
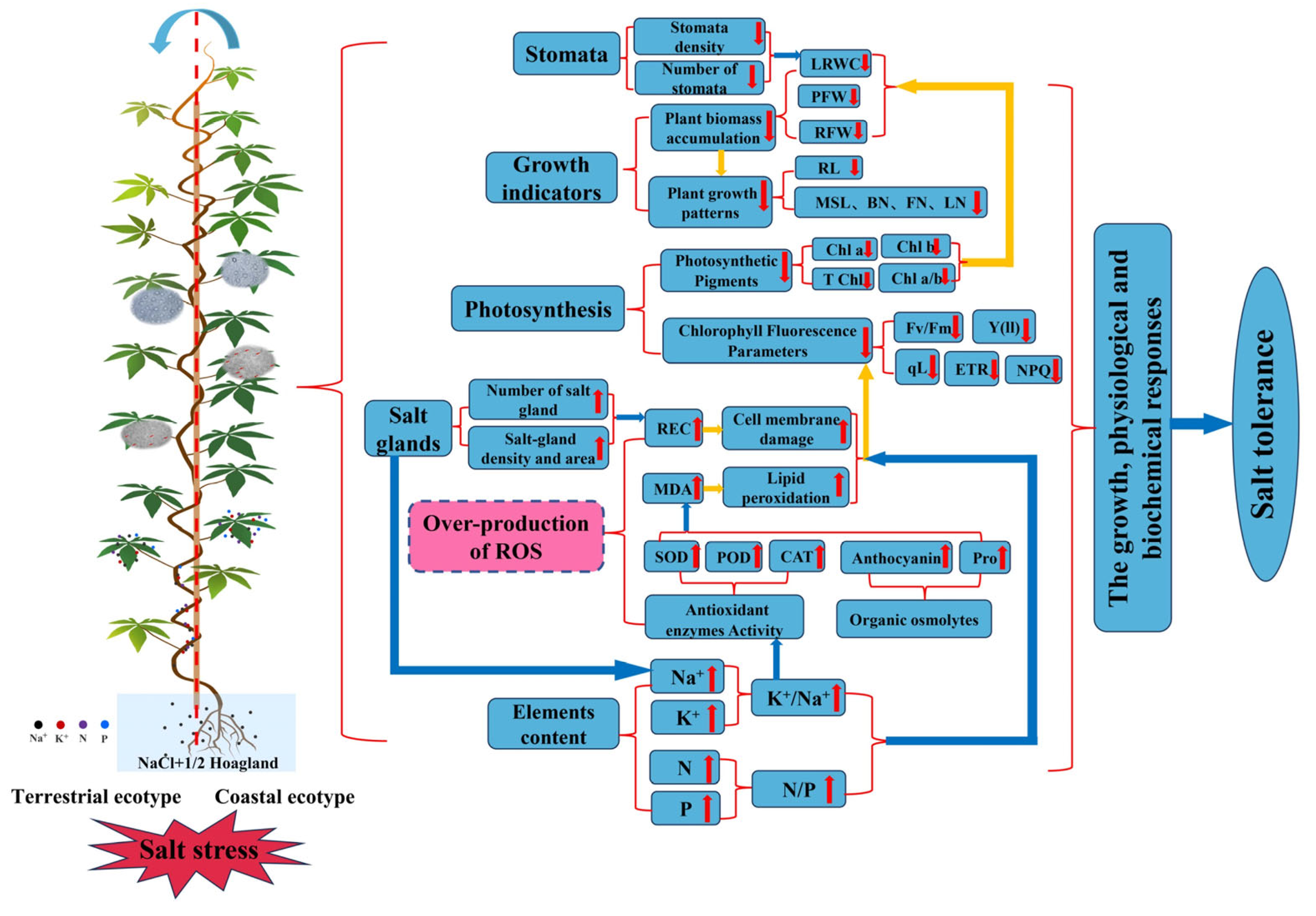
Ipomoea cairica (L.) from Mangrove Wetlands Acquired Salt Tolerance through Phenotypic Plasticity
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Plant Harvest and Growth Parameters
2.3. Salt Rejection and Ionic Balance
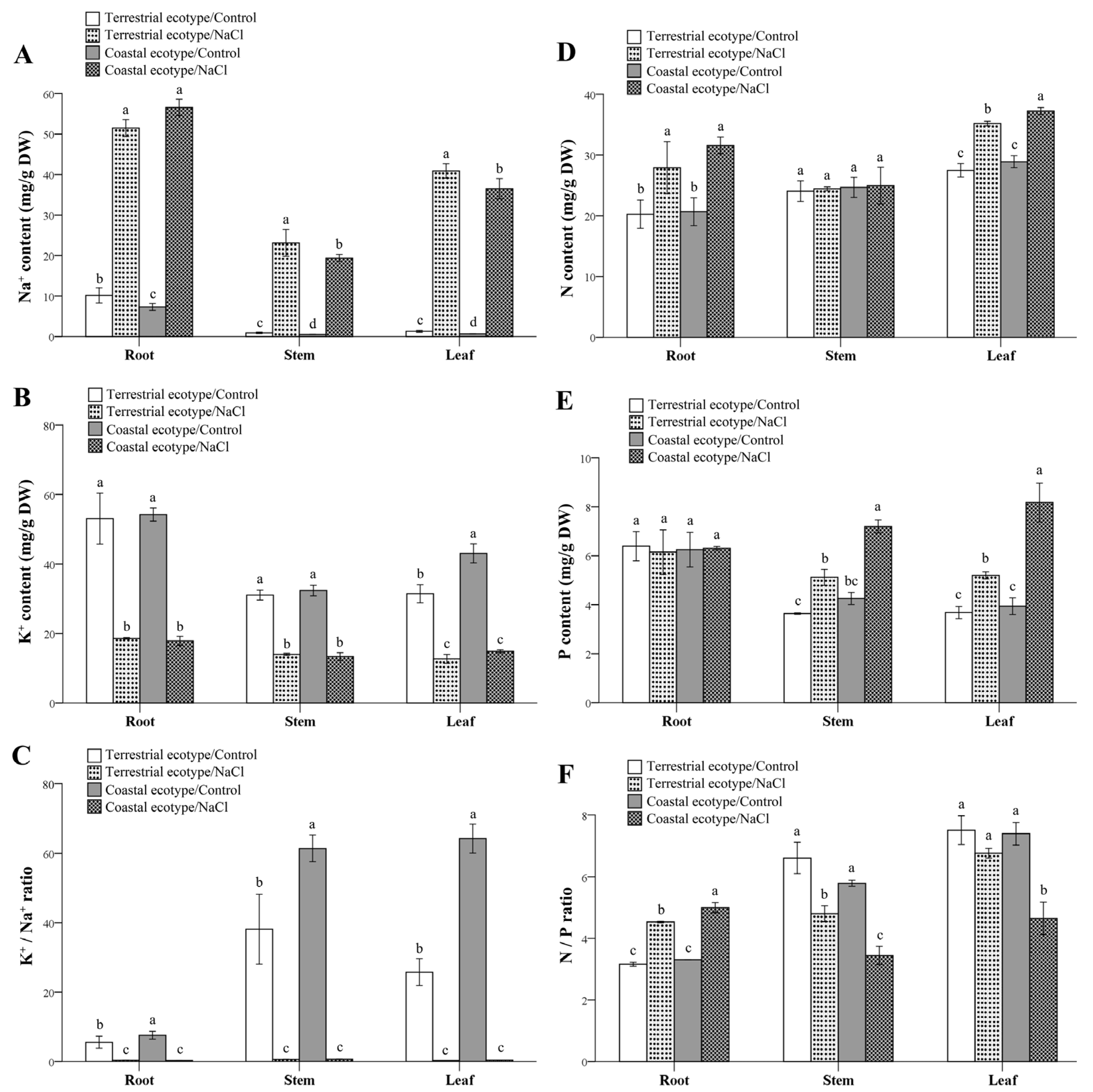
2.3.1. Determination of the Contents of Nitrogen, Phosphorus, Potassium, and Sodium in the Roots, Stems, and Leaves
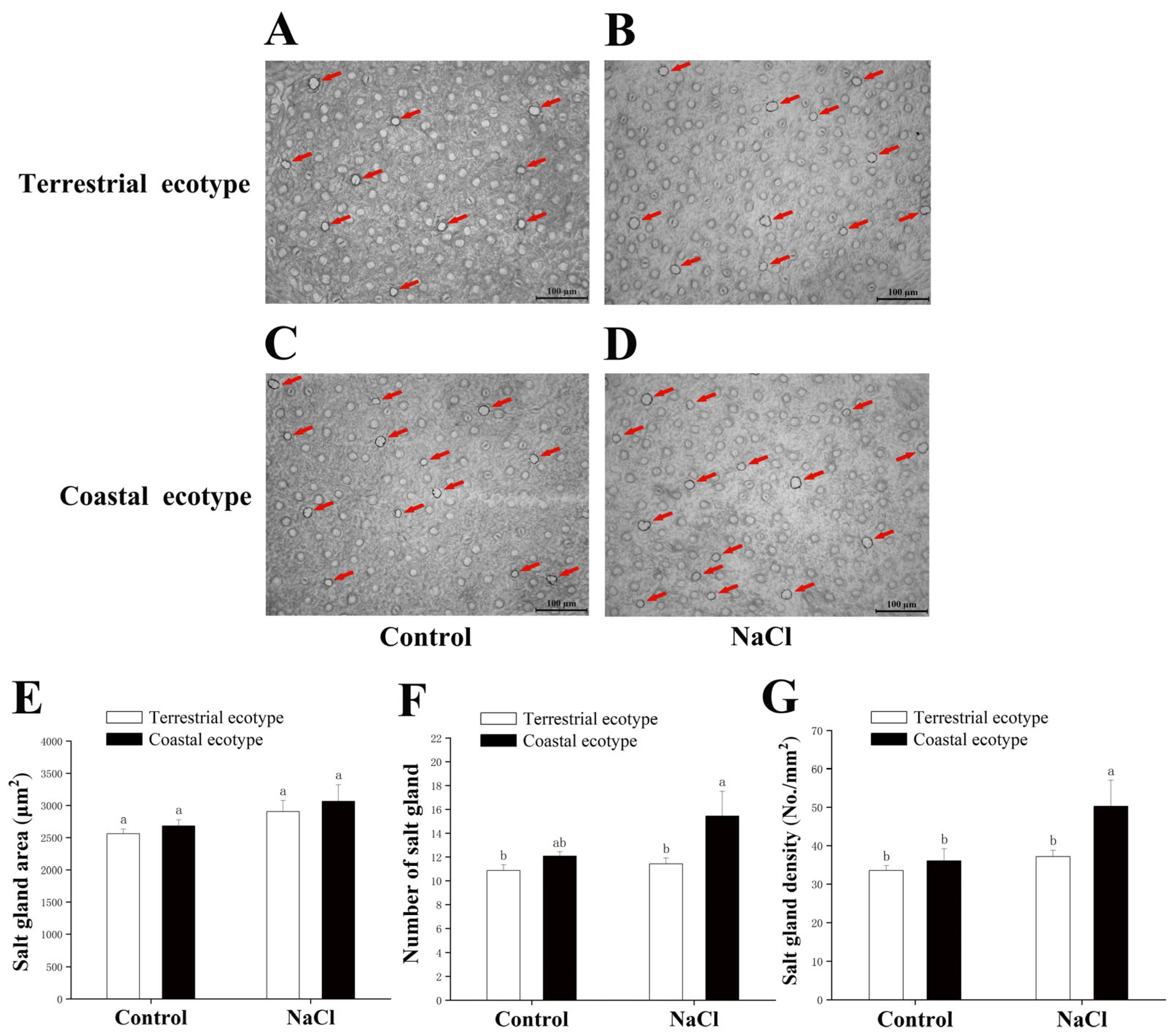
2.3.2. Measurement of the Density and Area of the Salt Glands
2.4. Determination of the Relative Electrical Conductivity (REC) and Content of Malondialdehyde (MDA)
2.4.1. Determination of the Relative Electrical Conductivity
2.4.2. Determination of the Relative Content of Malondialdehyde (MDA)
2.5. Detection of Apoptosis and the Integrity of Root Cell Membranes
2.6. Determination of the Anthocyanin Content and Antioxidant Enzyme Activities
2.6.1. Assays of the Antioxidant Enzymes
2.6.2. Determination of the Anthocyanin Content
2.7. Determination of the Osmotic Adjustment Indicators
2.7.1. Determination of the Content of Proline
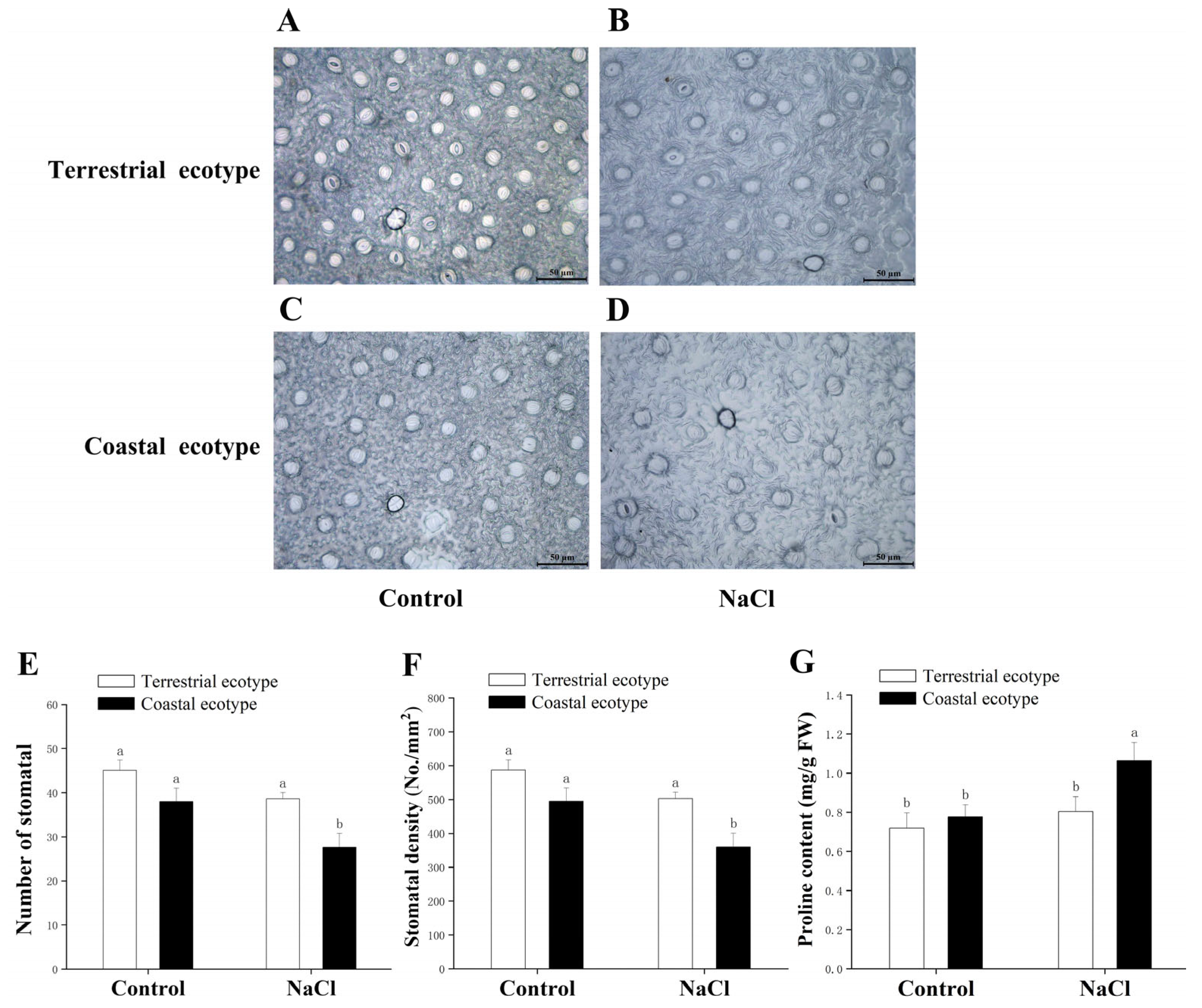
2.7.2. Measurement of Stomatal Density
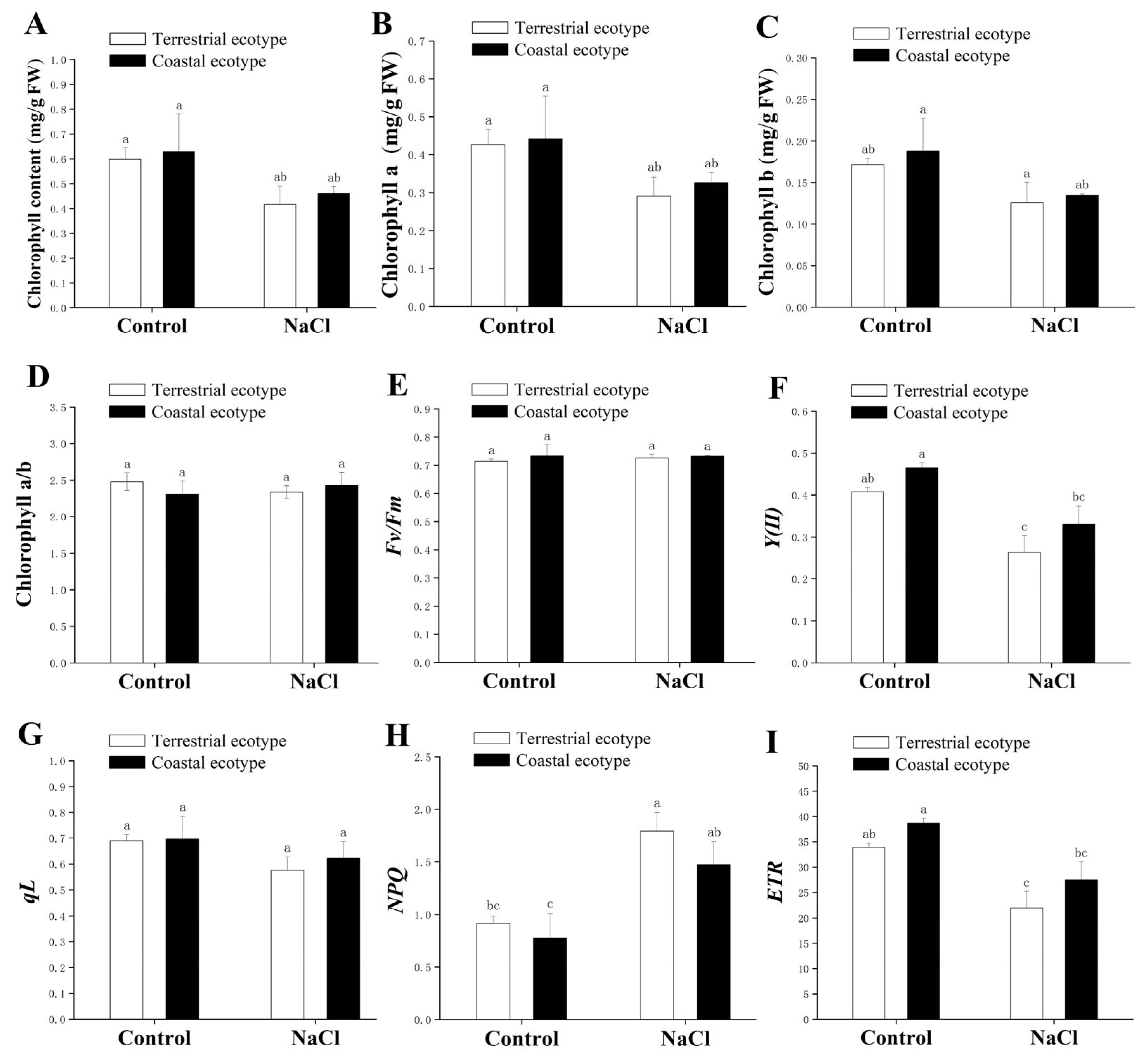
2.8. Measurement of the Content of Chlorophyll and Parameters of Chlorophyll Fluorescence
2.9. Statistical Analysis
3. Results
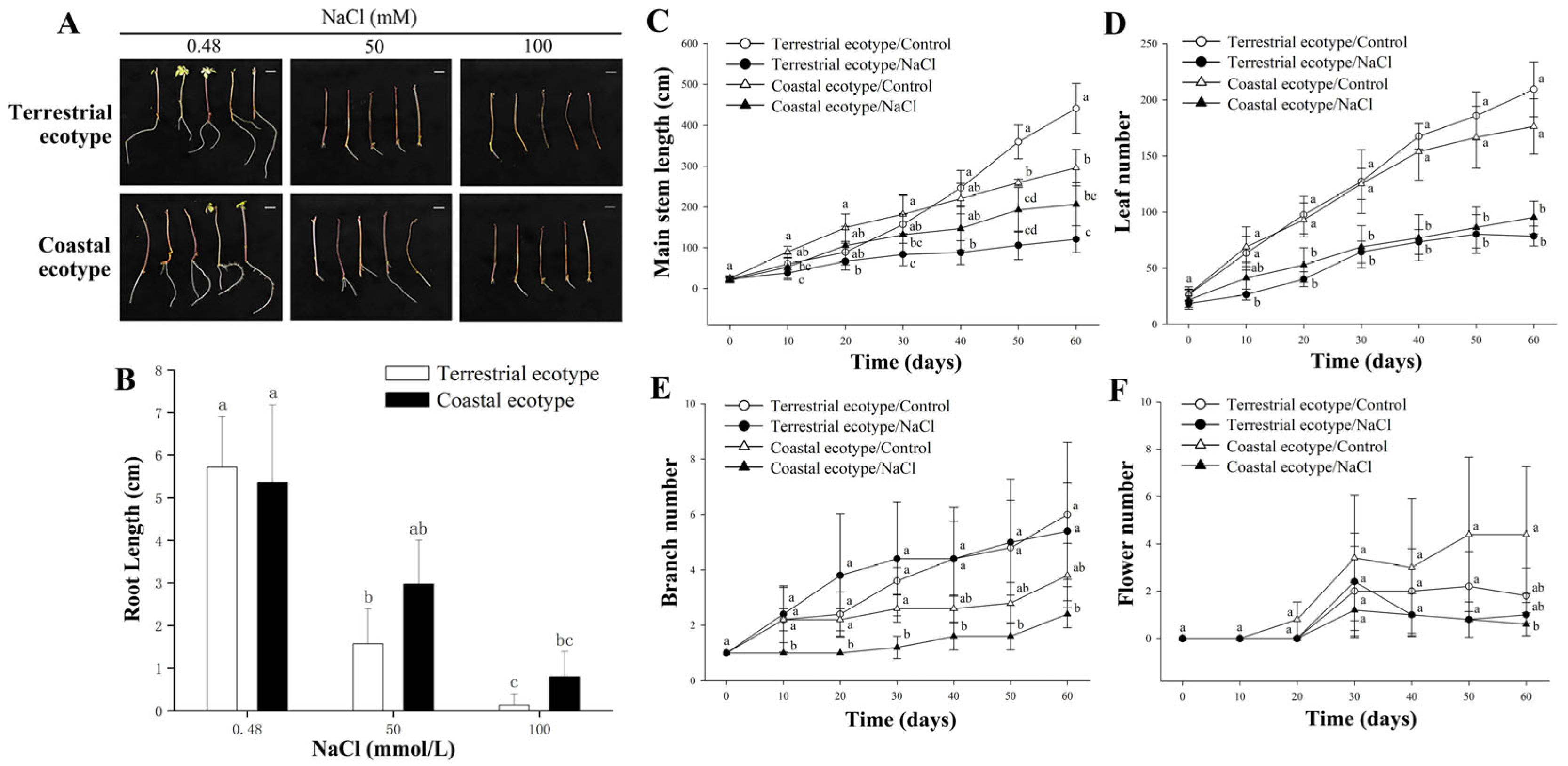
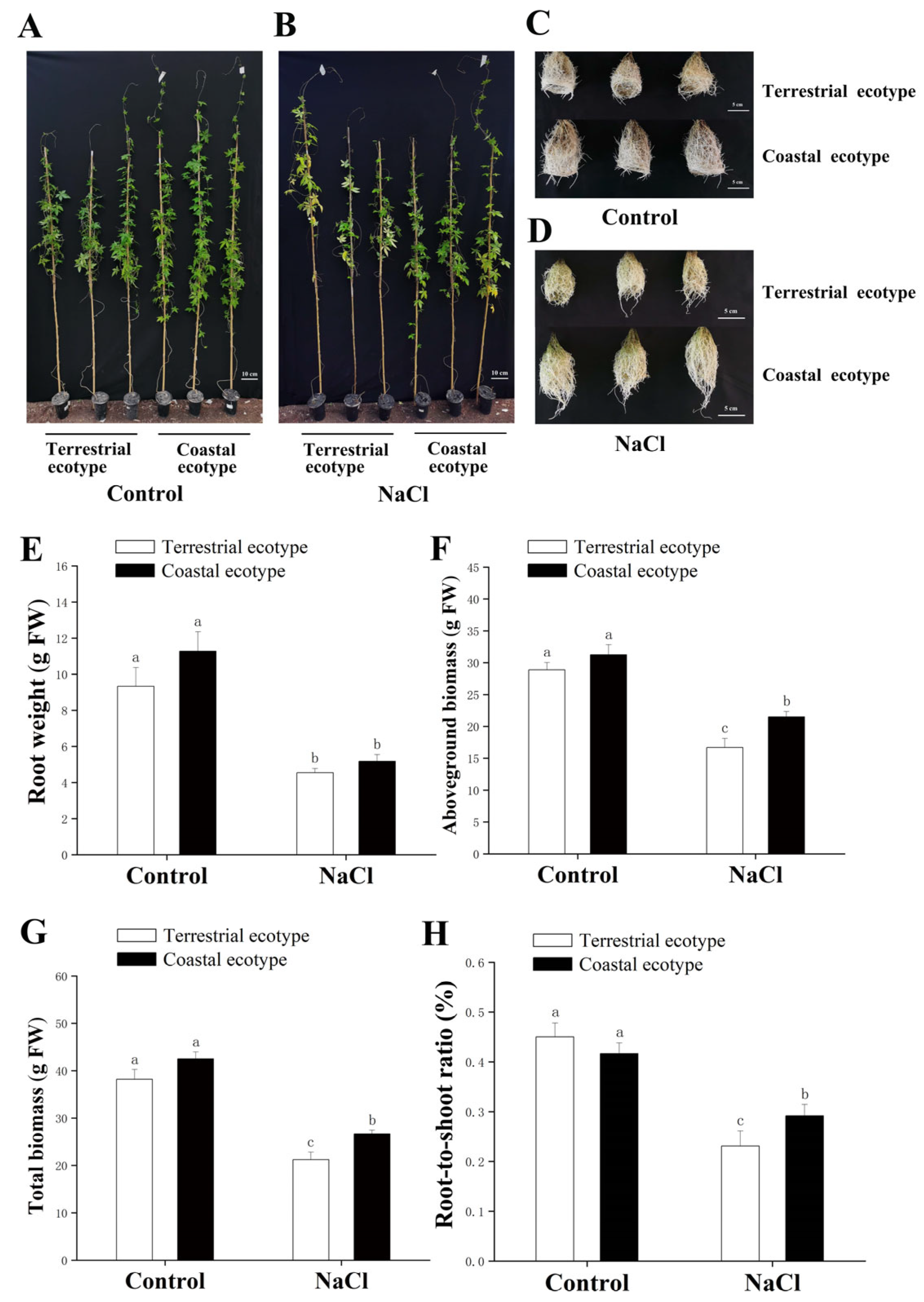
3.1. Growth of Both Lines under Salt Stress
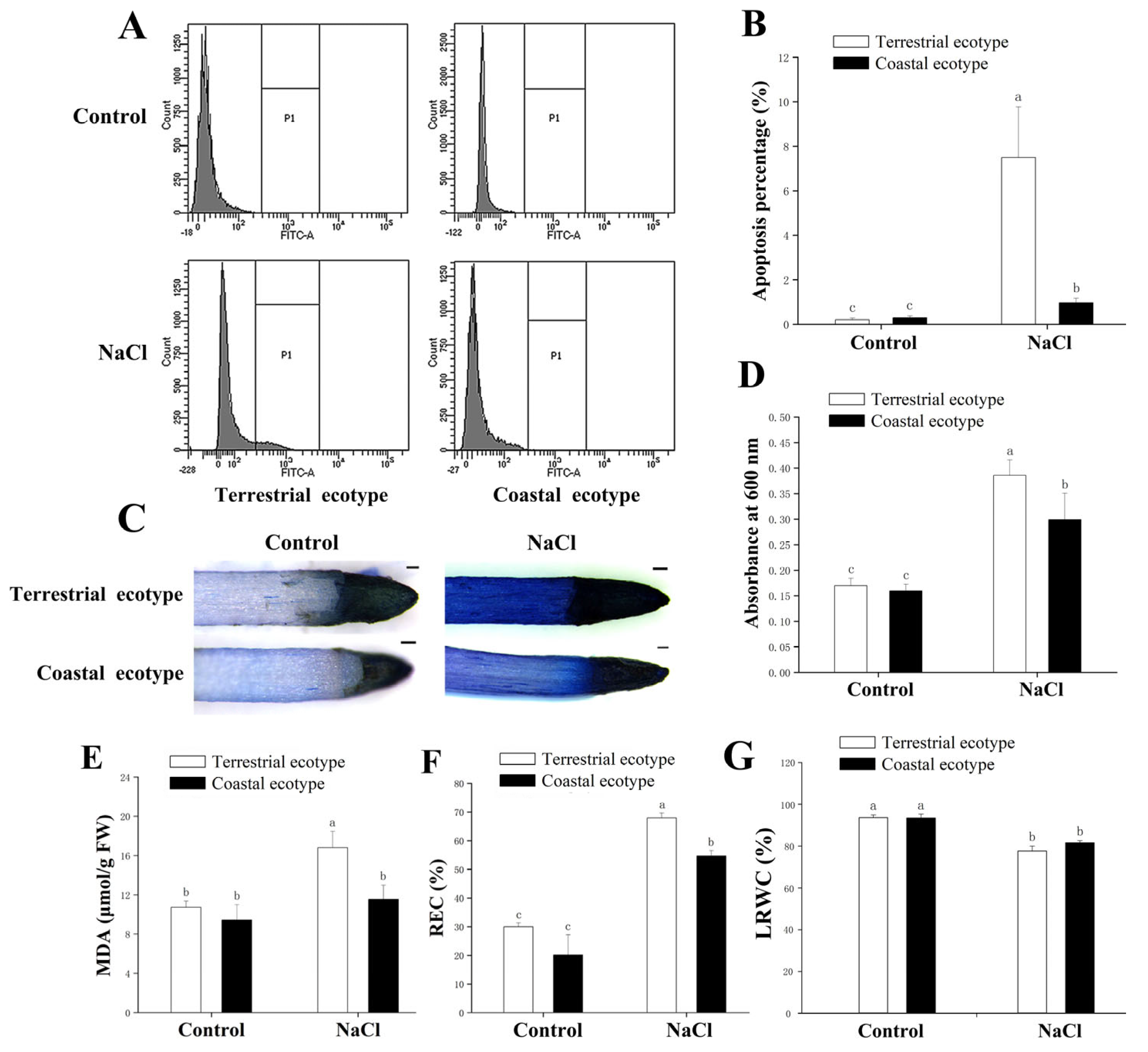
3.2. Cell Damage of Both Ecotypes under Salt Stress
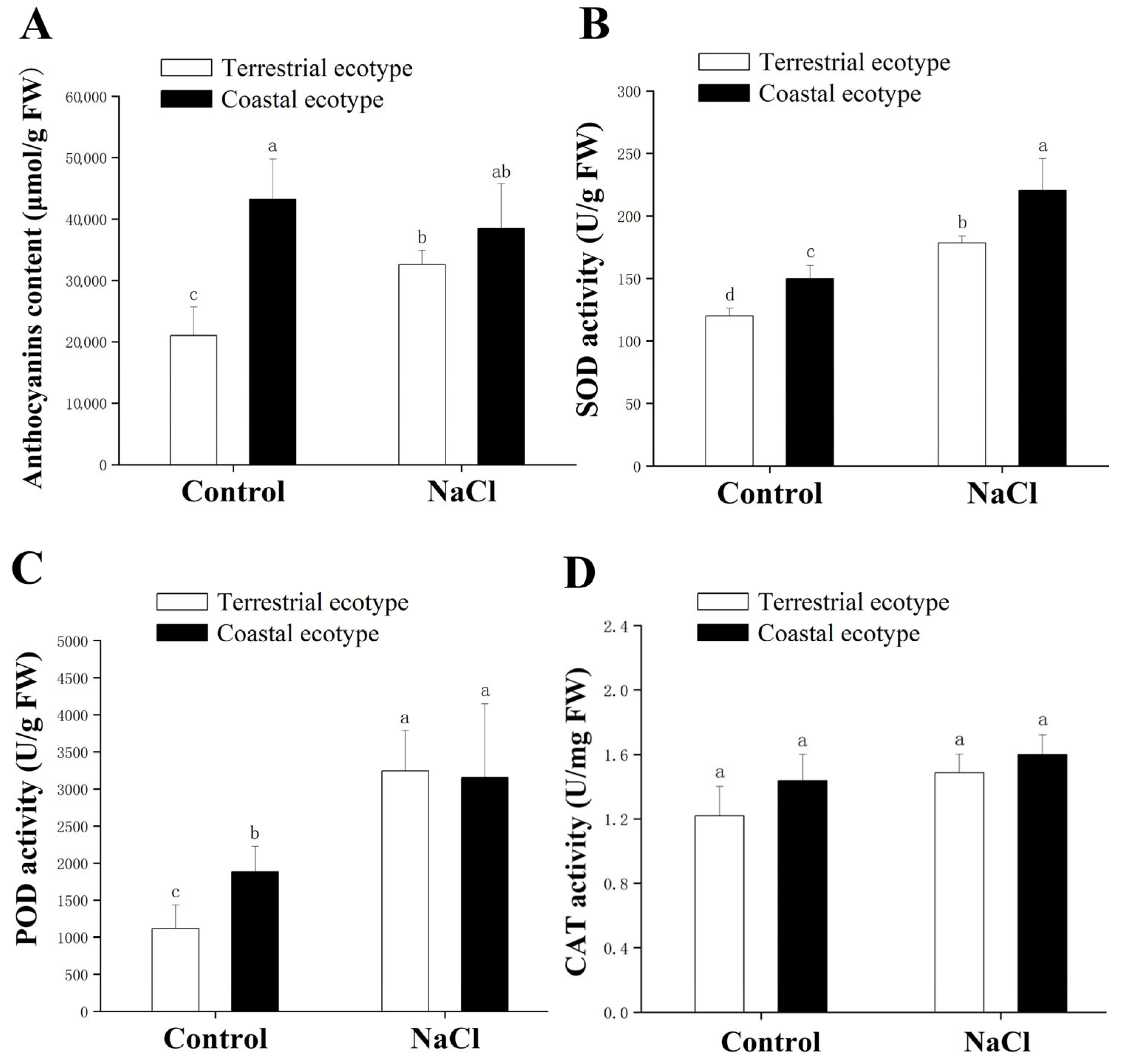
3.3. Antioxidant Capacity of I. cairica in Response to Salt Stress
3.4. Salt Rejection and Transport
3.5. Osmotic Regulation
3.6. Photosynthetic Performance
4. Discussion
4.1. Morphological Changes in the Two Lines under Salt Stress
4.2. Osmotic Adjustment Responses to Salt Stress
4.3. Salt Exclusion and Nutrient Management
4.4. Salt Stress Response to the Cell Damage and Antioxidant Capacity
4.5. Effects of Salt Stress on the Chlorophyll Content and Fluorescence Properties
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ding, H.; Xu, H.G.; Wu, J. Status and trends of invasive alien species in China. World Environ. 2009, 3, 39–40. [Google Scholar]
- Gu, Z.Y.; Wu, X.H.; Yang, G.; Chen, J.D. Status and Countermeasures against Invasion of Exotic Organisms in China. Jiangsu Agric. Sci. 2006, 6, 418–421. [Google Scholar]
- Bellard, C.; Cassey, P.; Blackburn, T.M. Alien species as a driver of recent extinctions. Biol. Lett. 2016, 12, 20150623. [Google Scholar] [CrossRef] [PubMed]
- Slingsby, J.A.; Merow, C.; Aiello-Lammens, M.; Allsopp, N.; Hall, S.; Kilroy Mollmann, H.; Turner, R.; Wilson, A.M.; Silander, J.A. Intensifying postfire weather and biological invasion drive species loss in a Mediterranean-type biodiversity hotspot. Proc. Natl. Acad. Sci. USA 2017, 114, 4697–4702. [Google Scholar] [CrossRef] [PubMed]
- Saenger, P.; Ragavan, P.; Sheue, C.R.; López-Portillo, J.; Yong, J.W.H.; Mageswaran, T. Mangrove biogeography of the Indo-Pacific. Sabkha Ecosyst. 2019, 43, 379–400. [Google Scholar]
- Wang, C.W.; Wong, S.L.; Liao, T.S.; Weng, J.H.; Chen, M.N.; Huang, M.Y.; Chen, C.I. Photosynthesis in response to salinity and submergence in two Rhizophoraceae mangroves adapted to different tidal elevations. Tree Physiol. 2022, 42, 1016–1028. [Google Scholar] [CrossRef]
- Ren, H.; Guo, Q.; Liu, H.; Li, J.; Zhang, Q.; Xu, H.; Xu, F. Patterns of alien plant invasion across coastal bay areas in southern China. J. Coast. Res. 2014, 295, 448–455. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, J.; Li, Y.; Liu, W.W.; Chen, Y.X.; Zhang, Y.H.; Li, Y.F. Spatially discontinuous relationships between salt marsh invasion and mangrove forest fragmentation. For. Ecol. Manag. 2021, 499, 0378–1127. [Google Scholar] [CrossRef]
- Liu, G.; Huang, Q.Q.; Lin, Z.G.; Huang, F.F.; Liao, H.X.; Peng, S.L. High tolerance to salinity and herbivory stresses may explain the expansion of Ipomoea cairica to salt marshes. PLoS ONE 2012, 7, e48829. [Google Scholar] [CrossRef] [PubMed]
- Bai, F.; Chisholm, R.; Sang, W.; Dong, M. Spatial risk assessment of alien invasive plants in China. Environ. Sci. Technol. 2013, 47, 7624–7632. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Chen, M.; Li, S. Isolation and Identification of Ipomoea cairica (L.) Sweet Gene IcSRO1 Encoding a SIMILAR TO RCD-ONE Protein, Which Improves Salt and Drought Tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2020, 21, 1017. [Google Scholar] [CrossRef] [PubMed]
- Morimaru, K.; Mitsutoshi, T.; Yasuo, I.; Kazutoshi, K.; Toshiyuki, O.; Nobuhide, F. High salinity leads to accumulation of soil organic carbon in mangrove soil. Chemosphere 2017, 177, 51–55. [Google Scholar] [CrossRef]
- Polash, M.A.S.; Sakil, M.A.; Hossain, M.A. Plants responses and their physiological and biochemical defense mechanisms against salinity: A review. Trop. Plant Res. 2019, 6, 250–274. [Google Scholar] [CrossRef]
- Wei, L.; Zhang, J.; Wei, S.; Hu, D.; Liu, Y.; Feng, L.; Li, C.; Qi, N.; Wang, C.; Liao, W. Nitric Oxide Enhanced Salt Stress Tolerance in Tomato Seedlings, Involving Phytohormone Equilibrium and Photosynthesis. Int. J. Mol. Sci. 2022, 23, 4539. [Google Scholar] [CrossRef]
- Hao, S.; Wang, Y.; Yan, Y.; Liu, Y.; Wang, J.; Chen, S. A Review on Plant Responses to Salt Stress and Their Mechanisms of Salt Resistance. Horticulturae 2021, 7, 132. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I. Optimising photosynthesis for environmental fitness. Funct Plant Biol. 2020, 47, iii–vii. [Google Scholar] [CrossRef]
- Guo, S.; Zhao, K. Possible Mechanism of NaCl Stress Inhibiting Photosynthesis of Maize Seedlings. Acta Plant Physiol. 2001, 27, 461–466. [Google Scholar]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef]
- Baker, N.R.; Rosenqvist, E. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef] [PubMed]
- Hessini, K.; Issaoui, K.; Ferchichi, S.; Saif, T.; Abdelly, C.; Siddique, K.H.M.; Cruz, C. Interactive effects of salinity and nitrogen forms on plant growth, photosynthesis and osmotic adjustment in maize. Plant Physiol. Biochem. 2019, 139, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Barbieri, G.; Vallone, S.; Orsini, F.; Paradiso, R.; De Pascale, S.; Negre-Zakharov, F.; Maggio, A. Stomatal density and metabolic determinants mediate salt stress adaptation and water use efficiency in basil (Ocimum basilicum L.). J. Plant Physiol. 2012, 169, 1737–1746. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Zhou, M.; Shabala, S. How Does Stomatal Density and Residual Transpiration Contribute to Osmotic Stress Tolerance? Plants 2023, 12, 494. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Leng, B.; Wang, B. Progress in Studying Salt Secretion from the Salt Glands in Recretohalophytes: How Do Plants Secrete Salt? Front. Plant Sci. 2016, 7, 977. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.Y.; Peng, S.L.; Chen, B.M.; Pang, J.X.; Chen, L.Y.; Xu, H.M.; Hou, Y.P. Do higher resource capture ability and utilization efficiency facilitate the successful invasion of native plants. Biol. Invasions 2011, 13, 869–881. [Google Scholar] [CrossRef]
- Hou, Y.P.; Peng, S.L.; Lin, Z.G.; Huang, Q.Q.; Ni, G.Y.; Zhao, N. Fast-growing and poorly shade-tolerant invasive species may exhibit higher physiological but not morphological plasticity compared with non-invasive species. Biol. Invasions 2014, 17, 1555–1567. [Google Scholar] [CrossRef]
- Tognon, G.B.; Petry, C.; Cuquel, F.L. Response to water deficit of Ipomoea cairica (L.) Sweet. Cienc. E Agrotecnolgia 2012, 36, 318–324. [Google Scholar] [CrossRef]
- Chen, M.H.; Mingling Cai, M.L.; Xiang, P.; Qin, Z.F.; Peng, C.L.; Li, S.S. Thermal adaptation of photosynthetic physiology of the invasive vine Ipomoea cairica (L.) enhances its advantage over native Paederia scandens (Lour.) Merr. in South China. Tree Physiol. 2023, 43, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, D.; Arnon, D. The Water-Culture Method for Growing Plants without Soil; University of California: Berkeley, CA, USA, 1950; pp. 13–57. [Google Scholar]
- Luo, B.; Wang, C.; Wang, X.; Zhang, H.; Zhou, Y.; Wang, W.; Song, P. Changes in photosynthesis and chlorophyll fluorescence in two soybean (Glycine max) varieties under NaCl stress. Int. J. Agric. Biol. Eng. 2021, 14, 76–82. [Google Scholar] [CrossRef]
- Marriboina, S.; Sengupta, D.; Kumar, S.; Reddy, A.R. Physiological and molecular insights into the high salinity tolerance of Pongamia pinnata (L.) Pierre, a potential biofuel tree species. Plant Sci. 2017, 258, 102–111. [Google Scholar] [CrossRef]
- Smith, V.R. A comparison between the H2SO4-H2O2-Li2SO4-Se oxidation method and alternative digestion procedures for plant nutrient analysis. Commun. Soil Sci. Plan 1979, 10, 1067–1077. [Google Scholar] [CrossRef]
- Tao, R.; Ding, J.; Li, C.; Zhu, X.; Guo, W.; Zhu, M. Evaluating and Screening of Agro-Physiological Indices for Salinity Stress Tolerance in Wheat at the Seedling Stage. Front. Plant Sci. 2021, 12, 646175. [Google Scholar] [CrossRef]
- Yuan, F.; Wang, X.; Zhao, B.; Xu, X.; Shi, M.; Leng, B.; Dong, X.; Lu, C.; Feng, Z.; Guo, J.; et al. The genome of the recretohalophyte Limonium bicolor provides insights into salt gland development and salinity adaptation during terrestrial evolution. Mol. Plant 2022, 15, 1024–1044. [Google Scholar] [CrossRef]
- Zhao, B.; Zhou, Y.; Jiao, X.; Wang, X.; Wang, B.; Yuan, F. Bracelet salt glands of the recretohalophyte Limonium bicolor: Distribution, morphology, and induction. J. Integr. Plant Biol. 2023, 65, 950–966. [Google Scholar] [CrossRef]
- Ding, F.; Chen, M.; Sui, N.; Wang, B.S. Ca2+ significantly enhanced development and salt-secretion rate of salt glands of Limonium bicolor under NaCl treatment. S. Afr. J. Bot. 2010, 76, 95–101. [Google Scholar] [CrossRef]
- Leng, B.Y.; Yuan, F.; Dong, X.X.; Wang, J.; Wang, B.S. Distribution pattern and salt excretion rate of salt glands in two recretohalophyte species of Limonium (Plumbaginaceae). S. Afr. J. Bot. 2018, 115, 74–80. [Google Scholar] [CrossRef]
- Fan, S.; Wu, H.; Gong, H.; Guo, J. The salicylic acid mediates selenium-induced tolerance to drought stress in tomato plants. Sci. Hortic. 2022, 300, 111092. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Kao, K.; Michayluk, M. A method for high-frequency intergeneric fusion of plant protoplasts. Planta 1974, 115, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.Y.; Fu, J.; Yin, X.B.; Qu, C.H.; Yang, C.J.; Li, H.Y.; He, G.Q.; Ni, J. Induction of Apoptosis in HepaRG Cell Line by Aloe-Emodin through Generation of Reactive Oxygen Species and the Mitochondrial Pathway. Cell. Physiol. Biochem. 2017, 42, 685–696. [Google Scholar] [CrossRef]
- Lequeux, H.; Hermans, C.; Lutts, S.; Verbruggen, N. Response to copper excess in Arabidopsis thaliana: Impact on the root system architecture, hormone distribution, lignin accumulation and mineral profile. Plant Physiol. Bioch. 2010, 48, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Khalid, M.F.; Hussain, S.; Anjum, M.A.; Ahmad, S.; Ali, M.A.; Ejaz, S.; Morillon, R. Better salinity tolerance in tetraploid vs. diploid volkamer lemon seedlings is associated with robust antioxidant and osmotic adjustment mechanisms. J. Plant Physiol. 2020, 244, 153071. [Google Scholar] [CrossRef]
- Liu, C.; Yan, M.; Huang, X.; Yuan, Z. Effects of salt stress on growth and physiological characteristics of pomegranate (Punica granatum L.) cuttings. Pak. J. Bot. 2018, 50, 457–464. [Google Scholar]
- Shahid, M.A.; Balal, R.M.; Khan, N.; Simón-Grao, S.; Alfosea-Simón, M.; Cámara-Zapata, J.M.; Mattson, N.S.; Garcia-Sanchez, F. Rootstocks influence the salt tolerance of Kinnow mandarin trees by altering the antioxidant defense system, osmolyte concentration, and toxic ion accumulation. Sci. Hortic. 2019, 250, 1–11. [Google Scholar] [CrossRef]
- Ghalati, R.E.; Shamili, M.; Homaei, A. Effect of putrescine on biochemical and physiological characteristics of guava (Psidium guajava L.) seedlings under salt stress. Sci. Hortic. 2020, 261, 108961–108967. [Google Scholar] [CrossRef]
- Hoch, W.; Singsaas, E.; Mccown, B. Anthocyanins facilitate nutrient recovery in autumn by shielding leaves from potentially damaging light levels. Plant Physiol. 2003, 133, 12961305. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhang, Y.; Han, W.; Feng, R.; Hu, Y.; Guo, J.; Gong, H. Silicon enhances water stress tolerance by improving root hydraulic conductance in Solanum lycopersicum L. Front. Plant Sci. 2016, 7, 196. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.K.; Huang, J.L. Principles and Techniques of Plant Physiological Biochemical Experiment, 3rd ed.; Higher Education Press: Beijing, China, 2015. [Google Scholar]
- Van Kleunen, M.; Schlaepfer, D.R.; Glaettli, M.; Fischer, M. Preadapted for invasiveness: Do species traits or their plastic response to shading differ between invasive and non-invasive plant species in their native range? J. Biogeogr. 2011, 38, 1294–1304. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Chourasia, K.N.; More, S.J.; Kumar, A.; Kumar, D.; Singh, B.; Bhardwaj, V.; Kumar, A.; Das, S.K.; Singh, R.K.; Zinta, G.; et al. Salinity responses and tolerance mechanisms in underground vegetable crops: An integrative review. Planta 2022, 255, 68. [Google Scholar] [CrossRef]
- Wang, Y.J.; Müller-Schärer, H.; van Kleunen, M.; Cai, A.M.; Zhang, P.; Yan, R.; Dong, B.C.; Yu, F.H. Invasive alien plants benefit more from clonal integration in heterogeneous environments than natives. New Phytol. 2017, 216, 1072–1078. [Google Scholar] [CrossRef] [PubMed]
- Amasino, R. Seasonal and developmental timing of flowering. Plant J. 2010, 61, 10011013. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Guan, R.; Chang, R.; Qiu, L. Correlation between Na+ Contents in Different Organs of Soybean and Salt Tolerance at the Seedling Stage. Acta Agron. Sin. 2011, 37, 1266–1273. [Google Scholar] [CrossRef]
- Menezes, R.V.; Azevedo Neto, A.D.D.; Ribeiro, M.D.O.; Cova, A.M.W. Growth and contents of organic and inorganic solutes in amaranth under salt stress. Pesqui. Agropecuária Trop. 2017, 47, 22–30. [Google Scholar] [CrossRef]
- Sahin, U.; Ekinci, M.; Ors, S.; Turan, M.; Yildiz, S.; Yildirim, E. Effects of individual and combined effects of salinity and drought on physiological, nutritional and biochemical properties of cabbage (Brassica oleracea var. capitata). Sci. Hortic. 2018, 240, 196–204. [Google Scholar] [CrossRef]
- Shirazi, M.; Khan, M.; Mahboob, W.; Khan, M.A.; Shereen, A.; Mujtaba, S.; Asad, A. Inconsistency in salt tolerance of some wheat (Triticum aestivium L.) genotypes evaluated under various growing environments. Pak. J. Bot. 2018, 50, 471–479. [Google Scholar]
- Zhang, H.; Gao, X.R.; Zhi, Y.H.; Li, X.; Zhang, Q.; Niu, J.B.; Wang, J.; Zhai, H.; Zhao, N.; Li, J.G.; et al. A non-tandem CCCH-type zinc-finger protein, IbC3H18, functions as a nuclear transcriptional activator and enhances abiotic stress tolerance in sweet potato. New Phytol. 2019, 223, 1918–1936. [Google Scholar] [CrossRef]
- Li, G.; Ye, Y.X.; Ren, X.Q.; Qi, M.Y.; Zhao, H.Y.; Zhou, Q.; Chen, X.H.; Wang, J.; Yuan, C.Y.; Wang, F.B. The rice Aux/IAA transcription factor gene OsIAA18 enhances salt and osmotic tolerance in Arabidopsis. Biol. Plant 2020, 64, 454–464. [Google Scholar] [CrossRef]
- Kang, C.; He, S.Z.; Zhai, H.; Li, R.J.; Zhao, N.; Liu, Q.C. A sweetpotato auxin response factor gene (IbARF5) is involved in carotenoid biosynthesis and salt and drought tolerance in transgenic Arabidopsis. Front. Plant Sci. 2018, 9, 1307. [Google Scholar] [CrossRef]
- Munns, R.; Passioura, J.B.; Colmer, T.D.; Byrt, C.S. Osmotic adjustment and energy limitations to plant growth in saline soil. New Phytol. 2020, 225, 1091–1096. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, W.; da Silva, J.A.T.; Liu, X.; Duan, J. Rice histone deacetylase HDA704 positively regulates drought and salt tolerance by controlling stomatal aperture and density. Planta 2021, 254, 79. [Google Scholar] [CrossRef]
- Hameed, M.; Ashraf, M.; Naz, N. Anatomical adaptations to salinity in cogon grass [Imperata cylindrica (L.) Raeuschel] from the Salt Range, Pakistan. Plant Soil 2009, 322, 229–238. [Google Scholar] [CrossRef]
- Yuan, F.; Wang, B. Adaptation of Recretohalophytes to Salinity. In Handbook of Halophytes; Springer: Cham, Switzerland, 2020; pp. 1–21. [Google Scholar]
- Wei, X.; Yan, X.; Yang, Z. Salt glands of recretohalophyte Tamarix under salinity: Their evolution and adaptation. Ecol. Evol. 2020, 10, 9384–9395. [Google Scholar] [CrossRef]
- Shabala, S.; Demidchik, V.; Shabala, L.; Cuin, T.; Smith, S.; Miller, A.; Davies, J.; Newman, I. Extracellular Ca2+ ameliorates NaCl-induced K+ loss from Arabidopsis root and leaf cells by controlling plasma membrane K+-permeable channels. Plant Physiol. 2006, 141, 1653–1665. [Google Scholar] [CrossRef]
- Zhang, Y.; Fang, J.; Wu, X.; Dong, L. Na+/K+ balance and transport regulatory mechanisms in weedy and cultivated rice (Oryza sativa L.) under salt stress. BMC Plant Biol. 2018, 18, 375. [Google Scholar] [CrossRef]
- Fang, S.; Hou, X.; Liang, X. Response mechanisms of plants under saline-alkali stress. Front. Plant Sci. 2021, 12, 667458. [Google Scholar] [CrossRef]
- Chen, H.N.; Tao, L.Y.; Shi, J.M.; Han, X.R.; Cheng, X.G. Exogenous salicylic acid signal reveals an osmotic regulatory role in priming the seed germination of Leymus chinensis under salt-alkali stress. Environ. Exp. Bot. 2021, 188, 104498. [Google Scholar]
- Rahman, A.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Calcium Supplementation Improves Na+/K+ Ratio, Antioxidant Defense and Glyoxalase Systems in Salt-Stressed Rice Seedlings. Front. Plant Sci. 2016, 7, 609. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.Q.; Li, H.; Zhu, Y.H.; Li, S.J. Comparative physiological response of sainfoin (Onobrychis viciaefolia) seedlings to alkaline and saline-alkaline stress. J. Anim. Plant Sci. 2021, 31, 1028–1035. [Google Scholar]
- Qin, Y.; Bai, J.; Wang, Y.; Liu, J.; Hu, Y.; Dong, Z.; Ji, L. Comparative effects of salt and alkali stress on photosynthesis and root physiology of oat at anthesis. Arch. Biol. Sci. 2018, 70, 329–338. [Google Scholar] [CrossRef]
- Wang, R.; Wang, C.; Feng, Q.; Liou, R.M.; Lin, Y.F. Biological inoculant of salt-tolerant bacteria for plant growth stimulation under different saline soil conditions. J. Microbiol. Biotechnol. 2021, 31, 398–407. [Google Scholar] [CrossRef]
- Zelm, E.V.; Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Xue, X.; Huan, C.; Peng, F.; Liao, J.; Ma, S.; You, Q.; Wang, T. Salt Tolerance Strategies of Nitraria tangutorum Bobr. and Elaeagnus angustifolia Linn. Determine the Inoculation Effects of Microorganisms in Saline Soil Conditions. Agronomy 2022, 12, 913. [Google Scholar] [CrossRef]
- Umar, S.; Diva, I.; Anjum, N.A.; Iqbal, M.; Ahmad, I.; Pereira, E. Potassium-induced alleviation of salinity stress in Brassica campestris L. Cent. Eur. J. Biol. 2011, 6, 1054–1063. [Google Scholar] [CrossRef]
- Ratnayake, M.; Leonard, R.; Menge, J. Root exudation in relation to supply of phosphorus and its possible relevance to mycorrhizal formation. New Phytol. 1978, 81, 543–552. [Google Scholar] [CrossRef]
- Yang, Z.; Li, J.L.; Liu, L.N. Photosynthetic regulation under salt stress and salt-tolerance mechanism of sweet sorghum. Front. Plant Sci. 2020, 10, 1722. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Feng, X.X.; Gao, J.J.; Peng, R.H.; Yao, Q.H.; Wang, L.J. VvMYBA6 in the promotion of anthocyanin biosynthesis and salt tolerance in transgenic Arabidopsis. Plant Biotechnol. Rep. 2017, 11, 299–314. [Google Scholar] [CrossRef]
- Naing, A.H.; Kim, C.K. Abiotic stress-induced anthocyanins in plants: Their role in tolerance to abiotic stresses. Physiol. Plant 2021, 172, 1711–1723. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, J.; Yan, D.; Yuan, H.; Wang, Y.; He, Y.; Wang, X.; Li, Z.; Mei, J.; Hu, M. Exogenous spermidine improves salt tolerance of pecan-grafted seedlings via activating antioxidant system and inhibiting the enhancement of Na+/K+ ratio. Acta Physiol. Plant 2020, 42, 83. [Google Scholar] [CrossRef]
- Banu, M.N.A.; Hoque, M.A.; Watanabe-Sugimoto, M. Proline and glycinebetaine induce antioxidant defense gene expression and suppress cell death in cultured tobacco cells under salt stress. J. Plant Physiol. 2009, 166, 146–156. [Google Scholar] [CrossRef]
- Naing, A.H.; Park, K.I.; Ai, T.N.; Chung, M.Y.; Han, J.S.; Kang, Y.W.; Lim, K.B.; Kim, C.K. Overexpression of snapdragon Delila (Del) gene in tobacco enhances anthocyanin accumulation and abiotic stress tolerance. BMC Plant Biol. 2017, 17, 65. [Google Scholar] [CrossRef] [PubMed]
- Katsuhara, M.; Kawasaki, T. Salt Stress Induced Nuclear and DNA Degradation in Meristematic Cells of Barley Roots. Plant Cell Physiol. 1996, 37, 169–173. [Google Scholar] [CrossRef]
- Ameisen, J. On the origin, evolution, and nature of programmed cell death: A timeline of four billion years. Cell Death Differ. 2002, 9, 367–393. [Google Scholar] [CrossRef]
- Zhao, Y.; Wu, M.; Deng, P.; Zhou, X.W.; Huang, S.Y. Effects of salt stress on growth and chlorophyII fluorescence parameters of Siraitia grosvenorii seedlings. South China Fruits 2021, 50, 103–107. [Google Scholar] [CrossRef]
- Athar, H.R.; Ashraf, M. Photosynthesis under Drought Stress. In Handbook of Photosynthesis, 2nd ed.; Pessarakli, M., Ed.; CRC Press: Boca Raton, FL, USA, 2005; pp. 793–809. [Google Scholar]
- Ehsen, S.; Abideen, Z.; Rizvi, R.F.; Gulzar, S.; Aziz, I.; Gul, B.; Khan, M.A.; Ansari, R. Ecophysiological adaptations and anti-nutritive status of sustainable cattle feed Haloxylon stocksii under saline conditions. Flora 2019, 257, 151425. [Google Scholar] [CrossRef]
- Abideen, Z.; Koyro, H.W.; Hussain, T.; Rasheed, A.; Alwahibi, M.S.; Elshikh, M.S.; Hussain, M.I.; Zulfiqar, F.; Mansoor, S.; Abbas, Z. Biomass Production and Predicted Ethanol Yield Are Linked with Optimum Photosynthesis in Phragmites karka under Salinity and Drought Conditions. Plants 2022, 11, 1657. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, W.; Ahmed, I.M.; Chen, X.; Cao, F.; Zhu, S.; Wu, F. Genotypic differences in photosynthetic performance, antioxidant capacity, ultrastructure and nutrients in response to combined stress of salinity and Cd in cotton. Biometals 2015, 28, 1063–1078. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, J.; Yuan, B.; Li, W.; Xie, X.; Chen, M.; Xiong, T. Ipomoea cairica (L.) from Mangrove Wetlands Acquired Salt Tolerance through Phenotypic Plasticity. Forests 2024, 15, 358. https://doi.org/10.3390/f15020358
Zou J, Yuan B, Li W, Xie X, Chen M, Xiong T. Ipomoea cairica (L.) from Mangrove Wetlands Acquired Salt Tolerance through Phenotypic Plasticity. Forests. 2024; 15(2):358. https://doi.org/10.3390/f15020358
Chicago/Turabian StyleZou, Jiatong, Benqi Yuan, Weihua Li, Xiaoting Xie, Minghao Chen, and Tiantian Xiong. 2024. "Ipomoea cairica (L.) from Mangrove Wetlands Acquired Salt Tolerance through Phenotypic Plasticity" Forests 15, no. 2: 358. https://doi.org/10.3390/f15020358