
Transcriptome Analysis Reveals the Hormone Signalling Coexpression Pathways Involved in Adventitious Root Formation in Populus
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Annotation and DEG Analysis
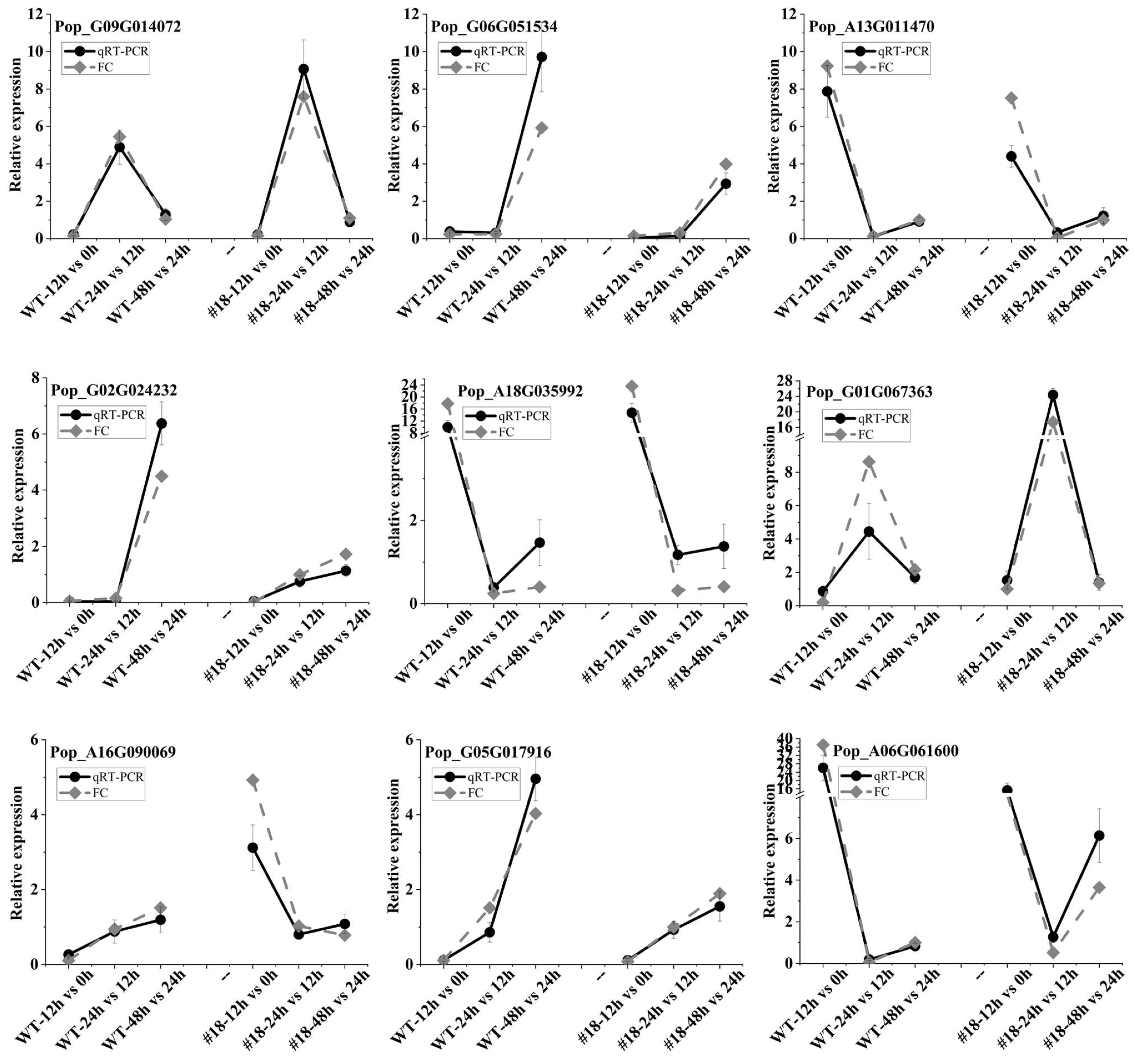
2.2. RNA Isolation and qRT-PCR
3. Results
3.1. PagFBL1-OE Stimulates the Remodelling of Gene Expression in Transgenic Poplar
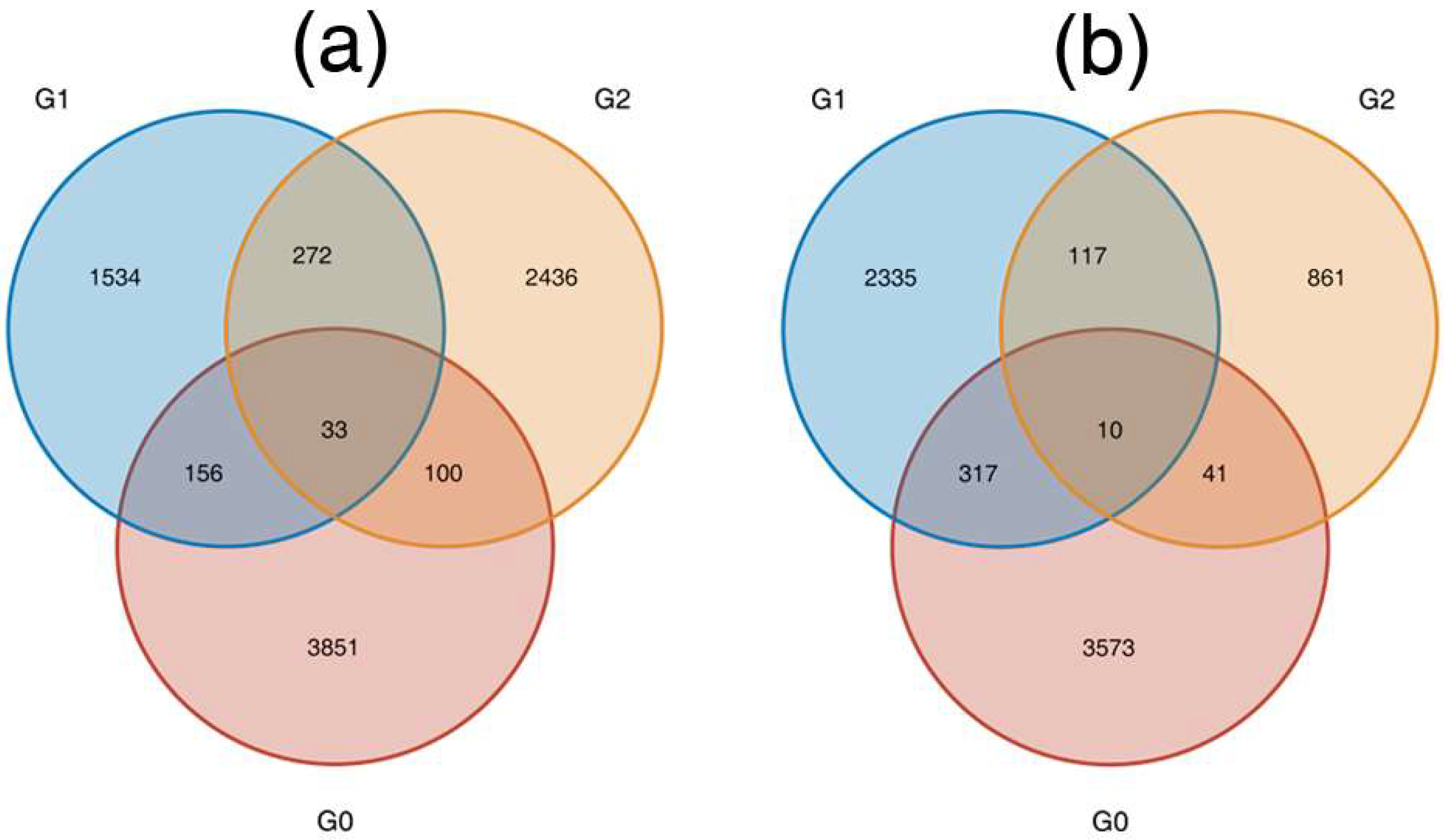
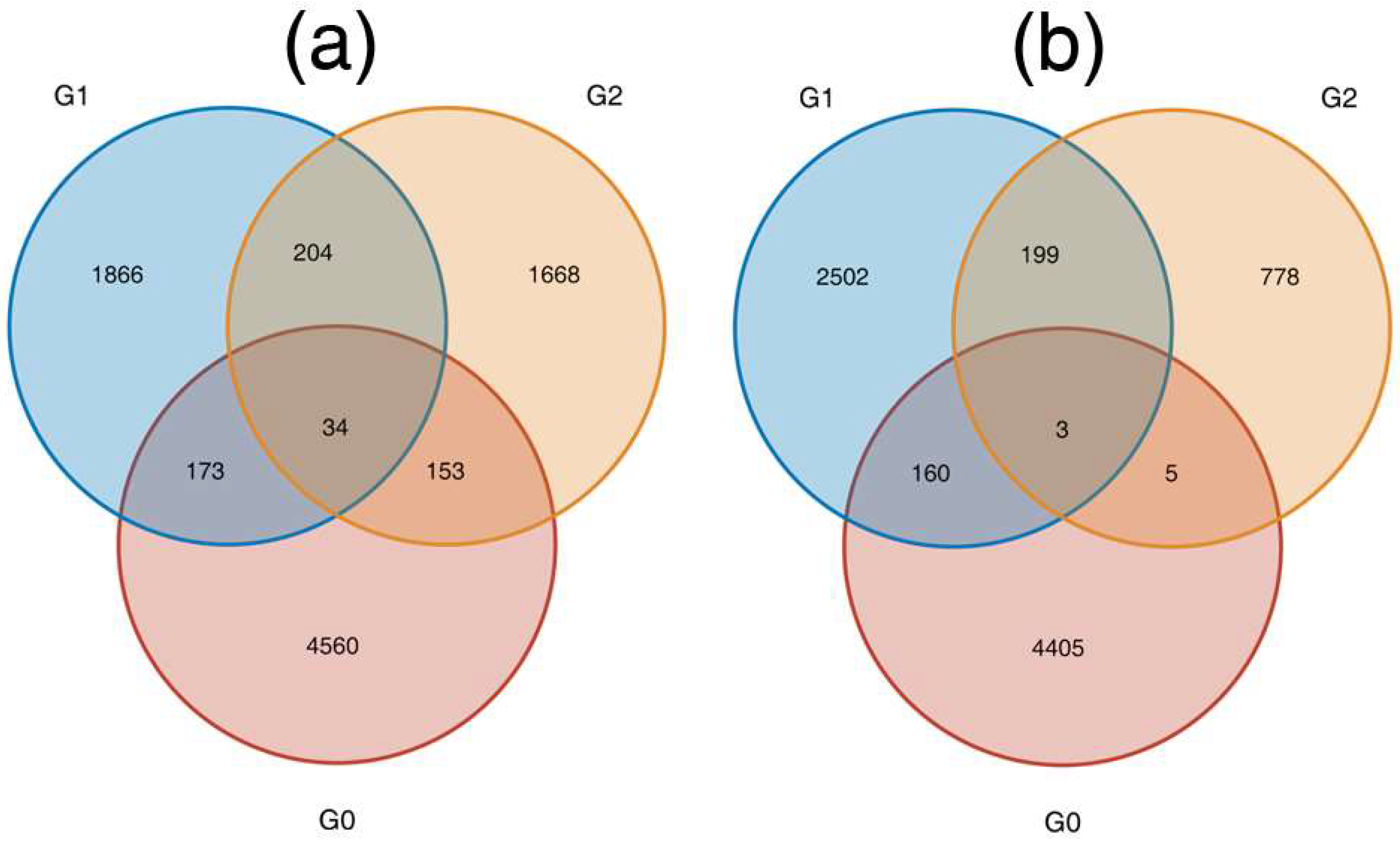
3.2. NOG and KEGG Pathway Enrichment Analysis
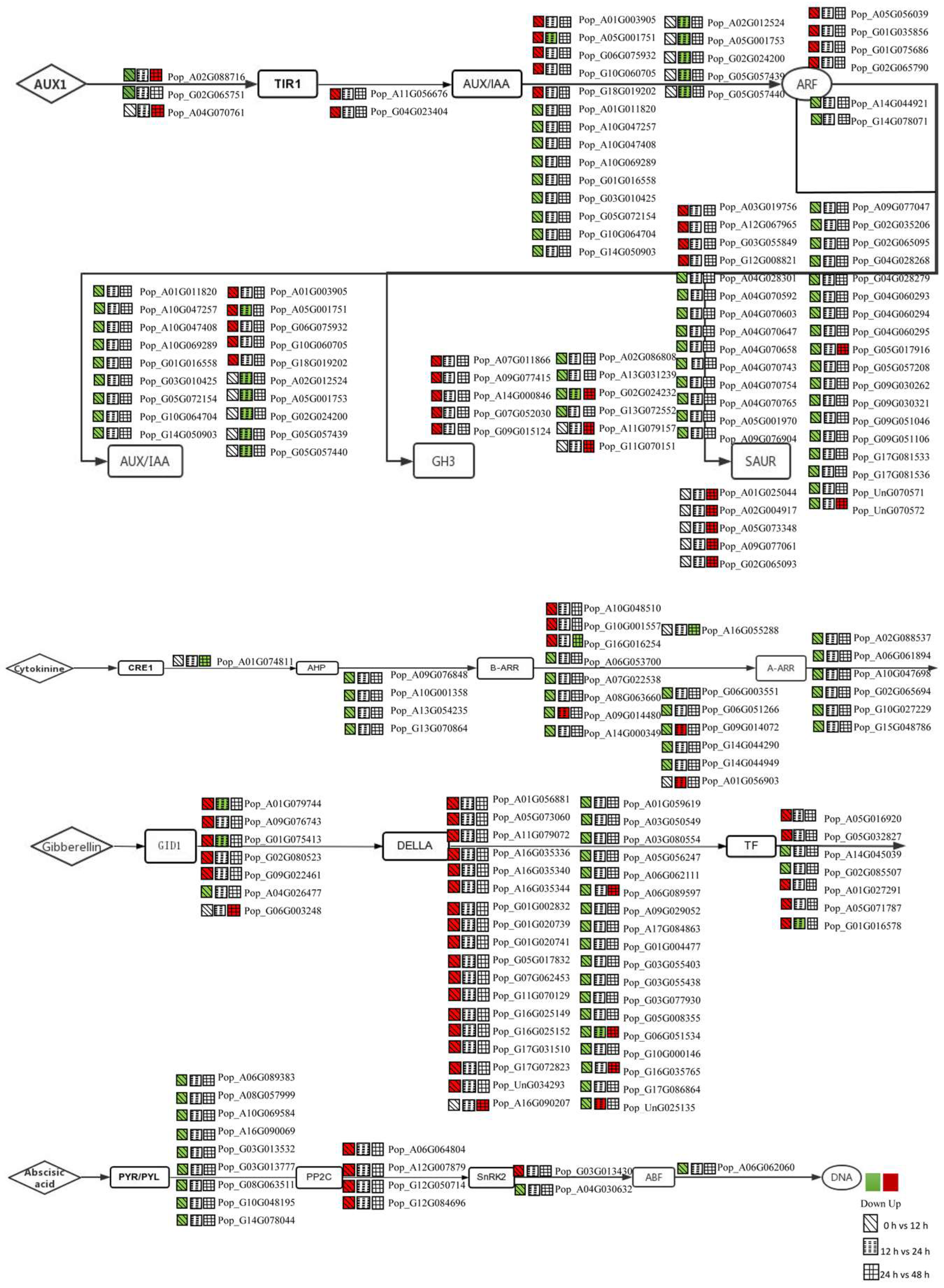
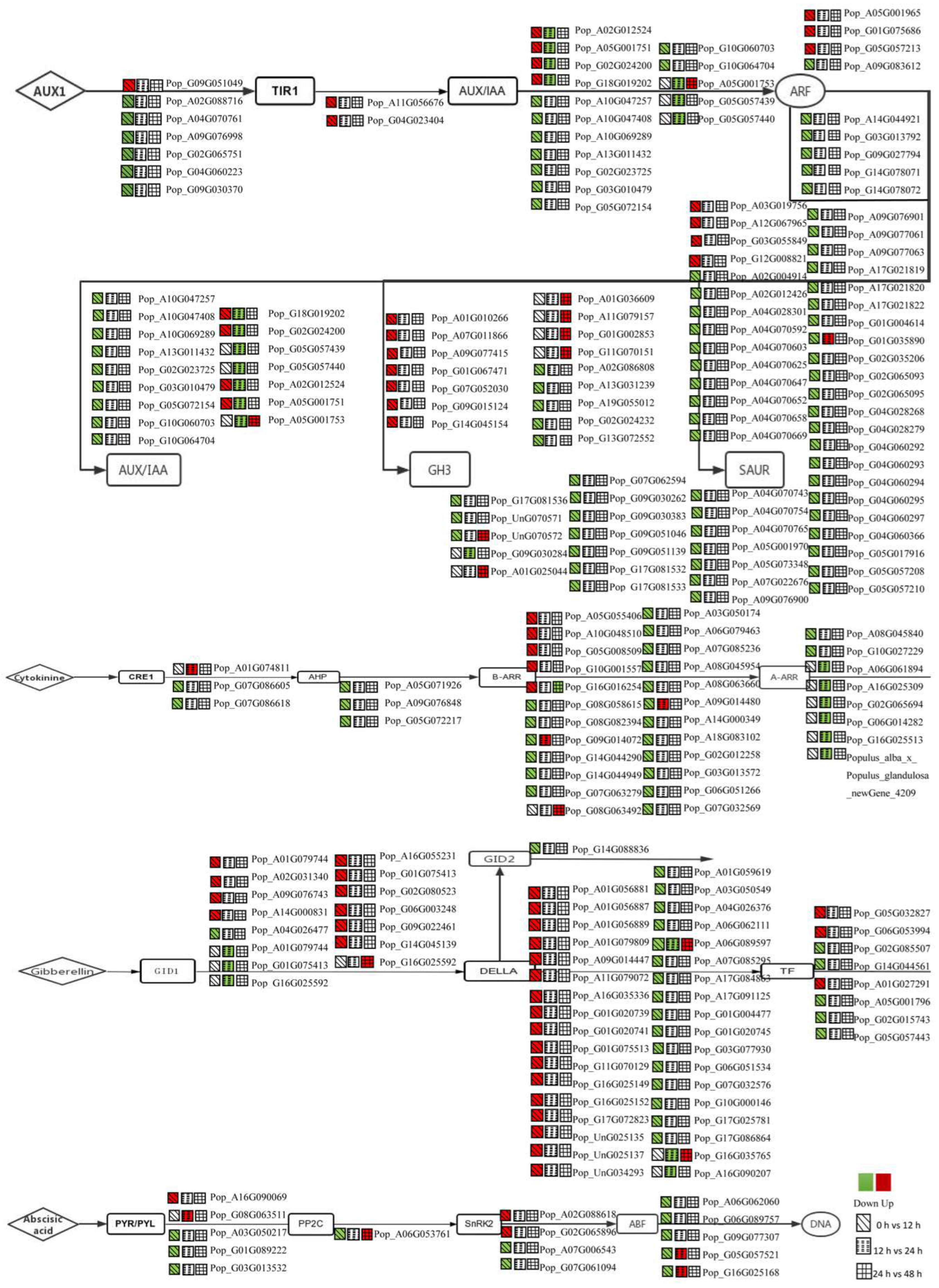
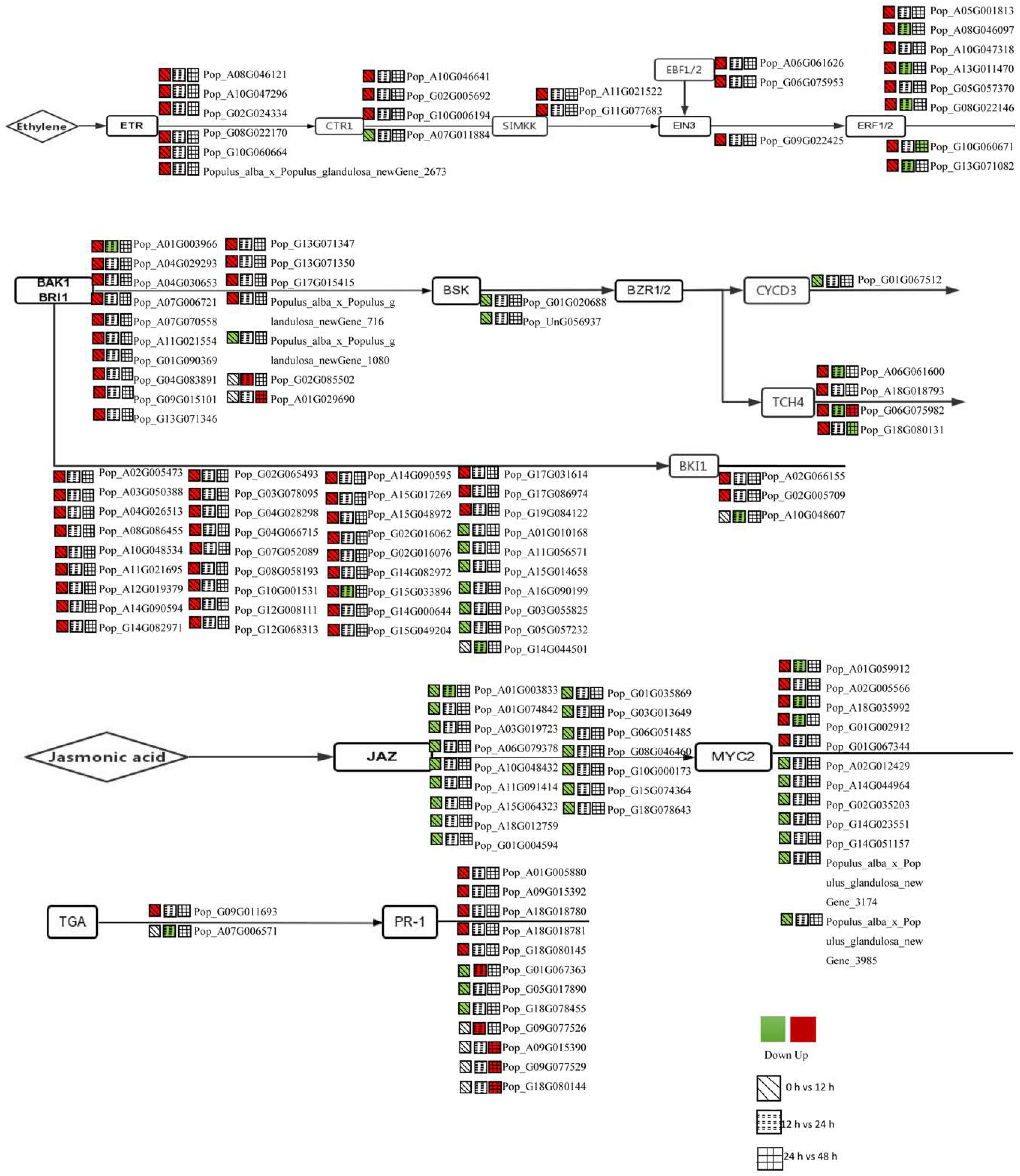
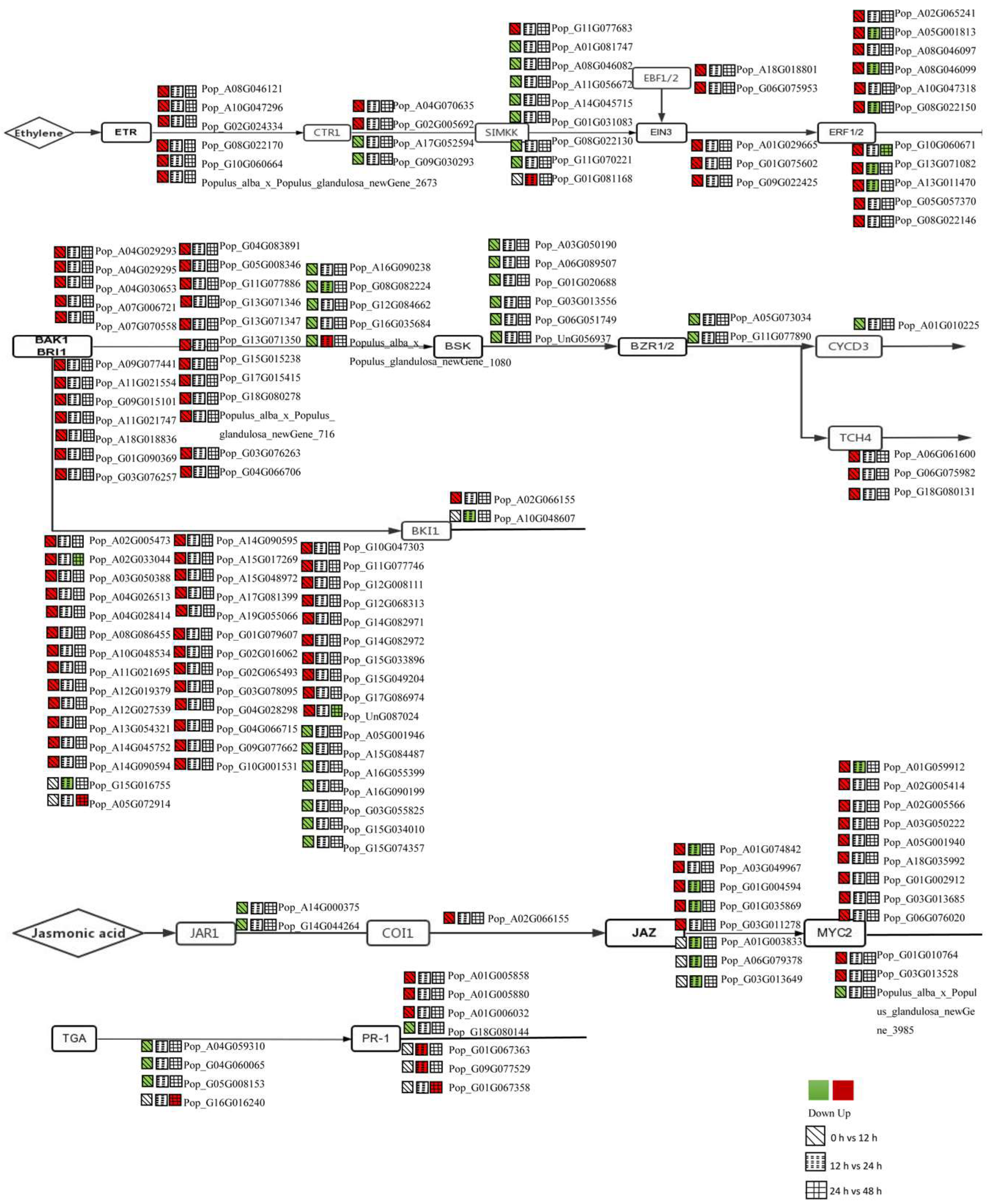
3.3. DEGs in the Plant Hormone Signalling Pathway Were Regulated during AR Formation
3.4. Expression Analysis of Auxin-Related Genes for the First 12 h
3.5. Expression Analysis of CTK-, GA- and ABA-Related Genes for the First 12 h
3.6. Expression Analysis of BR-, JA-, SA- and ETH-Related Genes for the First 12 h
3.7. Validation of Gene Expression by qRT-PCR
4. Discussion
4.1. Regulation of AR Formation by Hormone Signalling
4.2. Regulation of AR Formation by Auxin Signalling for the First 12 h
4.3. Regulation of AR Formation by JA, ETH, CTK and GA Signalling for the First 12 h
4.4. Regulation of AR Formation by BR, ABA and SA Signalling for the First 12 h
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shu, W.; Zhou, H.; Jiang, C.; Zhao, S.; Wang, L.; Li, Q.; Yang, Z.; Groover, A.; Lu, M.-Z. The auxin receptor TIR1 homolog (PagFBL1) regulates adventitious rooting through interactions with Aux/IAA28 in Populus. Plant Biotechnol. J. 2018, 17, 338–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakehal, A.; Bellini, C. Control of adventitious root formation: Insights into synergistic and antagonistic hormonal interactions. Physiol. Plant. 2018, 165, 90–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legué, V.; Rigal, A.; Bhalerao, R.P. Adventitious root formation in tree species: Involvement of transcription factors. Physiol. Plant. 2014, 151, 192–198. [Google Scholar] [CrossRef] [Green Version]
- Bai, Z.; Zhang, J.; Ning, X.; Gou, H.; Xu, X.; Huang, X.; Wang, Y.; Hu, Z.; Lu, C.; Zhang, L.; et al. A Ki-nase-Phosphatase-Transcription Factor Module Regulates Adventitious Root Emergence in Arabidopsis Root-Hypocotyl Junc-tions. Mol. Plant 2020, 13, 1162–1177. [Google Scholar] [CrossRef]
- Sengupta, S.; Chaudhuri, R.N. ABI3 plays a role in de-novo root regeneration from Arabidopsis thaliana callus cells. Plant Signal. Behav. 2020, 15, 1794147. [Google Scholar] [CrossRef]
- Joshi, M.; Ginberg, I. Adventitious root formation in crops-Potato as an example. Physiol. Plant. 2021, 172, 124–133. [Google Scholar] [CrossRef]
- Druege, U.; Hilo, A.; Pérez-Pérez, J.M.; Klopotek, Y.; Acosta, M.; Shahinnia, F.; Zerche, S.; Franken, P.; Hajirezaei, M.R. Mo-lecular and physiological control of adventitious rooting in cuttings: Phytohormone action meets resource allocation. Ann. Bot. 2019, 123, 929–949. [Google Scholar] [CrossRef] [Green Version]
- Koike, I.; Watanabe, S.; Okazaki, K.; Hayashi, K.-I.; Kasahara, H.; Shimomura, K.; Umehara, M. Endogenous auxin determines the pattern of adventitious shoot formation on internodal segments of ipecac. Planta 2020, 251, 73. [Google Scholar] [CrossRef]
- Lakehal, A.; Dob, A.; Novák, O.; Bellini, C. A DAO1-Mediated Circuit Controls Auxin and Jasmonate Crosstalk Robustness during Adventitious Root Initiation in Arabidopsis. Int. J. Mol. Sci. 2019, 20, 4428. [Google Scholar] [CrossRef] [Green Version]
- Shu, W.; Liu, Y.; Guo, Y.; Zhou, H.; Zhang, J.; Zhao, S.; Lu, M. A Populus TIR1 gene family survey reveals differential expression patterns and responses to 1-naphthaleneacetic acid and stress treatments. Front. Plant Sci. 2015, 6, 719. [Google Scholar] [CrossRef] [Green Version]
- Blázquez, M.A.; Nelson, D.C.; Weijers, D. Evolution of Plant Hormone Response Pathways. Annu. Rev. Plant Biol. 2020, 71, 327–353. [Google Scholar] [CrossRef] [Green Version]
- Rozhon, W.; Akter, S.; Fernandez, A.; Poppenberger, B. Inhibitors of Brassinosteroid Biosynthesis and Signal Transduction. Molecules 2019, 24, 4372. [Google Scholar] [CrossRef] [Green Version]
- Aoyama, T.; Oka, A. Cytokinin signal transduction in plant cells. J. Plant Res. 2003, 116, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-F.; Etheridge, N.; Schaller, G.E. Ethylene Signal Transduction. Ann. Bot. 2005, 95, 901–915. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Zhou, Y.; Zhou, M.; Yan, J.; Khurshid, M.; Weng, W.; Cheng, J.; Zhang, K. Jasmonic Acid Signaling Pathway in Plants. Int. J. Mol. Sci. 2019, 20, 2479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, L.J. Signaling Crosstalk between Salicylic Acid and Ethylene/Jasmonate in Plant Defense: Do We Understand What They Are Whispering? Int. J. Mol. Sci. 2019, 20, 671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.; Pardo, J.M.; Yun, D.-J. Desensitization of ABA-Signaling: The Swing From Activation to Degradation. Front. Plant Sci. 2020, 11, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, D.; Bai, S.; Ma, J.; Zhang, L.; Shao, F.; Zhang, K.; Yang, Y.; Sun, T.; Huang, J.; Zhou, Y.; et al. The genome of Populus alba x Populus tremula var. glandulosa clone 84K. DNA Res. 2019, 26, 423–431. [Google Scholar] [CrossRef]
- Liu, B.; Wang, L.; Zhang, J.; Li, J.; Zheng, H.; Chen, J.; Lu, M. WUSCHEL-related Homeobox genes in Populus tomentosa: Di-versified expression patterns and a functional similarity in adventitious root formation. BMC Genom. 2014, 15, 296. [Google Scholar] [CrossRef] [Green Version]
- Tahir, M.M.; Chen, S.; Ma, X.; Li, S.; Zhang, X.; Shao, Y.; Shalmani, A.; Zhao, C.; Bao, L.; Zhang, D. Transcriptome analysis reveals the promotive effect of potassium by hormones and sugar signaling pathways during adventitious roots formation in the apple rootstock. Plant Physiol. Biochem. 2021, 165, 123–136. [Google Scholar] [CrossRef]
- Steffens, B.; Rasmussen, A. The Physiology of Adventitious Roots. Plant Physiol. 2015, 170, 603–617. [Google Scholar] [CrossRef] [Green Version]
- Devi, J.; Kaur, E.; Swarnkar, M.K.; Acharya, V.; Bhushan, S. De novo transcriptome analysis provides insights into formation of in vitro adventitious root from leaf explants of Arnebia euchroma. BMC Plant Biol. 2021, 21, 414. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, Z.; Zhan, C.; Liu, M.; Xia, W.; Wang, N. Comprehensive analysis of dynamic gene expression and investigation of the roles of hydrogen peroxide during adventitious rooting in poplar. BMC Plant Biol. 2019, 19, 99. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, Y.; Hirao, T.; Mishima, K.; Ohira, M.; Hiraoka, Y.; Takahashi, M.; Watanabe, A. Transcriptome dynamics of rooting zone and aboveground parts of cuttings during adventitious root formation in Cryptomeria japonica D. Don. BMC Plant Biol. 2018, 18, 201. [Google Scholar] [CrossRef]
- Díaz-Sala, C. A Perspective on Adventitious Root Formation in Tree Species. Plants 2020, 9, 1789. [Google Scholar] [CrossRef] [PubMed]
- Bannoud, F.; Bellini, C. Adventitious Rooting in Populus Species: Update and Perspectives. Front. Plant Sci. 2021, 12, 668837. [Google Scholar] [CrossRef]
- Rigal, A.; Yordanov, Y.S.; Perrone, I.; Karlberg, A.; Tisserant, E.; Bellini, C.; Busov, V.B.; Martin, F.; Kohler, A.; Bhalerao, R.; et al. The AINTEGUMENTA LIKE1 Homeotic Transcription Factor PtAIL1 Controls the Formation of Adventitious Root Primordia in Poplar. Plant Physiol. 2012, 160, 1996–2006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez-Carvajal, G.A.; Morse, A.M.; Dervinis, C.; Davis, J.M. The Cytokinin Type-B Response Regulator PtRR13 Is a Negative Regulator of Adventitious Root Development in Populus. Plant Physiol. 2009, 150, 759–771. [Google Scholar] [CrossRef] [Green Version]
- Wei, M.; Liu, Q.; Wang, Z.; Yang, J.; Li, W.; Chen, Y.; Lu, H.; Nie, J.; Liu, B.; Lv, K.; et al. PuHox52-mediated hierarchical mul-tilayered gene regulatory network promotes adventitious root formation in Populus ussuriensis. New Phytol. 2020, 228, 1369–1385. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chen, Y.; Jiang, C.; Lu, M.-Z.; Zhang, J. Exogenous hormones supplementation improve adventitious root formation in woody plants. Front. Bioeng. Biotechnol. 2022, 10, 1009531. [Google Scholar] [CrossRef]
- Bellini, C.; Pacurar, D.I.; Perrone, I. Adventitious Roots and Lateral Roots: Similarities and Differences. Annu. Rev. Plant Biol. 2014, 65, 639–666. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Meng, N.; Chen, S.; Zhang, H.; Liu, Z.; Wang, Y.; Jing, Y.; Wang, Y.; Chen, S. Transcriptomic profiles of poplar (Populus simonii × P. nigra) cuttings during adventitious root formation. Front. Genet. 2022, 13, 968544. [Google Scholar] [CrossRef] [PubMed]
- Cai, K.; Zhang, D.; Li, X.; Zhang, Q.; Jiang, L.; Li, Y.; Song, R.; Sun, S.; Guo, R.; Han, R.; et al. Exogenous phytohormone ap-plication and transcriptome analysis provides insights for adventitious root formation in Taxus cuspidata S. et Z. Plant Growth Regul. 2022, 100, 33–53. [Google Scholar] [CrossRef]
- Xiong, Y.; Chen, S.; Wei, Z.; Chen, X.; Guo, B.; Zhang, T.; Yin, Y.; Yu, X.; Pang, J.; Niu, M.; et al. Transcriptomic analyses provide insight into adventitious root formation of Euryodendron excelsum H. T. Chang during ex vitro rooting. Plant Cell Tissue Organ. Cult. 2022, 148, 649–666. [Google Scholar] [CrossRef]
- Ranjan, A.; Perrone, I.; Alallaq, S.; Singh, R.; Rigal, A.; Brunoni, F.; Chitarra, W.; Guinet, F.; Kohler, A.; Martin, F.; et al. Mo-lecular basis of differential adventitious rooting competence in poplar genotypes. J. Exp. Bot. 2022, 73, 4046–4064. [Google Scholar] [CrossRef]
- Ji, X.-L.; Li, H.-L.; Qiao, Z.-W.; Zhang, J.-C.; Sun, W.-J.; You, C.-X.; Hao, Y.-J.; Wang, X.-F. The BTB protein MdBT2 recruits auxin signaling components to regulate adventitious root formation in apple. Plant Physiol. 2022, 189, 1005–1020. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-W.; Leng, Y.; Shi, R.-F. Transcriptomic profiling provides molecular insights into hydrogen peroxide-induced adventitious rooting in mung bean seedlings. BMC Genom. 2017, 18, 188. [Google Scholar] [CrossRef] [Green Version]
- de Klerk, G.-J.; van der Krieken, W.; de Jong, J.C. Review the formation of adventitious roots: New concepts, new possibilities. Vitr. Cell. Dev. Biol. Plant 1999, 35, 189–199. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Shi, M.; Tang, F.; Su, N.; Jin, F.; Pan, Y.; Chu, L.; Lu, M.; Shu, W.; Li, J. Transcriptome Analysis Reveals the Hormone Signalling Coexpression Pathways Involved in Adventitious Root Formation in Populus. Forests 2023, 14, 1436. https://doi.org/10.3390/f14071436
Zhang Q, Shi M, Tang F, Su N, Jin F, Pan Y, Chu L, Lu M, Shu W, Li J. Transcriptome Analysis Reveals the Hormone Signalling Coexpression Pathways Involved in Adventitious Root Formation in Populus. Forests. 2023; 14(7):1436. https://doi.org/10.3390/f14071436
Chicago/Turabian StyleZhang, Qiqi, Meirong Shi, Fang Tang, Nan Su, Feiyang Jin, Yining Pan, Liwei Chu, Mengzhu Lu, Wenbo Shu, and Jingcai Li. 2023. "Transcriptome Analysis Reveals the Hormone Signalling Coexpression Pathways Involved in Adventitious Root Formation in Populus" Forests 14, no. 7: 1436. https://doi.org/10.3390/f14071436