
Comparative Transcriptome Analysis Reveals Key Genes and Pathways Associated with Phosphate-Sensitive Behaviors in Cunninghamia lanceolata (Lamb.) Hook.

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Pi Treatment
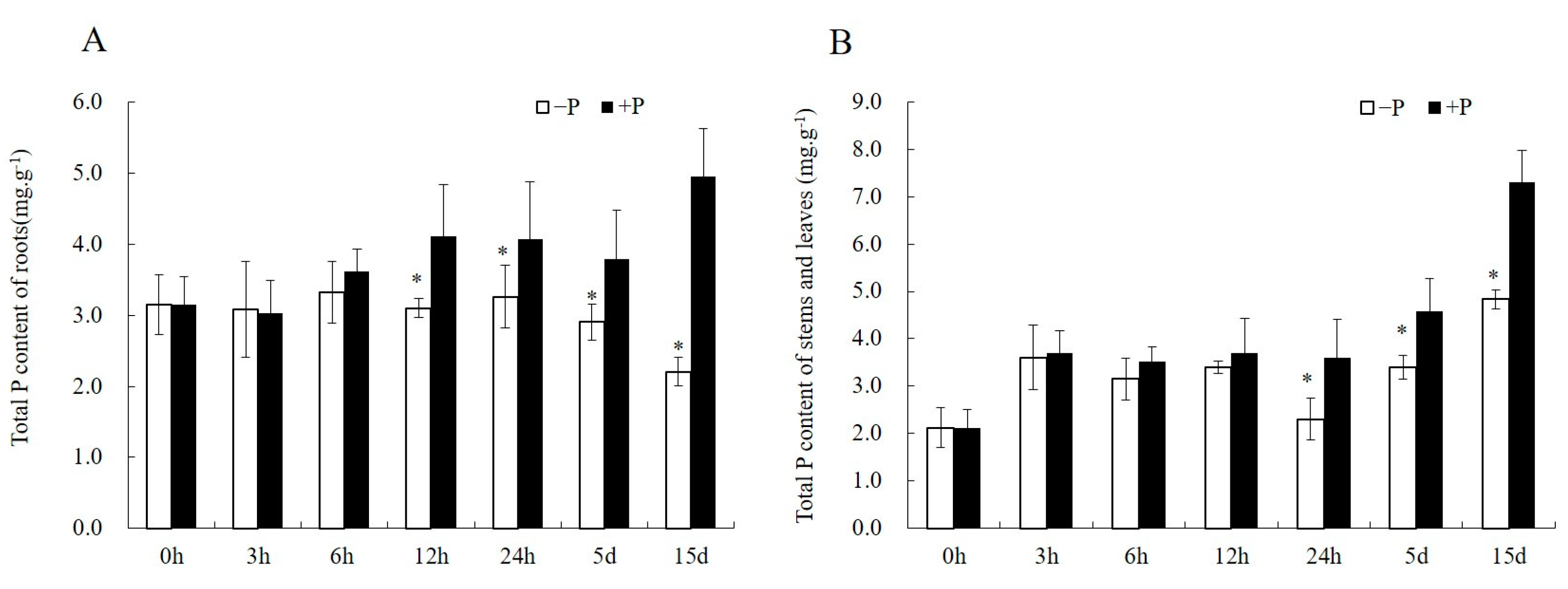
2.2. Morphology Traits and Nutrient Analyses
2.3. Total RNA Extraction, Library Construction, and RNA-Seq
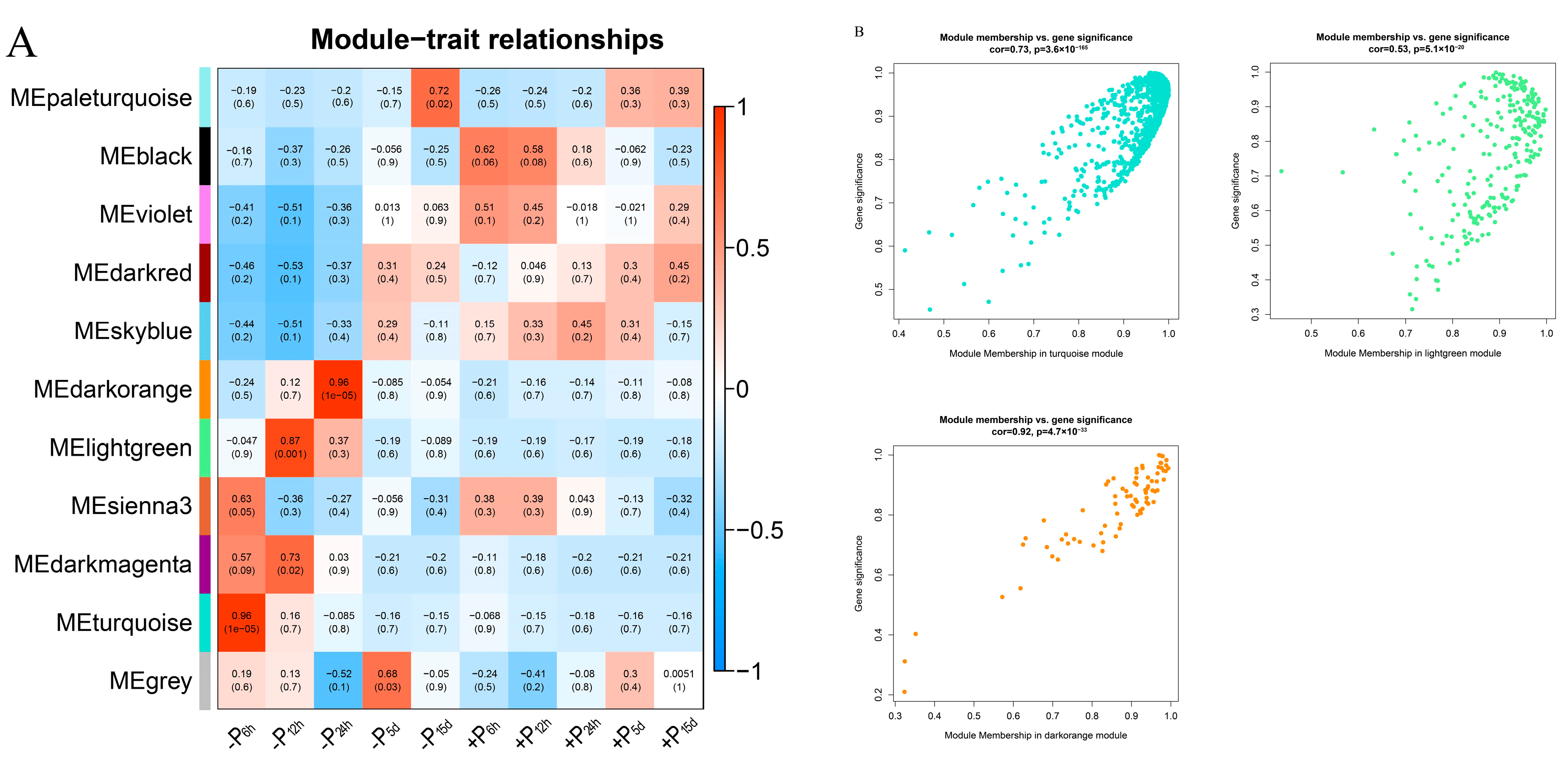
2.4. Construction of Module Genes Network and HUB Genes Identification
2.5. Quantitative Real-Time-PCR Analysis
3. Results
3.1. Evaluation of Clones with Different Phosphorus Efficiency Based on Morphological Changes during Pi-Deficiency Stress
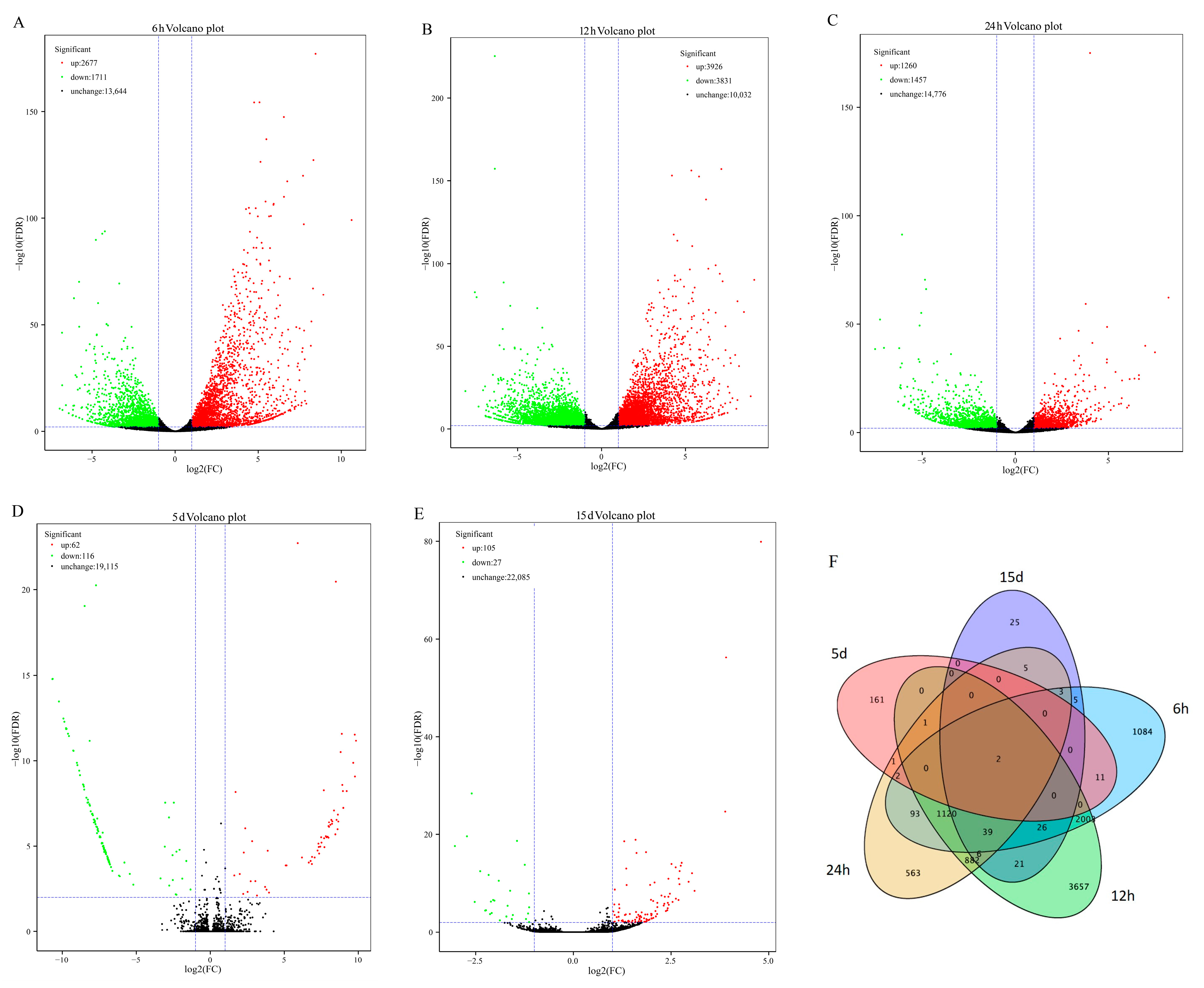
3.2. Global Profiling and Functional Annotation of the Pi-Responsive Genes in Pi-Sensitive Clones
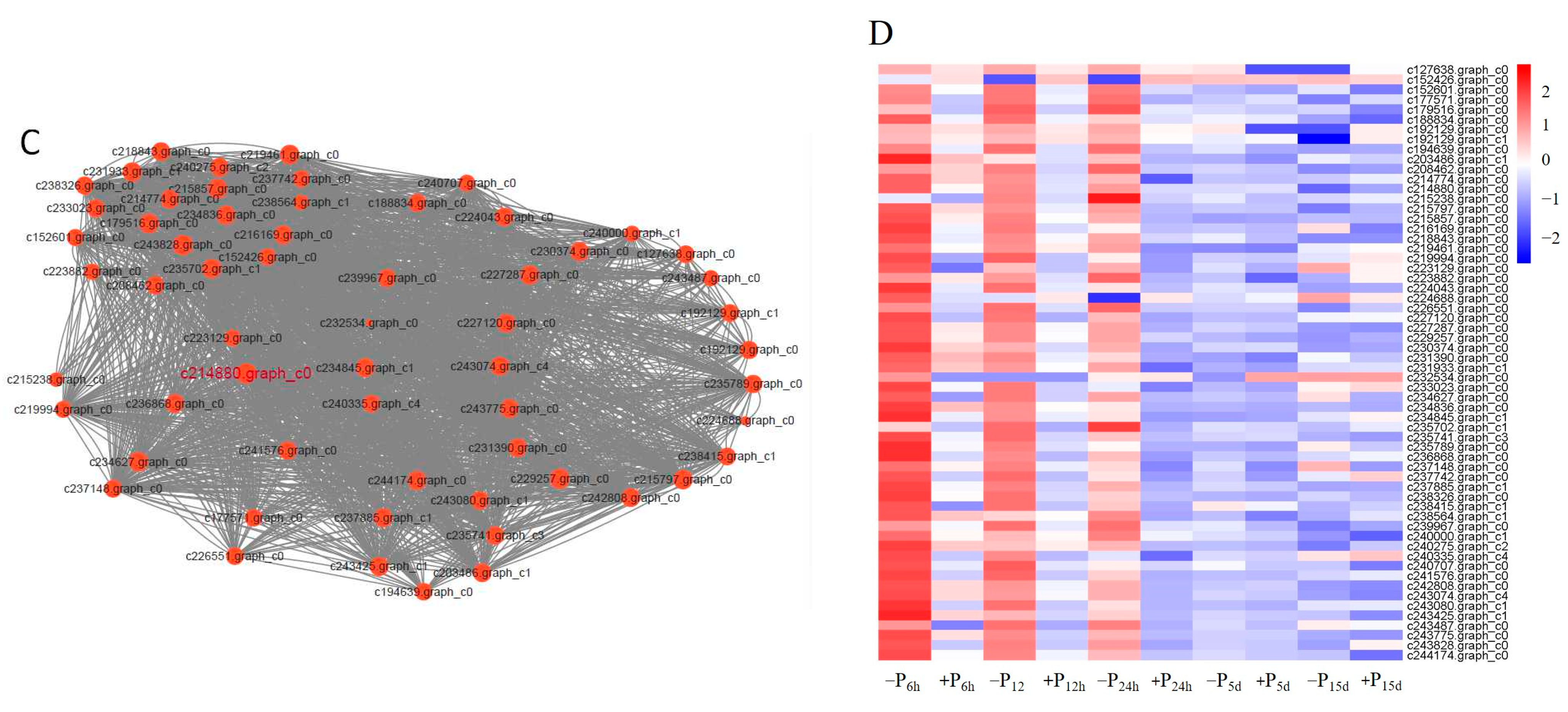
3.3. Analysis of the Gene Co-Expression Network and Selection of Hub Genes in Pi-Sensitive Clones
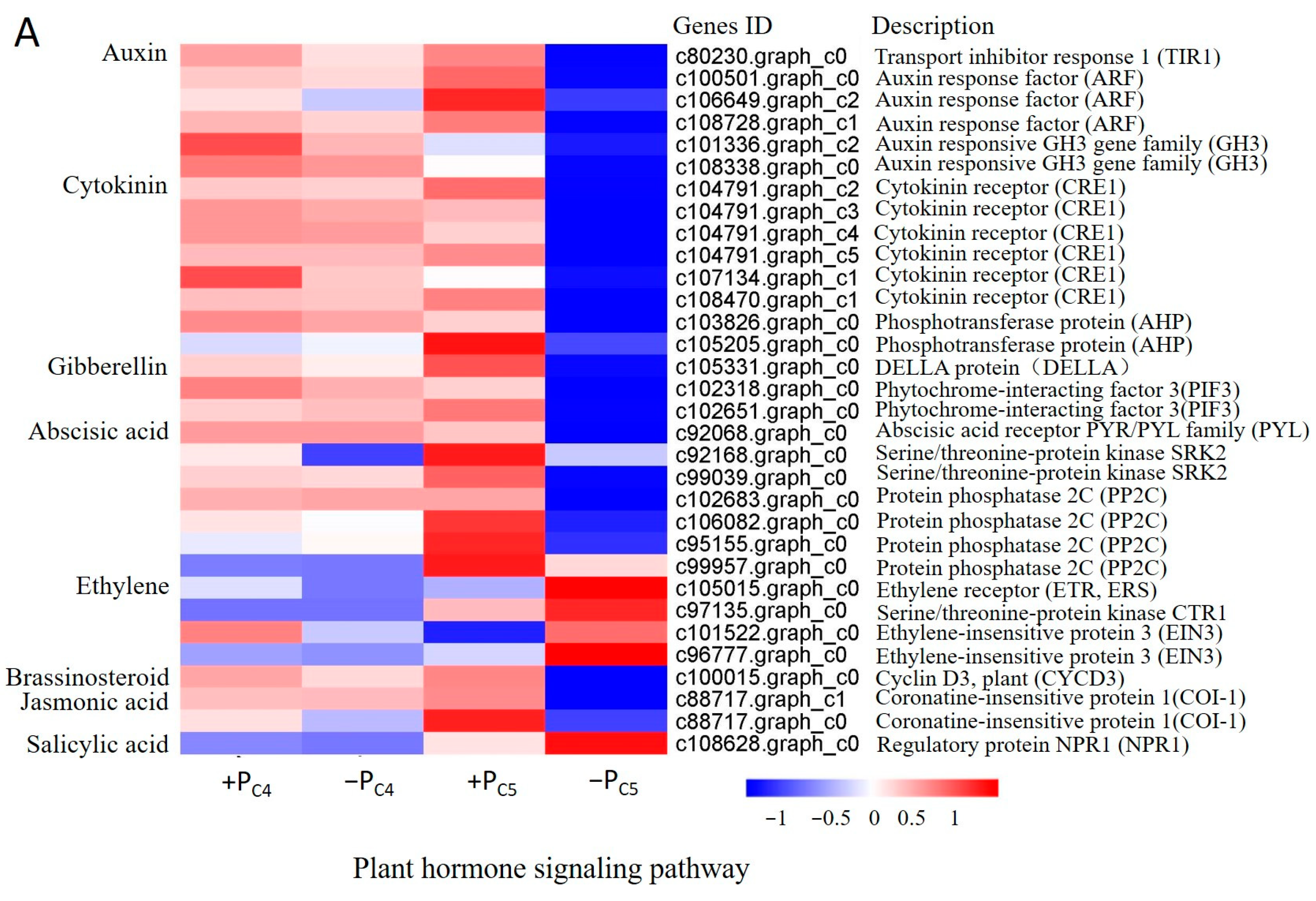
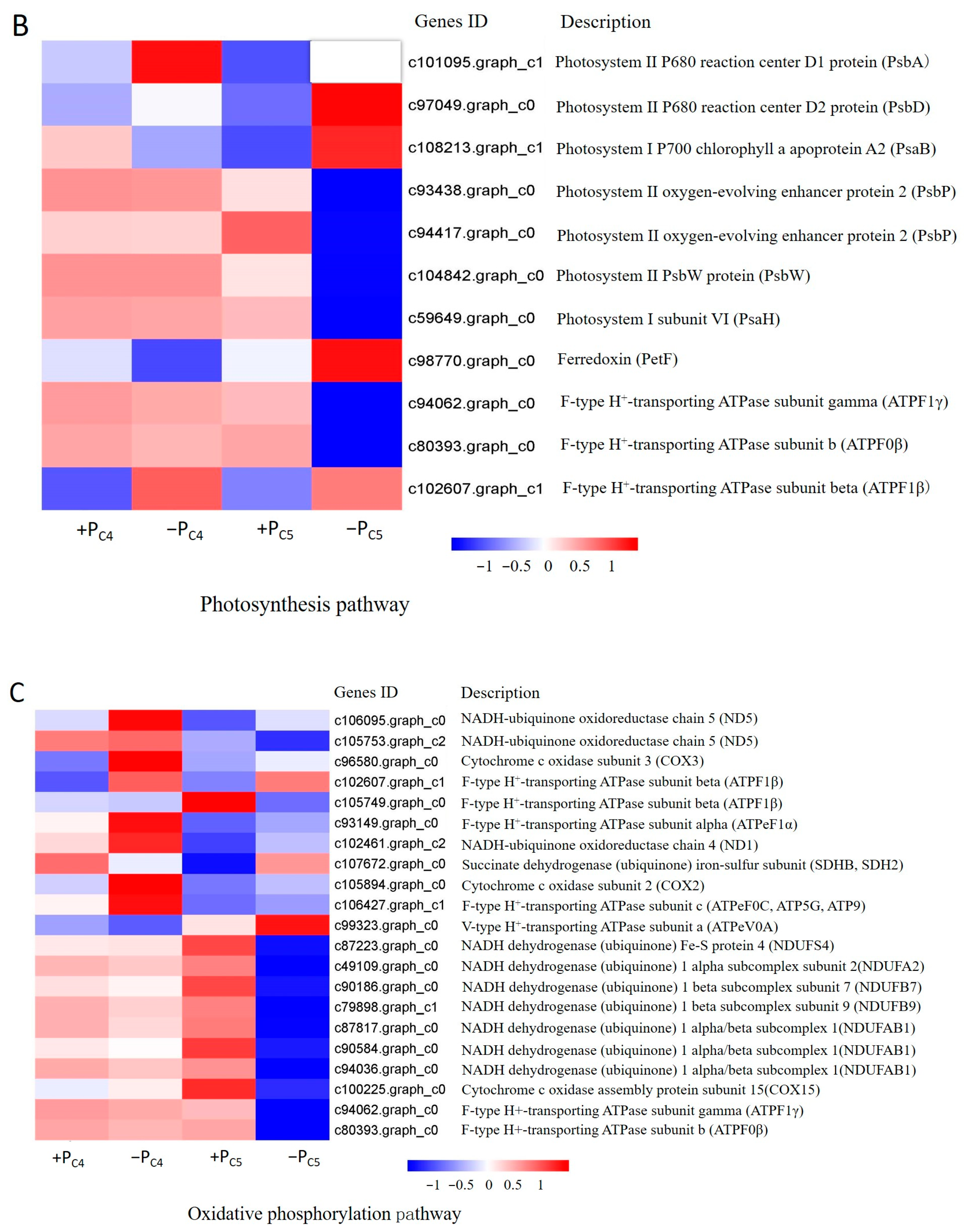
3.4. Gene-Specific Expression Analysis of Pi-Sensitive Clones
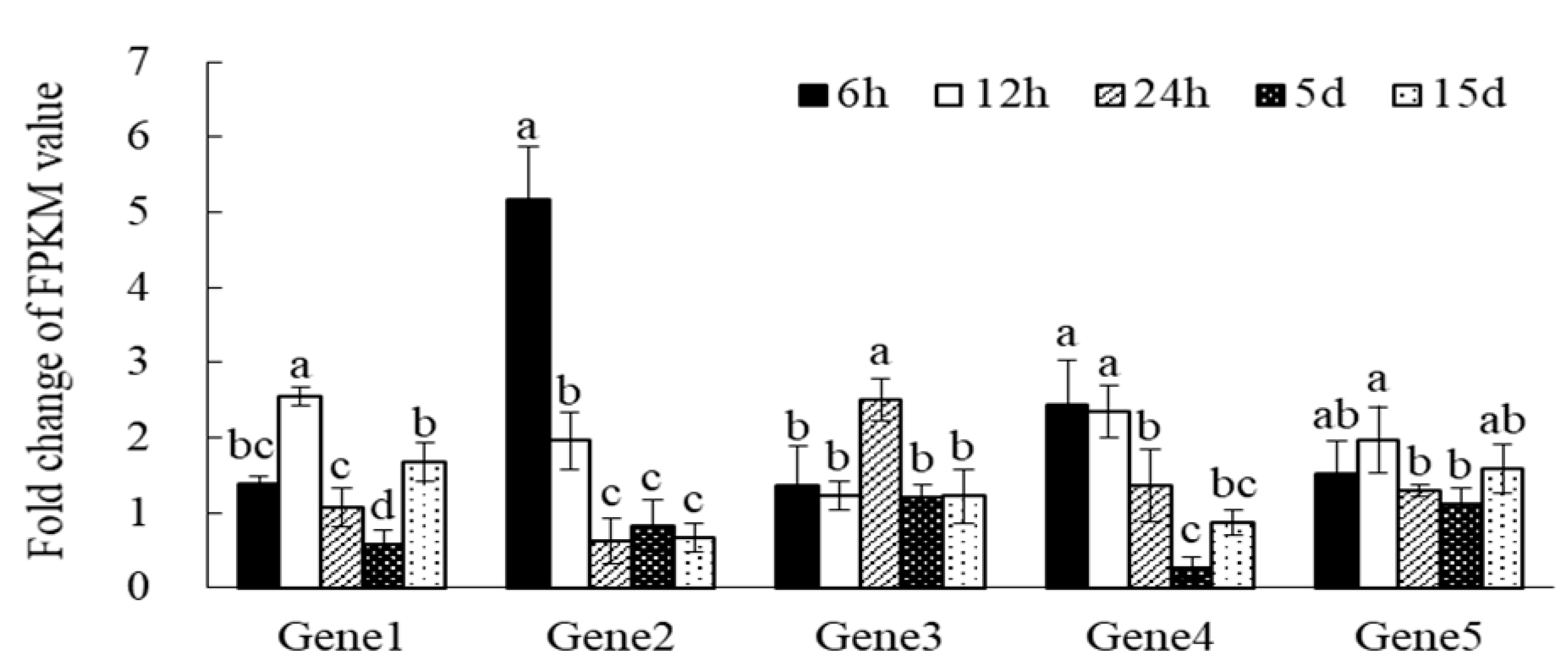
3.5. qRT-PCR Analysis of Chinese Fir Clone Genes
4. Discussion
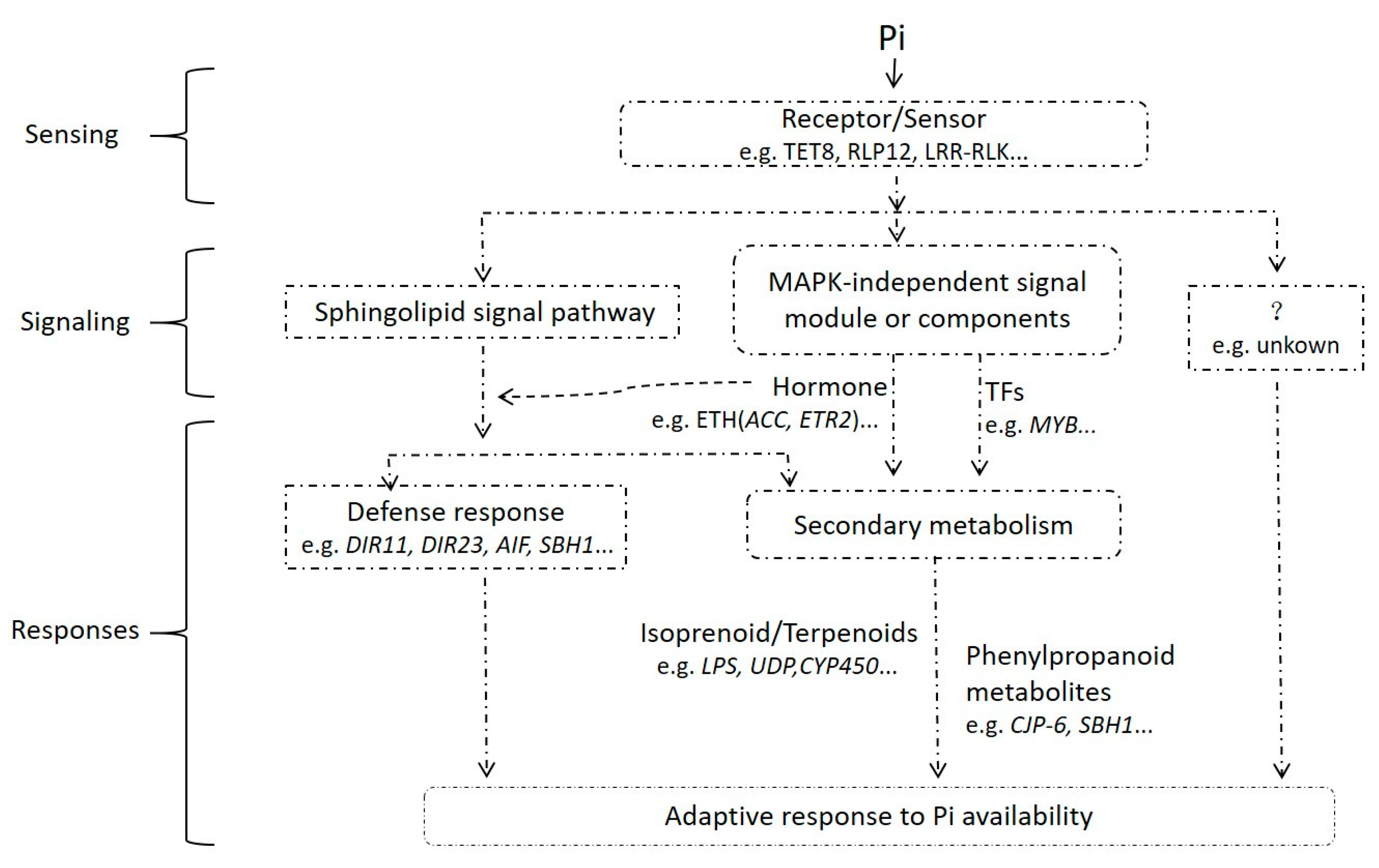
4.1. Ethylene Played an Important Role in the Early Response to Pi-Deficiency Stress of Pi-Sensitive Clone
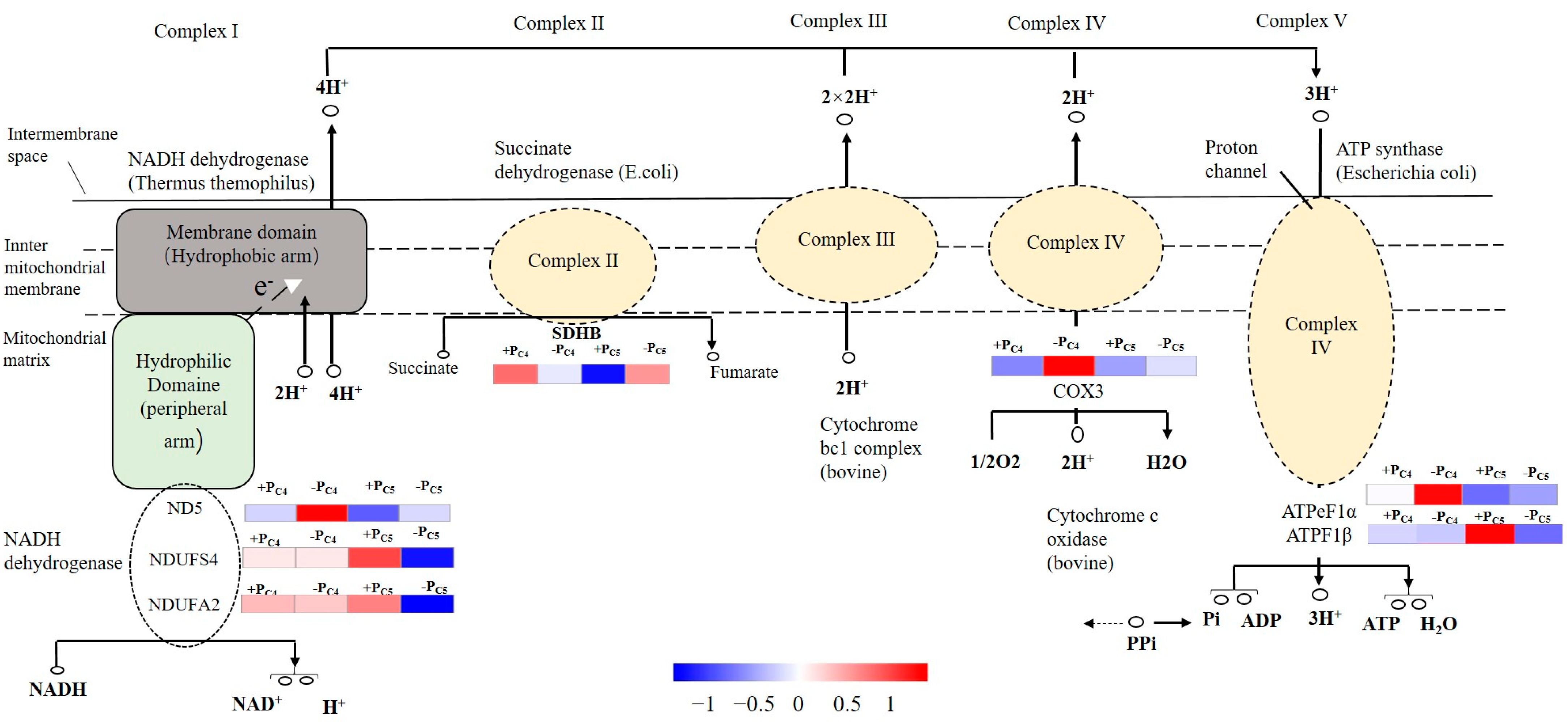
4.2. Pi-Deficiency Stress Suppressed the More Pi-Deficiency Response Genes Expression in Pi-Sensitive Clones than That in Pi-Tolerant Clones, Especially the Genes Involving in Plant Endogenous Hormone and ATP Synthase
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shen, J.; Yuan, L.; Zhang, J.; Li, H.; Bai, Z.; Chen, X.; Zhang, W.; Zhang, F. Phosphorus dynamics: From soil to plant. Plant Physiol. 2011, 156, 997–1005. [Google Scholar] [CrossRef] [Green Version]
- Zeng, H.; Wang, G.; Zhang, Y.; Hu, X.; Pi, E.; Zhu, Y.; Wang, H.; Du, L. Genome-wide identification of phosphate-deficiency-responsive genes in soybean roots by high-throughput sequencing. Plant Soil 2016, 398, 207–227. [Google Scholar] [CrossRef]
- O’Rourke, J.; Yang, S.; Miller, S.; Bucciarelli, B.; Liu, J.; Rydeen, A.; Bozsoki, Z.; Uhde-Stone, C.; Tu, Z.; Allan, D.; et al. An RNA-Seq transcriptome analysis of orthophosphate-deficient white lupin reveals novel insights into phosphorus acclimation in plants. Plant Physiol. 2013, 161, 705–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiou, T.; Lin, S. Signaling network in sensing phosphate availability in plants. Annu. Rev. Plant Biol. 2011, 62, 185–206. [Google Scholar] [CrossRef] [Green Version]
- Plaxton, W.; Tran, H. Metabolic adaptations of phosphate-starved plants. Plant Physiol. 2011, 156, 1006–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, A.E.; Lynch, J.P.; Ryan, P.R.; Delhaize, E.; Smith, F.A.; Smith, S.E.; Harvey, P.R.; Ryan, M.H.; Veneklaas, E.J.; Lambers, H.; et al. Plant and microbial strategies to improve the phosphorus efficiency of agriculture. Plant Soil 2011, 349, 121–156. [Google Scholar] [CrossRef]
- Cordell, D.; Drangert, J.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Cheraghi, M.; Lorestani, B.; Merrikhpour, H. Investigation of the effects of phosphate fertilizer application on the heavy metal content in agricultural soils with different cultivation patterns. Biol. Trace Elem. Res. 2012, 145, 87–92. [Google Scholar] [CrossRef]
- Chen, W.; Chen, R.; Zhang, Y.; Li, J.; Tigabu, M.; Ma, X.; Li, M. Cloning, Characterization and expression analysis of the phosphate starvation response gene, ClPHR1, from Chinese Fir. Forests 2020, 11, 104. [Google Scholar] [CrossRef] [Green Version]
- Hao, L.; Zhang, J.; Christie, P.; Li, X. Response of two maize inbred lines with contrasting phosphorus efficiency and root morphology to mycorrhizal colonization at different soil phosphorus supply levels. J. Plant Nutr. 2008, 31, 1059–1073. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Wang, J.; Zhu, M.; Jiang, J. Molecular mechanism of carbohydrate metabolism participation in tomato response to low phosphorus stress. Mol. Plant Breed. 2015, 13, 2833–2842. [Google Scholar]
- Zeng, J.; Liu, J.; Liang, L.; Xu, A.; Guo, X.; Zhang, L.; Zhang, W.; Hu, D. Effects of scion variety on the phosphorus efficiency of grafted Camellia oleifera seedlings. Forests 2022, 13, 203. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, Z.; Yang, Q. Genetic variations in root morphology and phosphorus efficiency of Pinus massoniana under heterogeneous and homogeneous low phosphorus conditions. Plant Soil 2013, 364, 93–104. [Google Scholar] [CrossRef]
- Chen, Z.; Wu, P.; Zou, X.; Wang, P.; Ma, J.; Ma, X. Relationship between growth and endogenous hormones of Chinese Fir seedlings under low phosphorus stress. Sci. Silvae Sin. 2016, 52, 57–67. [Google Scholar]
- Zou, X.; Wu, P.; Chen, N.; Wang, P.; Ma, X. Chinese fir root response to spatial and temporal heterogeneity of phosphorus availability in the soil. Can. J. For. Res. 2015, 45, 402–410. [Google Scholar] [CrossRef]
- Wu, P.; Ma, X.; Tigabu, M.; Wang, C.; Liu, A.; Oden, P. Root morphological plasticity and biomass production of two Chinese fir clones with high phosphorus efficiency under low phosphorus stress. Can. J. For. Res. 2011, 41, 228–234. [Google Scholar] [CrossRef]
- Chen, W.; Zhou, M.; Zhao, M.; Chen, R.; Tigabu, M.; Wu, P.; Li, M.; Ma, X. Transcriptome analysis provides insights into the root response of Chinese fir to phosphorus deficiency. BMC Plant Biol. 2021, 21, 525. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, D.; Arnon, D. The water-culture method for growing plants without soil. Calif. Agric. Exp. Stn. Circ. 1950, 347, 4–32. [Google Scholar]
- Bao, S.D. Soil Agro-Chemistrical Analysis, 3rd ed.; Agricultural Press of China: Beijing, China, 2000; pp. 40–54. (In Chinese) [Google Scholar]
- Yan, C.; Song, S.; Wang, W.; Wang, C.; Li, H.; Wang, F.; Li, S.; Sun, X. Screening diverse soybean genotypes for drought tolerance by membership function value based on multiple traits and drought-tolerant coefficient of yield. BMC Plant Biol. 2020, 20, 321. [Google Scholar] [CrossRef]
- Grabherr, M.; Haas, B.; Yassour, M.; Levin, J.; Thompson, D.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.; Madden, T.; Schäffer, A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Kawashima, S.; Okuda, S.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Mao, X.; Cai, T.; Olyarchuk, J.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, J.; Liu, W.; Luo, Z.; Wang, P.; Zhang, Y.; Zheng, R.; Shi, J. Transcriptome characteristics and six alternative expressed genes positively correlated with the phase transition of annual cambial activities in Chinese Fir (Cunninghamia lanceolata (Lamb.) Hook). PLoS ONE 2013, 8, e71562. [Google Scholar] [CrossRef] [Green Version]
- Shiu, S.; Bleecker, A. Expansion of the receptor-like kinase/pelle gene family and receptor-like proteins in Arabidopsis. Plant Physiol. 2003, 132, 530–543. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, S.; Ohta, M.; Usui, A.; Shinshi, H.; Ohme-Takagi, M. Arabidopsis ethylene-responsive element binding factors act as transcriptional activators or repressors of GCC box-mediated gene expression. Plant Cell 2000, 12, 393–404. [Google Scholar]
- Ouaked, F.; Rozhon, W.; Lecourieux, D.; Hirt, H. A MAPK pathway mediates ethylene signaling in plants. Embo. J. 2003, 22, 1282–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strader, L.C.; Chen, G.L.; Bartel, B. Ethylene directs auxin to control root cell expansion. Plant J. 2010, 64, 874–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Lai, H.; Zheng, S.; Li, M.; Ma, X.; Wu, P. Effects of exogenous ethylene on growth and phosphorus use efficiency of Chinese fir seedlings under phosphorus stress. Sci. Silvae Sin. 2021, 57, 43–50. [Google Scholar]
- Clay, N.; Adio, A.; Denoux, C.; Jander, G.; Ausubel, F. Glucosinolate metabolites required for an Arabidopsis innate immune response. Science 2009, 323, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Pilar López-Gresa, M.; Torres, C.; Campos, L.; Lisón, P.; Rodrigo, I.; María Bellés, J.; Conejero, V. Identification of defence metabolites in tomato plants infected by the bacterial pathogen Pseudomonas syringae. Env. Exp. Bot. 2011, 74, 216–228. [Google Scholar] [CrossRef]
- Diezel, C.; Allmann, S.; Baldwin, I. Mechanisms of optimal defense patterns in Nicotiana attenuata: Flowering attenuates herbivory-elicited ethylene and jasmonate signaling. J. Integr. Plant Biol. 2011, 53, 971–983. [Google Scholar] [CrossRef] [PubMed]
- Dubos, C.; Le Gourrierec, J.; Baudry, A.; Huep, G.; Lanet, E.; Debeaujon, I.; Routaboul, J.; Alboresi, A.; Weisshaar, B.; Lepiniec, L. MYBL2 is a new regulator of flavonoid biosynthesis in Arabidopsis thaliana. Plant J. 2008, 55, 940–953. [Google Scholar] [CrossRef] [PubMed]
- Jeong, C.; Chakrabarty, D.; Hahn, E.; Lee, H.; Paek, K. Effects of oxygen, carbon dioxide and ethylene on growth and bioactive compound production in bioreactor culture of ginseng adventitious roots. Biochem. Eng. J. 2006, 27, 252–263. [Google Scholar] [CrossRef]
- Dangl, J.; Jones, J. Plant pathogens and integrated defence responses to infection. Nature 2001, 411, 826–833. [Google Scholar] [CrossRef]
- Jeong, S.; Das, P.; Jeoung, S.; Song, J.; Lee, H.; Kim, Y.; Kim, W.; Park, Y.; Yoo, S.; Choi, S.; et al. Ethylene suppression of sugar-induced anthocyanin pigmentation in Arabidopsis. Plant Physiol. 2010, 154, 1514–1531. [Google Scholar] [CrossRef] [Green Version]
- Michaelson, L.; Napier, J.; Molino, D.; Faure, J. Plant sphingolipids: Their importance in cellular organization and adaption. Biochim. Biophys. Acta 2016, 1861, 1329–1335. [Google Scholar] [CrossRef]
- Nadira, U.; Ahmed, I.; Wu, F.; Zhang, G. The regulation of root growth in response to phosphorus deficiency mediated by phytohormones in a Tibetan wild barley accession. Acta Physiol. Plant. 2016, 38, 105. [Google Scholar] [CrossRef]
- Lei, K.; Zhou, H.; Gu, D.; An, G. The involvement of abscisic acid-insensitive mutants in low phosphate stress responses during rhizosphere acidification, anthocyanin accumulation and Pi homeostasis in Arabidopsis. Plant Sci. 2022, 322, 111358. [Google Scholar] [CrossRef] [PubMed]
- Perez-torres, C.; Lopezbucio, J.; Cruzramirez, A.; Ibarralaclette, E.; Dharmasiri, S.; Estelle, M.; Herrera-Estrella, L. Phosphate availability alters lateral root development in Arabidopsis by modulating auxin sensitivity via a mechanism involving the TIR1 auxin receptor. Plant Cell 2008, 20, 3258–3272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredeen, A.; Raab, T.; Rao, I.; Terry, N. Effects of phosphorus nutrition on photosynthesis in Glycine max (L.) Merr. Planta 1990, 181, 399–405. [Google Scholar] [CrossRef]
- Gniazdowska, A.; Rychter, A. Nitrate uptake by bean (Phaseolus vulgaris L.) roots under phosphate deficiency. Plant Soil 2000, 226, 79–85. [Google Scholar] [CrossRef]
- Xu, H.; Weng, X.; Yang, Y. Effect of phosphorus deficiency on the photosynthetic characteristics of rice plants. Russ. J. Plant Physiol. 2007, 54, 741–748. [Google Scholar] [CrossRef]
- Meng, X.; Chen, W.; Wang, Y.; Huang, Z.; Ye, X.; Chen, L.; Yang, L. Effects of phosphorus deficiency on the absorption of mineral nutrients, photosynthetic system performance and antioxidant metabolism in Citrus grandis. PLoS ONE 2021, 16, e0246944. [Google Scholar] [CrossRef]
- Li, L.; Liu, C.; Lian, X. Gene expression profiles in rice roots under low phosphorus stress. Plant Mol. Biol. 2010, 72, 423–432. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Source of Variation | ||
|---|---|---|---|
| Genotype | Pi Concentration | Genotype × Pi Concentration | |
| Plantlets height increment | 8.76 ** | 9.77 ** | 0.32 |
| Plantlets ground-diameter increment | 7.41 ** | 5.50 * | 0.11 |
| Root dry mass | 6.63 ** | 8.34 ** | 1.59 |
| Stem dry mass | 9.59 ** | 11.65 ** | 2.86 * |
| Leaf dry mass | 9.12 ** | 8.88 ** | 1.64 |
| Total dry mass | 10.19 ** | 13.82 ** | 2.08 |
| Root-shoot ratio | 6.85 ** | 1.33 | 1.55 |
| Total root length | 61.31 ** | 12.83 ** | 1.12 |
| Total root surface area | 23.85 ** | 5.24 * | 0.42 |
| Total root volume | 17.27 ** | 8.03 ** | 0.48 |
| Average root diameter | 6.71 ** | 24.66 ** | 1.41 |
| Leaf thickness | 11.24 ** | 2.68 | 3.40 * |
| Leaf length | 9.96 ** | 6.25 * | 0.87 |
| Leaf surface area | 8.84 ** | 8.26 ** | 3.01 * |
| Leaf volume | 7.94 ** | 8.31 ** | 3.20 * |
| Leaf mass per area | 5.90 ** | 8.15 ** | 0.41 |
| Root P content | 2.52 * | 10.97 ** | 1.26 |
| Total P content | 6.63 ** | 15.57 ** | 1.51 |
| Total N content | 13.54 ** | 20.20 ** | 0.85 |
| Total K content | 19.11 ** | 22.74 ** | 1.32 |
| Total Ca content | 7.63 ** | 16.11 ** | 1.79 |
| Total Mg content | 7.30 ** | 16.43 ** | 0.77 |
| Root P utilization efficiency | 5.40 ** | 17.90 ** | 4.47 ** |
| Total P utilization efficiency | 2.59 * | 4.69 * | 0.86 |
| Gene Set | Total Number of DEGs | Up-Regulated | Down-Regulated |
|---|---|---|---|
| +Pc5 _vs._ −Pc5 | 8128 | 4048 | 4080 |
| +Pc4 _vs._ −Pc4 | 48 | 22 | 26 |
| +Pc5 _vs._+Pc4 | 4981 | 2692 | 2289 |
| −Pc5 _vs._ −Pc4 | 8371 | 4114 | 4257 |
| Gene Set | KEGG Category | Pathway ID | UP | Down | p-Value |
|---|---|---|---|---|---|
| +Pc4 _vs._ −Pc4 | Oxidative phosphorylation | ko00190 | 4 | 0.00013 | |
| Photosynthesis | ko00195 | 2 | 0.00743 | ||
| Diterpenoid biosynthesis | ko00904 | 1 | 0.04714 | ||
| Zeatin biosynthesis | ko00908 | 1 | 0.01785 | ||
| +Pc5 _vs._ −Pc5 | Endocytosis | ko04144 | 39 | 3.30 × 10−5 | |
| Plant-pathogen interaction | ko04626 | 29 | 0.00024 | ||
| Glycerophospholipid metabolism | ko00564 | 21 | 0.00041 | ||
| alpha-Linolenic acid metabolism | ko00592 | 17 | 0.00043 | ||
| Terpenoid backbone biosynthesis | ko00900 | 15 | 0.00049 | ||
| Plant hormone signal transduction | ko04075 | 52 | 0.00018 | ||
| Cutin, suberine and wax biosynthesis | ko00073 | 11 | 0.00031 | ||
| Other glycan degradation | ko00511 | 10 | 0.00034 | ||
| Starch and sucrose metabolism | ko00500 | 51 | 0.00067 | ||
| Carotenoid biosynthesis | ko00906 | 14 | 0.00084 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, R.; Hu, D.; Chen, J.; Zheng, H.; Shi, J. Comparative Transcriptome Analysis Reveals Key Genes and Pathways Associated with Phosphate-Sensitive Behaviors in Cunninghamia lanceolata (Lamb.) Hook. Forests 2023, 14, 1203. https://doi.org/10.3390/f14061203
Wei R, Hu D, Chen J, Zheng H, Shi J. Comparative Transcriptome Analysis Reveals Key Genes and Pathways Associated with Phosphate-Sensitive Behaviors in Cunninghamia lanceolata (Lamb.) Hook. Forests. 2023; 14(6):1203. https://doi.org/10.3390/f14061203
Chicago/Turabian StyleWei, Ruping, Dehuo Hu, Jinhui Chen, Huiquan Zheng, and Jisen Shi. 2023. "Comparative Transcriptome Analysis Reveals Key Genes and Pathways Associated with Phosphate-Sensitive Behaviors in Cunninghamia lanceolata (Lamb.) Hook." Forests 14, no. 6: 1203. https://doi.org/10.3390/f14061203