


Different Species Proportions Influence Silvicultural Heterogeneity of Trees in a Restoration of a Ombrophilous Dense Forest in Lowlands
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
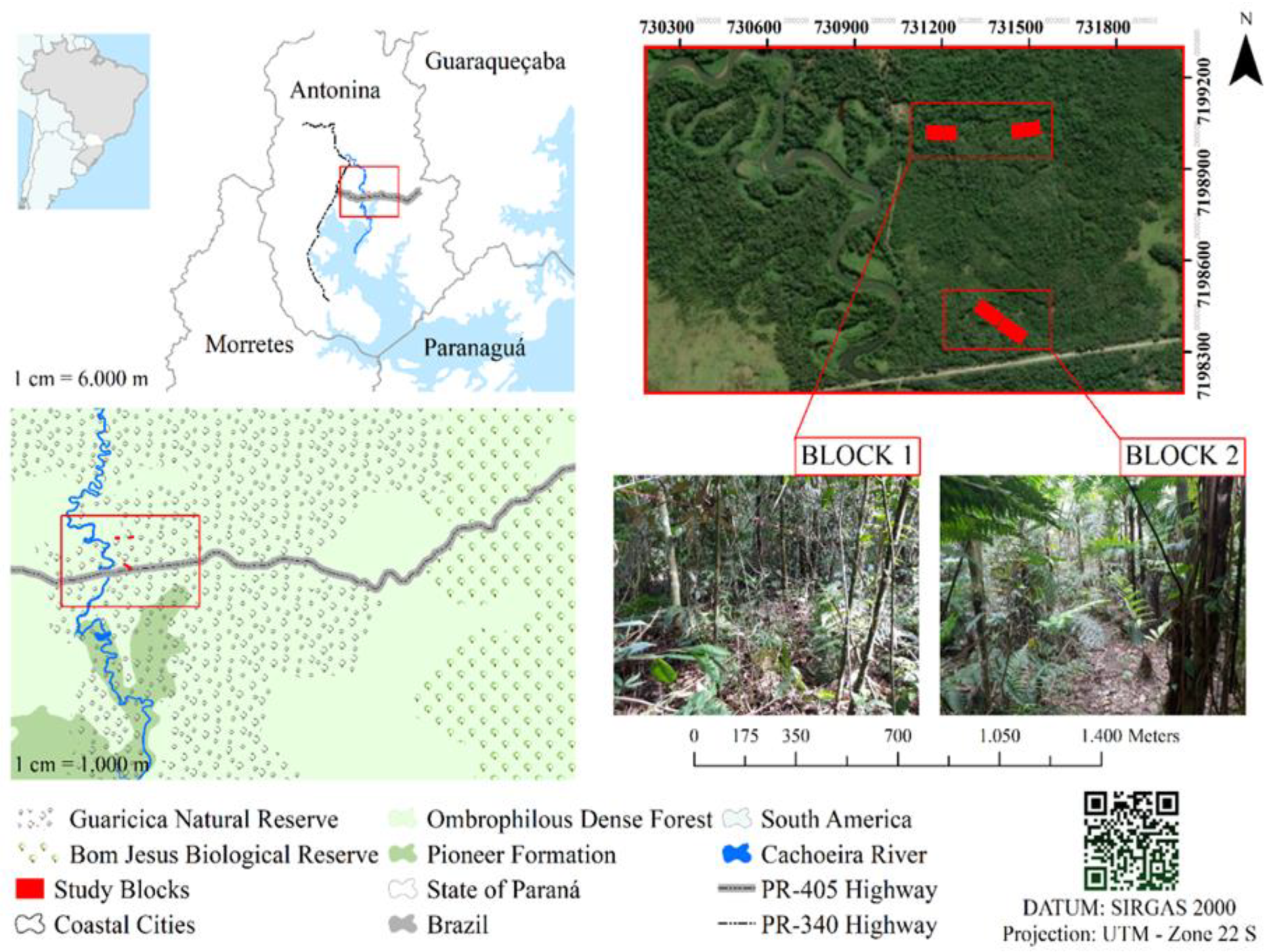
2.1. Characterization of the Study Area
2.2. Implementation of the Restoration Area
2.3. Data Collection and Analysis after 13 Years
3. Results
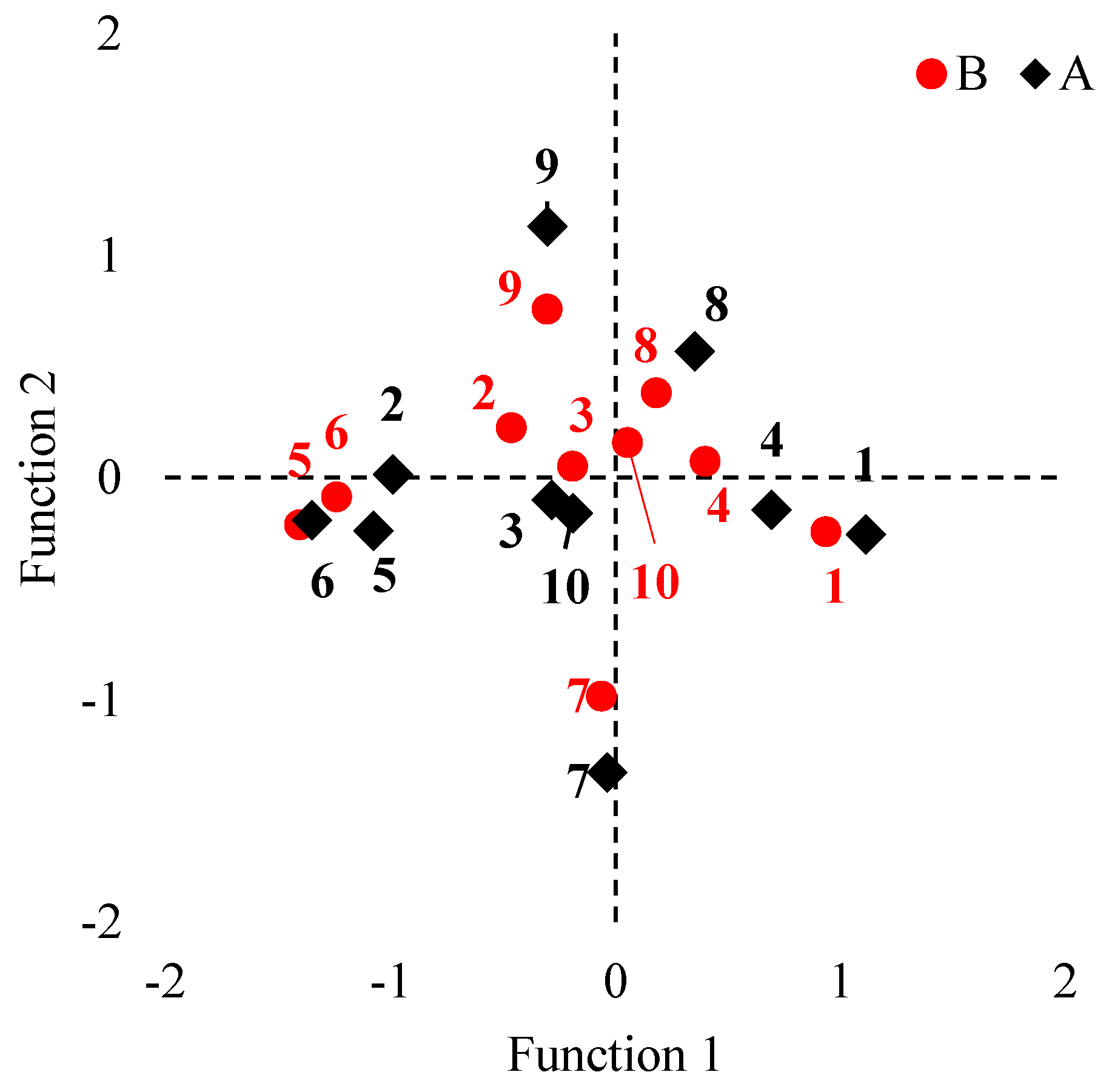
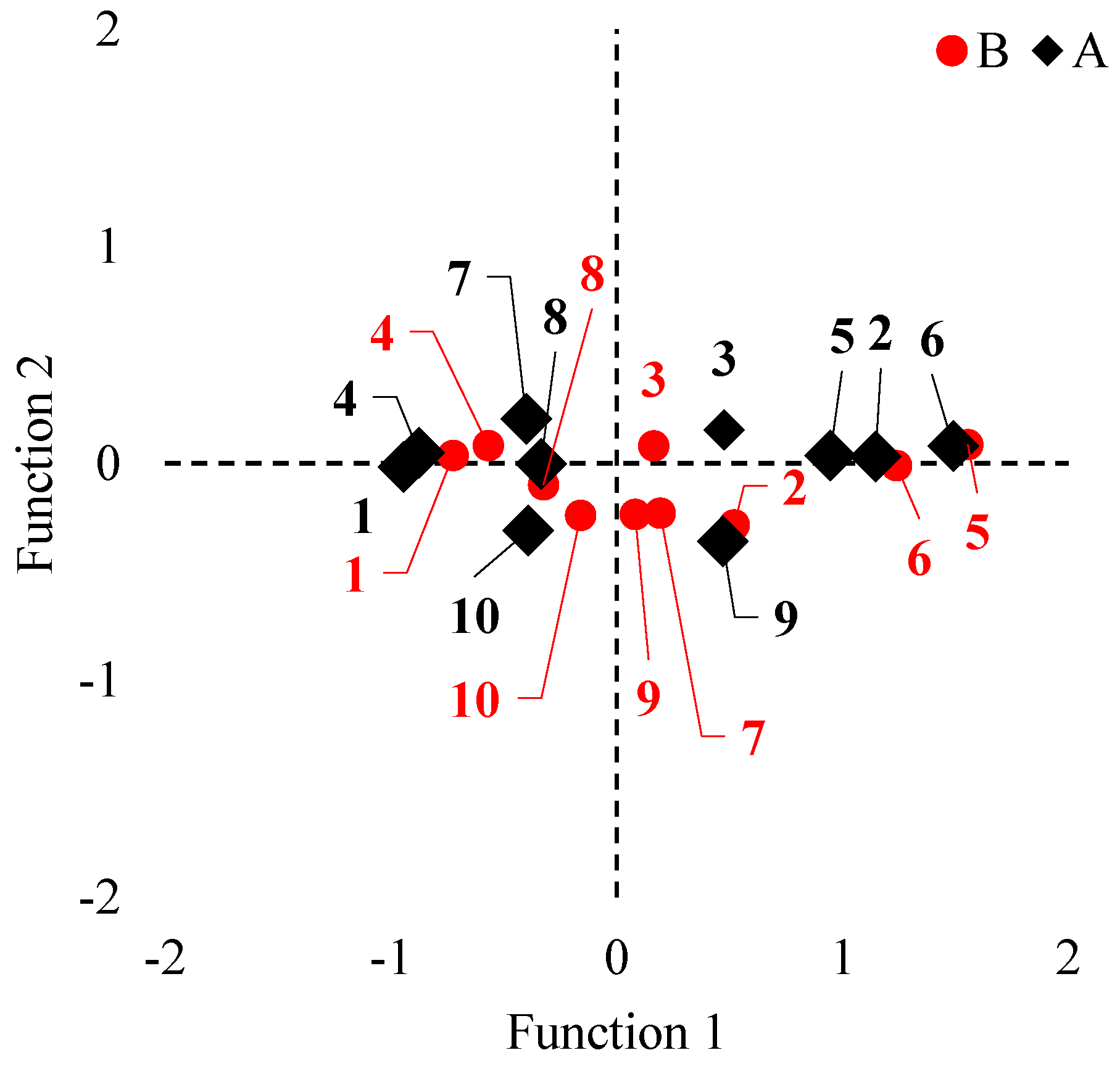
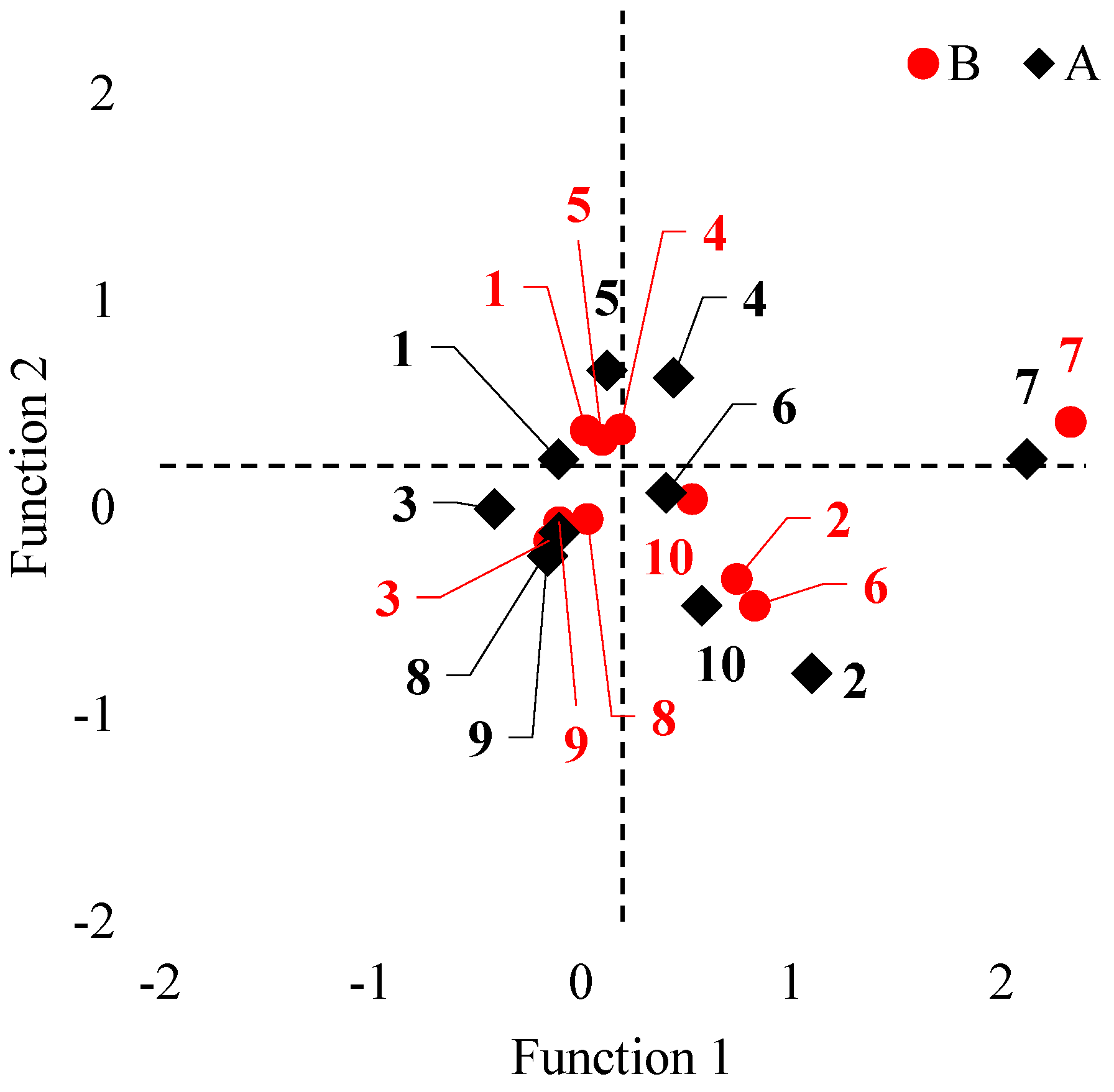
3.1. Multivariate Analysis of Variance (MANOVA)
3.2. VC1
3.3. VC2
3.4. VC3
4. Discussion
5. Conclusions
- Species 1, 4, and 8 are highly competitive to the presence of exotic grasses and are resilient to the transitional circumstances of the restoration process. In addition, they contributed to canopy formation and forest health associated with lower maintenance costs.
- Species 2, 3, 5, 6, and 9 are suitable for use in ecological restoration and contribute to different lower forest strata.
- Species 7 and 10 played an important role in the initial overlay, and their senescence indicates the evolution of the restoration process.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Test | VC1 | VC2 | VC3 | |||
|---|---|---|---|---|---|---|---|
| Value | α | Value | α | Value | α | ||
| Proportions | 0.992 | 0.003 | 0.995 | 0.009 | 0.993 | 0.005 | |
| 0.008 | 0.003 | 0.005 | 0.009 | 0.007 | 0.005 | ||
| Species | 0.492 | <0.001 | 0.596 | <0.001 | 0.732 | <0.001 | |
| 0.657 | <0.001 | 0.658 | <0.001 | 0.223 | <0.001 | ||
| Interaction | 0.975 | 0.008 | 0.967 | <0.001 | 0.957 | <0.001 | |
| 0.014 | 0.002 | 0.030 | <0.001 | 0.022 | <0.001 | ||
| VC | P | F | SEM | |||||
|---|---|---|---|---|---|---|---|---|
| F | 1 | 2 | 3 | F | 1 | 2 | ||
| VC 1 | A | AU | 0.755 | 0.263 | 0.069 | TH | 0.890 * | −0.061 |
| CV% | 69.50 | 93.70 | 100 | MD | 0.842 * | −0.254 | ||
| CC | 0.656 | 0.456 | 0.253 | CH | 0.589 | 0.804 * | ||
| B | AU | 0.634 | 0.115 | 0.040 | MD | 0.869 * | −0.313 | |
| CV% | 80.30 | 95.00 | 100 | TH | 0.854 * | 0.086 | ||
| CC | 0.623 | 0.322 | 0.196 | CH | 0.579 | 0.806 * | ||
| VC 2 | A | AU | 0.762 | 0.016 | SP | 0.939 * | −0.343 | |
| CV% | 97.90 | 100 | CL | 0.770 * | 0.638 | |||
| CC | 0.658 | 0.126 | ||||||
| B | AU | 0.682 | 0.014 | SP | 0.948 * | −0.319 | ||
| CV% | 97.90 | 100 | CL | 0.807 * | 0.591 | |||
| CC | 0.637 | 0.119 | ||||||
| VC 3 | A | AU | 0.258 | 0.156 | 0.062 | SH | 0.843 * | −0.379 |
| CV% | 54.20 | 87.00 | 100 | SQ | 0.585 | 0.792 * | ||
| CC | 0.453 | 0.368 | 0.241 | CQ | 0.467 | −0.460 | ||
| B | AU | 0.213 | 0.066 | 0.028 | SH | 0.913 * | −0.164 | |
| CV% | 69.50 | 90.90 | 100 | SQ | 0.486 | 0.843 * | ||
| CC | 0.419 | 0.248 | 0.164 | CQ | 0.514 | −0.405 | ||
| VC | P | F | Species | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |||
| VC1 | A | 1 | 1.111 | −0.989 | −0.285 | 0.693 | −1.076 | −1.351 | −0.037 | 0.352 | −0.303 | −0.190 |
| 2 | −0.257 | 0.015 | −0.101 | −0.146 | −0.240 | −0.192 | −1.328 | 0.568 | 1.132 | −0.160 | ||
| B | 1 | 0.934 | −0.464 | −0.191 | 0.398 | −1.404 | −1.240 | −0.063 | 0.181 | −0.305 | 0.052 | |
| 2 | −0.243 | 0.224 | 0.052 | 0.074 | −0.213 | −0.086 | −0.985 | 0.382 | 0.758 | 0.158 | ||
| VC2 | A | 1 | −0.944 | 1.146 | 0.476 | −0.875 | 0.945 | 1.490 | −0.400 | −0.335 | 0.470 | −0.392 |
| 2 | −0.017 | 0.032 | 0.154 | 0.050 | 0.037 | 0.080 | 0.206 | −0.002 | −0.360 | −0.311 | ||
| B | 1 | −0.724 | 0.521 | 0.164 | −0.568 | 1.555 | 1.239 | 0.193 | −0.324 | 0.082 | −0.159 | |
| 2 | 0.037 | −0.284 | 0.080 | 0.082 | 0.087 | −0.011 | −0.229 | −0.099 | −0.235 | −0.240 | ||
| VC3 | A | 1 | −0.307 | 0.897 | −0.611 | 0.240 | −0.076 | 0.206 | 1.920 | −0.304 | −0.359 | 0.375 |
| 2 | 0.033 | −1.002 | −0.207 | 0.426 | 0.463 | −0.129 | 0.034 | −0.321 | −0.434 | −0.675 | ||
| B | 1 | −0.176 | 0.541 | −0.350 | −0.013 | −0.099 | 0.627 | 2.128 | −0.168 | −0.302 | 0.328 | |
| 2 | 0.173 | −0.546 | −0.361 | 0.177 | 0.126 | −0.675 | 0.213 | −0.255 | −0.271 | −0.160 | ||
References
- Engel, V.L.; Parrota, J.A. Definindo a restauração ecológica: Tendências e mundiais. In Restauração Ecológica de Ecossistemas Naturais; Kageyama, P.Y., de Oliveira, R.E., de Moraes, L.F.D., Engel, V.L., e Mendes, F.B.G., Eds.; FEPAF: Botucatu, Brasil, 2003; pp. 1–26. [Google Scholar]
- Mcdonald, T.; Gann, G.D.; Jonson, J.; Dixon, K.W. International Standards for the Practice of Ecological Restoration—Including Principles and Key Concepts, 1st ed.; Society for Ecological Restoration: Washington, DC, USA, 2016. [Google Scholar]
- Society for Ecological Restoration International Science (SER); Policy Working Group (PWG). The SER International Primer on Ecological Restoration; Society for Ecological Restoration International: Tucson, AZ, USA, 2004; pp. 1–14. [Google Scholar]
- Prach, K. Tertiary succession—Do we really need the new term? Response to Rapson (2023). Restor. Ecol. 2023, 1, e13892. [Google Scholar] [CrossRef]
- Ren, J.W.F.; Coffman, G.C. Integrating the resilience concept into ecosystem restoration. Restor. Ecol. 2023, e13907. [Google Scholar] [CrossRef]
- Barragán, G.; Wang, T.; Rhemtulla, J.M. Árvores plantadas sob uma promessa de restauração global têm futuros mistos sob a mudança climática. Restor. Ecol. 2023, 31, e13764. [Google Scholar] [CrossRef]
- Evans, K.; Meli, P.; Zamora-Cristales, R.; Schweizer, D.; Méndez-Toribio, M.; Gómez-Ruiz, P.A.; Guariguata, M.R. Drivers of success in collaborative monitoring in forest landscape restoration: An indicative assessment from Latin America. Restor. Ecol. 2022, e13803. [Google Scholar] [CrossRef]
- Pierri, N.; Ângulo, R.J.; Souza, M.C.; Kim, M.K. A ocupação e o uso do solo no litoral paranaense: Condicionantes, conflitos e tendencias. Desenvolv. Meio Ambiente 2006, 13, 137–167. [Google Scholar] [CrossRef]
- Raynaut, C.; Zanoni, M.; Lana, P.C. O desenvolvimento sustentável regional: O que proteger? Quem desenvolver? Desenvolv. Meio Ambiente 2018, 47, 275–289. [Google Scholar] [CrossRef]
- Arraes, R.A.; Mariano, F.Z.; Simonassi, A.G. Causas do desmatamento no Brasil e seu ordenamento no contexto mundial. Rev. Econ. Sociol. Rural. 2012, 50, 119–140. [Google Scholar] [CrossRef]
- Cheung, K.C.; Marques, M.C.M.; Liebsch, D. Relação entre a presença de vegetação herbácea e a regeneração natural de espécies lenhosas em pastagens abandonadas na Floresta Ombrófila Densa do Sul do Brasil. Acta Bot. Bras. 2009, 23, 1048–1056. [Google Scholar] [CrossRef]
- Instituto de Pesquisa Selvagem e Educação Ambiental (SPVS) (Org.). Reservas Naturais da SPVS—20 anos de História; InVerso: Curitiba, Brazil, 2020. [Google Scholar]
- Rodrigues, R.R.; Gandolfi, S.; Nave, A.G.; Aronson, J.; Barreto, T.E.; Vidal, C.Y.; Brancalion, P.H.S. Large-scale ecological restoration of high-diversity tropical forests in SE Brazil. For. Ecol. Manag. 2011, 261, 1605–1613. [Google Scholar] [CrossRef]
- Souza, P.B.D.; Meira Neto, J.A.A.; Souza, A.L.D. Diversidade florística e estrutura fitossociológica de um gradiente topográfico em Floresta Estacional Semidecidual Submontana, MG. Cerne 2013, 19, 489–499. [Google Scholar] [CrossRef]
- Rodrigues, L.A.; Carvalho, D.A.; Oliveira Filho, A.T.; Curi, N. Efeitos de solos e topografia sobre a distribuição de espécies arbóreas em um fragmento de floresta estacional semidecidual, em Luminárias, MG. Rev. Árvore 2007, 31, 25–35. [Google Scholar] [CrossRef]
- Rosa, M.R.; Brancalion, P.H.S.; Crouzeilles, R.; Tambosi, L.R.; Piffer, P.R.; Lenti, F.E.B.; Hirota, M.; Santiami, E.; Metzger, J.P. Hidden destruction of older forests threatens Brazil’s Atlantic Forest and challenges restoration programs. Sci. Adv. 2021, 7, eabc4547. [Google Scholar] [CrossRef]
- Empresa Brasileira de Pesquisas Agropecuária—EMBRAPA Solos. Sistema Brasileiro de Classificação de Solos, 5th ed.; Santos, H.G., Jacomine, P.K.T., Anjos, L.H.C., Oliveira, V.A., Lumbreras, J.F., Coelho, M.R., Almeida, J.A., Araújo Filho, J.C., Oliveira, J.B., Cunha, T.J.F., et al., Eds.; Embrapa: Brasília, Brazil, 2018; Available online: https://www.embrapa.br/solos/sibcs (accessed on 14 June 2022).
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Gonçalves, J.L.M.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef]
- Roderjan, C.V.; Galvão, F.; Kuniyoshi, Y.S.; Hatschbach, G.G. As unidades fitogeográficas do estado do Paraná. Ciência Ambiente 2002, 24, 75–92. [Google Scholar]
- Carpanezzi, A.A.; Carpanezzi, O.T.B. Espécies nativas recomendadas para recuperação ambiental no Estado do Paraná: Em solos não degradados. Colombo, Paraná, Brasil. Colombo: Embrapa Florestas, Documentos, 136. 2006. 57. Available online: http://www.bibliotecaflorestal.ufv.br/handle/123456789/5951 (accessed on 14 June 2022).
- IBM Corp. Lançado em 2021. In IBM SPSS Statistics para Windows, Versão 28.0; IBM Corp: Endicott, NY, USA, 2021. [Google Scholar]
- Higuchi, P.; Silva, A.C.; Ferreira, T.S.; Souza, S.T.; Gomes, J.P.; Silva, K.M.; Santos, K.F.; Linke, C.; Paulino, O.S. Influência de variáveis ambientais sobre o padrão estrutural e florístico do componente arbóreo, em um fragmento de Floresta Ombrófila Mista Montana em Lages, SC. Ciência Florest. 2012, 22, 79–90. [Google Scholar] [CrossRef]
- Silva, J.O.; Silva, A.C.; Higuchi, P.; Mafra, A.L.; Loebens, R.; Rodrigues Júnior, L.C.; Rosa, A.D.; Lima, C.L.; Buzzi Júnior, F. Heterogeneidade ambiental e regeneração natural em uma Floresta Ombrófila Mista Aluvial. Sci. For. 2016, 44, 787–797. [Google Scholar] [CrossRef]
- Schaffer, L.H.; Mattar, E.A.; Nakajima, N.V.; Silva, A.S.; Borges, R.A.; Borges, A.V.P.; Carpanezzi, A.A.; Neves, E.I.M.; Angelo, A.C.; Britez, R.M. Crescimento de espécies arbóreas nativas em recuperação de área degradada no litoral do Paraná. Pesqui. Florest. Bras. 2020, 40, 1–9. [Google Scholar] [CrossRef]
- Siminski, A.; Zambiazi, D.C.; Santos, K.L.; Fantini, A.C. Dynamics of Natural Regeneration: Implications for Landscape Restoration in the Atlantic Forest, Brazil. Front. For. Glob. Chang. 2021, 4, 576908. [Google Scholar] [CrossRef]
- Piaia, B.B.; Rovedder, A.P.M.; Procknow, D.; Camargo, B. Avaliação de indicadores ecológicos na restauração por plantio em núcleo com diferentes idades. Ciênc. Florest. 2021, 31, 1512–1534. [Google Scholar] [CrossRef]
- Silva, F.A.M.; Leite, G.S.; Bim, O.J.B. Florística e fitossociologia de áreas de restauração com diferentes históricos de uso. Pesqui. Florest. Bras. 2021, 41, 1–14. [Google Scholar] [CrossRef]
- Freire, J.M.; Jesus, E.C.; Rouws, J.R.C.; Faria, S.M.; Zilli, J.E. Efeito do substrato sobre o crescimento de mudas de Mimosa bimucronata inoculadas com estirpes de rizóbio. Pesqui. Florest. Bras. 2017, 37, 131–138. [Google Scholar] [CrossRef]
- Bitencourt, F.; Zocche, J.J.; Costa, S.; Souza, P.Z.; Mendes, A. Nucleação por Mimosa bimucronata (DC.) O. Kuntze em áreas degradadas pela mineração de carvão. Rev. Bras. Biociênc. 2017, 5, 750–752. [Google Scholar]




| Identification and Taxonomy | FS% | SS | |||
|---|---|---|---|---|---|
| NI | Scientific Name | Family | A | B | |
| 1 | Alchornea glandulosa Poepp. & Endl. | Euphorbiaceae | 10 | 7 | Pi, Es |
| 2 | Citharexylum myrianthum Cham. | Verbenaceae | 10 | 10 | Pi, Es |
| 3 | Hieronyma alchorneoides Allemão | Phyllanthaceae | 10 | 7 | Es |
| 4 | Inga edulis Mart. | Fabaceae | 10 | 17 | Pi, Es |
| 5 | Inga laurina (Sw.) Willd. | Fabaceae | 10 | 7 | Es |
| 6 | Inga marginata Willd. | Fabaceae | 10 | 10 | Pi, Es |
| 7 | Mimosa bimucronata (DC.) Kuntze | Mimosaceae | 10 | 12 | Pi |
| 8 | Myrsine coriacea (Sw.) R. Br. Ex Roem. & Schult. | Primulaceae | 10 | 7 | Pi |
| 9 | Schizolobium parahyba (Vell.) Blake | Fabaceae | 10 | 7 | Pi, Es |
| 10 | Senna multijuga (Rich.) H. S. Irwin & Barneby | Fabaceae | 10 | 17 | Pi |
| VC | Variables | Database Collection | ||
|---|---|---|---|---|
| Quantitative data | ||||
| VC1 | Dimensions | Mean diameter (MD) | Diameter at breast-height (DBH 1.30 cm) | |
| Commercial height (CH) | Of possible use (m) | |||
| Total height (TH) | From the base to the end of the crown (m) | |||
| Semi-quantitative data | ||||
| VC2 | Stratification | Sociological position (SP) | 1: canopy | 3: understory |
| 2: intermediate | ||||
| Canopy luminosity (CL) | 1: illuminated | 4: 75% shaded | ||
| 2: 25% shaded | 5: 100% shaded | |||
| 3: 50% shaded | ||||
| VC3 | Qualities | Stem quality (SQ) | 1: straight stem-tree | 4: forked stem-tree > DAP |
| 2: 50% tortuous stem-tree | 5: forked stem-tree < DAP | |||
| 3: >50% tortuous stem-tree | 6: prop root | |||
| Stem health (SH) | 1: sane | 4: dead | ||
| 2: deteriorated | 5: stump sprout | |||
| 3: hollow tree | ||||
| Crown quality (CQ) | 1: 100% existent | 4: 25%–50% lifeless crown | ||
| 2: species deciduous | 5: >50% lifeless crown | |||
| 3: 25% lifeless crown | 6: 100% nonexistent | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schneider, C.R.; Mastella, A.D.F.; Mattar, E.A.; Angelo, A.C.; Behling, A.; Rios, R.C.; Blum, C.T.; Carpanezzi, A.A.; Neves, E.J.M.; Souza, K.K.F.d.; et al. Different Species Proportions Influence Silvicultural Heterogeneity of Trees in a Restoration of a Ombrophilous Dense Forest in Lowlands. Forests 2024, 15, 447. https://doi.org/10.3390/f15030447
Schneider CR, Mastella ADF, Mattar EA, Angelo AC, Behling A, Rios RC, Blum CT, Carpanezzi AA, Neves EJM, Souza KKFd, et al. Different Species Proportions Influence Silvicultural Heterogeneity of Trees in a Restoration of a Ombrophilous Dense Forest in Lowlands. Forests. 2024; 15(3):447. https://doi.org/10.3390/f15030447
Chicago/Turabian StyleSchneider, Chaiane Rodrigues, Alexandre Dal Forno Mastella, Eduardo Abilhoa Mattar, Alessandro Camargo Angelo, Alexandre Behling, Román Carlos Rios, Christopher Thomas Blum, Antonio Aparecido Carpanezzi, Edinelson Jose Maciel Neves, Karen Koch Fernandes de Souza, and et al. 2024. "Different Species Proportions Influence Silvicultural Heterogeneity of Trees in a Restoration of a Ombrophilous Dense Forest in Lowlands" Forests 15, no. 3: 447. https://doi.org/10.3390/f15030447