


Ecological Potential of Mediterranean Habitats for Black Pine (Pinus nigra J.F. Arnold) in Croatia

Abstract
:1. Introduction
2. Materials and Methods
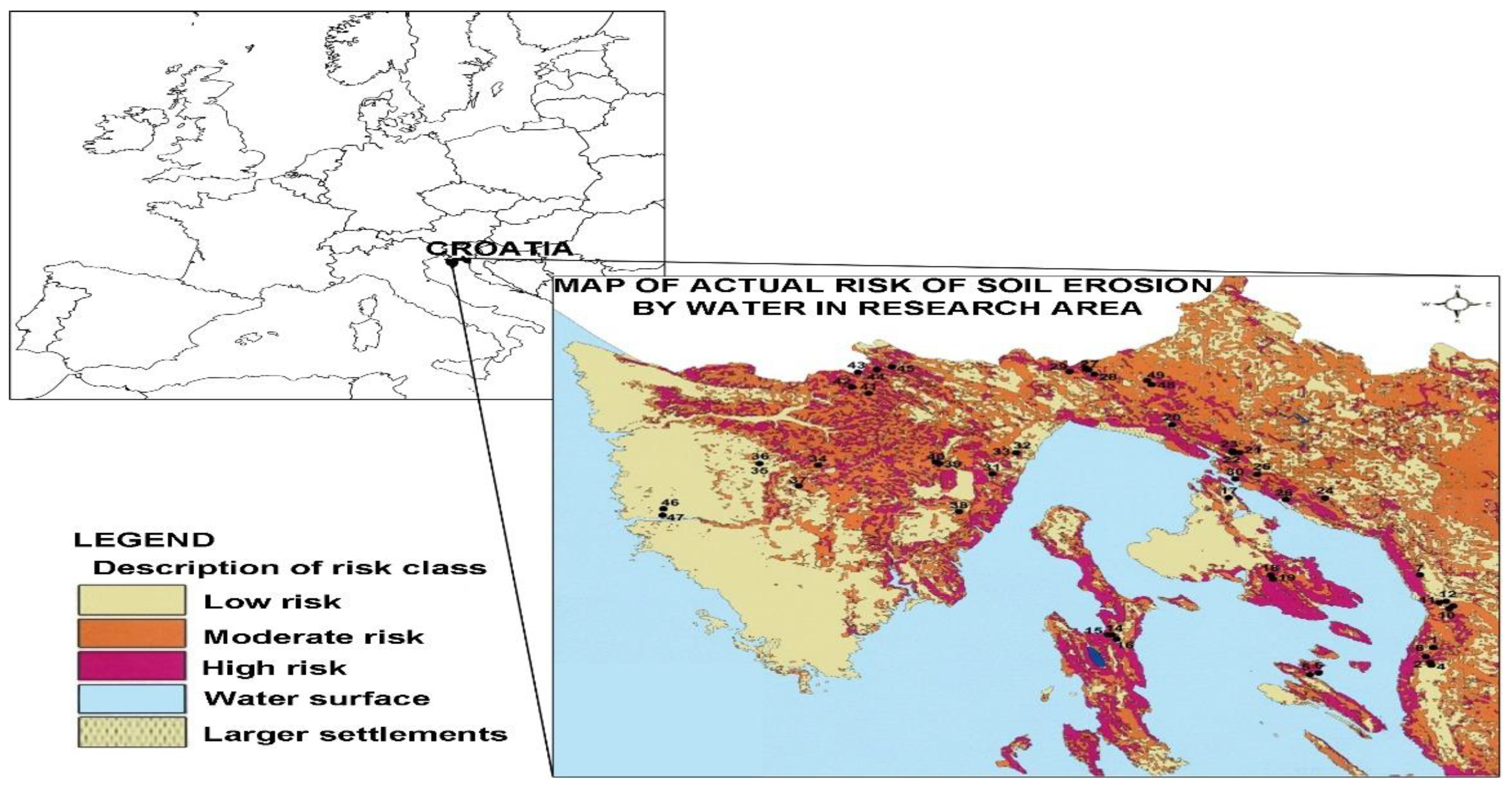
2.1. Study Area
2.2. Data Collection
2.3. Data Analyses
- Soil reaction was measured electrometrically with a combined electrode in a suspension of soil in water, i.e., in 0.01 M CaCl2 in the ratio of 1:2.5 for surface mineral and argyl-accumulative layers, and in the ratio of 1:10 for prominently humic surface horizons. A laboratory microprocessor pH meter (MA 5736, Metrel, accuracy ± 0.01 pH) was used for the measurements.
- Humus content was determined using the bichromatic method according to Tjurin.
- The total nitrogen content was determined by incineration according to the Kjeldajl procedure and distillation according to Bremner.
- Samples of the forest floor were ground and homogenised and incinerated in a microwave oven with HNO3 (Milestone Laboratory System). K content was determined by flame-photometry, and all other elements determined using the AAS method (atomic absorption spectrophotometry).
2.3.1. Statistical Analysis
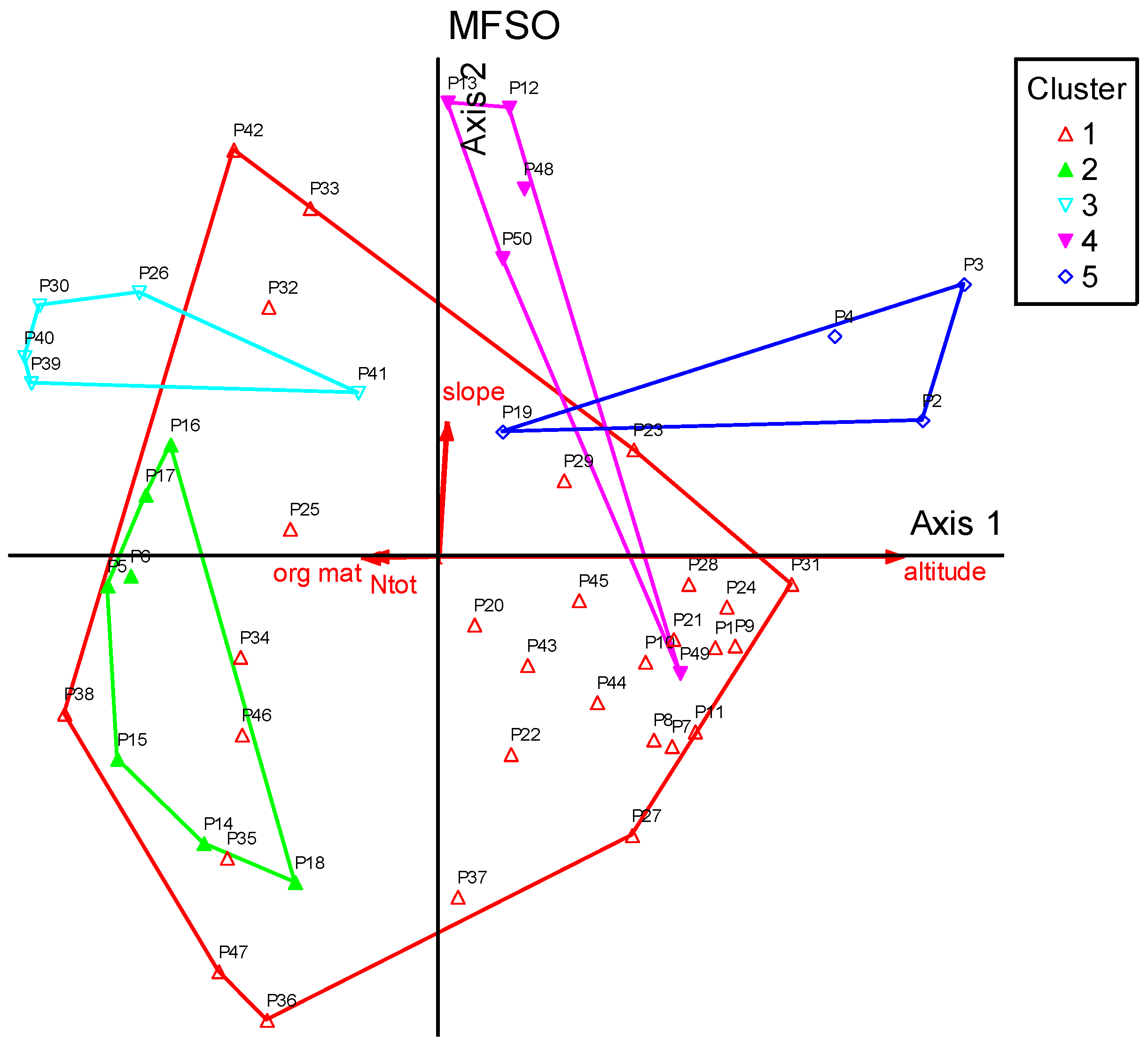
2.3.2. Multivariate Fuzzy Set Ordination
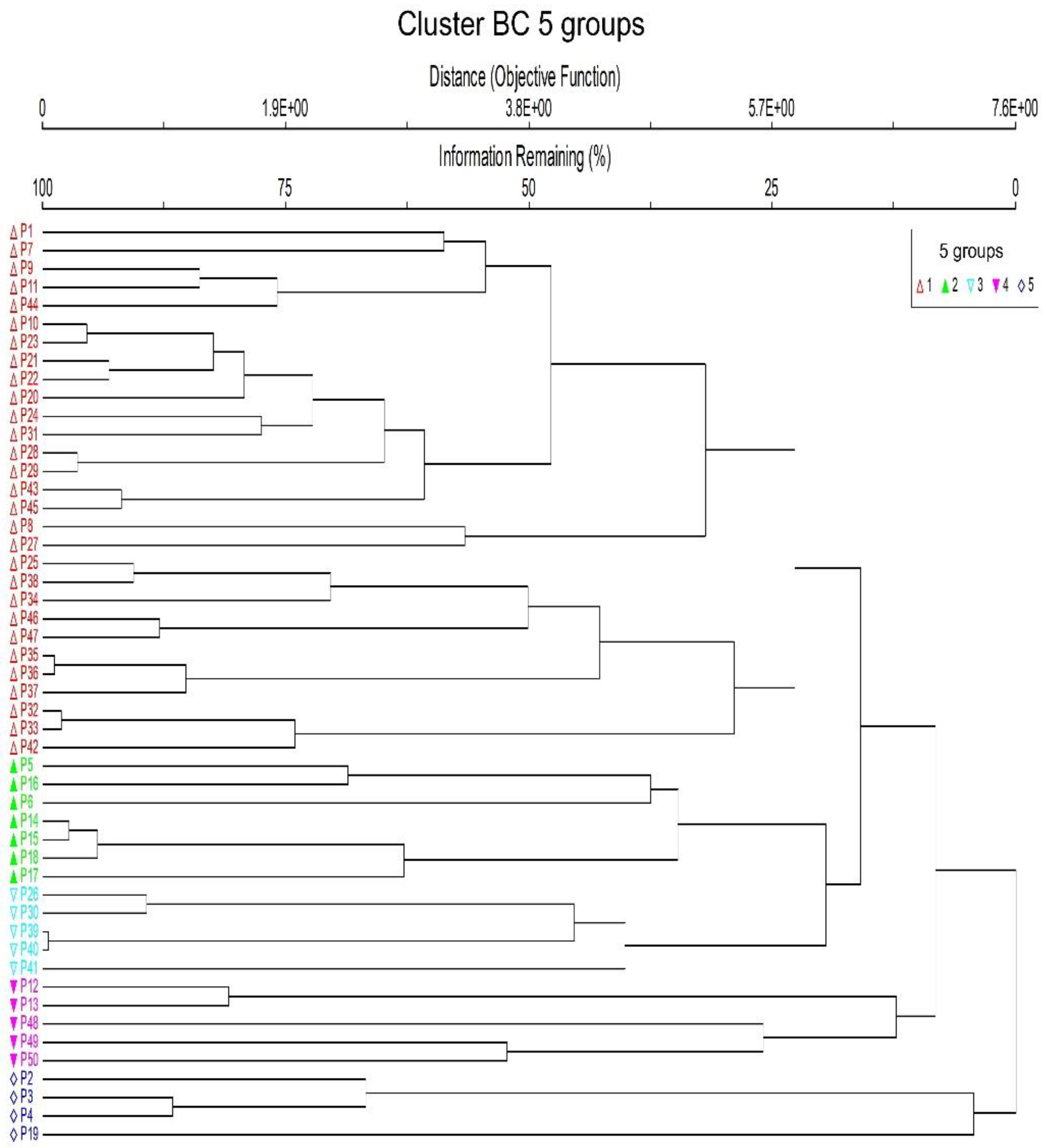
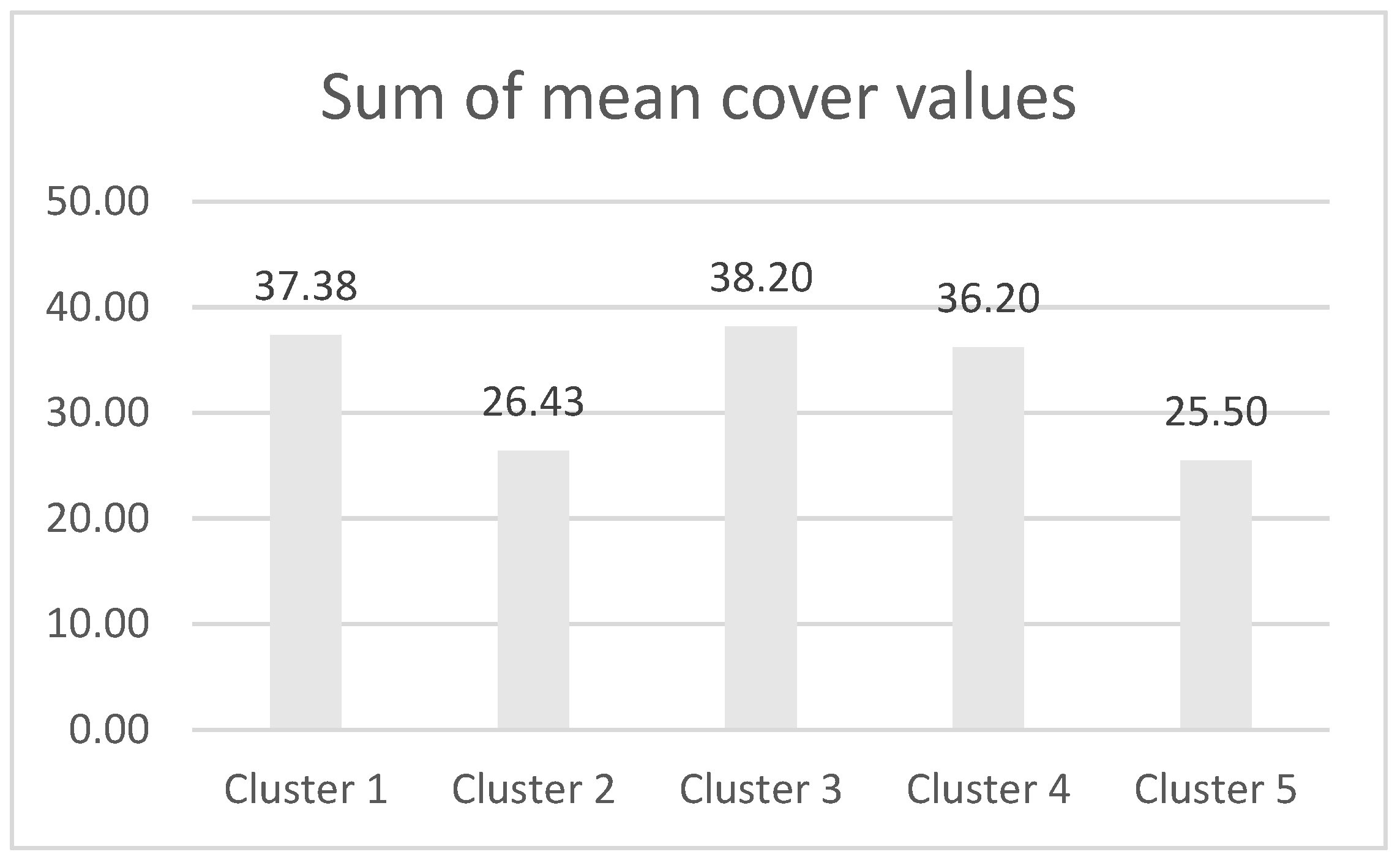
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quézel, P.; Médail, F. Ecologie et Biogéographie du Bassin Méditerranéen; Elsevier: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Barbéro, M.; Oisel, R.; Quézel, P.; Richardson, D.; Romane, F. Pines of the Mediterranean Basin. In Ecology and Biogeography of Pinus; Richardson, D.M., Ed.; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Lucas-Borja, M.E.; Jing, X.; Candel-Perez, D.; Parhizkar, M.; Rocha, F.; Heydari, M.; Muñoz-Rojas, M.; Zema, D.A. Afforestation with Pinus nigra Arn ssp salzmannii along an elevation gradient: Controlling factors and implications for climate change adaptation. Trees 2022, 36, 93–102. [Google Scholar] [CrossRef]
- Ivančević, V. Biološko-tehnički radovi na sanaciji Senjske bujice «torrente» i povećanje vodnog kapaciteta. Šum. List. 2005, 129, 91–109. (In Croatian) [Google Scholar]
- Piermattei, A.; Renzaglia, F.; Urbinati, C. Recent expansion of Pinus nigra Arn. above the timberline in the central Apennines, Italy. Ann. For. Sci. 2012, 69, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Piermattei, A.; Garbarino, M.; Urbinati, C. Structural attributes, tree-ring growth and climate sensitivity of Pinus nigra Arn. At high altitude: Common patterns of a possible treeline shift in the central Apennines (Italy). Dendrochronologia 2014, 32, 210–219. [Google Scholar]
- Piermattei, A.; Lingua, E.; Urbinati, C.; Garbarino, M. Pinus nigra anthropogenic treelines in the central Apennines show common pattern of tree recruitment. Eur. J. For. Res. 2016, 135, 1119–1130. [Google Scholar] [CrossRef] [Green Version]
- Santana, V.M.; Alday, J.G.; Baeza, M. Effects of fire regime shift in Mediterranean Basin ecosystems: Changes in soil seed bank composition among functional types. Plant Ecol. 2014, 215, 555–566. [Google Scholar] [CrossRef]
- Santana, V.M.; González-Pelayo, J.G.; Maia, P.E.E.; Varela, T.M.E.; Valdecantos, A.; Vallejo, V.R.; Keizeri, J.J. Effects of fire recurrence and different salvage logging techniques on carbon storage in Pinus pinaster forests from northern Portugal. Eur. J. For. Res. 2016, 135, 1107–1117. [Google Scholar] [CrossRef]
- Muñoz-Rojas, M.; De la Rosa, D.; Zavala, L.; Jordán, A.; Anaya-Romero, M. Changes in land cover and vegetation carbon stocks in Andalusia, Southern Spain (1956–2007). Sci. Total Environ. 2011, 409, 2796–2806. [Google Scholar] [CrossRef]
- Morales-Molino, C.; Tinner, W.; García-Antón, M.; Colombaroli, D. The historical demise of Pinus nigra forests in the Northern Iberian Plateau (south-western Europe). J. Ecol. 2016, 105, 634–646. [Google Scholar] [CrossRef] [Green Version]
- Sheffer, E. A review of the development of Mediterranean pine-oak ecosystems after land abandonment and afforestation: Are they novel ecosystems? Ann. For. Sci. 2012, 69, 429–443. [Google Scholar] [CrossRef] [Green Version]
- Espelta, J.M. La reconstruccio de paisatges forestals afectats per grans incendis. Projecte pilot a l’incendi del Bages-Bergueda del 1994. Silvicultura 1999, 27, 6–9. [Google Scholar]
- Espelta, J.M.; Retana, J.; Habrouk, A. An economic and ecological multi-criteria evaluation of reforestation methods to recover burned Pinus nigra forests in NE Spain. For. Ecol. Manag. 2003, 180, 185–198. [Google Scholar] [CrossRef]
- Vallejo, V.R.; Bautista, S.; Cortina, J. Restoration for soil protection after disturbances. In Life and Environment in the Mediterranean; Trabaud, L., Ed.; WIT Press: Boston, MA, USA, 2000; pp. 301–343. [Google Scholar]
- Allen, H.D. Response of past and present Mediterranean ecosystems to environmental change. Prog. Phys. Geogr. 2003, 27, 359–377. [Google Scholar] [CrossRef]
- González-Hidalgo, J.; De Luis, M.; Raventós, J.; Sánchez, J. Spatial distribution of seasonal rainfall trends in a western Mediterranean area. Int. J. Climatol. 2001, 21, 843–860. [Google Scholar] [CrossRef]
- Martinović, J. Gospodarenje Šumskim Tlima u Hrvatskoj; Šumarski Institut Jastrebarsko i Hrvatske Šume d.o.o.: Zagreb, Croatia, 2003; p. 521. (In Croatian) [Google Scholar]
- Trinajstić, I. Nomenklaturno-sintaksonomska revizija submediteranskih šuma crnoga bora (Pinus nigra Arnold) u Hrvatskoj. Šum. List. 1998, 122, 147–154. (In Croatian) [Google Scholar]
- Rauš, Đ.; Trinajstić, I.; Vukelić, J.; Medvedović, J. Biljni svijet hrvatskih šuma. In Šume u Hrvatskoj.; Rauš, Đ., Ed.; Šumarski Fakultet Sveučilišta u Zagrebu i Hrvatske Šume: Zagreb, Croatia, 1992; pp. 33–77. (In Croatian) [Google Scholar]
- Liber, Z. Filogenetski i Taksonomski Odnosi Populacija Crnoga Bora (Pinus nigra Arnold) u Hrvatskoj. Ph.D. Thesis, Prirodoslovno-Matematički Fakultet Sveučilišta u Zagrebu (Faculty of Science University of Zagreb), Zagreb, Croatia, 2000. [Google Scholar]
- Gaussen, H.; Heywood, V.H.; Chater, A.O.; Pinus, L. Flora Europaea, 2nd ed.; Tutin, I.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1993; p. 42. [Google Scholar]
- Vidaković, M. Oblici crnog bora u Jugoslaviji na temelju antomije iglica. Glas. Šum. Pokuse 1957, 13, 111–148. (In Croatian) [Google Scholar]
- Yildiz, O.; Altundağ, E.; Çetin, B.; Güner, S.T.; Sarginci, M.; Toprak, B. Experimental arid land afforestation in Central Anatolia, Turkey. Environ. Monit. Assess. 2018, 190, 355. [Google Scholar] [CrossRef]
- Bayar, E.; Deligöz, A. Ecophysiological behavior of Mediterranean woody species under summer drought. Bosque 2021, 42, 311–321. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.M.; Rodriguez-Vallejo, C.; Silveiro, E.; Hortal, A.; Palacios-Rodríguez, G.; Duque-Lazo, J.; Camarero, J.J. Cumulative Drought Stress Leads to a Loss of Growth Resilience and Explains Higher Mortality in Planted than in Naturally Regenerated Pinus pinaster Stands. Forests 2018, 9, 358. [Google Scholar] [CrossRef] [Green Version]
- Zaghi, D. Management of Natura 2000 Habitats. 9530 *(Sub)-Mediterranean Pine Forests with Endemic Black Pines; Technical Report 2008 24/24; European Commission: Brussels, Belgium, 2008. [Google Scholar]
- Bezak, K. Tablice drvnih masa cera, crnog bora i običnog bora. Radovi 1992, 5, 47–65. (In Croatian) [Google Scholar]
- Braun-Blanquet, J. Grundzüge der Vegetationskunde. In Pflanzensoziologie, 3rd ed.; Springer: Wien, Austria, 1964. [Google Scholar]
- Dierschke, H. Grundlagen und Methoden. In Pflanzensoziologie; Verlag Eugen Ulmer: Stuttgart, Germany, 1994. [Google Scholar]
- Barkman, J.J.; Doing, H.; Segal, S. Kritische Bemerkungen und Vorschläge zur Quantitativen Vegetationsanalysen. Acta Bot. Neerl. 1964, 13, 394–419. [Google Scholar] [CrossRef]
- van der Maarel, E. Transformation of cover-abundance values in phytosociology and its effect on community similarity. Vegetatio 1979, 39, 97–114. [Google Scholar]
- Nikolić, T. (Ed.) Popis flore Hrvatske, 1. dio. Flora Croatica. Index Florae Croaticae. Pars 1 Nat. Croat. 1994, 3 (Suppl. S2), 1–116. [Google Scholar]
- Nikolić, T. (Ed.) Popis flore Hrvatske, 2. dio. Flora Croatica. Index Florae Croaticae. Pars 2 Nat. Croat. 1997, 6 (Suppl. S1), 1–232. [Google Scholar]
- Nikolić, T. (Ed.) Popis flore Hrvatske, 3. dio. Flora Croatica. Index Florae Croaticae. Pars 3 Nat. Croat. 2000, 9 (Suppl. S1), 1–324. [Google Scholar]
- United Nations Economic Commission for Europe. Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; Part XI, Sampling and Analysis of Litterfall. 2004, pp. 1–18. Available online: https://www.icp-forests.org/pdf/manual/2000/Chapt11_compl2004.pdf (accessed on 17 September 2022).
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Dufrene, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 354–366. [Google Scholar] [CrossRef]
- Roberts, D.W. Statistical analysis of multidimensional fuzzy set ordinations. Ecology 2008, 89, 1246–1260. [Google Scholar] [CrossRef]
- Roberts, D.W. Comparison of multidimensional fuzzy set ordination with CCA and DB-RDA. Ecology 2009, 90, 2622–2634. [Google Scholar] [CrossRef]
- Ellenberg, H. Wege der Geobotanik zum Verständnis der Pflanzendecke. Naturwissenschaften 1968, 55, 462–470. [Google Scholar] [CrossRef]
- Tomašević, A. Meliorativni utjecaj kulture alepskog bora (Pinus halepensis Mill.) i pinije (Pinus pinea Endl.) na degradirano stanište hrasta medunca (Querco pubescentis-Carpinetum orientalis H-ić, Anić 1959) u zadarskom području. Glas. Šum. Pokuse 1994, 30, 223–298. (In Croatian) [Google Scholar]
- Enríquez-De-Salamanca, Á. Effects of Climate Change on Forest Regeneration in Central Spain. Atmosphere 2022, 13, 1143. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Trew, B.T.; Maclean, I.M.D. Vulnerability of global biodiversity hotspots to climate change. Glob. Ecol. Biogeogr. 2021, 30, 768–783. [Google Scholar] [CrossRef]
- Gil, L.; Prada Aránzazu, M. Los pinos como especies basicas de la restauracion forestal en el medio mediterráneo. Ecologia 1993, 7, 113–125. [Google Scholar]
- Serrada Hierro, R. Consideraciones sobre el impacto de la repoblacion forestal en el suelo. Ecologia 1990, 1, 453–462. [Google Scholar]
- Grau Corbi, J.M.; Cámara Obregón, A.; Montoto Quinteiro, J.L. Fitoclimatología básica de los Pinus nigra Arn., Pinus sylvestris L. Y Pinus pinaster Ait. Aplicación del modelo de idoneidad. Investig. Agrar. Sist. Recur. For. 1999, 1, 37–51. [Google Scholar]
- Quézel, P. Les forêts du pourtour méditerranéen. In Forêts et Maquis Méditerranéens: Écologie, Conservation et Aménagements; Note technique MAB; UNESCO: France, Paris, 1976; Volume 2, pp. 9–33e. [Google Scholar]
- M’hirit, O. El bosque mediterráneo: Espacio ecológico, riqueza económica y bien social. Unasylva (Mediterranean Forests, FAO) 1999, 50, 3–15. [Google Scholar]
- Cachinero-Vivar, A.M.; Palacios-Rodríguez, G.; Lara-Gómez, M.A.; Navarro-Cerrillo, R.M. Thinning Effect of C Sequestration along an Elevation Gradient of Mediterranean Pinus spp. Plantations. Forests 2021, 12, 1583. [Google Scholar] [CrossRef]
- Pardos, M.; Vázquez-Piqué, J.; Benito, L.; Madrigal, G.; Alejano, R.; Fernández, M.; Calama, R. Does the Age of Pinus sylvestris Mother Trees Influence Reproductive Capacity and Offspring Seedling Survival? Forests 2022, 13, 937. [Google Scholar] [CrossRef]
- Tesei, G.; Allegrezza, M.; Ballelli, S.; Ciaschetti, G.; Console, C.; Montecchiari, S.; Ottaviani, C.; Pirone, G.; Frattaroli, A.R. The oldest Pinus nigra plantations in the central Apennines: Distribution and floristic-vegetational characterisation. Plant Sociol. 2021, 58, 15–28. [Google Scholar] [CrossRef]
- Zerbe, S. Restoration of natural broad-leaved woodland in Central Europe on sites with coniferous forest plantations. For. Ecol. Manag. 2002, 167, 27–42. [Google Scholar] [CrossRef]
- Hasenauer, H. Terms and Definitions Relevant for Conservation. Abstracts of the International Conference. In Proceedings of the Question of Conversion of Coniferous Forests, Freiburg, Germany, 27 September–2 October 2003; p. 16. [Google Scholar]
- Spiecker, H.; Hansen, J.; Klima, E.; Skovsgaard, J.; Sterba, H.; Teuffel, K. Norway Spruce Conversion-Options and Consequences; European Forest Institute Research Report: Joensuu, Finland, 2004; p. 18. [Google Scholar]
- Vizcaíno-Palomar, N.; Revuelta-Eugercios, B.; Zavala, M.A.; Alía, R.; González-Martínez, S.C. The role of population origin and micro environment in seedling emergence and early survival in Mediterranean maritime pine (Pinus pinaster Aiton). PLoS ONE 2014, 9, e109132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zlatanov, T.; Velichkov, I.; Lexer, M.J.; Dubravac, T. Regeneration dynamics in aging black pine (Pinus nigra Arn.) plantations on the south slopes of the Middle Balkan Range in Bulgaria. New For. 2010, 40, 289–303. [Google Scholar] [CrossRef]
- Lüpke, B.; Ammer, C.; Bruciamacchie, M.; Brunner, A.; Ceitel, J.; Collet, C.; Deuleuze, C.; Placido, J.; Huss, J.; Jankovič, J.; et al. Silvicultural Strategies for Conversion. Abstract of the International Conference. In Proceedings of the Question of Conversion of Coniferous Forests, Freiburg, Germany, 27 September–2 October 2003; pp. 25–26. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Belt | Vegetation Zone | Phytocenosis | Average Annual Precipitation | Average Annual Temperature | Pioneer Species | Climatogenic Species |
|---|---|---|---|---|---|---|
| Mediterranean-litoral | Sub-Mediterranean | Querco pubescenti-Carpinetum orientalis Horvatić 1939 | 1200 mm | 12–14 °C | Pinus nigra | Carpinus orientalis, Quercus pubescens |
| Mediterranean-montan | Epi-Mediterranean | Aristolochio luteae-Quercetum pubescentis (Horvat 1959) Poldini 2008 | 1400 mm | 10–12 °C | Pinus nigra | Ostrya carpinifolia, Quercus pubescens |
| Variables | Cum-R2 | Inc.-R2 | Random p | Gamma | R2-Max | R2-Min |
|---|---|---|---|---|---|---|
| Altitude | 0.1973 | 0.1973 | 0.0010 | 1.0000 | 0.1911 | 0.0001 |
| Slope | 0.3210 | 0.1072 | 0.0690 | 0.8972 | 0.3411 | 0.0023 |
| Org mat | 0.3922 | 0.0712 | 0.1960 | 0.2251 | 0.3685 | 0.0186 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barčić, D.; Hršak, V.; Rosavec, R.; Ančić, M. Ecological Potential of Mediterranean Habitats for Black Pine (Pinus nigra J.F. Arnold) in Croatia. Forests 2022, 13, 1900. https://doi.org/10.3390/f13111900
Barčić D, Hršak V, Rosavec R, Ančić M. Ecological Potential of Mediterranean Habitats for Black Pine (Pinus nigra J.F. Arnold) in Croatia. Forests. 2022; 13(11):1900. https://doi.org/10.3390/f13111900
Chicago/Turabian StyleBarčić, Damir, Vladimir Hršak, Roman Rosavec, and Mario Ančić. 2022. "Ecological Potential of Mediterranean Habitats for Black Pine (Pinus nigra J.F. Arnold) in Croatia" Forests 13, no. 11: 1900. https://doi.org/10.3390/f13111900