
Changes in the Potential Habitat Distribution of Typical Fire-Resistant Forest Species under Climate Change in the Subtropical Regions of China
Abstract
:1. Introduction
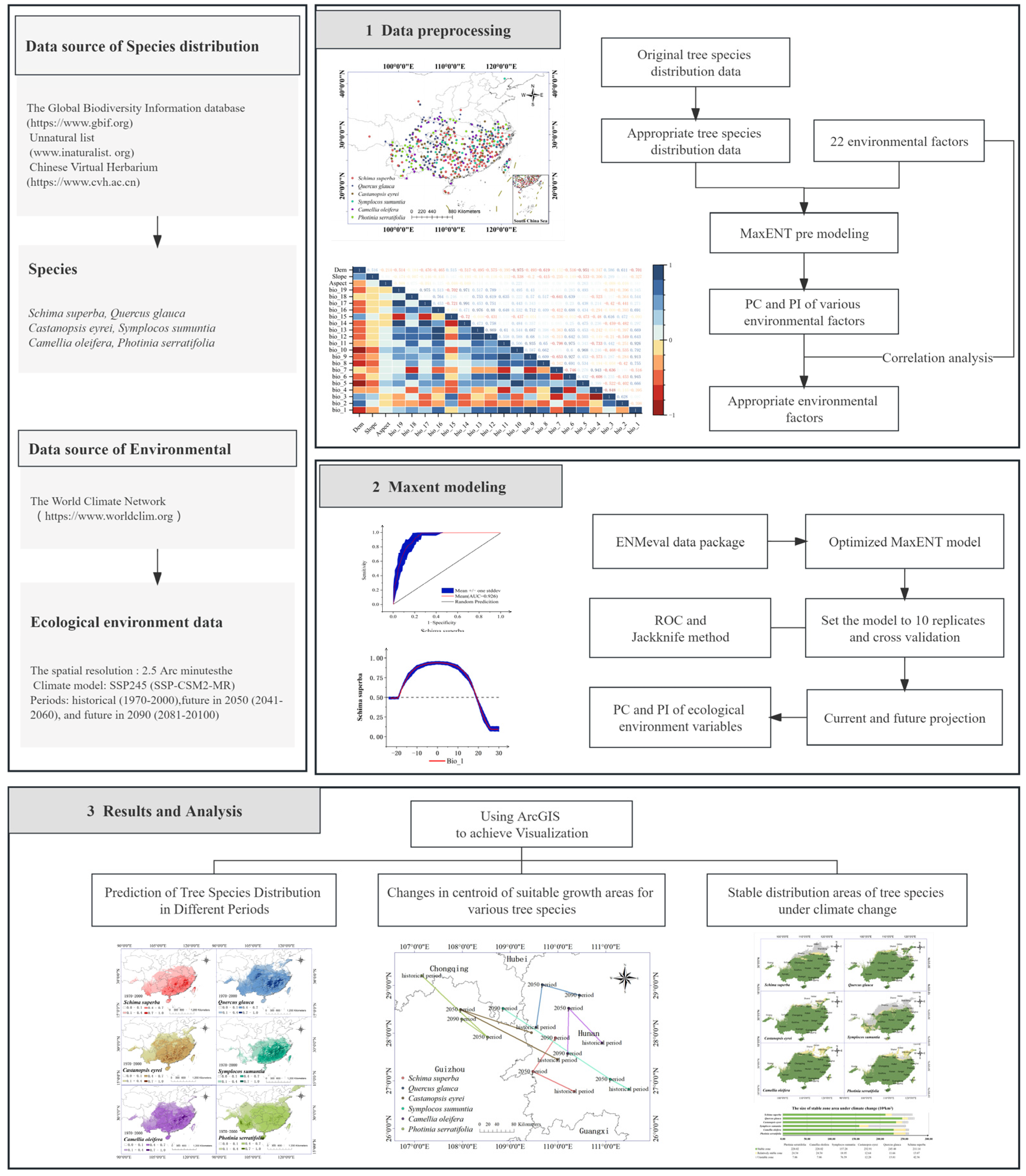
2. Materials and Methods
2.1. Materials
2.1.1. Overview of the Research Area
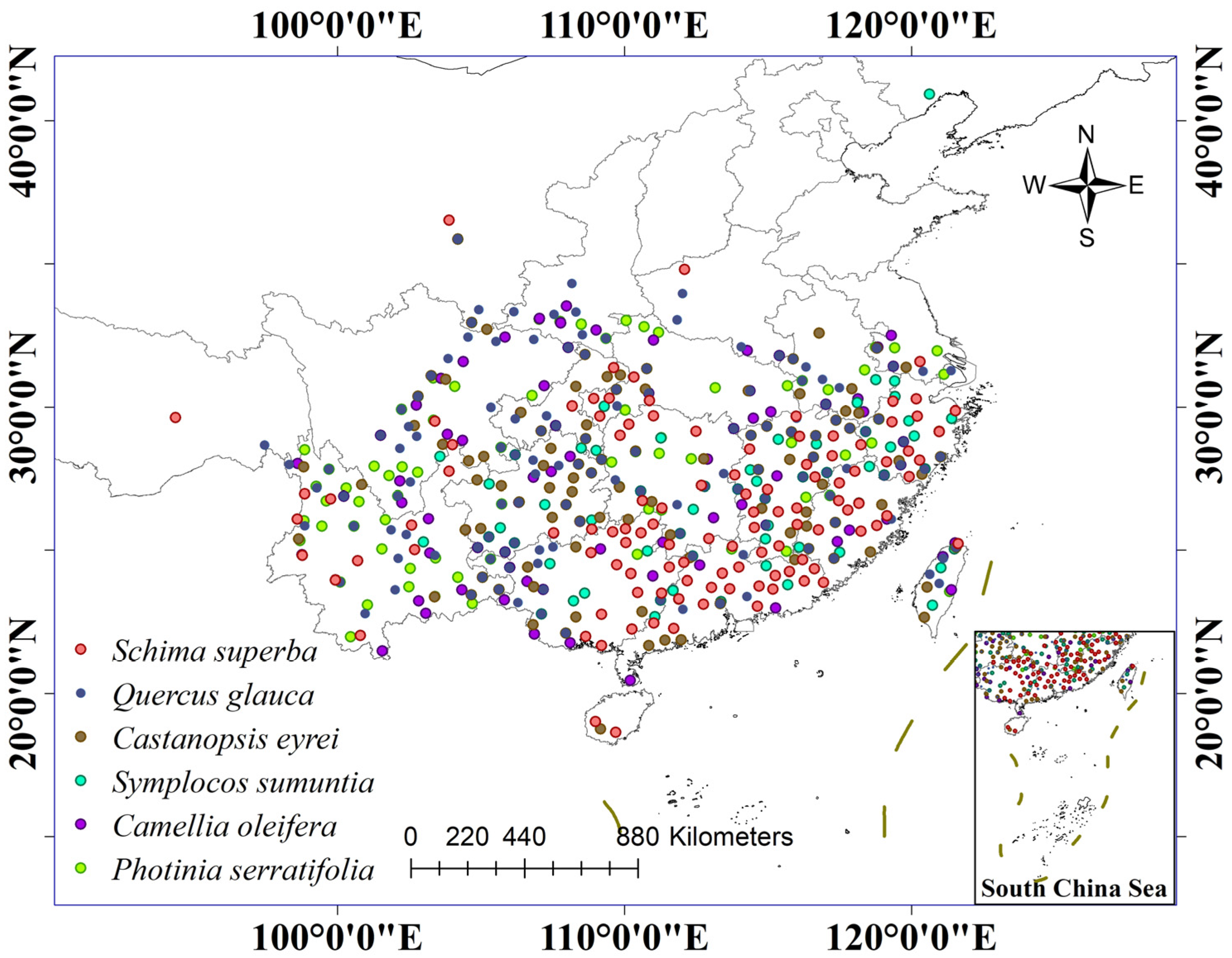
2.1.2. Species Distribution Data
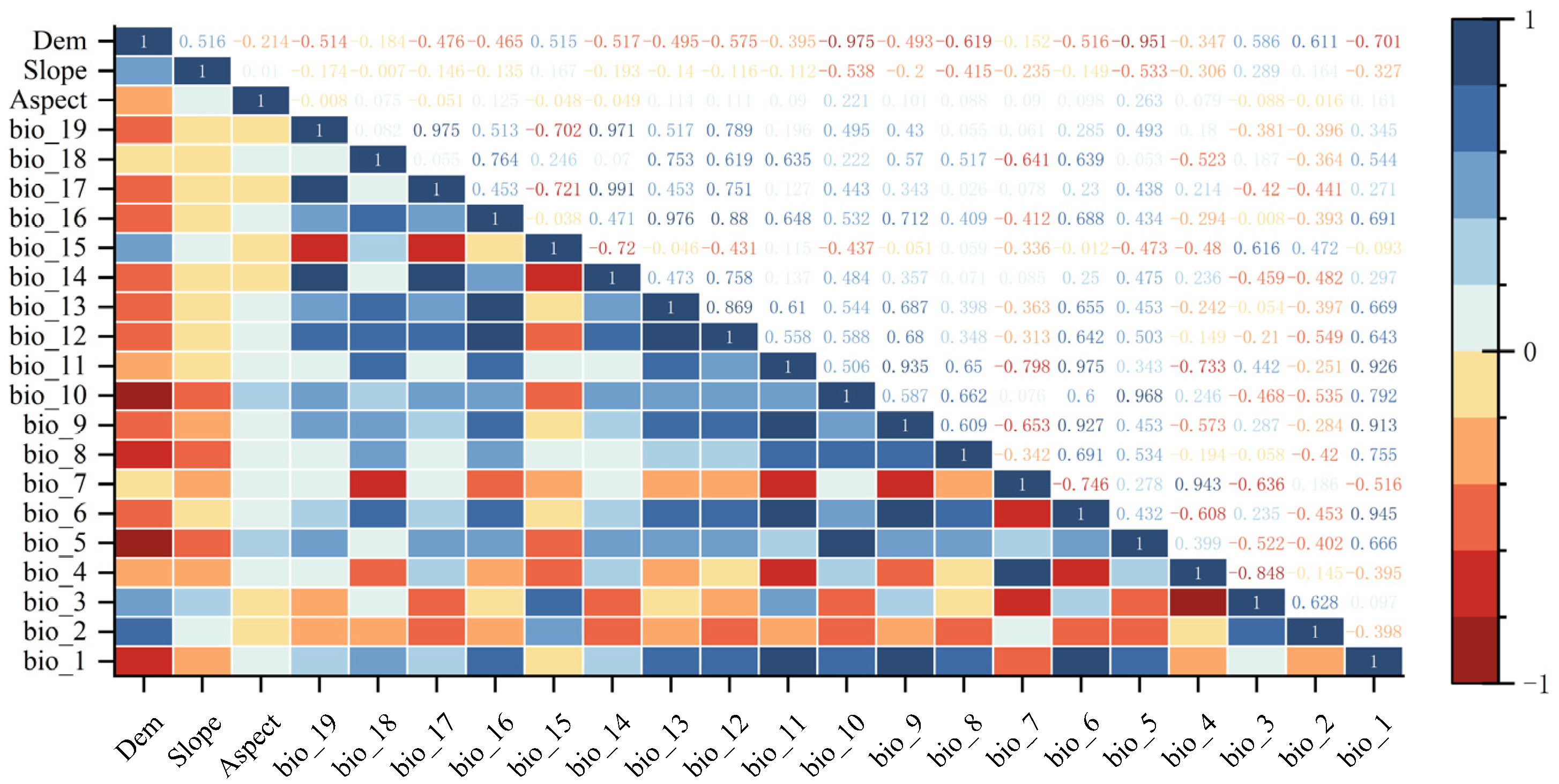
2.1.3. Ecological Environment Data
2.1.4. MaxEnt Model
2.2. Methods
2.2.1. Data Preprocessing
2.2.2. Precision Evaluation
2.2.3. Model Evaluation
2.2.4. Model Evaluation and Analytical Methods
3. Results and Analysis
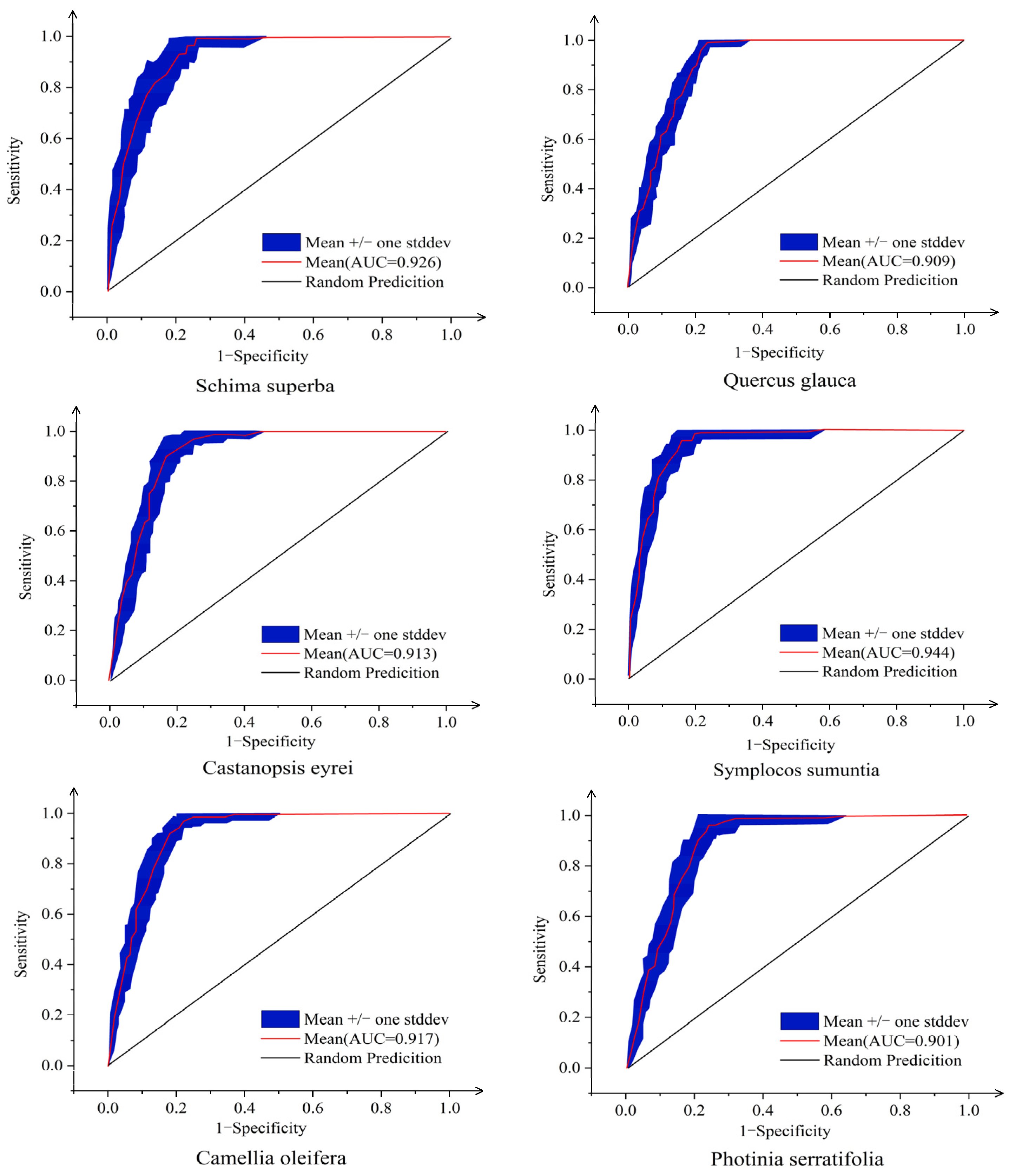
3.1. Model Accuracy
3.2. Main Environmental Factors Affecting the Distribution of Tree Species during Historical Periods
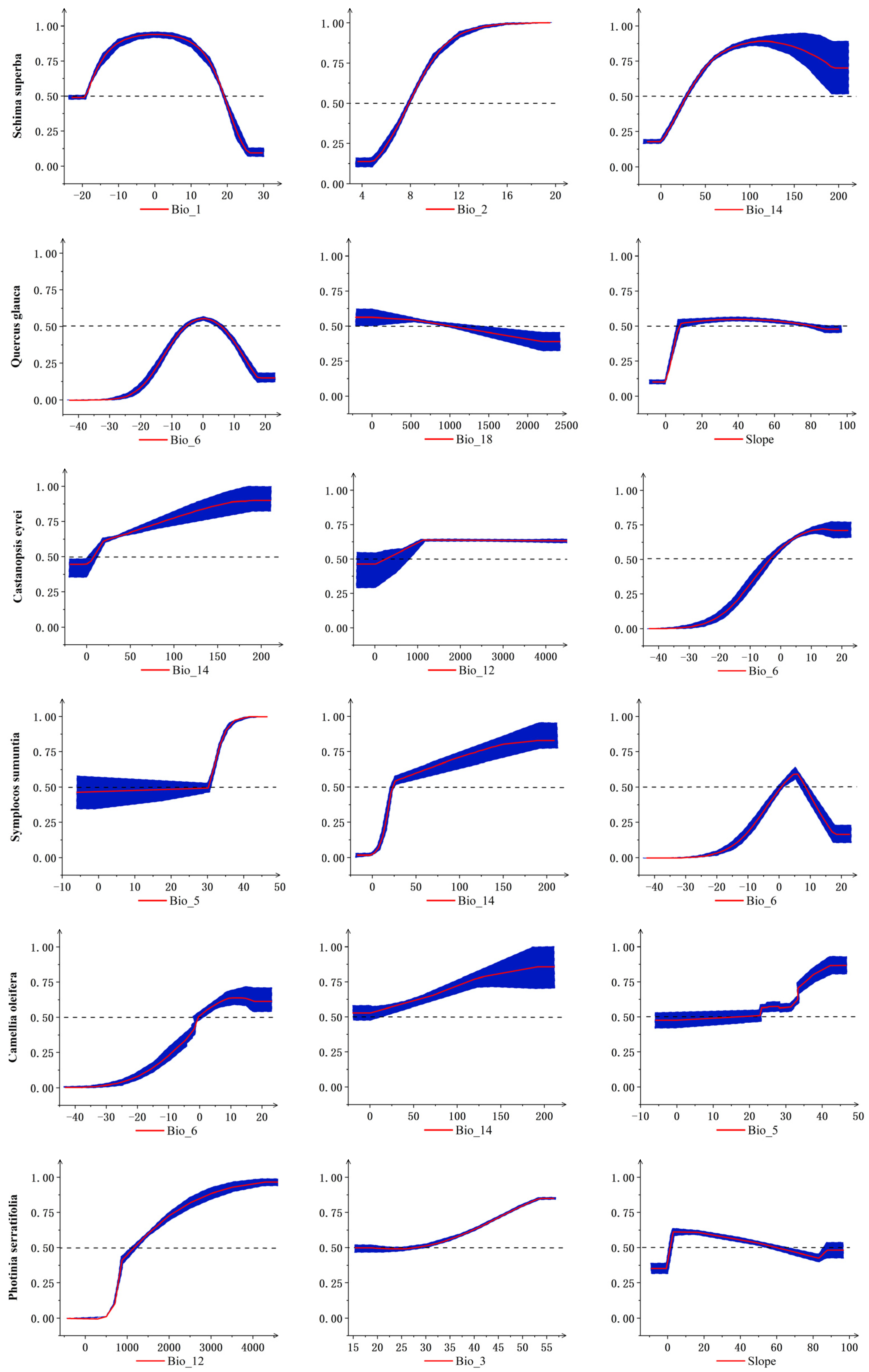
3.3. Response of Tree Species to Major Environmental Factors during Historical Periods
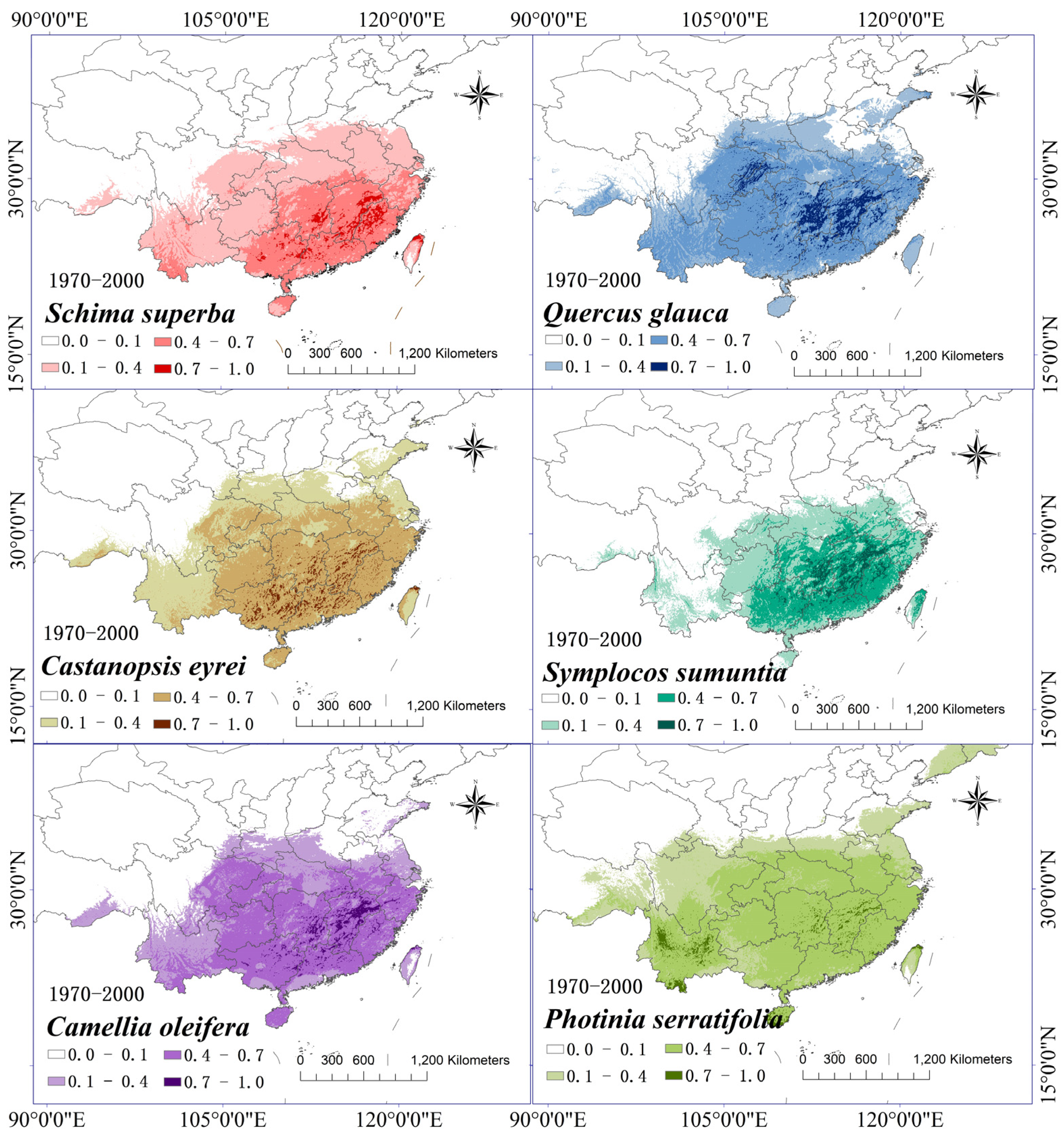
3.4. Distribution Prediction of Tree Species during Historical Periods
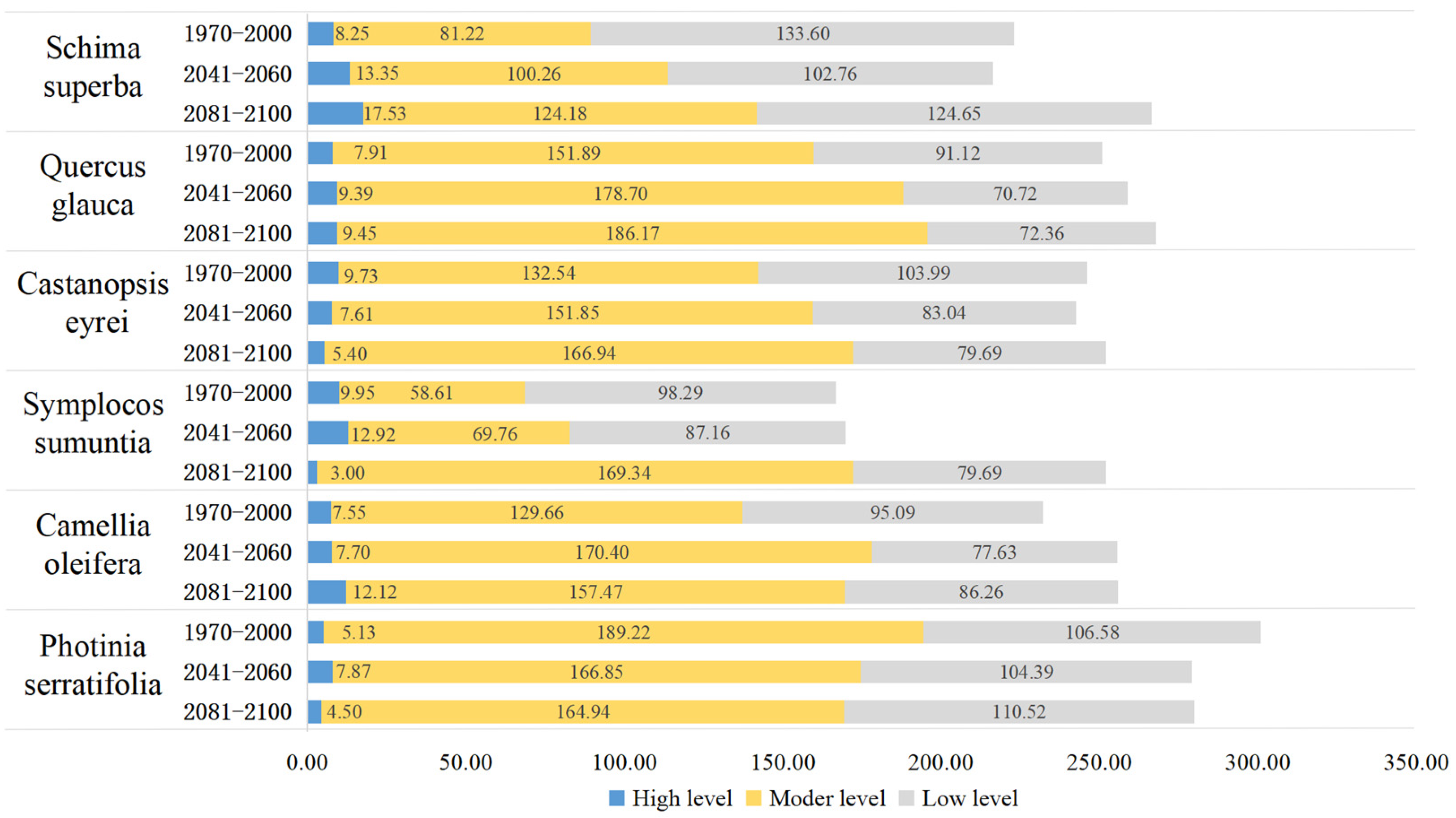
3.5. Changes in the Area of Suitable Growth Areas for Tree Species under Climate Change
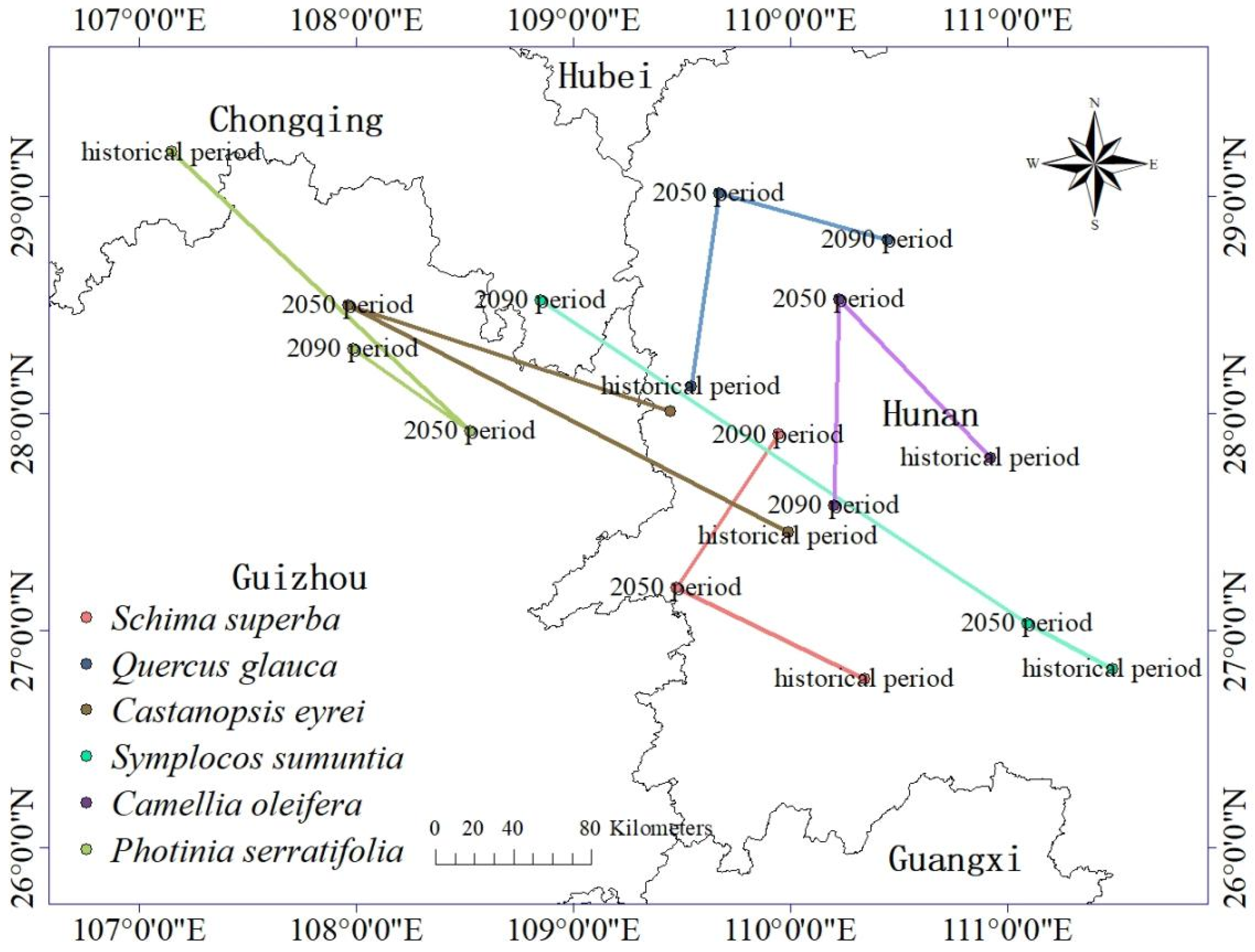
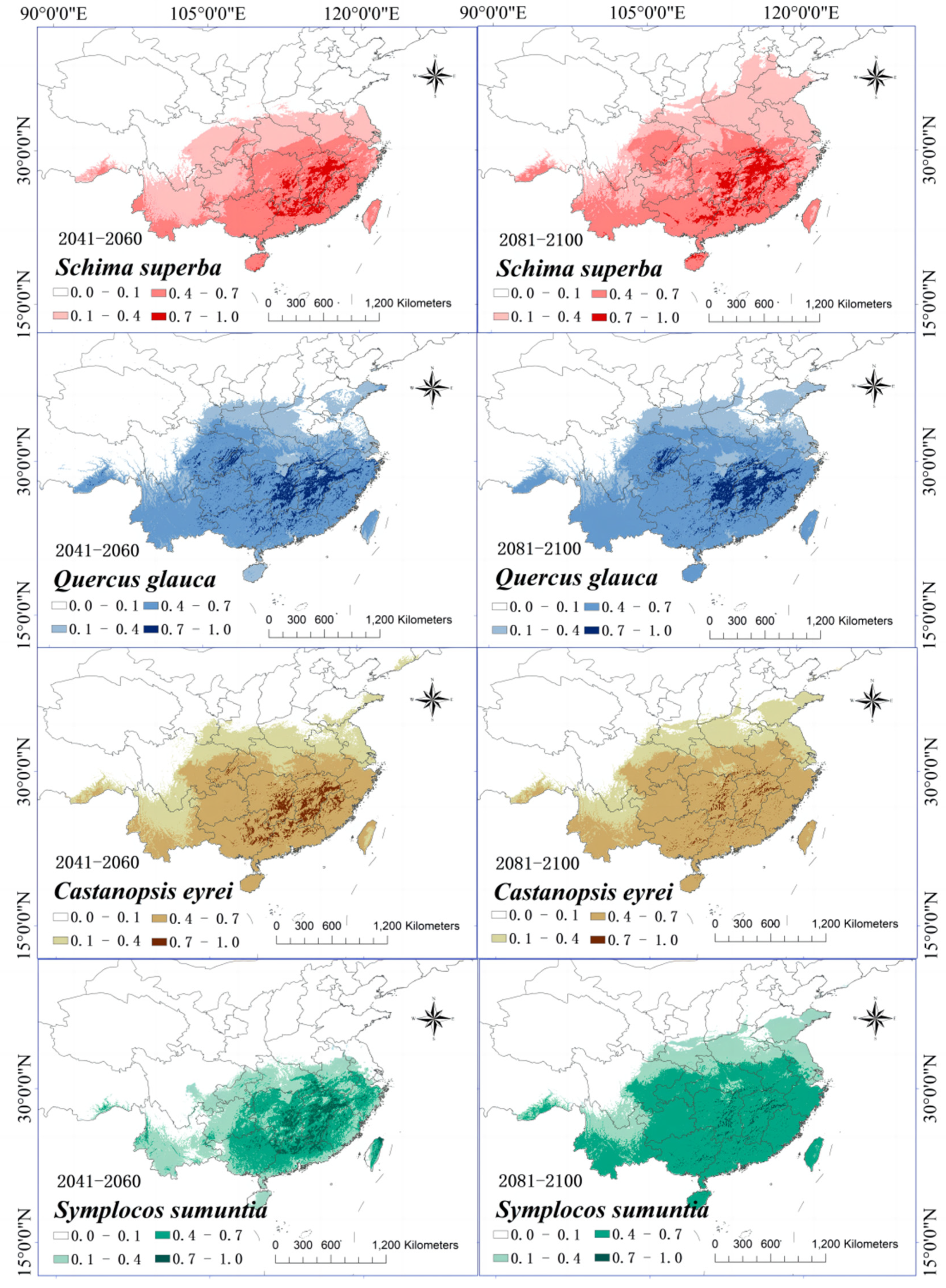
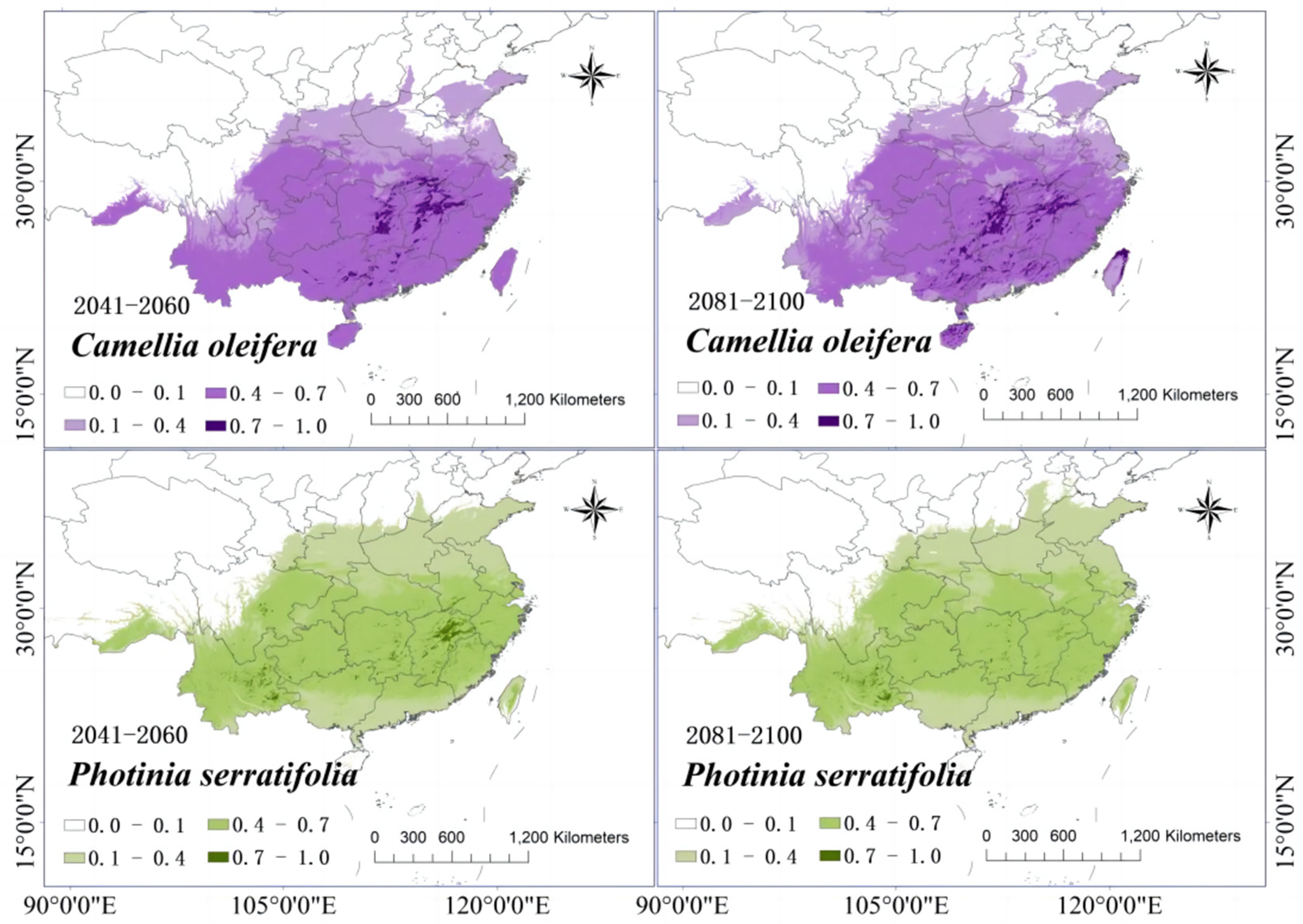
3.6. Changes in the Distribution of Suitable Growth Areas of Tree Species When under Climate Change
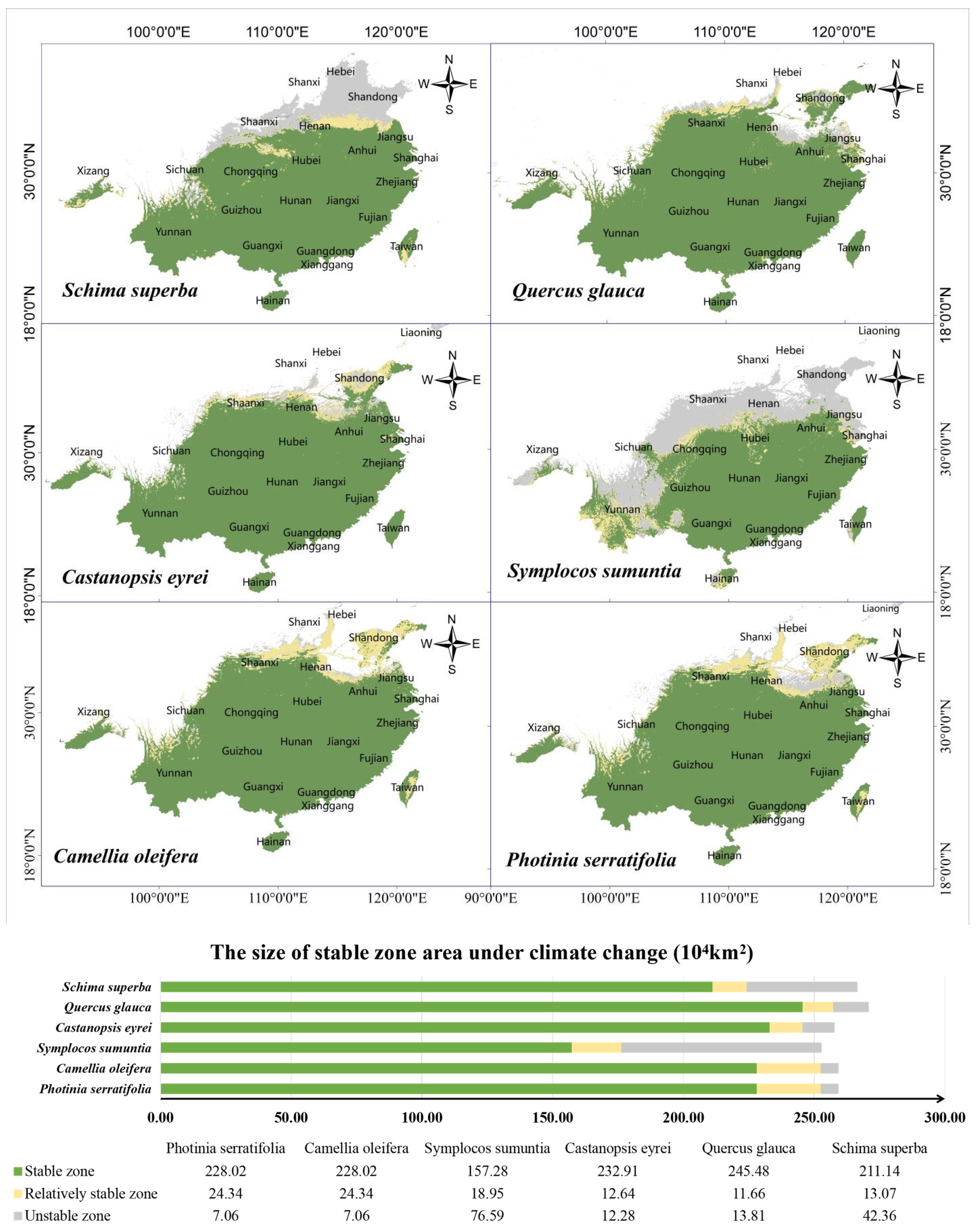
3.7. Stable and Suitable Distribution of Tree Species under Climate Change
4. Discussion
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Curran, T.J.; Perry GL, W.; Wyse, S.V.; Alam, M.A. Managing fire and biodiversity in the wildland-urban interface: A role for green firebreaks. Fire 2017, 1, 3. [Google Scholar] [CrossRef]
- Si, L.; Shu, L.; Wang, M.; Zhao, F. Review on the Difference of Fireproof Efficiency of Biological Fire-resistance Forest Belt. Terr. Ecosyst. Conserv. 2022, 2, 53–59. [Google Scholar]
- Platt, K.; Jackman, E.R. The Cheatgrass Problem in Oregon; Federal Cooperative Extension Service, Oregon State College: Corvallis, OR, USA, 1946. [Google Scholar]
- Shu, L.; Tian, X.; Li, H. Research progress on fire resistant forest belts. For. Sci. 1999, 35, 80–85. [Google Scholar]
- Ryu, S.R.; Choi, H.T.; Lim, J.H.; Lee, I.-K.; Ahn, Y.-S. Post-fire restoration plan for sustainable forest management in South Korea. Forests 2017, 8, 188. [Google Scholar] [CrossRef]
- Xiong, D.; Xiaoming, L.; Ximing, Z.; Wanhui, Y.; Forezki, A.; Runge, M. Studies on gas exchange of Tamarix ramosissima Labd. Acta Ecol. Sin. 2003, 23, 180–187. [Google Scholar]
- Cui, X.; Alam, M.A.; Perry, G.L.; Paterson, A.M.; Wyse, S.V.; Curran, T.J. Green firebreaks as a management tool for wildfires: Lessons from China. J. Environ. Manag. 2019, 233, 329–336. [Google Scholar] [CrossRef]
- Deng, Y. Characteristics and Prevention and Control Strategies of Forest Fires in Zijin County. Low Carbon World 2022, 12, 187–189. [Google Scholar]
- Lai, G. Exploration on the Management and Benefits of Oil Tea Fireproof Forest Belt Tending in Sanming Mountain Area. J. Green Sci. Technol. 2017, 147–148. [Google Scholar]
- Xia, C.; Wang, C. Analysis on biological fire prevention project in Greater Khingan forest area. For. Fire Prev. 2004, 24–25. [Google Scholar]
- Ou, T.-H.; Qian, W.-H. Vegetation variations along the monsoon boundary zone in East Asia. Chin. J. Geophys. 2006, 49, 627–636. [Google Scholar] [CrossRef]
- Tian, X.; Shu, L. The application and research of fire break forest belts. World For. Res. 2000, 13, 20–26. [Google Scholar]
- Gu, W.; Lu, Z.; Huang, C.; Li, Y.; Guan, Y. Screening study of fire resistant tree species in Jianshui County, Yunnan Province of southwestern China. J. Beijing For. Univ. 2020, 42, 49–60. [Google Scholar]
- Zeng, S.-P.; Liu, F.-L.; Zhao, M.-F.; Ai, Y.; Chen, X.-W. Age-and organ-related variances in fire resistance traits of typical tree species in subtropical China. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2020, 31, 1063–1072. [Google Scholar]
- Wang, X.; Liu, G.; Xiao, T. Suitability Characteristics of Camellia oleifera Growth under Climate Change Scenarios. Trop. Geogr. 2020, 40, 868–880. [Google Scholar]
- Liu, X.-T.; Yuan, Q.; Ni, J. Research advances in modelling plant species distribution in China. Chin. J. Plant Ecol. 2019, 43, 273–283. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Liu, L.; Nie, Y. Predicting Potential Distribution of Tibetan Spruce (Picea smithiana) in Qomolangma (Mount Everest) National Nature Preserve Using Maximum Entropy Niche-based Model. Chin. Geogr. Sci. 2011, 21, 417–426. [Google Scholar] [CrossRef]
- Miao, J.; Wang, Y.; Wang, L.; Xu, X. Prediction of potential geographical distribution pattern change for Castanopsis sclerophylla on MaxEnt. J. Nanjing Fore. Univ. 2021, 45, 193–198. [Google Scholar]
- Halvorsen, R. A strict maximum likelihood explanation of MaxEnt, and some implications for distribution modelling. Sommerfeltia 2013, 36, 1–132. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENM eval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Zhang, S.-H.; Bai, H.-Y.; Qi, G.-Z.; Liang, J. Changes of climate zone boundary of the Qinling Mountains from 1960 to 2019. J. Nat. Resour. 2021, 36, 2491–2506. [Google Scholar] [CrossRef]
- Yan, X.; MacKinnon, J.; Li, D. Study on biogeographical divisions of China. Biodivers. Conserv. 2004, 13, 1391–1417. [Google Scholar]
- Du, L.; Wu, Z.; Wu, Q.; Guo, L.; Dong, X.; Lin, S. Temporal and Spatial Correlation Between Forest Loss and Forest Fire in the Ecological Region of Southern China. J. Southwest For. Univ. 2022, 42, 138–147. [Google Scholar]
- Tan, J.; Huang, A.; Shi, X.; Zhang, Y.; Zhang, Y.; Cao, L.; Wu, Y. Evaluating the Performance of BCC-CSM2-MR Model in Simulating the Land Surface Processes in China. Plateau Meteorol. 2022, 41, 1335–1347. [Google Scholar]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. A maximum entropy approach to species distribution modeling. In Proceedings of the Twenty-First International Conference on Machine Learning, Banff, AB, Canada, 4–8 July 2004. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Kumar, P. Assessment of impact of climate change on Rhododendrons in Sikkim Himalayas using Maxent modelling: Limitations and challenges. Biodivers. Conserv. 2012, 21, 1251–1266. [Google Scholar] [CrossRef]
- Zhao, G.; Cui, X.; Sun, J.; Li, T.; Wang, Q.; Ye, X.; Fan, B. Analysis of the distribution pattern of Chinese Ziziphus jujuba under climate change based on optimized biomod2 and MaxEnt models. Ecol. Ind. 2021, 132, 108256. [Google Scholar] [CrossRef]
- Syfert, M.M.; Smith, M.J.; Coomes, D.A. The effects of sampling bias and model complexity on the predictive performance of MaxEnt species distribution models. PLoS ONE 2013, 8, e55158. [Google Scholar] [CrossRef]
- Kumar, S.; Stohlgren, T.J. Maxent modeling for predicting suitable habitat for threatened and endangered tree Canacomyrica monticola in New Caledonia. J. Ecol. Nat. Environ. 2009, 1, 94–98. [Google Scholar]
- Wang, Y.; Xie, B.; Wan, F.; Xiao, Q.; Dai, L. Application of ROC curve analysis in evaluating performance of alien species’ potential distribution models. Biodivers. Sci. 2007, 15, 365–372. [Google Scholar]
- Bradley, A.P. The use of the area under the ROC curve in the evaluation of machine learning algorithms. Pattern Recognit. 1997, 30, 1145–1159. [Google Scholar] [CrossRef]
- Liu, B.; Gao, X.; Zheng, K.; Ma, J.; Jiao, Z.; Xiao, J.; Wang, H. The potential distribution and dynamics of important vectors Culex pipiens pallens and Culex pipiens quinquefasciatus in China under climate change scenarios: An ecological niche modelling approach. Pest Manag. Sci. 2020, 76, 3096–3107. [Google Scholar] [CrossRef] [PubMed]
- Pulliam, H.R. On the relationship between niche and distribution. Ecol. Lett. 2000, 3, 349–361. [Google Scholar] [CrossRef]
- Bowers, A.J.; Zhou, X. Receiver operating characteristic (ROC) area under the curve (AUC): A diagnostic measure for evaluating the accuracy of predictors of education outcomes. J. Educ. Stud. Placed Risk 2019, 24, 20–46. [Google Scholar] [CrossRef]
- Carrington, A.M.; Manuel, D.G.; Fieguth, P.W.; Ramsay, T.; Osmani, V.; Wernly, B.; Bennett, C.; Hawken, S.; McInnes, M.; Magwood, O. Deep ROC analysis and AUC as balanced average accuracy to improve model selection, understanding and interpretation. arXiv 2021, arXiv:2103.11357. [Google Scholar]
- Arias, P.; Bellouin, N.; Coppola, E.; Jones, G.; Krinner, J.; Marotzke, V.; Naik, M.D.; Palmer, G.-K.; Plattner, J.; Rogelj, M.; et al. Climate Change 2021: The Physical Science Basis. In Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Technical Summary; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Shaohong, W.; Qingchen, C.; Jiangbo, G.; Liu, L.; Feng, A.; Deng, H.; Zuo, L.; Liu, W. Identification of regional pattern of climate change risk in China under different global warming targets. J. Geogr. Sci. 2023, 33, 429–448. [Google Scholar]
- Brooker, R.; Young, J.C.; Watt, A.D. Climate change and biodiversity: Impacts and policy development challenges–a European case study. Int. J. Biodivers. Sci. Manag. 2007, 3, 12–30. [Google Scholar] [CrossRef]
- Hays, G.C.; Richardson, A.J.; Robinson, C. Climate change and marine plankton. Trends Ecol. Evol. 2005, 20, 337–344. [Google Scholar] [CrossRef]
- Sun, S.; Zhang, Y.; Huang, D.; Wang, H.; Cao, Q.; Fan, P.; Yang, N.; Zheng, P.; Wang, R. The effect of climate change on the richness distribution pattern of oaks (Quercus L.) in China. Sci. Total Environ. 2020, 744, 140786. [Google Scholar] [CrossRef] [PubMed]
- Jian, N. Relationship between geographical distribution of Schima superba, its forest and climate in China. J. Plant Resour. Environ. 1996, 5, 28–34. [Google Scholar]
- Ouyang, Z.Y.; Li, Z.H.; Ouyang, S.L.; Chen, Y.; Zhou, Z.; Wu, J. Prediction of the potential distribution of Cyclobalanopsis gilva in China based on the Maxent and ArcGIS model. J. Central South Univ. For. Technol. 2023, 43, 19–26. [Google Scholar]
- Cao, M.C.; Zhou, G.S.; Weng, E.S. Application and comparison of generalized models and classification and regression tree in simulating tree species distribution. Acta Ecol. Sin. 2005, 25, 2031–2040. [Google Scholar]
- Pan, L.; Yang, T.; Liao, N.; Tong, X.; He, L.; Liu, B. Medicinal Value and Biological Prospect Prediction of Symplocos. J. Anhui Agric. Sci. 2018, 46, 8–11. [Google Scholar]
- Wen, K.; Ouyang, Y. Study on bark fire resistance of 11 evergreen broadleaved tree species. South China For. Sci. 2018, 46, 50–53. [Google Scholar]
- Jing, M.; Zhu, L.; Cherubini, P.; Yuan, D.; Li, Z.; Wang, X.; Liu, S. Responses of radial growth of Pinus massoniana and Castanopsis eyrei to climate change at different elevations in south China. Ecol. Ind. 2022, 145, 109602. [Google Scholar] [CrossRef]
- Su, X.; Chen, S.; Tong, J. The preliminary study of the correlation between the distribution of main tree species and the climate factors in Fujian Province. J. Fujian Coll. For. 2001, 21, 371–375. [Google Scholar]
- Pan, X.; Chi, H.; Lin, H.; Du, H.; Huang, S.; Jin, K.; Lu, L.; Han, S. Preliminary report on the introduction experiment of angular alum in coastal areas of Wenzhou. J. Green Sci. Technol. 2019, 45–49. [Google Scholar]
- Hu, J.; Wu, W.; Cao, Z.; Wen, J.; Shu, Q.; Fu, S. Morphological, physiological and biochemical responses of Camellia oleifera to low-temperature stress. Pak. J. Bot. 2016, 48, 899–905. [Google Scholar]
- Liang, H.-H.; Chen, S.; Zhao, B.; Zhong, Y.-T.; Ma, W.-J.; Li, Y. A Comparative Study on Inundation Tolerance of 7 hrub Seedling under Waterlogging Stress. J. Northwest For. Univ. 2020, 35, 61–67. [Google Scholar]
- Wang, Q.; Yu, W.; Li, Y.; Wang, H.; Sun, J. Comprehensive evaluation of cold tolerance on 6 evergreen broad—Left tree specifications. J. Shandong For. Sci. Technol. 2016, 46, 14–19+6. [Google Scholar]
- Wu, H.; Hou, W.; Qian, Z.-H.; Hu, J.-G. The research on the sensitivity of climate change in China in recent 50 years based on composite index. Acta Phys. Sin. 2012, 61, 149205. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Environmental Factor | Unit |
|---|---|---|
| Bio_1 | Annual Mean Temperature | °C |
| Bio_2 | Mean Diurnal Range | °C |
| Bio_3 | Isothermality | % |
| Bio_4 | Temperature Seasonality | °C |
| Bio_5 | Max. Temperature of Warmest Month | °C |
| Bio_6 | Min. Temperature of Coldest Month | °C |
| Bio_7 | Temperature Annual Range | °C |
| Bio_8 | Mean Temperature of Wettest Quarter | °C |
| Bio_9 | Mean Temperature of Driest Quarter | °C |
| Bio_10 | Mean Temperature of Warmest Quarter | °C |
| Bio_11 | Mean Temperature of Coldest Quarter | °C |
| Bio_12 | Annual Precipitation | mm |
| Bio_13 | Precipitation of Wettest Month | mm |
| Bio_14 | Precipitation of Driest Month | mm |
| Bio_15 | Precipitation Seasonality | % |
| Bio_16 | Precipitation of Wettest Quarter | mm |
| Bio_17 | Precipitation of Driest Quarter | mm |
| Bio_18 | Precipitation of Warmest Quarter | mm |
| Bio_19 | Precipitation of Coldest Quarter | mm |
| Dem | m | |
| Aspect | ° | |
| Slope | ° |
| FCs | RM | |||||||
|---|---|---|---|---|---|---|---|---|
| L | 0.5 | 1 | 1.5 | 2 | 2.5 | 3 | 3.5 | 4 |
| LQ | 0.5 | 1 | 1.5 | 2 | 2.5 | 3 | 3.5 | 4 |
| H | 0.5 | 1 | 1.5 | 2 | 2.5 | 3 | 3.5 | 4 |
| LQH | 0.5 | 1 | 1.5 | 2 | 2.5 | 3 | 3.5 | 4 |
| LQHP | 0.5 | 1 | 1.5 | 2 | 2.5 | 3 | 3.5 | 4 |
| LQHPT | 0.5 | 1 | 1.5 | 2 | 2.5 | 3 | 3.5 | 4 |
| Tree Species | Ecological Environment Variables |
|---|---|
| Schima superba | Bio_1, Bio_2, Bio_3, Bio_5, Bio_7, Bio_14, Bio_18, Aspect, Slope |
| Quercus glauca | Bio_5, Bio_6, Bio_7, Bio_8, Bio_12, Bio_18, Aspect, Slope |
| Castanopsis eyrei | Bio_2, Bio_4, Bio_6, Bio_12, Bio_14, Aspect, Slope |
| Symplocos sumuntia | Bio_2, Bio_3, Bio_5, Bio_8, Bio_14, Bio_15, Aspect, Slope, dem |
| Camellia oleifera | Bio_2, Bio_4, Bio_5, Bio_6, Bio_8, Bio_14, Bio_18, Aspect, Slope |
| Photinia serratifolia | Bio_3, Bio_12, Bio_18, Aspect, Slope |
| Tree Species | Type | FC | RM | Delta AICc | OR10 |
|---|---|---|---|---|---|
| Schima superba | Optimum | LQ | 1 | 0 | 0.14 |
| Default | LQHPT | 1 | 55.55 | 0.21 | |
| Quercus glauca | Optimum | LQHP | 1.5 | 0 | 0.14 |
| Default | LQHPT | 1 | 28.07 | 0.27 | |
| Castanopsis eyrei | Optimum | LQH | 2 | 0 | 0.18 |
| Default | LQHPT | 1 | 30.43 | 0.23 | |
| Symplocos sumuntia | Optimum | LQH | 1.5 | 0 | 0.21 |
| Default | LQHPT | 1 | 66.56 | 0.28 | |
| Camellia oleifera | Optimum | LQHPT | 2 | 0 | 0.19 |
| Default | LQHPT | 1 | 47.10 | 0.24 | |
| Photinia serratifolia | Optimum | LQHP | 2 | 0 | 0.40 |
| Default | LQHPT | 1 | 25.99 | 0.54 |
| Tree Species | Variable | PC | PI |
|---|---|---|---|
| Schima superba | Bio_1 | 59.8 | 2.5 |
| Bio_14 | 16.1 | 3 | |
| Bio_2 | 12.5 | 3.8 | |
| Quercus glauca | Bio_6 | 66.8 | 33.4 |
| Bio_18 | 13.8 | 0.2 | |
| Slope | 7.2 | 6.3 | |
| Castanopsis eyrei | Bio_14 | 57.9 | 3.6 |
| Bio_12 | 20.7 | 3.6 | |
| Bio_6 | 7.7 | 67.4 | |
| Symplocos sumuntia | Bio_14 | 73.4 | 39.5 |
| Bio_5 | 9.3 | 6 | |
| Bio_6 | 5.7 | 47.9 | |
| Camellia oleifera | Bio_6 | 68.4 | 59.7 |
| Bio_14 | 7.8 | 1 | |
| Bio_5 | 5.6 | 3.1 | |
| Photinia serratifolia | Bio_12 | 82.3 | 89.5 |
| Bio_3 | 7.7 | 3 | |
| Slope | 4.4 | 2.1 |
| Tree Species | Historic Period | Historic Period to 2050 | 2050 | Future 2050 to 2090 | 2090 | Historic Period to 2090 | |||
|---|---|---|---|---|---|---|---|---|---|
| Area Change | Change Amplitude | Area Change | Change Amplitude | Area Change | Change Amplitude | ||||
| Schima superba | 223.06 | −6.70 | −3.00% | 216.37 | 49.99 | 23.10% | 266.36 | 43.29 | 19.41% |
| Quercus glauca | 250.92 | 7.90 | 3.15% | 258.82 | 9.17 | 3.54% | 267.99 | 17.07 | 6.80% |
| Castanopsis eyrei | 246.26 | −3.75 | −1.52% | 242.51 | 9.51 | 3.92% | 252.02 | 5.76 | 2.34% |
| Symplocos sumuntia | 166.84 | 3.00 | 1.80% | 169.85 | 82.17 | 48.38% | 252.02 | 85.17 | 51.05% |
| Camellia oleifera | 232.29 | 23.44 | 10.09% | 255.73 | 0.12 | 0.05% | 255.85 | 23.56 | 10.14% |
| Photinia serratifolia | 300.92 | −21.81 | −7.25% | 279.11 | 0.84 | 0.30% | 279.95 | −20.97 | −6.97% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouyang, W.; Qiu, H.; Chen, Z.; Wu, Y.; Li, J. Changes in the Potential Habitat Distribution of Typical Fire-Resistant Forest Species under Climate Change in the Subtropical Regions of China. Forests 2023, 14, 1897. https://doi.org/10.3390/f14091897
Ouyang W, Qiu H, Chen Z, Wu Y, Li J. Changes in the Potential Habitat Distribution of Typical Fire-Resistant Forest Species under Climate Change in the Subtropical Regions of China. Forests. 2023; 14(9):1897. https://doi.org/10.3390/f14091897
Chicago/Turabian StyleOuyang, Wenxin, Hanqing Qiu, Zhiming Chen, Yiheng Wu, and Jianjun Li. 2023. "Changes in the Potential Habitat Distribution of Typical Fire-Resistant Forest Species under Climate Change in the Subtropical Regions of China" Forests 14, no. 9: 1897. https://doi.org/10.3390/f14091897