. Eur. J. For. Res. 2010, 129, 249–259. [Google Scholar] [CrossRef]
- Bottero, A.; Garbarino, M.; Dukić, V.; Govedar, Z.; Lingua, E.; Nagel, T.A.; Motta, R. Gap-phase dynamics in the old-growth forest of Lom, Bosnia and Herzegovina. Silva Fenn. 2011, 45, 875–887. [Google Scholar] [CrossRef]
- Dubravac, T.; Dekanić, S.; Novotny, V.; Milašinčić, J. Natural Regeneration of Beech Forests in the Strict Protected Area of the Plitvice Lakes National Park. South-East Eur. For. 2013, 4, 95–103. [Google Scholar] [CrossRef]
- Tabaku, V.; Meyer, P. Gap patterns of Albanian and Central European beech forests. Forstarchiv 1999, 70, 87–97. [Google Scholar]
- Drössler, L.; von Lüpke, B. Canopy gaps in two virgin beech forest reserves in Slovakia. J. For. Sci. 2005, 51, 446–457. [Google Scholar] [CrossRef]
- Drössler, L.; von Lüpke, B. Stand structure, regeneration and site conditions in two virgin beech forest reserves in Slovakia. Allg. Forst Jagdztg. 2007, 178, 121–135. [Google Scholar]
- Krejči, V.; Dubravac, T. Mogućnost obnove šuma Nacionalnih parkova. Rad. Šumar. Inst. 2001, 36, 113–122. [Google Scholar]
- Dubravac, T.; Vuletić, D.; Vrbek, B. Natural reforestation and future of beech and fir forests in the Risnjak NP. Period Biol. 2005, 107, 73–79. [Google Scholar]
- Dubravac, T.; Čavlović, J.; Roth, V.; Vrbek, B.; Novotny, V.; Dekanić, S. The structure and possibility of natural regeneration in managed and non-managed beech and fir forests in Croatia. Period Biol. 2007, 109, 21–27. [Google Scholar]
- Novotny, V.; Balenović, I.; Dubravac, T.; Vuletić, D.; Dekanić, S. First results of monitoring of stand structure changes in unmanaged beech stands in NP Plitvice Lakes. South-East Eur. For. 2010, 2, 91–98. [Google Scholar] [CrossRef]
- Juriček, M. Razvoj Šumske Vegetacije na Trajnim Pokusnim Plohama Nacionalnog Parka “Plitvička jezera”. Diplomski Rad, Šumarski fakultet Sveučilišta u Zagrebu, Zagreb, Croatia, 2014; p. 77. [Google Scholar]
- Alberdi, I.; Michalak, R.; Fischer, C.; Gasparini, P.; Brändli, U.-B.; Tomter, S.M.; Kuliesis, A.; Snorrason, A.; Redmond, J.; Hernández, L. Towards Harmonized Assessment of European Forest Availability for Wood Supply in Europe. For. Policy Econ. 2016, 70, 20–29. [Google Scholar] [CrossRef]
- Pucher, C.; Erber, G.; Hasenauer, H. Europe’s Potential Wood Supply by Harvesting System. Forests 2023, 14, 398. [Google Scholar] [CrossRef]
- Forest Europe. State of Europe’s Forests 2020. Available online: https://foresteurope.org/wp-content/uploads/2016/08/SoEF_2020.pdf (accessed on 25 August 2023).
- Vukelić, J. Šumska Vegetacija Hrvatske; Sveučilište u Zagrebu, Šumarski fakultet Zagreb, Državni Zavod za Zaštitu Prirode: Zagreb, Hrvatska, 2012; p. 403. [Google Scholar]
- Bertović, S. Prilog poznavanju odnosa klime i vegetacije u Hrvatskoj. Acta Biol. 1975, VII/2, 89–215. [Google Scholar]
- Šegulja, N.; Rauš, Đ. Sto trajnih ploha Republike Hrvatske (ekološka istraživanja). Glas. Šum. Pokuse 1993, 29, 133–148. [Google Scholar]
- Rauš, Đ. Sto trajnih ploha Repulike Hrvatske (ekološka istraživanja). Glas. Šum. Pokuse 1995, 32, 225–376. [Google Scholar]
- Hayakawa, Y.S.; Oguchi, T.; Komatsubara, J.; Ito, K.; Hori, K.; Nishiaki, Y. Rapid on-site topographic mapping with a handheld laser ranger finder for a geoarcheological survey in Syria. Geogr. Res. 2007, 45, 95–104. [Google Scholar] [CrossRef]
- USDA Forest Sevice. Available online: https://www.fia.fs.usda.gov (accessed on 10 November 2023).
- Braun-Blanquet, J. Pflanzensoziologie—Grundzuge der Vegetationskunde; Springer: Wien, Austria; New York, NY, USA, 1964. [Google Scholar]
- Westhoff, V.; van der Maarel, E. The Braun-Blanquet Approach. In Ordination and Classification of Communities; Whittaker, R.H., Ed.; W. Junk: The Hague, The Netherlands, 1973; pp. 617–726. [Google Scholar]
- Nikolić, T. (Ed.) Flora Croatica, Baza Podataka. Botanički Zavod, Prirodoslovno-Matematički Fakultet, Sveučilište u Zagrebu. 2015. Available online: http://hirc.botanic.hr/fcd (accessed on 10 November 2023).
- Friedrichs, M.; Hermoso, V.; Bremerich, V.; Langhans, S.D. Evaluation of habitat protection under the European Natura 2000 conservation network—The example for Germany. PLoS ONE 2018, 13, e0208264. [Google Scholar] [CrossRef] [PubMed]
- Cent, J.; Grodzińska-Jurczak, M.; Pietrzyk-Kaszyńska, A. Emerging multilevel environmental governance—A case of public participation in Poland. J. Nat. Conserv. 2014, 22, 93–102. [Google Scholar] [CrossRef]
- Butchart, S.H.M.; Walpole, M.; Collen, B.; van Strien, A.; Scharlemann, J.P.W.; Almond, R.E.A. Global Biodiversity: Indicators of recent declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef] [PubMed]
- Bormann, B.T.; Cunningham, P.G.; Gordon, J.C. Best Management Practices, Adaptive Management, or Both? In Proceedings of the 1996, National Society of American Foresters Convention, Portland, ME, USA, 28 October–1 November 1996. [Google Scholar]
- Salafsky, N.; Margoluis, R.; Redford, K. Adaptive Management: A Tool for Conservation Practitioners. Available on Biodiversity Support Program. Washington, DC. 2001. Available online: www.worldwildlife.org/bsp/ (accessed on 3 October 2023).
- Schreiber, E.S.G.; Bearlin, A.R.; Nicol, S.J.; Todd, C.R. Adaptive management: A synthesis of current understanding and effective application. Ecol. Manag. Restor. 2004, 5, 177–182. [Google Scholar] [CrossRef]
- Williams, B.K. Passive and active adaptive management: Approaches and an example. J. Environ. Manag. 2011, 92, 1371–1378. [Google Scholar] [CrossRef]
- Magdić, N. Small-Scale Natural Regeneration of the Beech Forests in the Plitvice Lakes National Park. Master’s Thesis, Faculty of Forestry, University of Zagreb, Zagreb, Croatia, 2010; p. 62, (In Croatian with English summary). [Google Scholar]
- Gaston, K.J.; Jackson, S.F.; Cantú-Salazar, L.; Cruz-Pinón, G. The ecological performance of protected areas. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 93–113. [Google Scholar] [CrossRef]
- Guedes-Santos, J.; Correia, R.A.; Jepson, P.; Ladle, R.J. Evaluating public interest in protected areas using Wikipedia page views. J. Nat. Conserv. 2021, 63, 126040. [Google Scholar] [CrossRef]
- Dasgupta, S. Do Protected Areas Work in the Tropics? Mangabay Series: Conservation Effectiveness. 2017. Available online: https://news.mongabay.com/2017/12/do-protected-areas-work-in-the-tropics/ (accessed on 5 October 2023).
- Castro, A.J.; Martín-López, B.; López, E.; Plieninger, T.; Alcaraz-Segura, D.; Vaughn, C.C. Do protected areas networks ensure the supply of ecosystem services? Spatial patterns of two nature reserve systems in semi-arid Spain. Appl. Geogr. 2015, 60, 1–9. [Google Scholar] [CrossRef]
- von Oheimb, G.; Westphal, C.; Tempel, H.; Härdtle, W. Structural pattern of a near-natural beech forest (Fagus sylvatica) (Serrahn, North-east Germany). For. Ecol. Manag. 2005, 212, 253–263. [Google Scholar] [CrossRef]
- Anić, I. The Influence of Stand Structure and Regeneration on the Sustainability of Fir-Beech and Beech Forests in the National Park Plitvice Lakes. 2007. Available online: http://bib.irb.hr/datoteka/326894.Microsoft_Word_-_plitvice_2006_konacno_izvj.pdf (accessed on 15 November 2023).
- Anić, I.; Mikac, S. Structure, texture and regeneration of Dinaric beech-fir Virgin Forest of Ćorkova Uvala. Šum. List 2008, 132, 505–515, (In Croatian with English summary). [Google Scholar]
- Łysik, M. Ten Years of Change in Ground-Layer Vegetation of European Beech Forest in the Protected Area (Ojców National Park, South Poland). Pol. J. Ecol. 2008, 56, 17–31. [Google Scholar]
- Madsen, P.; Hahn, K. Natural regeneration in a beech-dominated forest managed by close-to-nature principles—A gap cutting based experiment. Can. J. For. Res. 2008, 38, 1716–1729. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
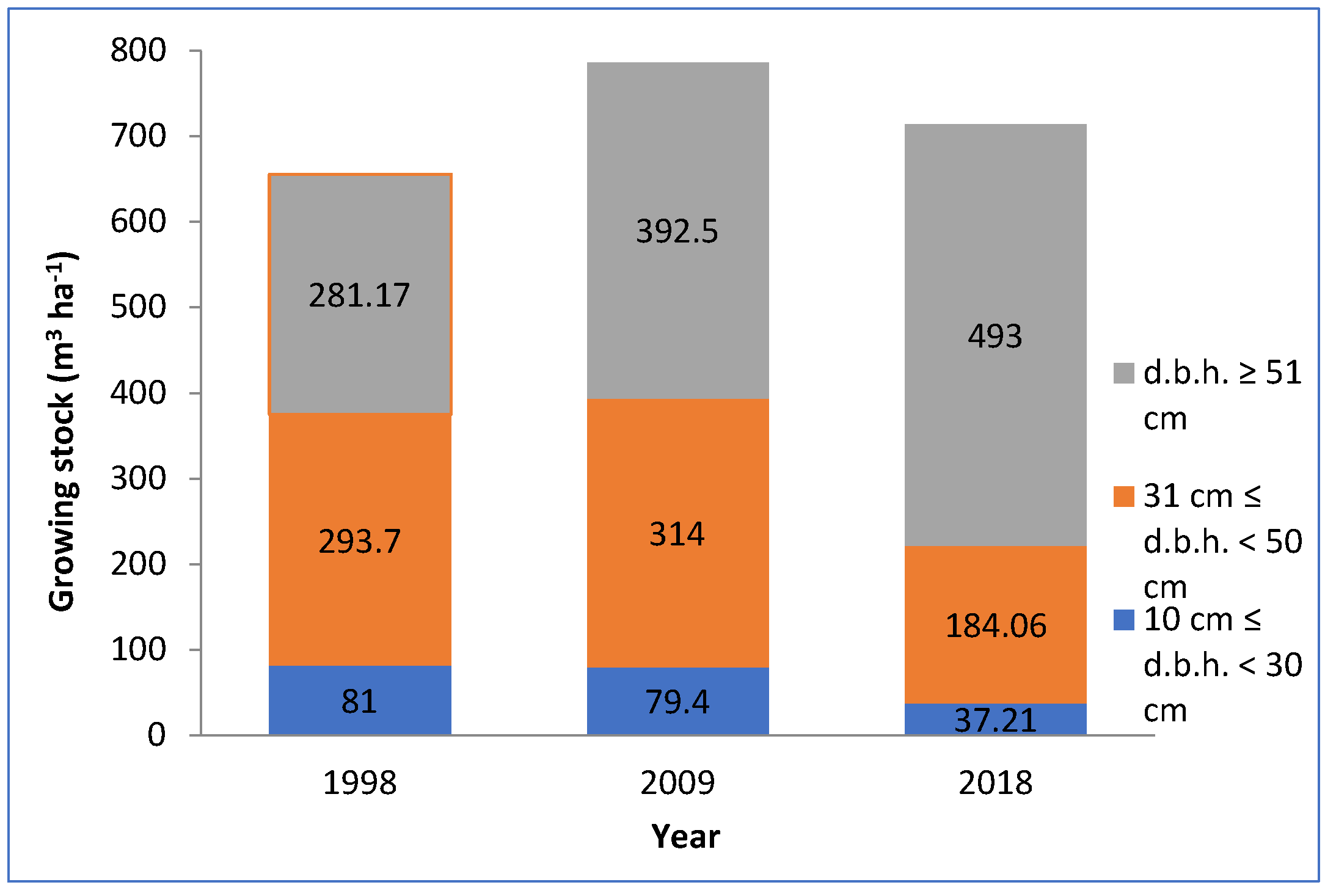{kind=link}
{kind=link}
{kind=link}
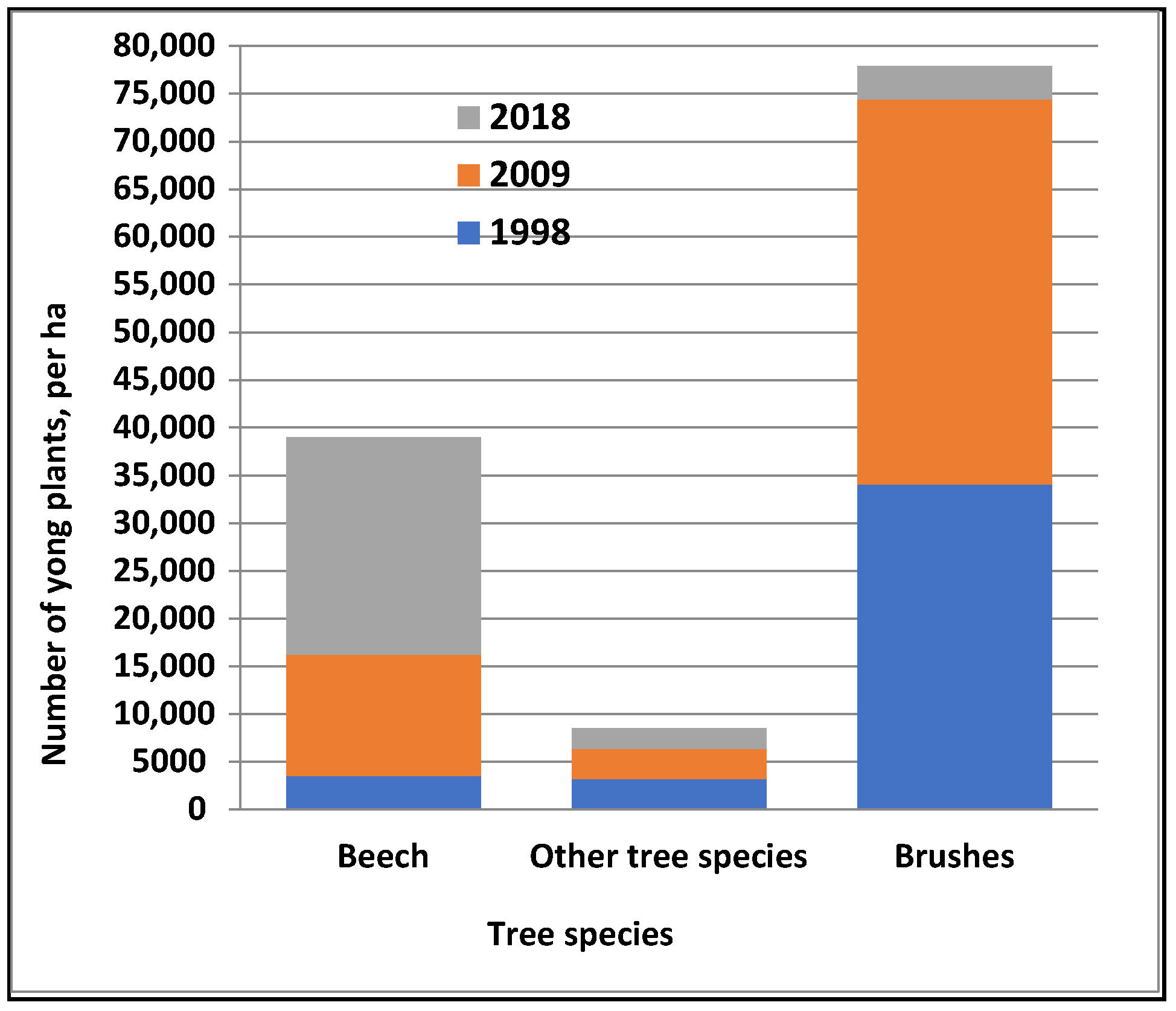{kind=link}
{kind=link}
| Measurement | N | G | V | DBH | h | g | v |
|---|---|---|---|---|---|---|---|
| Year | trees/ha | m2ha−1 | m3/ha−1 | cm | m | m2 | m3 |
| 1998 | 301 | 43.11 | 656.41 | 39.3 | 26.9 | 0.14 | 2.18 |
| 2009 | 291 | 45.68 | 803.07 | 41.1 | 27.1 | 0.16 | 2.76 |
| 2018 | 196 | 37.80 | 695.80 | 46.8 | 32.0 | 0.19 | 3.55 |
| Measurement Years | 1998. | 2009. | 2018. |
|---|---|---|---|
| Mean | 39.10 | 41.11 | 46.83 |
| Standard Error | 0.928867 | 0.983053 | 2.09356 |
| Median | 38.00 | 40.50 | 44.30 |
| Mode | 37.00 | 35.50 | 39.75 |
| Standard Deviation | 16.12 | 16.77 | 17.64 |
| Sample Variance | 259.7007 | 281.2206 | 311.1924 |
| Kurtosis | 0.287316 | 0.141323 | −0.60844 |
| Skewness | 0.671723 | 0.587233 | 0.258718 |
| Range | 89 | 89.5 | 78.05 |
| Minimum | 11.00 | 11.00 | 10.90 |
| Maximum | 100.00 | 100.50 | 88.95 |
| Sum | 11768 | 11963 | 3324.853 |
| Count | 301 | 291 | 196 |
| Plot number: | TPP 31 | Soil: | Classification by Braun-Blanquet—in abundance and cover | ||||
| Date: | 1980, 1988, 2004 | Brown soil on limestone | |||||
| Locality: | Medvjeđak | ||||||
| Area: | 20 × 20 m (400 m2) | Growing form: | |||||
| Elevation: | 825 m | Regular tall forest | +—insignificant | ||||
| Coordinates: | N 44°53.683′ | 1—1%–10% | |||||
| E 15°38.308′ | 2—10%–25% | ||||||
| Bedrock: | Limestone | 3—25%–50% | |||||
| Exposition: | N | 4—50%–75% | |||||
| Inclination: | 10% | 5—75%–100% | |||||
| Cover: | |||||||
| I. Tree layer: | |||||||
| II. Shrub layer: | |||||||
| III. Ground layer: | |||||||
| Species: | Survey 1980 | Survey 1988 | Survey 2004 | Species: | Survey 1980 | Survey 1988 | Survey 2004 |
| I. | 95% | 90% | 85% | Asplenium trichomanes L. | + | + | |
| Fagus sylvatica L. | 5 | 5 | 5 | Lunaria rediviva L. | + | + | 1 |
| II. | 5% | 5% | 15% | Actea spicata L. | + | + | |
| Sambucus racemosa L. | 2 | 2 | Polistichum aculeatum (L.) Roth. | + | |||
| Rhamnus falax Boiss. | 2 | 1 | 1 | Sanicula euroaea L. | 1 | + | |
| Fagus sylvatica L. | 1 | 1 | 1 | Impatiens noli tengere L. | + | 1 | |
| Daphne mezereum L. | 1 | + | + | Glechoma hirsuta W.K. | + | ||
| Sambucus nigra L. | 1 | Lamium galeobdolon Huds. | 1 | ||||
| Lonicera xylosteum L. | + | Festuca sylvatica Huds. | 1 | ||||
| Corylus avellana L. | + | Scolopendrium vulgare Sm. | 1 | ||||
| Euonymus verrucosa Scop. | + | Epilobium hirsutum L. | + | ||||
| Crataegus monogyna Jacq. | + | Viola sylvestris Lam. | + | ||||
| Abies alba Mill. | + | Solanum dulcamara L. | + | ||||
| Lonicera alpigena L. | + | + | Aconitum vulparia Rchb. | + | + | ||
| Acer pseudoplatanus L. | + | 1 | + | Arum maculatum L. | + | + | |
| Fraxinus excelsior L. | + | + | + | Mercurialis perennis L. | + | + | + |
| III. | 80% | 80% | 80% | Milium effusum L. | + | + | |
| Asperula odorata L. | 2 | 3 | Pulmonaria officinalis L. | + | 1 | + | |
| Poligonatum multiflorum (L.) All. | + | + | + | Eupatorium cannabinum L. | + | + | + |
| Viola sylvatica Lam. | + | + | Allium ursinum L. | + | |||
| Driopteris filix mas (L.) Rich. | 2 | 1 | 2 | Asarum europaeum L. | + | ||
| Circea lutetiana L. | 1 | 2 | + | Phyllitis scolopendrium (L.) Newm. | 1 | ||
| Lamium orvala L. | 1 | + | + | Rubus hirtus L. | + | + | |
| Cardamine savensis Shultz. | + | 1 | + | Acotinum licoctonum L. | + | ||
| Mycelis muralis (L.) Rchb. | + | + | + | Anemone nemorosa L. | 1 | ||
| Solanum dulcamara L. | + | + | Stellaria nemorum L. | + | |||
| Geranium robertianum L. | + | + | 1 | Cardamine enneaphyllos (L.) Cr. | + | + | |
| Urtica dioica L. | + | + | + | Veratrum album L. | + | ||
| Senecio nemorensis L. | 1 | 1 | + | Stellaria nemorum L. | |||
| Galium rotundifolium L. | + | 1 | Euphorbia dulcis L. | + | |||
| Brachypodium sylvaticum (Huds.) R.B. | + | + | + | Epilobium montanum L. | + | ||
| Atropa belladonna L. | + | Lathyrus vernus (L.) Bernh. | + | ||||
| Galeobdolon luteum Huds. | 1 | 1 | Doronicum austriacum Jacq. | + | |||
| Fagus sylvatica L. | + | 1 | + | Carex pilosa Scop. | 1 | ||
| Carex sylvatica Huds. | + | + | Cardamine tryfolia L. | + | |||
| Heracleum spondylum L. | + | Scrophularia nodosa L. | + | ||||
| Veronica montana L. | + | + | Fragaria vesca L. | + | |||
| Galeopsis tetrahit L. | 1 | Atropa belladona L. | + | ||||
| Athyrium filix femina (L.) Roth. | + | 1 | 1 | Neottia nidus avis (L.) Rich. | + | ||
| Paris quadrifolia L. | 1 | Euphorbia carniolica Jacq. | + | ||||
| Salvia glutinosa L. | + | 1 | Stachys sylvatica L. | + | |||
| Euphorbia amygdaloides L. | + | + | Hacquetia epipactis (Scop.) DC. | + | |||
| Oxalis acetosella L. | + | + | Prenanthes purpurea L. | + | |||
| Melica uniflora | + | Cyclamen europaeum Mill. | + | ||||
| Height Class | Beech | Other Trees * | Shrubs ** | Total | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (cm) | 1998 | 2009 | 2018 | 1998 | 2009 | 2018 | 1998 | 2009 | 2018 | 1998 | 2009 | 2018 |
| to 30 | 1528 | 10,194 | 15,650 | 1556 | 1028 | 1136 | 28,167 | 36,194 | 3130 | 31,251 | 47,416 | 19,916 |
| 31–60 | 1417 | 1111 | 4016 | 1056 | 1306 | 621 | 5500 | 3917 | 111 | 7973 | 6334 | 4748 |
| 61–130 | 556 | 1139 | 1856 | 361 | 611 | 305 | 361 | 222 | 166 | 1278 | 1972 | 2327 |
| 131–150 | 28 | 139 | 378 | 56 | 28 | 55 | 84 | 167 | 433 | |||
| 151–200 | 28 | 83 | 526 | 167 | 111 | 55 | 56 | 195 | 250 | 581 | ||
| 201–250 | 28 | 250 | 28 | 83 | 28 | 111 | 250 | |||||
| 251> | 55 | 55 | 110 | 110 | ||||||||
| Total | 3557 | 12,694 | 22,731 | 3224 | 3167 | 2172 | 34,028 | 40,389 | 3462 | 40,809 | 56,360 | 28,365 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dubravac, T.; Barčić, D.; Rosavec, R.; Španjol, Ž.; Vojniković, S. The Dynamics of Stand Structure Development and Natural Regeneration of Common Beech (Fagus sylvatica L.) in Plitvice Lakes National Park. Forests 2024, 15, 357. https://doi.org/10.3390/f15020357
Dubravac T, Barčić D, Rosavec R, Španjol Ž, Vojniković S. The Dynamics of Stand Structure Development and Natural Regeneration of Common Beech (Fagus sylvatica L.) in Plitvice Lakes National Park. Forests. 2024; 15(2):357. https://doi.org/10.3390/f15020357
Chicago/Turabian StyleDubravac, Tomislav, Damir Barčić, Roman Rosavec, Željko Španjol, and Sead Vojniković. 2024. "The Dynamics of Stand Structure Development and Natural Regeneration of Common Beech (Fagus sylvatica L.) in Plitvice Lakes National Park" Forests 15, no. 2: 357. https://doi.org/10.3390/f15020357