
Effects of Microbial Communities on Elevational Gradient Adaptation Strategies of Pinus yunnanensis Franch. and Pinus densata Mast. in a Mixed Zone
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Sample Processing
2.2. Soil and Root Chemical Properties
2.3. DNA Isolation and Sequencing
2.4. Bioinformatic Analysis
2.5. Data and Statistical Analyses
3. Results
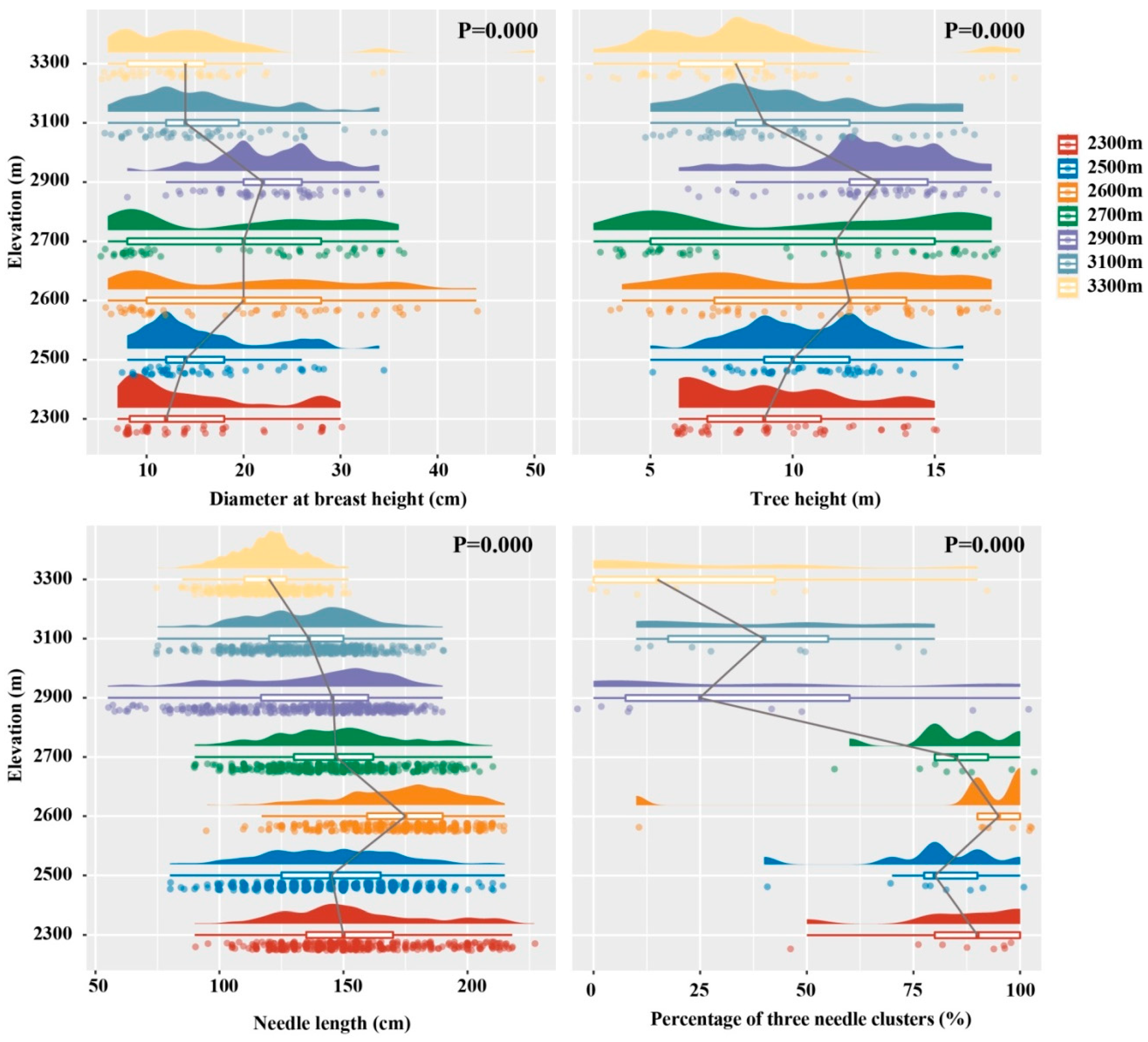
3.1. Morphological Features
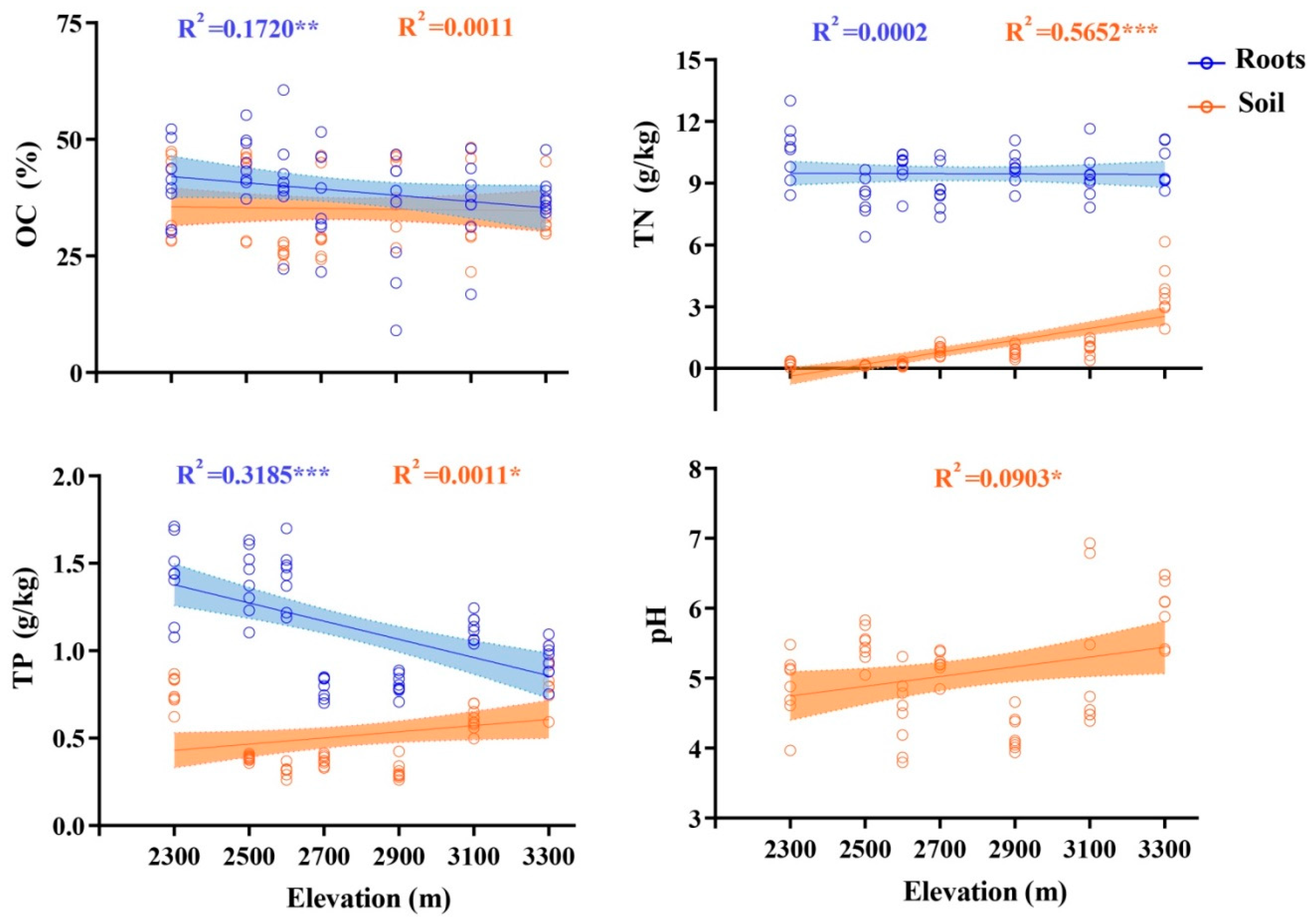
3.2. Soil Chemical Properties and Plant Nutrient Contents
3.3. Soil Chemical Properties and Plant Nutrient Contents
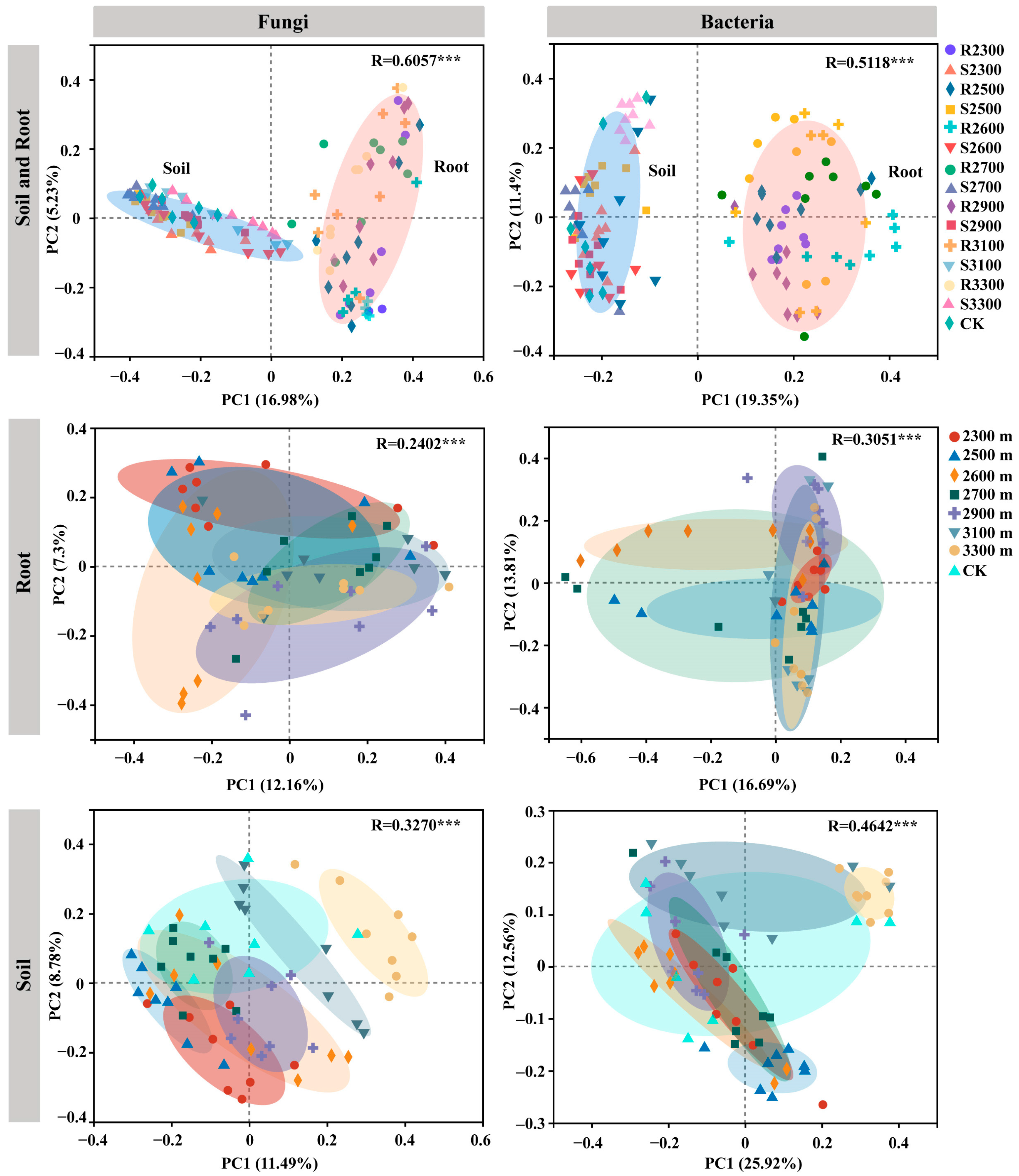
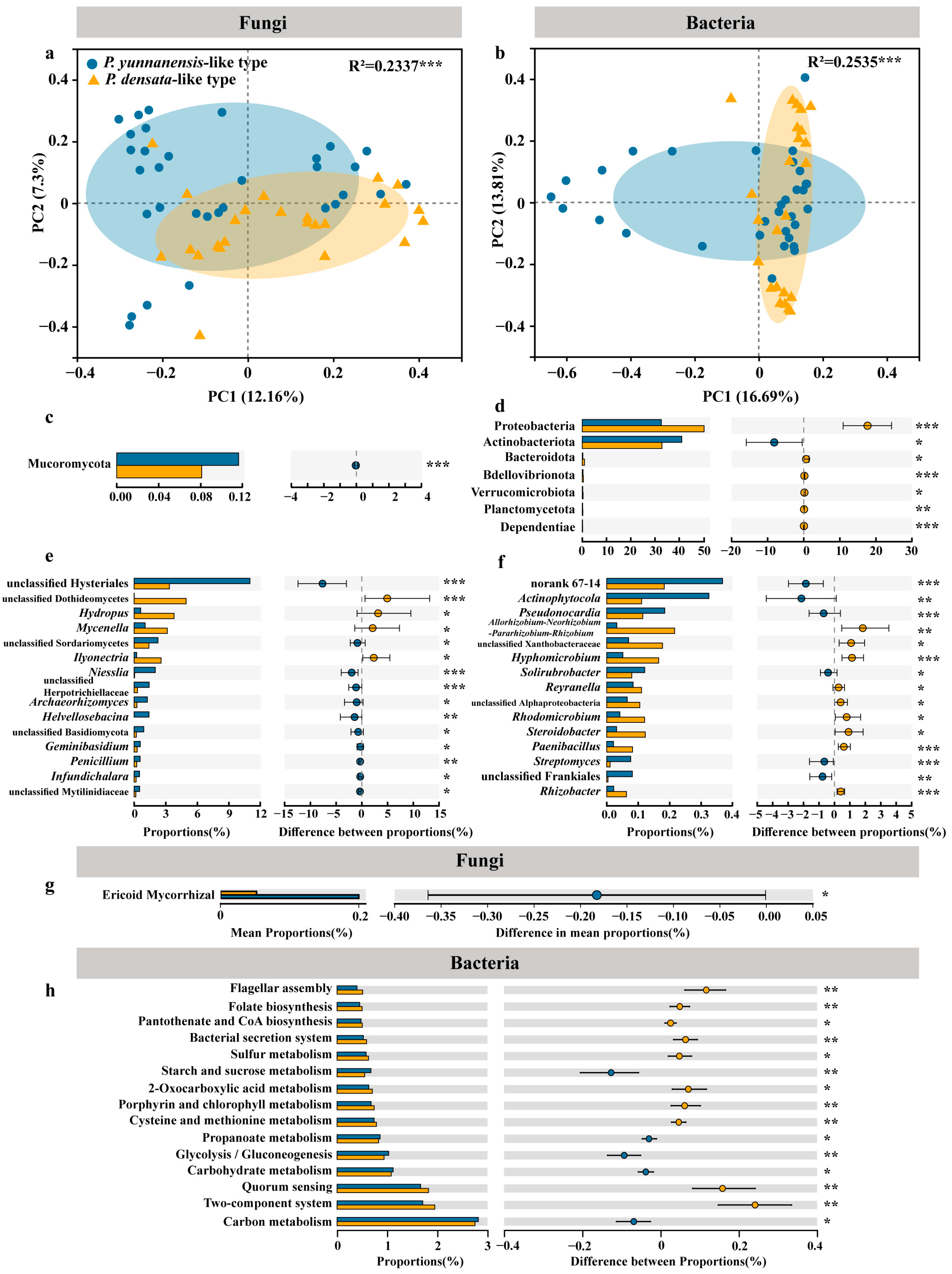
3.4. Differentiation of the Pine Root Microbiome at Different Elevations
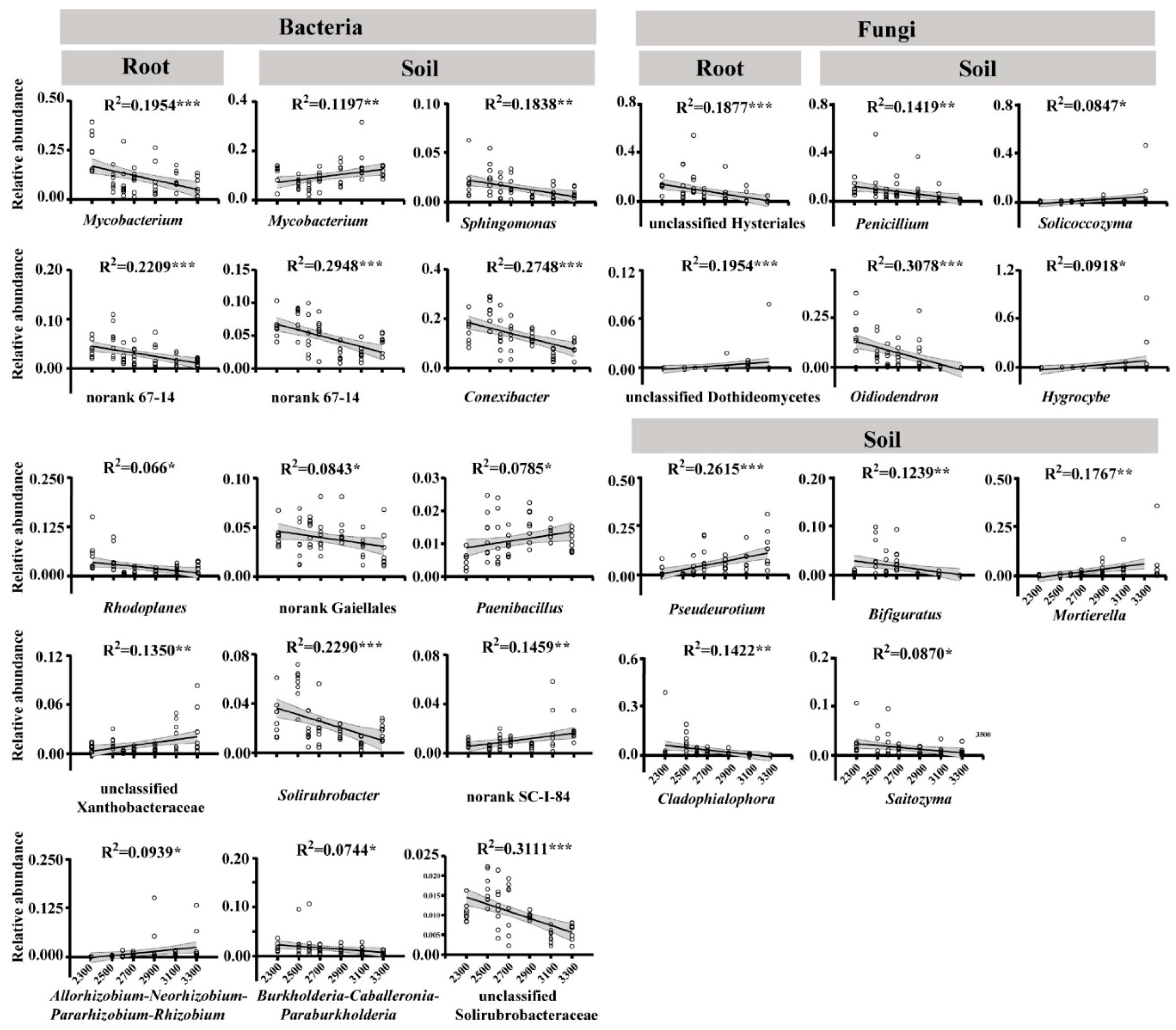
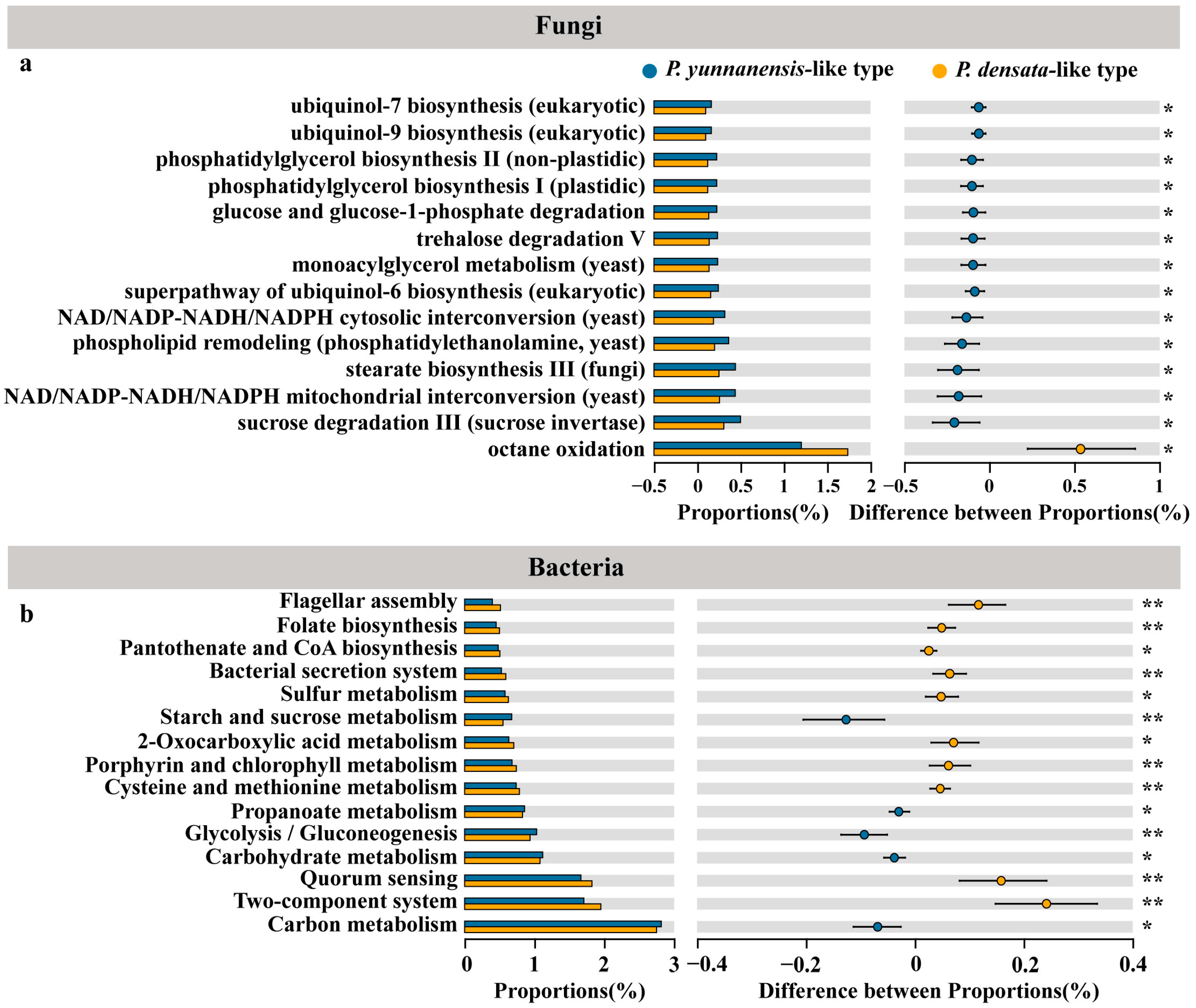
3.5. Strategies for the Differential Recruitment of Root Endophytes by Related Pines
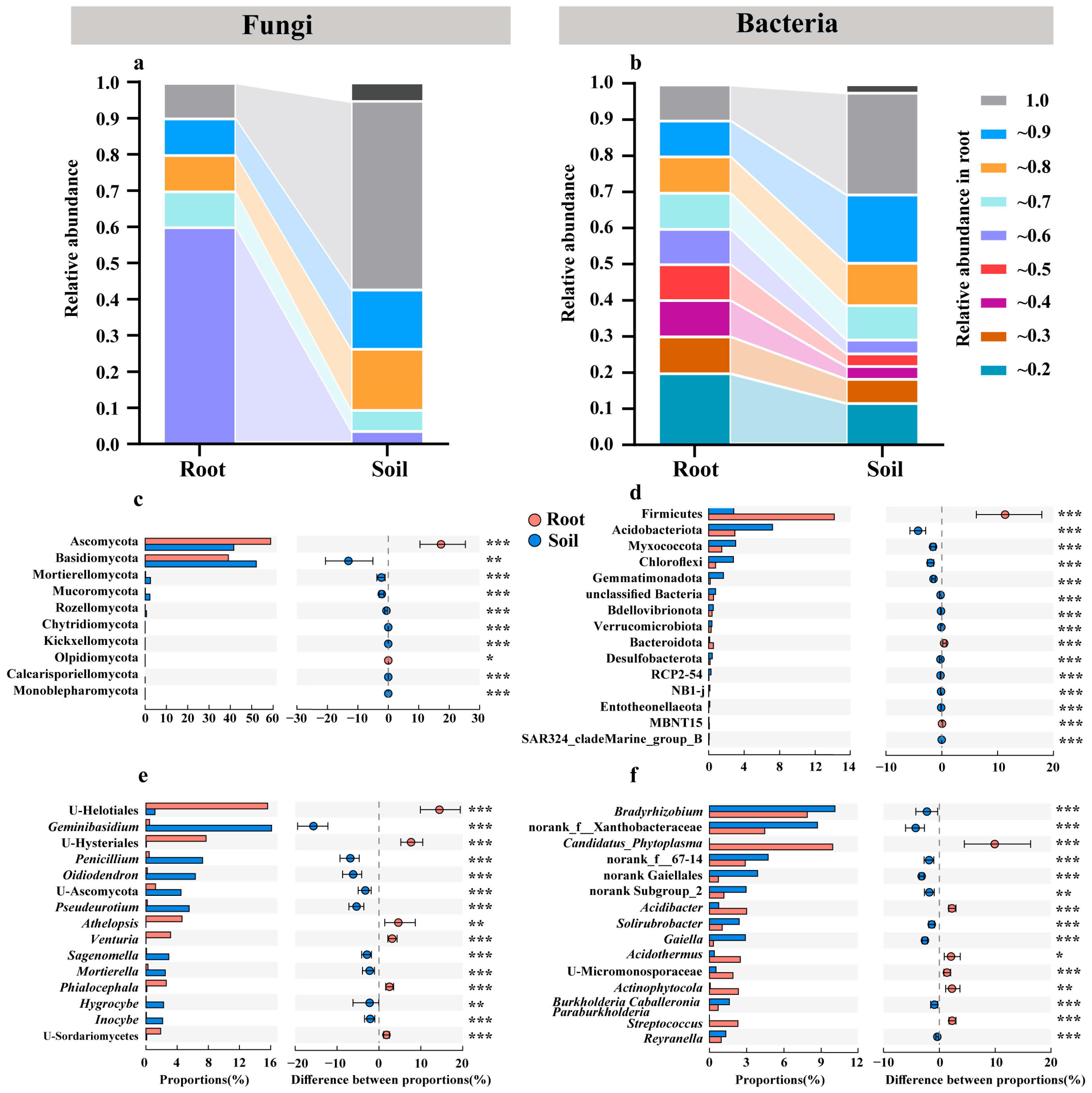
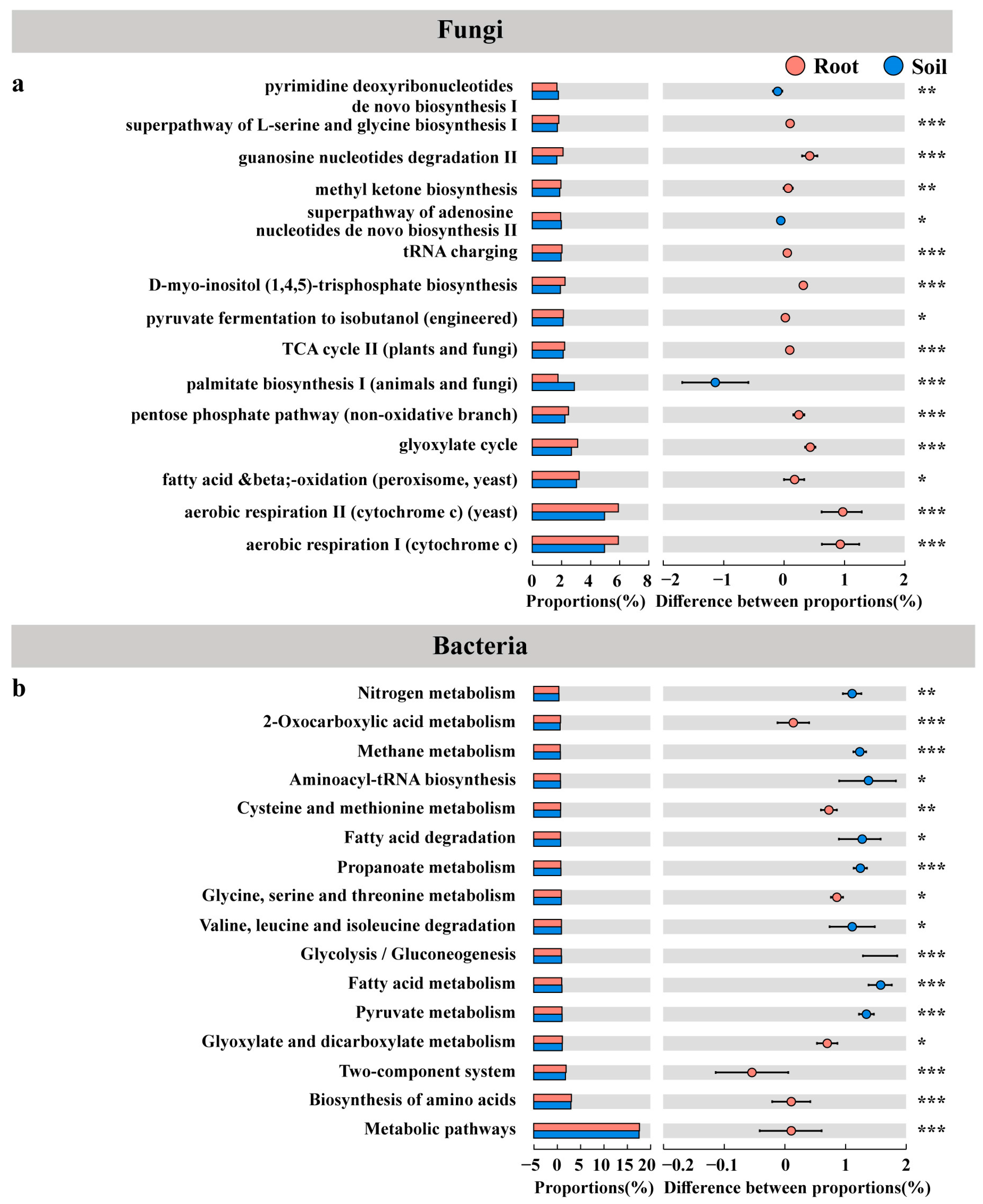
3.6. Pines Recruited More Diverse Microbial Communities from Similar Soil Microbial Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Merino-Martín, L.; Hernández-Cáceres, D.; Reverchon, F.; Angeles-Alvarez, G.; Zhang, G.; Dunoyer de Segonzac, D.; Dezette, D.; Stokes, A. Habitat partitioning of soil microbial communities along an elevation gradient: From plant root to landscape scale. Oikos 2023, 2023, e09034. [Google Scholar] [CrossRef]
- Zuo, Y.W.; He, P.; Zhang, J.H.; Li, W.Q.; Ning, D.H.; Zeng, Y.L.; Yang, Y.; Xia, C.Y.; Zhang, H.; Deng, H.P. Contrasting responses of multispatial soil fungal communities of Thuja sutchuenensis Franch., an extremely endangered conifer in Southwestern China. Microbiol. Spectr. 2022, 10, e0026022. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Li, L.; Wang, X.; You, J.; Li, J.; Chen, X. Elevational is the main factor controlling the soil microbial community structure in alpine tundra of the Changbai Mountain. Sci. Rep. 2020, 10, 12442. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, A.; Meng, F.; Zhao, W.; Yang, Y.; Soininen, J.; Shen, J.; Zhou, J. Embracing mountain microbiome and ecosystem functions under global change. New Phytol. 2022, 234, 1987–2002. [Google Scholar] [CrossRef]
- Shen, C.; Gunina, A.; Luo, Y.; Wang, J.; He, J.Z.; Kuzyakov, Y.; Hemp, A.; Classen, A.T.; Ge, Y. Contrasting patterns and drivers of soil bacterial and fungal diversity across a mountain gradient. Environ. Microbiol. 2020, 22, 3287–3301. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yang, T.; Shi, Y.; Zhu, Y.; He, M.; Zhao, Y.; Adams, J.M.; Chu, H. Strong partitioning of soil bacterial community composition and co-occurrence networks along a small-scale elevational gradient on Zijin Mountain. Soil Ecol. Lett. 2021, 3, 290–302. [Google Scholar] [CrossRef]
- Praeg, N.; Seeber, J.; Leitinger, G.; Tasser, E.; Newesely, C.; Tappeiner, U.; Illmer, P. The role of land management and elevation in shaping soil microbial communities: Insights from the Central European Alps. Soil Biol. Biochem. 2020, 150, 107951. [Google Scholar] [CrossRef]
- Odriozola, I.; Navrátilová, D.; Tláskalová, P.; Klinerová, T.; Červenková, Z.; Kohout, P.; Větrovský, T.; Čížková, P.; Starý, M.; Baldrian, P. Predictors of soil fungal biomass and community composition in temperate mountainous forests in Central Europe. Soil Biol. Biochem. 2021, 161, 108366. [Google Scholar] [CrossRef]
- Puissant, J.; Cécillon, L.; Mills, R.T.E.; Robroek, B.J.M.; Gavazov, K.; De Danieli, S.; Spiegelberger, T.; Buttler, A.; Brun, J.-J. Seasonal influence of climate manipulation on microbial community structure and function in mountain soils. Soil Biol. Biochem. 2015, 80, 296–305. [Google Scholar] [CrossRef] [Green Version]
- Cai, Z.; Wang, X.; Bhadra, S.; Gao, Q. Distinct factors drive the assembly of quinoa-associated microbiomes along elevation. Plant Soil. 2020, 448, 55–69. [Google Scholar] [CrossRef]
- Wu, S.; Wang, Y.; Wang, Z.; Shrestha, N.; Liu, J. Species divergence with gene flow and hybrid speciation on the Qinghai-Tibet Plateau. New Phytol. 2022, 234, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Armenise, L.; Simeone, M.C.; Piredda, R.; Schirone, B. Validation of DNA barcoding as an efficient tool for taxon identification and detection of species diversity in Italian conifers. Eur. J. Forest. Res. 2012, 131, 1337–1353. [Google Scholar] [CrossRef]
- Jin, W.T.; Gernandt, D.S.; Wehenkel, C.; Xia, X.M.; Wei, X.X.; Wang, X.Q. Phylogenomic and ecological analyses reveal the spatiotemporal evolution of global pines. Proc. Natl. Acad. Sci. USA 2021, 118, e2022302118. [Google Scholar] [CrossRef]
- Meng, J.; Mao, J.F.; Zhao, W.; Xing, F.; Chen, X.; Liu, H.; Xing, Z.; Wang, X.R.; Li, Y. Adaptive differentiation in seedling traits in a hybrid pine species complex, Pinus densata and its parental species, on the Tibetan plateau. PLoS ONE 2015, 10, e0118501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.R.; Szmidt, A.E. Hybridization and chloroplast dna variation in a pinus species complex from Asia. Evolution 1994, 48, 1020–1031. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.R.; Szmidt, A.E.; Savolainen, O. Genetic composition and diploid hybrid speciation of a high mountain pine, Pinus densata, native to the Tibetan plateau. Genetics 2001, 159, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.R.; Szmidt, A.E.; Lewandowski, A.; Wang, Z.R. Evolutionary analysis of Pinus densata Masters, a putative Tertiary hybrid: 1. Allozyme variation. Theor. Appl. Genet. 1990, 80, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Ge, S.; Hong, D.Y. Allozyme diversity and population genetic structure of Pinus densata master in Northwestern Yunnan, China. Biochem. Genet. 2000, 38, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.L.; Zhang, D.; Hong, D.Y.; Wang, X.R. Chromosomal localization of 5S and 18S-5.8S-25S ribosomal DNA sites in five Asian pines using fluorescence in situ hybridization. Theor. Appl. Genet. 2003, 106, 198–204. [Google Scholar] [CrossRef]
- Song, B.H.; Wang, X.Q.; Wang, X.R.; Ding, K.Y.; Hong, D.Y. Cytoplasmic composition in Pinus densata and population establishment of the diploid hybrid pine. Mol. Ecol. 2003, 12, 2995–3001. [Google Scholar] [CrossRef] [Green Version]
- Song, B.H.; Wang, X.Q.; Wang, X.R.; Sun, L.J.; Hong, D.Y.; Peng, P.H. Maternal lineages of Pinus densata, a diploid hybrid. Mol. Ecol. 2002, 11, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Fort, M. The Himalayas: From mountain building to landform evolution in a changing world. Geogr. Pol. 2011, 84, 15–37. [Google Scholar] [CrossRef] [Green Version]
- Mao, J.F.; Wang, X.R. Distinct niche divergence characterizes the homoploid hybrid speciation of Pinus densata on the Tibetan plateau. Am. Nat. 2011, 177, 424–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.; Meng, J.; Wang, B.; Zhang, L.; Xu, Y.; Zeng, Q.Y.; Li, Y.; Mao, J.F.; Wang, X.R. Weak crossability barrier but strong juvenile selection supports ecological speciation of the hybrid pine Pinus densata on the Tibetan plateau. Evolution. 2014, 68, 3120–3133. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Gao, J.; Bai, Z.; Wu, S.; Li, X.; Wang, N.; Du, X.; Fan, H.; Zhuang, G.; Bohu, T.; et al. Unraveling mechanisms and impact of microbial recruitment on oilseed rape (Brassica napus L.) and the rhizosphere mediated by plant growth-promoting rhizobacteria. Microorganisms 2021, 9, 161. [Google Scholar] [CrossRef]
- Zhou, J.; Chai, X.; Zhang, L.; George, T.S.; Wang, F.; Feng, G. Different arbuscular mycorrhizal fungi cocolonizing on a single plant root system recruit distinct microbiomes. mSystems 2020, 5, e00929-20. [Google Scholar] [CrossRef]
- Hartman, K.; Tringe, S.G. Interactions between plants and soil shaping the root microbiome under abiotic stress. Biochem. J. 2019, 476, 2705–2724. [Google Scholar] [CrossRef] [Green Version]
- Dilla-Ermita, C.J.; Lewis, R.W.; Sullivan, T.S.; Hulbert, S.H. Wheat genotype-specific recruitment of rhizosphere bacterial microbiota under controlled environments. Front. Plant Sci. 2021, 12, 718264. [Google Scholar] [CrossRef]
- Gong, S.; Feng, B.; Jian, S.P.; Wang, G.S.; Ge, Z.W.; Yang, Z.L. Elevation matters more than season in shaping the heterogeneity of soil and root associated ectomycorrhizal fungal community. Microbiol. Spectr. 2022, 10, e0195021. [Google Scholar] [CrossRef]
- Zi, H.; Jiang, Y.; Cheng, X.; Li, W.; Huang, X. Change of rhizospheric bacterial community of the ancient wild tea along elevational gradients in Ailao mountain, China. Sci. Rep. 2020, 10, 9203. [Google Scholar] [CrossRef]
- Park, K.H.; Yoo, S.; Park, M.S.; Kim, C.S.; Lim, Y.W. Different patterns of belowground fungal diversity along altitudinal gradients with respect to microhabitat and guild types. Environ. Microbiol. Rep. 2021, 13, 649–658. [Google Scholar] [CrossRef]
- Institute of Botany. Flora of China. The Chinese Academy of Sciences. 2019. Available online: http://www.iplant.cn/foc (accessed on 8 August 2022).
- Chen, L.; Fang, K.; Zhou, J.; Yang, Z.P.; Dong, X.F.; Dai, G.H.; Zhang, H.B. Enrichment of soil rare bacteria in root by an invasive plant Ageratina adenophora. Sci. Total Environ. 2019, 683, 202–209. [Google Scholar] [CrossRef]
- Yeomans, J.C.; Bremner, J.M. A rapid and precise method for routine determination of organic carbon in soil. Commun. Soil Sci. Plant Anal. 1988, 19, 1467–1476. [Google Scholar] [CrossRef]
- Dong, J.; Gruda, N.; Li, X.; Tang, Y.; Duan, Z. Impacts of elevated CO2 on nitrogen uptake of cucumber plants and nitrogen cycling in a greenhouse soil. Appl. Soil. Ecol. 2020, 145, 103342. [Google Scholar] [CrossRef]
- Liang, F.; Xu, L.; Ji, L.; He, Q.; Wu, L.; Yan, S. A new approach for biogas slurry disposal by adopting CO2-rich biogas slurry as the flower fertilizer of Spathiphyllum: Feasibility, cost and environmental pollution potential. Sci. Total Environ. 2021, 770, 145333. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic. Acids. Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [Green Version]
- Bulgarelli, D.; Garrido-Oter, R.; Münch, P.C.; Weiman, A.; Dröge, J.; Pan, Y.; McHardy, A.C.; Schulze-Lefert, P. Structure and function of the bacterial root microbiota in wild and domesticated barley. Cell Host Microbe 2015, 17, 392–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundberg, D.S.; Lebeis, S.L.; Paredes, S.H.; Yourstone, S.; Gehring, J.; Malfatti, S.; Tremblay, J.; Engelbrektson, A.; Kunin, V.; Del Rio, T.G.; et al. Defining the core Arabidopsis thaliana root microbiome. Nature 2012, 488, 86–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, R.I.; Miletto, M.; Taylor, J.W.; Bruns, T.D. Dispersal in microbes: Fungi in indoor air are dominated by outdoor air and show dispersal limitation at short distances. ISME J. 2013, 7, 1262–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyon, D.; Tsai, S.Q.; Khayter, C.; Foden, J.A.; Sander, J.D.; Joung, J.K. FLASH assembly of TALENs for high-throughput genome editing. Nat. Biotechnol. 2012, 30, 460–465. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.; Poggiali, D.; Whitaker, K.; Marshall, T.R.; Kievit, R.A. Raincloud plots: A multi-platform tool for robust data visualization. Wellcome Open Res. 2019, 4, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 23 September 2022).
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package; R Package Version 2.6-2. Available online: https://github.com/vegandevs/vegan/ (accessed on 23 September 2022).
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Xing, F.; Mao, J.F.; Meng, J.; Dai, J.; Zhao, W.; Liu, H.; Xing, Z.; Zhang, H.; Wang, X.R.; Li, Y. Needle morphological evidence of the homoploid hybrid origin of Pinus densata based on analysis of artificial hybrids and the putative parents, Pinus tabuliformis and Pinus yunnanensis. Ecol. Evol. 2014, 4, 1890–1902. [Google Scholar] [CrossRef]
- Buck, R.; Hyasat, S.; Hossfeld, A.; Flores-Rentería, L. Patterns of hybridization and cryptic introgression among one- and four-needled pinyon pines. Ann. Bot. 2020, 126, 401–411. [Google Scholar] [CrossRef]
- Ma, F.; Zhao, C.; Milne, R.; Ji, M.; Chen, L.; Liu, J. Enhanced drought-tolerance in the homoploid hybrid species Pinus densata: Implication for its habitat divergence from two progenitors. New Phytol. 2010, 185, 204–216. [Google Scholar] [CrossRef]
- Reinhold-Hurek, B.; Bünger, W.; Burbano, C.S.; Sabale, M.; Hurek, T. Roots shaping their microbiome: Global hotspots for microbial activity. Annu. Rev. Phytopathol. 2015, 53, 403–424. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Rott, M.; Schlaeppi, K.; Ver Loren van Themaat, E.; Ahmadinejad, N.; Assenza, F.; Rauf, P.; Huettel, B.; Reinhardt, R.; Schmelzer, E.; et al. Revealing structure and assembly cues for Arabidopsis root-inhabiting bacterial microbiota. Nature 2012, 488, 91–95. [Google Scholar] [CrossRef]
- Gottel, N.R.; Castro, H.F.; Kerley, M.; Yang, Z.; Pelletier, D.A.; Podar, M.; Karpinets, T.; Uberbacher, E.; Tuskan, G.A.; Vilgalys, R.; et al. Distinct microbial communities within the endosphere and rhizosphere of Populus deltoides roots across contrasting soil types. Appl. Environ. Microb. 2011, 77, 5934–5944. [Google Scholar] [CrossRef] [Green Version]
- Dombrowski, N.; Schlaeppi, K.; Agler, M.T.; Hacquard, S.; Kemen, E.; Garrido-Oter, R.; Wunder, J.; Coupland, G.; Schulze-Lefert, P. Root microbiota dynamics of perennial Arabis alpina are dependent on soil residence time but independent of flowering time. ISME J. 2017, 11, 43–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakya, M.; Gottel, N.; Castro, H.; Yang, Z.K.; Gunter, L.; Labbé, J.; Muchero, W.; Bonito, G.; Vilgalys, R.; Tuskan, G.; et al. A multifactor analysis of fungal and bacterial community structure in the root microbiome of mature Populus deltoides trees. PLoS ONE 2013, 8, e76382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bálint, M.; Tiffin, P.; Hallström, B.; O’Hara, R.B.; Olson, M.S.; Fankhauser, J.D.; Piepenbring, M.; Schmitt, I. Host genotype shapes the foliar fungal microbiome of balsam poplar (Populus balsamifera). PLoS ONE 2013, 8, e53987. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Müller, D.B.; Srinivas, G.; Garrido-Oter, R.; Potthoff, E.; Rott, M.; Dombrowski, N.; Münch, P.C.; Spaepen, S.; Remus-Emsermann, M.; et al. Functional overlap of the Arabidopsis leaf and root microbiota. Nature 2015, 528, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Coleman-Derr, D.; Desgarennes, D.; Fonseca-Garcia, C.; Gross, S.; Clingenpeel, S.; Woyke, T.; North, G.; Visel, A.; Partida-Martinez, L.P.; Tringe, S.G. Plant compartment and biogeography affect microbiome composition in cultivated and native Agave species. New Phytol. 2016, 209, 798–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere microbiome assemblage is affected by plant development. ISME J. 2014, 8, 790–803. [Google Scholar] [CrossRef] [Green Version]
- Lebeis, S.L.; Paredes, S.H.; Lundberg, D.S.; Breakfield, N.; Gehring, J.; McDonald, M.; Malfatti, S.; Glavina del Rio, T.; Jones, C.D.; Tringe, S.G.; et al. Salicylic acid modulates colonization of the root microbiome by specific bacterial taxa. Science 2015, 349, 860–864. [Google Scholar] [CrossRef] [Green Version]
- Hamonts, K.; Trivedi, P.; Garg, A.; Janitz, C.; Grinyer, J.; Holford, P.; Botha, F.C.; Anderson, I.C.; Singh, B.K. Field study reveals core plant microbiota and relative importance of their drivers. Environ. Microbiol. 2018, 20, 124–140. [Google Scholar] [CrossRef]
- Huang, C.L.; Sarkar, R.; Hsu, T.W.; Yang, C.F.; Chien, C.H.; Chang, W.C.; Chiang, T.Y. Endophytic microbiome of biofuel plant Miscanthus sinensis (Poaceae) interacts with environmental gradients. Microb. Ecol. 2020, 80, 133–144. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Bonito, G.; Reynolds, H.; Robeson II, M.S.; Nelson, J.; Hodkinson, B.P.; Tuskan, G.; Schadt, C.W.; Vilgalys, R. Plant host and soil origin influence fungal and bacterial assemblages in the roots of woody plants. Mol. Ecol. 2014, 23, 3356–3370. [Google Scholar] [CrossRef] [PubMed]
- Hiruma, K.; Gerlach, N.; Sacristán, S.; Nakano, R.T.; Hacquard, S.; Kracher, B.; Neumann, U.; Ramírez, D.; Bucher, M.; O’Connell Richard, J.; et al. Root endophyte Colletotrichum tofieldiae confers plant fitness benefits that are phosphate status dependent. Cell 2016, 165, 464–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almario, J.; Jeena, G.; Wunder, J.; Langen, G.; Zuccaro, A.; Coupland, G.; Bucher, M. Root-associated fungal microbiota of nonmycorrhizal Arabis alpina and its contribution to plant phosphorus nutrition. Proc. Natl. Acad. Sci. USA 2017, 114, e9403–e9412. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, P.J.P.; Colaianni, N.R.; Fitzpatrick, C.R.; Dangl, J.L. Beyond pathogens: Microbiota interactions with the plant immune system. Curr. Opin. Microbiol. 2019, 49, 7–17. [Google Scholar] [CrossRef]
- Wallace, J.; Laforest-Lapointe, I.; Kembel, S.W. Variation in the leaf and root microbiome of sugar maple (Acer saccharum) at an elevational range limit. PeerJ 2018, 6, e5293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Vries, F.T.; Griffiths, R.I.; Bailey, M.; Craig, H.; Girlanda, M.; Gweon, H.S.; Hallin, S.; Kaisermann, A.; Keith, A.M.; Kretzschmar, M.; et al. Soil bacterial networks are less stable under drought than fungal networks. Nat. Commun. 2018, 9, 3033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, H.; Guo, C.; Fan, B.; Fu, R.; Su, H.; Zhang, J.; Sun, L. Lingonberry (Vaccinium vitis-idaea L.) interact with Lachnum pygmaeum to mitigate drought and promote growth. Front. Plant Sci. 2022, 13, 920338. [Google Scholar] [CrossRef]
- Njeru, C.M.; Ekesi, S.; Mohamed, S.A.; Kinyamario, J.I.; Kiboi, S.; Maeda, E.E. Assessing stock and thresholds detection of soil organic carbon and nitrogen along an altitude gradient in an east Africa mountain ecosystem. Geoderma Reg. 2017, 10, 29–38. [Google Scholar] [CrossRef]
- Adamczyk, M.; Hagedorn, F.; Wipf, S.; Donhauser, J.; Vittoz, P.; Rixen, C.; Frossard, A.; Theurillat, J.P.; Frey, B. The soil microbiome of GLORIA Mountain summits in the Swiss Alps. Front. Microbiol. 2019, 10, 1080. [Google Scholar] [CrossRef]
- Fitzpatrick, C.R.; Copeland, J.; Wang, P.W.; Guttman, D.S.; Kotanen, P.M.; Johnson, M.T.J. Assembly and ecological function of the root microbiome across angiosperm plant species. Proc. Natl. Acad. Sci. USA 2018, 115, e1157–e1165. [Google Scholar] [CrossRef] [Green Version]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, D.; Tang, J.; Cai, N.; Chen, S.; He, Y.; Deng, Z.; Yang, Y.; Yang, D.; Xu, Y.; Chen, L. Effects of Microbial Communities on Elevational Gradient Adaptation Strategies of Pinus yunnanensis Franch. and Pinus densata Mast. in a Mixed Zone. Forests 2023, 14, 685. https://doi.org/10.3390/f14040685
Mu D, Tang J, Cai N, Chen S, He Y, Deng Z, Yang Y, Yang D, Xu Y, Chen L. Effects of Microbial Communities on Elevational Gradient Adaptation Strategies of Pinus yunnanensis Franch. and Pinus densata Mast. in a Mixed Zone. Forests. 2023; 14(4):685. https://doi.org/10.3390/f14040685
Chicago/Turabian StyleMu, Dejin, Junrong Tang, Nianhui Cai, Shi Chen, Yingnian He, Zijun Deng, Yi Yang, Dan Yang, Yulan Xu, and Lin Chen. 2023. "Effects of Microbial Communities on Elevational Gradient Adaptation Strategies of Pinus yunnanensis Franch. and Pinus densata Mast. in a Mixed Zone" Forests 14, no. 4: 685. https://doi.org/10.3390/f14040685