


Disturbance-Dependent Variation in Functional Redundancy Drives the Species Versus Functional Diversity Relationship across Spatial Scales and Vegetation Layers


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
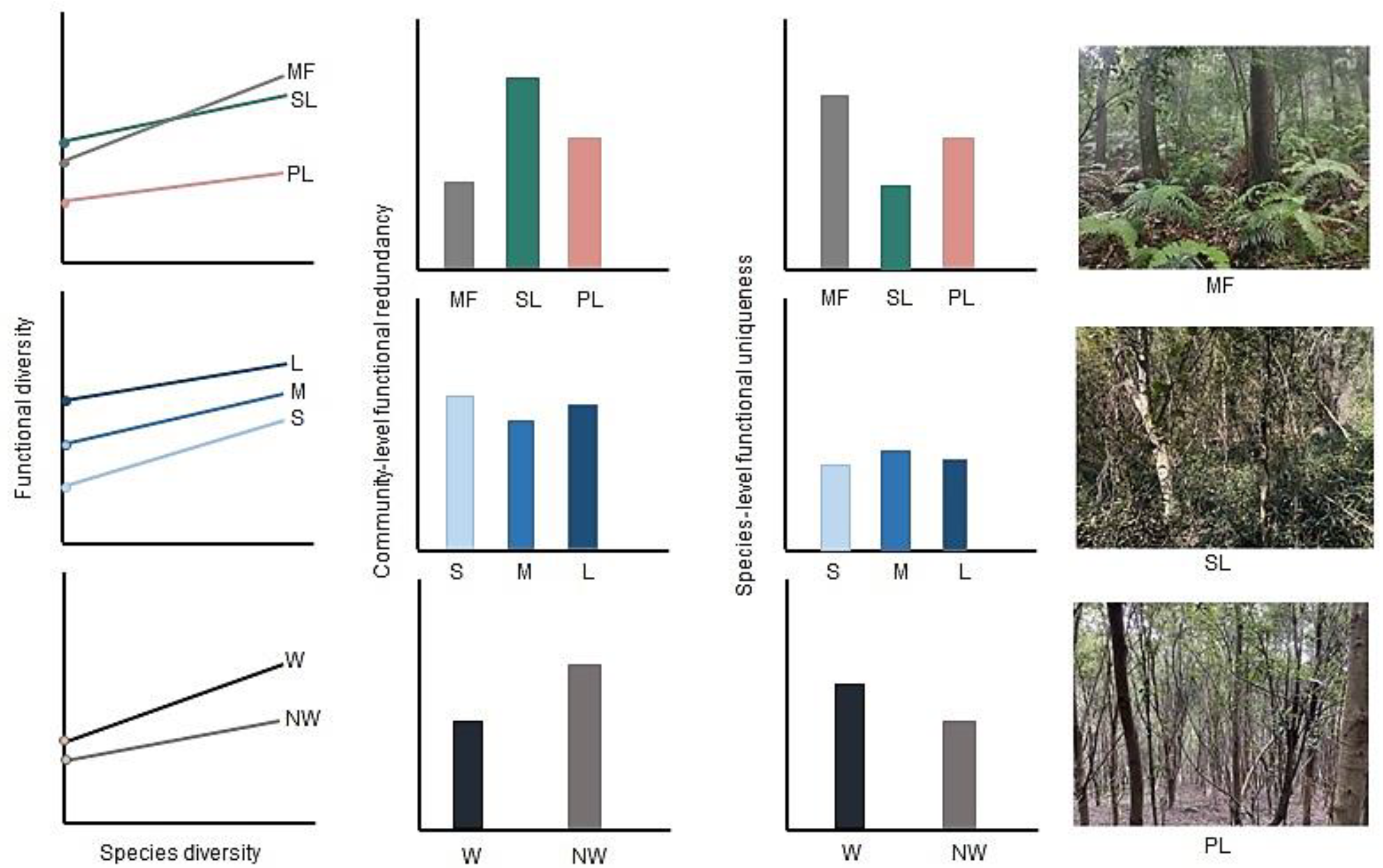
- Do SD–FD relationships (i.e., relationships between species richness and functional richness, species evenness and functional evenness, and Shannon’s species diversity and Rao’s functional diversity) vary among intact mature forests versus disturbed shrublands and plantations?
- Do conclusions to question 1 vary among spatial scales of observation (i.e., small, medium versus large plots) and vegetation layers (i.e., woody overstory versus non-woody understory vegetation)?
- Can community-level functional redundancy and species-level functional uniqueness explain any observed variation in the relationship between different indices of species versus functional diversity?
2. Materials and Methods
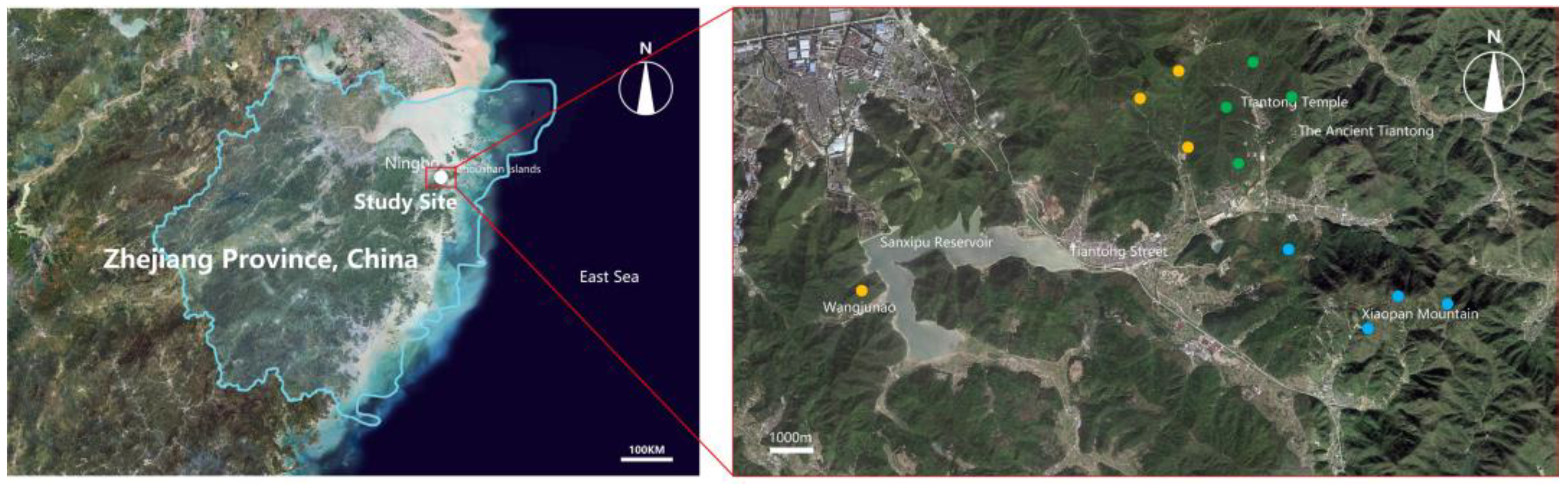
2.1. Study Site and Disturbance History
2.2. Plot Establishment, Spatial Scaling, and Vegetation Sampling
2.3. Functional Traits
2.4. Quantifying Species and Functional Diversity and Composition
2.5. Quantifying Community-Level Functional Redundancy and Species-Level Functional Uniqueness
2.6. Statistical Analysis
3. Results
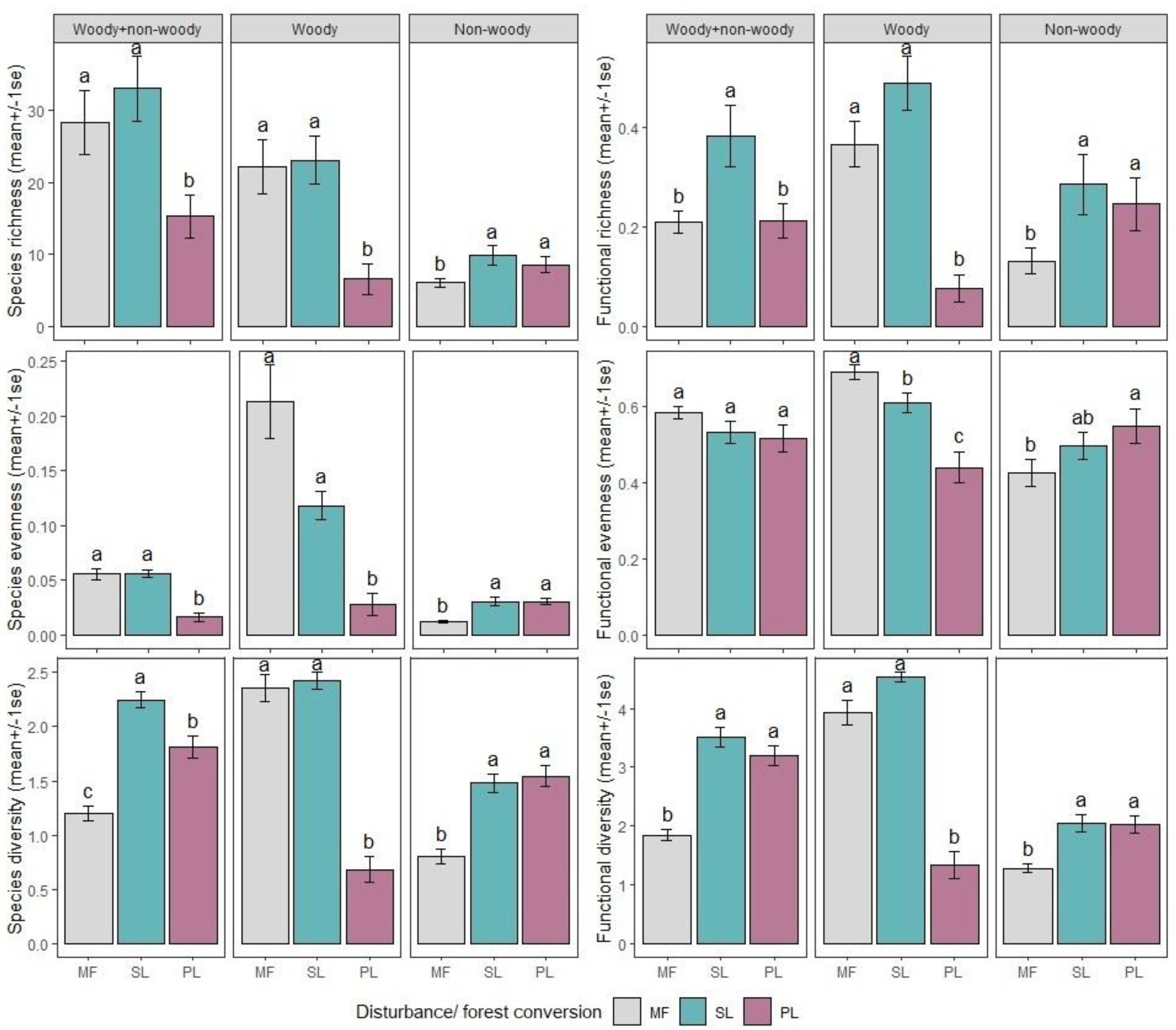
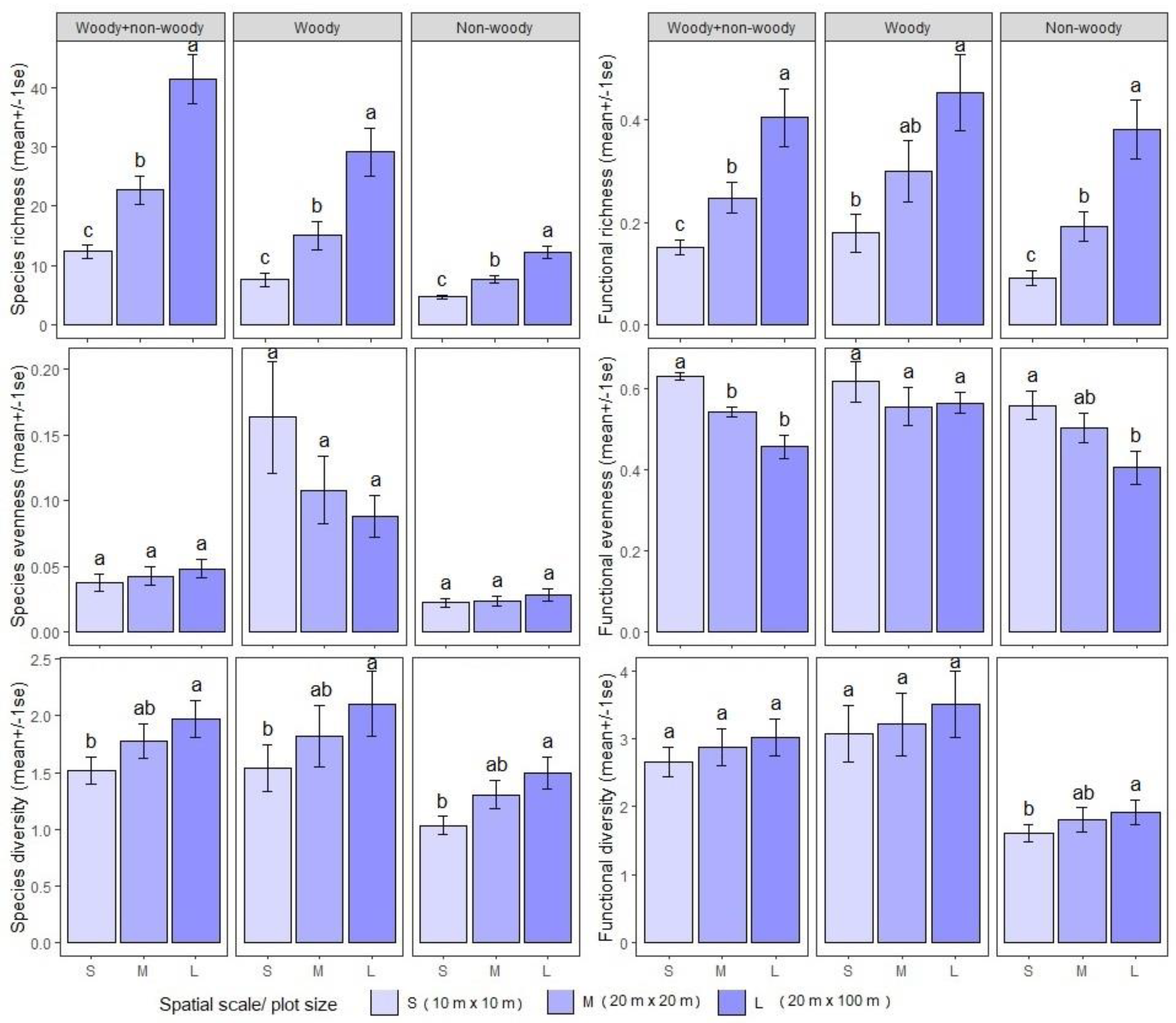
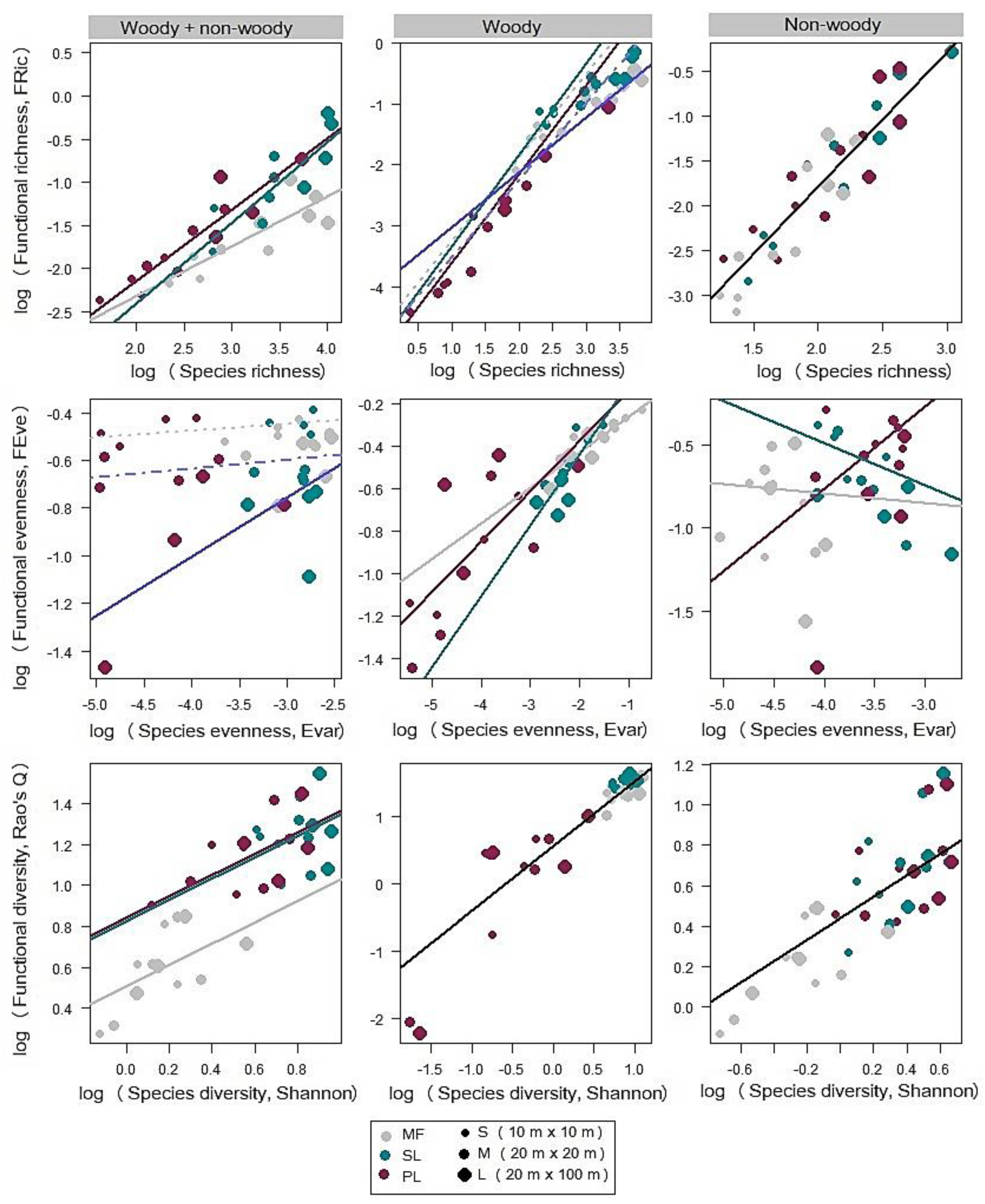
3.1. Species Richness Versus Functional Richness (FRic) Relationship
3.2. Species Evenness Versus Functional Evenness (FEve) Relationship
3.3. Species Diversity Versus Functional Diversity (RaoQ) Relationship
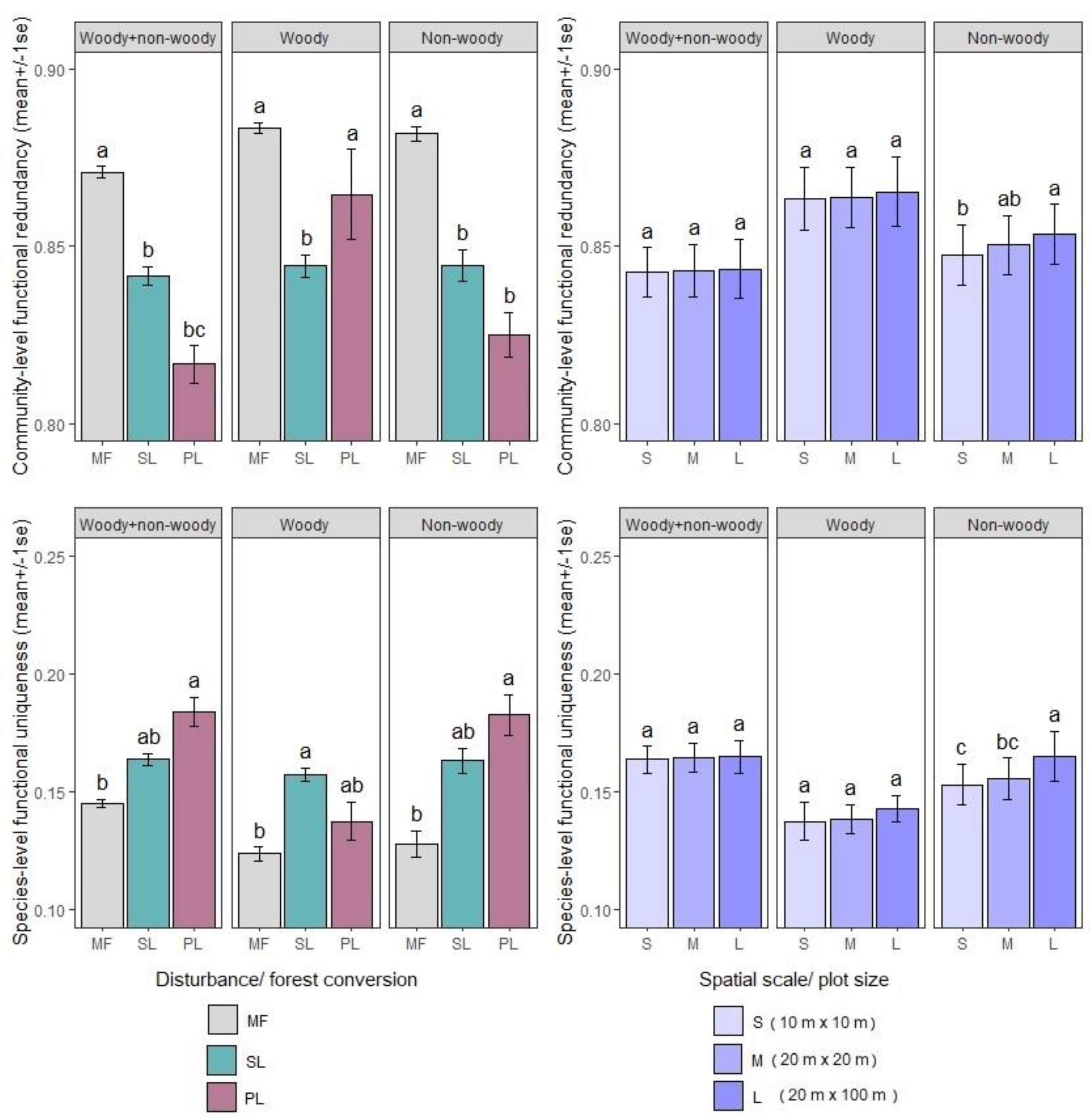
3.4. Community-Level Functional Redundancy and Species-Level Functional Uniqueness
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pereira, H.M.; Leadley, P.W.; Proença, V.; Alkemade, R.; Scharlemann, J.P.W.; Fernandez-Manjarrés, J.F.; Araújo, M.B.; Balvanera, P.; Biggs, R.; Cheung, W.W.L.; et al. Scenarios for Global Biodiversity in the 21st Century. Science 2010, 330, 1496–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadotte, M.W.; Carscadden, K.; Mirotchnick, N. Beyond species: Functional diversity and the maintenance of ecological processes and services. J. Appl. Ecol. 2011, 48, 1079–1087. [Google Scholar]
- Hector, A.; Bagchi, R. Biodiversity and ecosystem multifunctionality. Nature 2007, 448, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Wedin, D.; Knops, J. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature 1996, 379, 718–720. [Google Scholar] [CrossRef]
- Biswas, S.R.; Mallik, A.U.; Braithwaite, N.T.; Biswas, P.L. Effects of disturbance type and microhabitat on species and functional diversity relationship in stream-bank plant communities. For. Ecol. Manag. 2019, 432, 812–822. [Google Scholar] [CrossRef]
- Mayfield, M.M.; Boni, M.F.; Daily, G.C.; Ackerly, D. Species and functional diversity of native and human-dominated plant communities. Ecology 2005, 86, 2365–2372. [Google Scholar] [CrossRef] [Green Version]
- Mayfield, M.M.; Bonser, S.P.; Morgan, J.W.; Aubin, I.; McNamara, S.; Vesk, P.A. What does species richness tell us about functional trait diversity? Predictions and evidence for responses of species and functional trait diversity to land-use change. Glob. Ecol. Biogeogr. 2010, 19, 423–431. [Google Scholar] [CrossRef]
- Sasaki, T.; Okubo, S.; Okayasu, T.; Jamsran, U.; Ohkuro, T.; Takeuchi, K. 1-208. Two-phase functional redundancy in plant communities along a grazing gradient in Mongolian rangelands. Ecology 2009, 90, 2598–2608. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.R.; Mallik, A.U. Species diversity and functional diversity relationship varies with disturbance intensity. Ecosphere 2011, 2, art52. [Google Scholar] [CrossRef]
- Carmona, C.P.; Azcárate, F.M.; de Bello, F.; Ollero, H.S.; Lepš, J.; Peco, B. Taxonomical and functional diversity turnover in Mediterranean grasslands: Interactions between grazing, habitat type and rainfall. J. Appl. Ecol. 2012, 49, 1084–1093. [Google Scholar] [CrossRef]
- Dovrat, G.; Meron, E.; Shachak, M.; Moshe, Y.; Osem, Y. The relationship between species diversity and functional diversity along aridity gradients in semi-arid rangeland. J. Arid Environ. 2021, 195, 104632. [Google Scholar] [CrossRef]
- Ouyang, J.; Biswas, S.R.; Yin, C.; Qing, Y.; Biswas, P.L. Shifting Importance of Abiotic versus Biotic Filtering from Intact Mature Forests to Post-Clearcut Secondary Forests. Forests 2022, 13, 672. [Google Scholar] [CrossRef]
- Chazdon, R.L. Second Growth: The Promise of Tropical Forest Regeneration in an Age of Deforestation; University of Chicago Press: Chicago, IL, USA, 2014. [Google Scholar]
- Ricotta, C.; de Bello, F.; Moretti, M.; Caccianiga, M.; Cerabolini, B.E.L.; Pavoine, S. Measuring the functional redundancy of biological communities: A quantitative guide. Methods Ecol. Evol. 2016, 7, 1386–1395. [Google Scholar] [CrossRef] [Green Version]
- Li, H. Scale-Dependent Effects of Forest Conversion on Species and Functional Diversity in Zhejiang Tiantong National Forest Park; East China Normal University: Shanghai, China, 2021. [Google Scholar]
- Wiens, J.A. Spatial Scaling in Ecology. Funct. Ecol. 1989, 3, 385–397. [Google Scholar] [CrossRef]
- Levin, S.A. The Problem of Pattern and Scale in Ecology: The Robert H. MacArthur Award Lecture. Ecology 1992, 73, 1943–1967. [Google Scholar] [CrossRef]
- Xiang, J. Effects of Forest Conversion on Spatial Aotocorrelation in Plant Species and Functional Diversity in Tiantong National Forest Park; East China Normal University: Shanghai, China, 2021. [Google Scholar]
- Fletcher, R.; Fortin, M.-J. Scale. In Spatial Ecology and Conservation Modeling: Applications with R; Springer International Publishing: Cham, Switzerland, 2018; pp. 17–53. [Google Scholar] [CrossRef]
- Albert, C.H.; Grassein, F.; Schurr, F.M.; Vieilledent, G.; Violle, C. When and how should intraspecific variability be considered in trait-based plant ecology? Perspect. Plant Ecol. Evol. Syst. 2011, 13, 217–225. [Google Scholar] [CrossRef]
- McGuinness, K.A. Species–area curves. Biol. Rev. 1984, 59, 423–440. [Google Scholar] [CrossRef]
- Chen, H.Y.H.; Biswas, S.R.; Sobey, T.M.; Brassard, B.W.; Bartels, S.F. Reclamation strategies for mined forest soils and overstorey drive understorey vegetation. J. Appl. Ecol. 2018, 55, 926–936. [Google Scholar] [CrossRef]
- Stirling, G.; Wilsey, B. Empirical Relationships between Species Richness, Evenness, and Proportional Diversity. Am. Nat. 2001, 158, 286–299. [Google Scholar] [CrossRef]
- Villeger, S.; Mason, N.W.H.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.-Y.; Guo, C.-Z.; Ni, J. Dynamics of major forest vegetations in Tiantong National Forest Park during the last 30 years. Chin. J. Appl. Ecol. 2014, 25, 1547–1554. (In Chinese) [Google Scholar]
- Zhang, S.; Zeng, Y.; He, Y.; Chen, Y.; Lu, J.; Liu, H.; Xu, M.; Wang, X. Spatiotemporal distribution characteristics of leaf-litter nutrients in a dynamic plot of subtropical evergreen broad-leaved forest in Tiantong, Zhejiang Province. Acta Ecol. Sin. 2020, 40, 7335–7342. [Google Scholar]
- Wang, X.-H.; Kent, M.; Fang, X.-F. Evergreen broad-leaved forest in Eastern China: Its ecology and conservation and the importance of resprouting in forest restoration. For. Ecol. Manag. 2007, 245, 76–87. [Google Scholar] [CrossRef]
- Yan, E.-R.; Wang, X.-H.; Huang, J.-J. Shifts in plant nutrient use strategies under secondary forest succession. Plant Soil 2006, 289, 187–197. [Google Scholar] [CrossRef]
- Yan, E.-R.; Wang, X.-H.; Guo, M.; Zhong, Q.; Zhou, W.; Li, Y.-F. Temporal patterns of net soil N mineralization and nitrification through secondary succession in the subtropical forests of eastern China. Plant Soil 2009, 320, 181–194. [Google Scholar] [CrossRef]
- Fridley, J.D.; Stachowicz, J.J.; Naeem, S.; Sax, D.F.; Seabloom, E.W.; Smith, M.D.; Stohlgren, T.J.; Tilman, D.; Holle, B.V. The invasion paradox: Reconciling pattern and process in species invasions. Ecology 2007, 88, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Adler, P.B.; Salguero-Gomez, R.; Compagnoni, A.; Hsu, J.S.; Ray-Mukherjee, J.; Mbeau-Ache, C.; Franco, M. Functional traits explain variation in plant life history strategies. Proc. Natl. Acad. Sci. USA 2014, 111, 740–745. [Google Scholar] [CrossRef] [Green Version]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. From tropics to tundra: Global convergence in plant functioning. Proc. Natl. Acad. Sci. USA 1997, 94, 13730–13734. [Google Scholar] [CrossRef] [Green Version]
- Garnier, E.; Cortez, J.; Billes, G.; Navas, M.L.; Roumet, C.; Debussche, M.; Laurent, G.; Blanchard, A.; Aubry, D.; Bellmann, A.; et al. Plant functional markers capture ecosystem properties during secondary succession. Ecology 2004, 85, 2630–2637. [Google Scholar] [CrossRef]
- Gratani, L.; Varone, L.; Crescente, M.F.; Catoni, R.; Ricotta, C.; Puglielli, G. Leaf thickness and density drive the responsiveness of photosynthesis to air temperature in Mediterranean species according to their leaf habitus. J. Arid Environ. 2018, 150, 9–14. [Google Scholar] [CrossRef]
- Pauli, D.; White, J.W.; Andrade-Sanchez, P.; Conley, M.M.; Heun, J.; Thorp, K.R.; French, A.N.; Hunsaker, D.J.; Carmo-Silva, E.; Wang, G.; et al. Investigation of the Influence of Leaf Thickness on Canopy Reflectance and Physiological Traits in Upland and Pima Cotton Populations. Front. Plant Sci. 2017, 8, 1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eggli, U.; Nyffeler, R.; Eggli, U.; Nyffeler, R. Living under temporarily arid conditions-succulence as an adaptive strategy. Bradleya 2009, 27, 13–36. [Google Scholar] [CrossRef]
- Hunt, E.R.; Weber, J.A.; Gates, D.M. Effects of Nitrate Application on Amaranthus powellii Wats. III. Optimal Allocation of Leaf Nitrogen for Photosynthesis and Stomatal Conductance. Plant Physiol. 1985, 79, 619–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Diaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; ter Steege, H.; Morgan, H.D.; van der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, A. A method to improve leaf succulence quantification. Braz. Arch. Biol. Technol. 1999, 42, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Licon, C. Proximate and other chemical analyses. In Encyclopedia of Dairy Sciences, 3rd ed.; McSweeney, P.L.H., McNamara, J.P., Eds.; Academic Press: Oxford, UK, 2022; pp. 521–529. [Google Scholar] [CrossRef]
- Guo, H.-N.; Wang, L.-X.; Liu, H.-T. Potential mechanisms involving the immobilization of Cd, As and Cr during swine manure composting. Sci. Rep. 2020, 10, 16632. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L. A Rapid and Accurate Procedure for Estimation of Organic Carbon in Soils; Indiana Academy of Science: Indianapolis, IN, USA, 1974; pp. 456–462. [Google Scholar]
- Smith, B.; Wilson, J.B. A consumer’s guide to evenness indices. Oikos 1996, 76, 70–82. [Google Scholar] [CrossRef]
- Tuomisto, H. An updated consumer’s guide to evenness and related indices. Oikos 2012, 121, 1203–1218. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Albert, C.H.; de Bello, F.; Boulangeat, I.; Pellet, G.; Lavorel, S.; Thuiller, W. On the importance of intraspecific variability for the quantification of functional diversity. Oikos 2012, 121, 116–126. [Google Scholar] [CrossRef]
- Violle, C.; Enquist, B.J.; Mcgill, B.J.; Jiang, L.; Albert, C.H.; Hulshof, C.; Jung, V.; Messier, J. The return of the variance: Intraspecific variability in community ecology. Trends Ecol. Evol. 2012, 27, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Ross, S.R.P.-J.; Hassall, C.; Hoppitt, W.J.E.; Edwards, F.A.; Edwards, D.P.; Hamer, K.C. Incorporating intraspecific trait variation into functional diversity: Impacts of selective logging on birds in Borneo. Methods Ecol. Evol. 2017, 8, 1499–1505. [Google Scholar] [CrossRef] [Green Version]
- Laliberte, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Gower, J.C. Principal coordinates analysis. In Encyclopedia of Biostatistics; Armitage, P., Colton, T., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2005. [Google Scholar]
- de Bello, F.; Lepš, J.; Lavorel, S.; Moretti, M. Importance of species abundance for assessment of trait composition: An example based on pollinator communities. Community Ecol. 2007, 8, 163–170. [Google Scholar] [CrossRef]
- Warring, B.; Cardoso, F.C.G.; Marques, M.C.M.; Varassin, I.G. Functional diversity of reproductive traits increases across succession in the Atlantic forest. Rodriguésia 2016, 67, 321–333. [Google Scholar] [CrossRef] [Green Version]
- Camilo, G.d.S.; Terra, B.d.F.; Araujo, F.G. Using the relationship between taxonomic and functional diversity to assess functional redundancy in streams of an altered tropical watershed. Environ. Biol. Fishes 2018, 101, 1395–1405. [Google Scholar] [CrossRef]
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional richness, functional evenness and functional divergence: The primary components of functional diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Hillebrand, H.; Bennett, D.M.; Cadotte, M.W. Consequences of dominance: A review of evenness effects on local and regional ecosystem processes. Ecology 2008, 89, 1510–1520. [Google Scholar] [CrossRef]
- Bergholz, K.; May, F.; Giladi, I.; Ristow, M.; Ziv, Y.; Jeltsch, F. Environmental heterogeneity drives fine-scale species assembly and functional diversity of annual plants in a semi-arid environment. Perspect. Plant Ecol. Evol. Syst. 2017, 24, 138–146. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biswas, S.R.; Yin, C.; Gong, L.; Qing, Y.; Li, J. Disturbance-Dependent Variation in Functional Redundancy Drives the Species Versus Functional Diversity Relationship across Spatial Scales and Vegetation Layers. Forests 2023, 14, 408. https://doi.org/10.3390/f14020408
Biswas SR, Yin C, Gong L, Qing Y, Li J. Disturbance-Dependent Variation in Functional Redundancy Drives the Species Versus Functional Diversity Relationship across Spatial Scales and Vegetation Layers. Forests. 2023; 14(2):408. https://doi.org/10.3390/f14020408
Chicago/Turabian StyleBiswas, Shekhar R., Chaoqin Yin, Li Gong, Yanxia Qing, and Jialin Li. 2023. "Disturbance-Dependent Variation in Functional Redundancy Drives the Species Versus Functional Diversity Relationship across Spatial Scales and Vegetation Layers" Forests 14, no. 2: 408. https://doi.org/10.3390/f14020408