
Effects of Forest Gaps on Forest Floor Microbial Community Composition in Pinus tabulaeformis Forests in a Rocky Mountain Area, Beijing, China
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design and Litter Samples
2.3. Laboratory Analysis
2.3.1. Litter Physicochemical Properties
2.3.2. Litter Microbial Community
2.3.3. Litter Enzyme Activities and Hydrolase–Based Vector Lengths and Angles
2.4. Data Analysis
3. Results
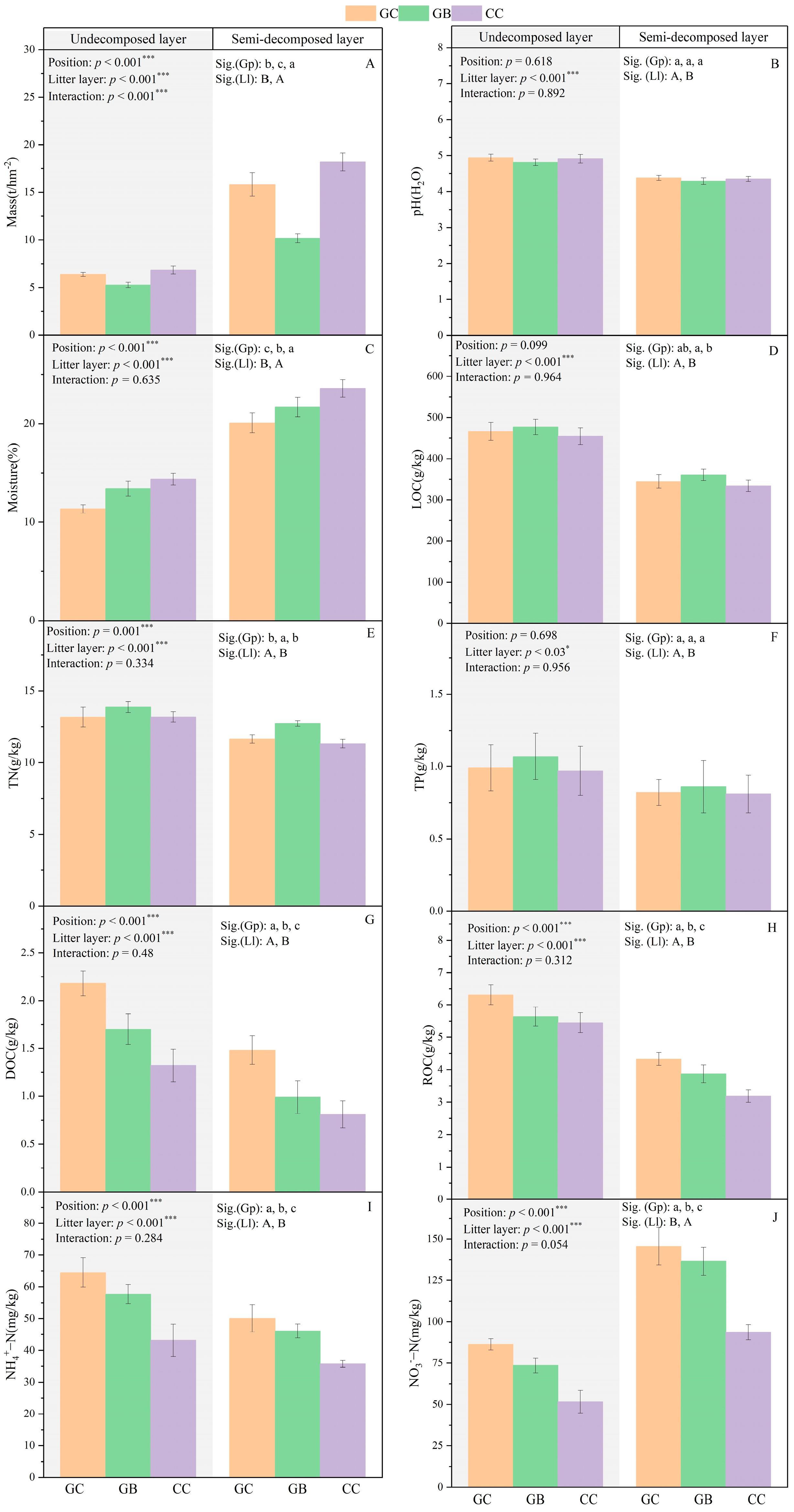
3.1. Litter Physicochemical Properties
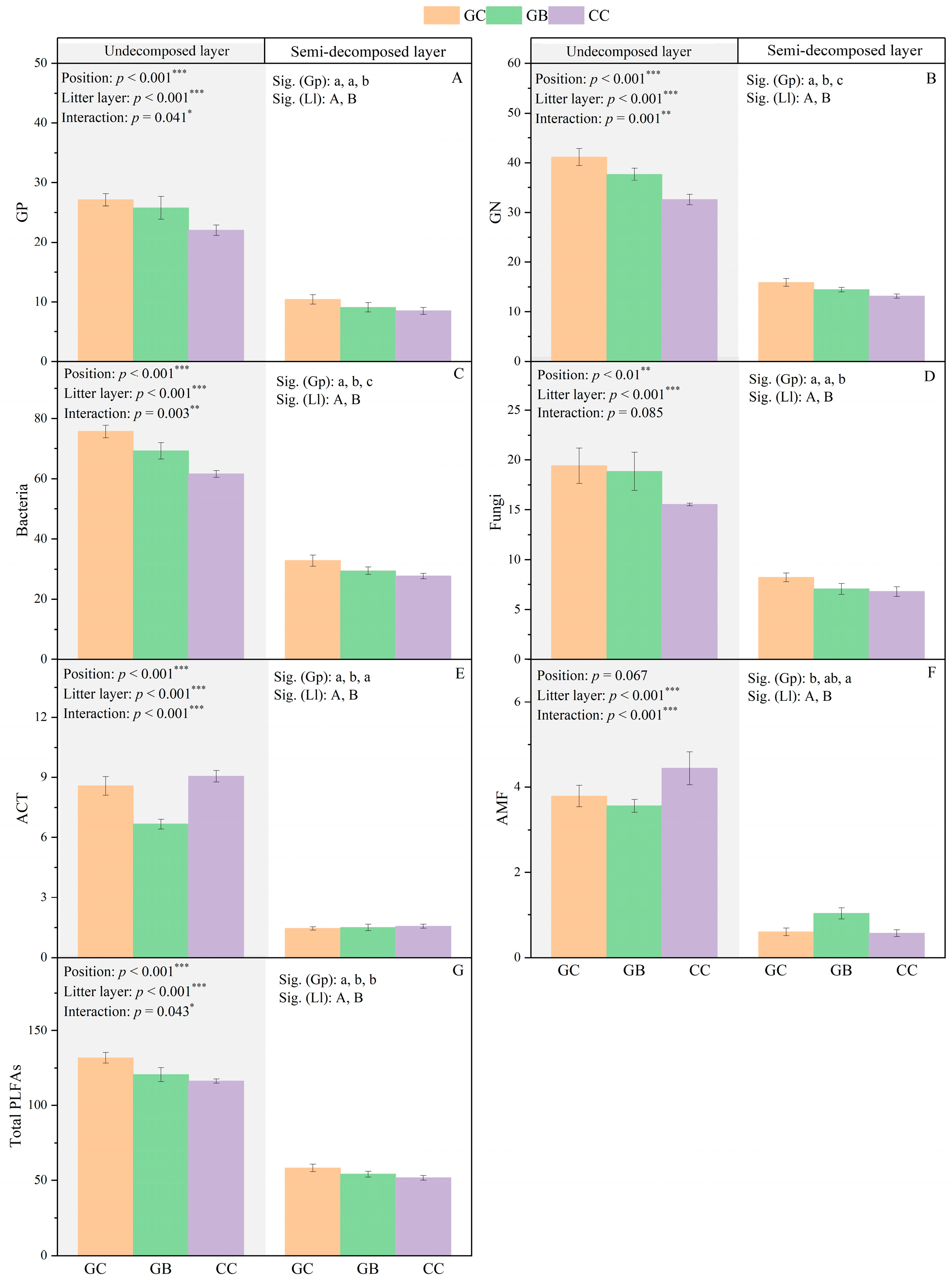
3.2. Litter Microbial Community Composition
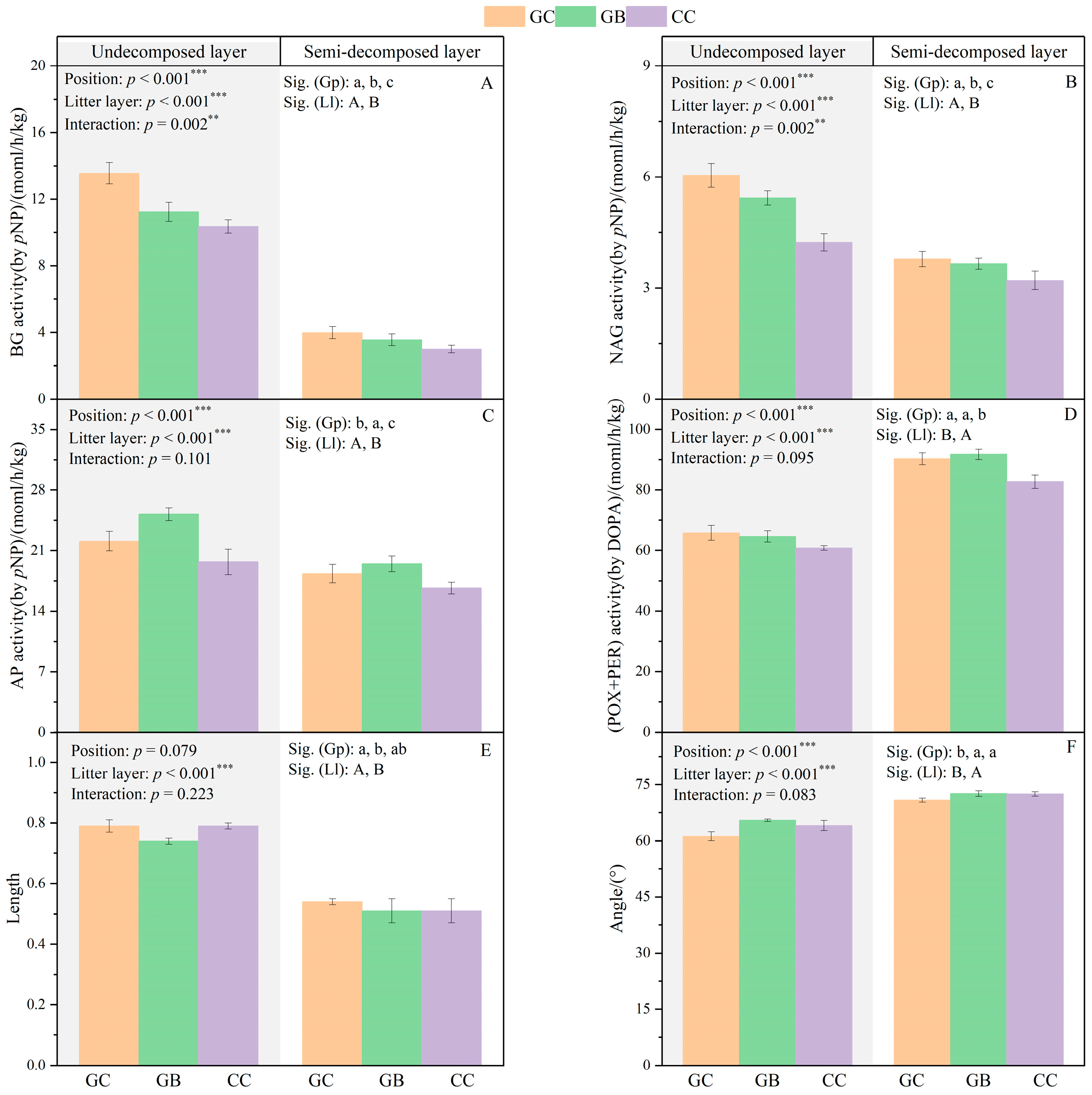
3.3. Litter Enzyme Activities and Hydrolase–Based Vector Lengths and Angles
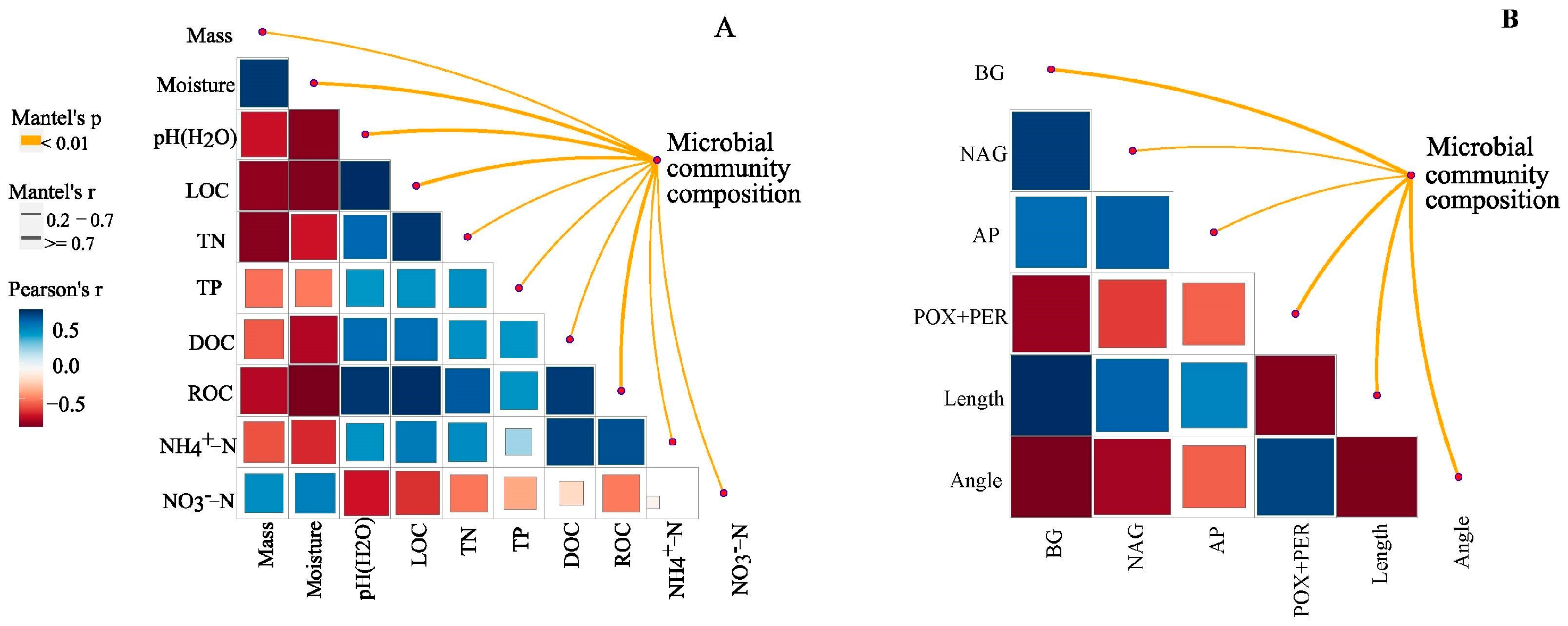
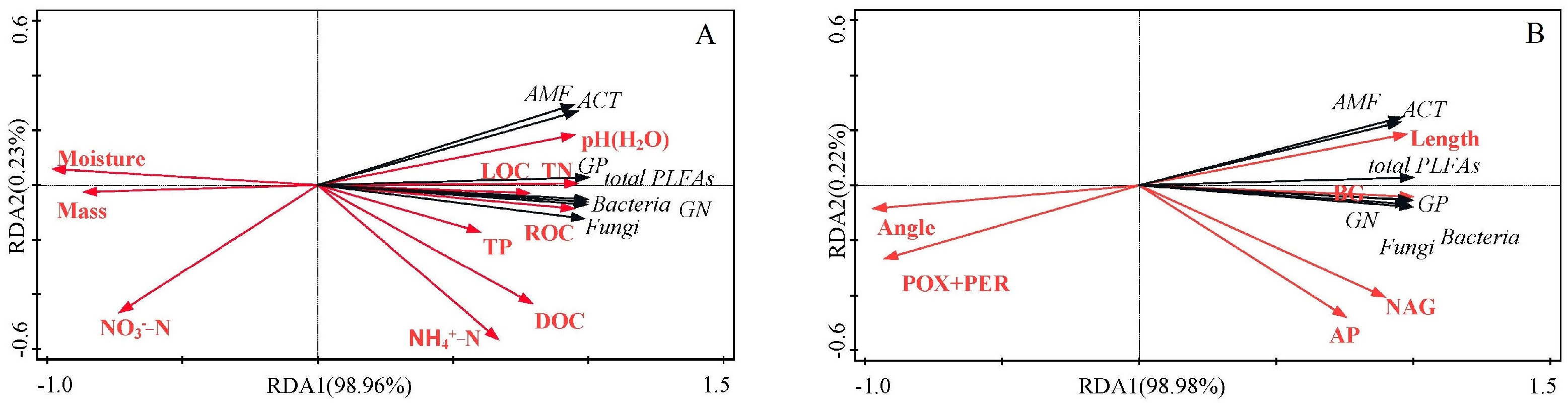
3.4. Factors Affecting Litter Microbial Community Composition
4. Discussion
4.1. Effect of Gap Position on Litter Microbial Community Composition
4.2. Effect of Litter Layers on Microbial Community Composition
4.3. Factors Affecting Microbial Community Composition
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muscolo, A.; Bagnato, S.; Sidari, M.; Mercurio, R. A review of the roles of forest canopy gaps. J. For. Res. 2014, 25, 725–736. [Google Scholar] [CrossRef]
- Mallik, A.U.; Kreutzweiser, D.P.; Spalvieri, C.M. Forest regeneration in gaps seven years after partial harvesting in riparian buffers of boreal mixed wood streams. For. Ecol. Manag. 2014, 312, 117–128. [Google Scholar] [CrossRef]
- Ritter, E.; Dalsgaard, L.; Einhorn, K.S. Light, temperature and soil moisture regimes following gap formation in a semi-natural beech dominated forest in Denmark. For. Ecol. Manag. 2005, 206, 15–33. [Google Scholar] [CrossRef]
- Guan, Y.; Fei, F.; Guan, Q.; Chen, B. Advances in studies of forest gap ecology. Sci. Silvae Sin. 2016, 52, 91–99. [Google Scholar]
- Gray, A.N.; Spies, T.A.; Pabst, R.J. Canopy gaps affect long-term patterns of tree growth and mortality in mature and old-growth forests in the Pacific Northwest. For. Ecol. Manag. 2012, 281, 111–120. [Google Scholar] [CrossRef]
- Terborgh, J.; Huanca Nuñez, N.; Alvarez Loayza, P.; Cornejo Valverde, F. Gaps contribute tree diversity to a tropical floodplain forest. Ecology 2017, 98, 2895–2903. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Wu, F.; Zhang, D.; Yang, W.; Tan, B.; Zhao, Y.; Wu, Q. The effects of forest gaps on cellulose degradation in the foliar litter of two shrub species in an alpine fir forest. Plant Soil 2015, 393, 109–122. [Google Scholar] [CrossRef]
- He, W.; Wu, F.; Yang, W.; Zhang, D.; Xu, Z.; Tan, B.; Zhao, Y.; Justine, M.F. Gap locations influence the release of carbon, nitrogen and phosphorus in two shrub foliar litter in an alpine fir forest. Sci. Rep. 2016, 6, 22014. [Google Scholar] [CrossRef]
- Hu, R.; Lin, B.; Liu, Q. Effects of forest gaps and litter on the early regeneration of Picea asperata plantations. Sci. Silvae Sin. 2011, 47, 23–29. [Google Scholar]
- Alexander, H.D.; Mack, M.C. Gap regeneration within mature deciduous forests of Interior Alaska: Implications for future forest change. For. Ecol. Manag. 2017, 396, 35–43. [Google Scholar] [CrossRef]
- Zhu, J.; Lu, D.; Zhang, W. Effects of gaps on regeneration of woody plants: A meta-analysis. J. For. Res. 2014, 25, 501–510. [Google Scholar] [CrossRef]
- Stan, A.B.; Daniels, L.D. Growth releases across a natural canopy gap-forest gradient in old-growth forests. For. Ecol. Manag. 2014, 313, 98–103. [Google Scholar] [CrossRef]
- Xie, J.; Zhang, G.; Li, Y.; Yan, X.; Zang, L.; Liu, Q.; Chen, D.; Sui, M.; He, Y. A bibliometric analysis of forest gap research during 1980–2021. Sustainability 2023, 15, 1994. [Google Scholar] [CrossRef]
- Vilhar, U.; Roženbergar, D.; Simončič, P.; Diaci, J. Variation in irradiance, soil features and regeneration patterns in experimental forest canopy gaps. Ann. For. Sci. 2014, 72, 253–266. [Google Scholar] [CrossRef]
- Muscolo, A.; Sidari, M.; Bagnato, S.; Mallamaci, C.; Mercurio, R. Gap size effects on above- and below-ground processes in a silver fir stand. Eur. J. For. Res. 2010, 129, 355–365. [Google Scholar] [CrossRef]
- Yang, Y.; Geng, Y.; Zhou, H.; Zhao, G.; Wang, L. Effects of gaps in the forest canopy on soil microbial communities and enzyme activity in a Chinese pine forest. Pedobiologia 2017, 61, 51–60. [Google Scholar] [CrossRef]
- Lyu, Q.; Luo, Y.; Dong, Y.; Xiang, Y.; Zhao, K.; Chen, G.; Chen, Y.; Fan, C.; Li, X. Effects of forest gaps on the structure and diversity of soil bacterial communities in weeping cypress forest plantations. Front. Microbiol. 2022, 13, 882949. [Google Scholar] [CrossRef]
- Niu, Y.; Liang, W.; Wei, X.; Han, Y. Forest gaps modulate the composition and co-occurrence network of soil bacterial community in Larix principis-rupprechtii Mayr plantation. Agronomy 2023, 13, 38. [Google Scholar] [CrossRef]
- Lin, N.; Bartsch, N.; Heinrich, S.; Vor, T. Long-term effects of canopy opening and liming on leaf litter production, and on leaf litter and fine-root decomposition in a European beech (Fagus sylvatica L.) forest. For. Ecol. Manag. 2015, 338, 183–190. [Google Scholar] [CrossRef]
- Duguid, M.C.; Frey, B.R.; Ellum, D.S.; Kelty, M.; Ashton, M.S. The influence of ground disturbance and gap position on understory plant diversity in upland forests of southern New England. For. Ecol. Manag. 2013, 303, 148–159. [Google Scholar] [CrossRef]
- Ritter, E.; Bjørnlund, L. Nitrogen availability and nematode populations in soil and litter after gap formation in a semi-natural beech-dominated forest. Appl. Soil. Ecol. 2005, 28, 175–189. [Google Scholar] [CrossRef]
- Li, H. Effects of forest gaps on chemical composition dynamics in decomposing litter in a subalpine coniferous forest. Sichuan Agri. Univ. 2018, 24–82. [Google Scholar] [CrossRef]
- Li, H.; Xu, L.Y.; Wu, F.Z.; Yang, W.Q.; Hu, Y. Forest gaps alter the total phenol dynamics in decomposing litter in an alpine fir forest. PLoS ONE 2016, 11, 148426. [Google Scholar] [CrossRef]
- Li, H.; Wu, F.; Yang, W.; Xu, L.; Ni, X.; He, J.; Tan, B.; Hu, Y.; Justin, M.F. The losses of condensed tannins in six foliar litters vary with gap position and season in an alpine forest. iForest-Biogeosci. For. 2016, 9, 910. [Google Scholar] [CrossRef]
- Li, D.; Li, X.; Wu, Q.; Yin, H.; Li, X. Effects of gap locations on the decomposition of fine root of Toona sinensis and soil fungal community diversity in cypress plantation forest. Acta Ecol. Sin. 2022, 42, 2784–2797. [Google Scholar]
- Šnajdr, J.; Valášková, V.; Merhautová, V.; Herinková, J.; Cajthaml, T.; Baldrian, P. Spatial variability of enzyme activities and microbial biomass in the upper layers of Quercus petraea forest soil. Soil Biol. Biochem. 2008, 40, 2068–2075. [Google Scholar] [CrossRef]
- Šnajdr, J.; Valášková, V.; Merhautová, V.; Cajthaml, T.; Baldrian, P. Activity and spatial distribution of lignocellulose-degrading enzymes during forest soil colonization by saprotrophic basidiomycetes. Enzym. Microb. Technol. 2008, 43, 186–192. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, D.; Li, X.; Liu, H.; Zhang, M.; Yang, W.; Zhang, J. Edge effects of forest gap in Pinus massoniana plantations on the decomposition of leaf litter recalcitrant components of Cinnamomum camphora and Toona ciliata. J. Appl. Ecol. 2016, 27, 1116–1124. [Google Scholar]
- Zhou, H.; Geng, Y.; Wang, L.; Yang, Y.; Yang, Y. Variation in forest floor enzyme activities among different forest gap positions in Pinus tabulaeformis plantations. Ecol. Environ. Sci. 2016, 25, 1621–1628. [Google Scholar]
- Zhang, Q.; Zak, J. Effects of gap size on litter decomposition and microbial activity in a subtropical forest. Ecology 1995, 76, 2196–2204. [Google Scholar] [CrossRef]
- Xu, L.; Yang, W.; Li, H.; Ni, X.; He, J.; Wu, F. Effects of forest gap on soluble nitrogen and soluble phosphorus of foliar litter decomposition in an alpine forest. J. Soil Water Conserv. 2014, 28, 214–221. [Google Scholar]
- Xu, L.; Yang, W.; Li, H.; Ni, X.; He, J.; Wu, F. Effects of forest gaps on soluble carbon during foliar litter decomposition in an alpine forest in the upper Yangtze River. Resour. Environ. Yangtze Basin 2015, 24, 882–891. [Google Scholar]
- Li, X.; Zhang, J.; Yang, W.; Zhang, Y.; Zhang, M.; Liu, H.; Liu, Y. Effect of forest gap on carbon release of Toona ciliata leaf litter. J. Nat. Resour. 2016, 31, 1114–1126. [Google Scholar]
- Zhang, Y.; Zhang, D.; Zhang, J.; Yang, W.; Deng, C.; Li, J.; Li, X.; Tang, S.; Zhang, M. Effects of forest gap size on litter recalcitrant components of two tree species in Pinus massoniana plantations. J. Plant Ecol. 2015, 39, 785–796. [Google Scholar]
- Li, D.; Li, X.; Su, Y.; Li, X.; Yin, H.; Li, X.; Guo, M.; He, Y. Forest gaps influence fungal community assembly in a weeping cypress forest. Appl. Microbiol. Biotechnol. 2019, 103, 3215–3224. [Google Scholar] [CrossRef]
- Muscolo, A.; Sidari, M.; Mercurio, R. Influence of gap size on organic matter decomposition, microbial biomass and nutrient cycle in Calabrian pine (Pinus laricio, Poiret) stands. For. Ecol. Manag. 2007, 242, 412–418. [Google Scholar] [CrossRef]
- Ou, J.; Zhang, J.; Cui, N.; Chen, Y.; Zhang, J.; Yang, W.; Liu, Y. The early effects of forest gap harvesting on soil microbial biomass in Pinus massoniana plantations. J. Nat. Resour. 2014, 29, 2036–2047. [Google Scholar]
- Steinberger, Y.; Stein, A.; Dorman, M.; Svoray, T.; Doniger, T.; Rinot, O.; Gil, E. A sensitive soil biological indicator to changes in land-use in regions with Mediterranean climate. Sci. Rep. 2022, 12, 22216. [Google Scholar] [CrossRef]
- Kong, Y.; Qin, H.; Zhu, C.; Tian, W.; Zhu, X.; Yu, Y.; Zhang, J. Research progress on the mechanism by which soil microorganisms affect soil health. Acta Pedol. Sin. 2023, 1–19. [Google Scholar] [CrossRef]
- Prescott, C.E.; Grayston, S.J. Tree species influence on microbial communities in litter and soil: Current knowledge and research needs. For. Ecol. Manag. 2013, 309, 19–27. [Google Scholar] [CrossRef]
- Kazuo, I.; Hiroaki, O.; Tsunehiro, W.; Ryunosuke, T.; Rieko, U.; Chao, L.; Senoo, K.; Shibata, H. High soil microbial activity in the winter season enhances nitrogen cycling in a cool-temperate deciduous forest. Soil Biol. Biochem. 2018, 124, 90–100. [Google Scholar]
- Bani, A.; Pioli, S.; Ventura, M.; Panzacchi, P.; Borruso, L.; Tognetti, R.; Tonon, G.; Brusetti, L. The role of microbial community in the decomposition of leaf litter and deadwood. Appl. Soil Ecol. 2018, 126, 75–84. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Koppen-Geiger climate classification. Hydrol. Earth. Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, Z.; Sun, G.; Chen, L.; Xu, H.; Chen, S. Biophysical controls on nocturnal sap flow in plantation forests in a semiarid region of northern China. Agric. For. Meteorol. 2020, 284, 107904. [Google Scholar] [CrossRef]
- Tan, X.; Sun, X.; Yan, H.; Wang, T.; Ren, Y.; Dong, J. Canopy gaps in the pine forests of the West Mountain in Beijing. J. Beijing For. Univ. 2000, 22, 64–68. [Google Scholar]
- Bao, S.D. Soil Agrochemical Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000; pp. 263–268. [Google Scholar]
- Vogt, D.J.; Tilley, J.P.; Edmonds, R.L. Soil and Plant Analysis for Forest Ecosystem Characterization; Higher Education Press: Beijing, China, 2015. [Google Scholar]
- Frostegård, Å.; Tunlid, A.; Bååth, E. Microbial biomass measured as total lipid phosphate in soils of different organic content. J. Microbiol. Meth. 1991, 14, 151–163. [Google Scholar] [CrossRef]
- Kaiser, C.; Koranda, M.; Kitzler, B.; Fuchslueger, L.; Schnecker, J.; Schweiger, P.; Rasche, F.; Zechmeister-Boltenstern, S.; Sessitsch, A.; Richter, A. Belowground carbon allocation by trees drives seasonal patterns of extracellular enzyme activities by altering microbial community composition in a beech forest soil. New Phytol. 2010, 187, 843–858. [Google Scholar] [CrossRef]
- Zelles, L. Fatty acid patterns of phospholipids and lipopolysaccharides in the characterisation of microbial communities in soil: A review. Biol. Fertil. Soils 1999, 29, 111–129. [Google Scholar] [CrossRef]
- Moorhead, D.L.; Sinsabaugh, R.L.; Hill, B.H.; Weintraub, M.N. Vector analysis of ecoenzyme activities reveal constraints on coupled C, N and P dynamics. Soil Biol. Biochem. 2016, 93, 1–7. [Google Scholar] [CrossRef]
- Moorhead, D.L.; Rinkes, Z.L.; Sinsabaugh, R.L.; Weintraub, M.N. Dynamics relationships between microbial biomass, respiration, inorganic nutrients and enzyme activities: Informing enzyme-based decomposition models. Front. Microbiol. 2013, 4, 223. [Google Scholar] [CrossRef]
- Zhao, Y. Effect of forest canopy on the structure of microbial community in decomposing foliar litter in a subalpine forest of western Sichuan, China. Sichuan Agri. Univ. 2016, 1–156. [Google Scholar]
- Du, T.; Liu, Y.; Yang, Y.; Zhang, Y.; You, C.; Zhang, L.; Tan, B.; Xu, Z.; Li, H. Effects of forest gaps on cellulose degradation during foliar litter decomposition in a subalpine forest of western Sichuan. J. Appl. Environ. Biol. 2021, 27, 617–624. [Google Scholar]
- Scharenbroch, B.C.; Bockheim, J.G. Gaps and soil C dynamics in old growth northern hardwood–hemlock forests. Ecosystems 2008, 11, 426–441. [Google Scholar] [CrossRef]
- Lan, X.; Ding, G. Effects of gaps on soil nutrients and soil microbial carbon in a Pinus massoniana forest, Southwestern, China. Pol. J. Environ. Stud. 2022, 31, 3653–3666. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, D.; Zhang, J.; Yang, W.; Li, X.; Zhang, Y.; Zhang, M. Effects of forest gap size on leaf litter weight loss and nutrient release of four species in Pinus massoniana plantations. Acta Ecol. Sin. 2017, 37, 513–522. [Google Scholar]
- Foudyl-Bey, S.; Brais, S.; Drouin, P. Litter heterogeneity modulates fungal activity, C mineralization and N retention in the boreal forest floor. Soil Biol. Biochem. 2016, 100, 264–275. [Google Scholar] [CrossRef]
- Wang, C.; Lu, X.; Mori, T.; Mao, Q.; Zhou, K.; Zhou, G.; Nie, Y.; Mo, J. Responses of soil microbial community to continuous experimental nitrogen additions for 13 years in a nitrogen-rich tropical forest. Soil Biol. Biochem. 2018, 121, 103–112. [Google Scholar] [CrossRef]
- Zheng, H.; Chen, Y.; Liu, Y.; Zhang, J.; Yang, W.; Yang, L.; Li, H.; Wang, L.; Wu, F.; Guo, L. Litter quality drives the differentiation of microbial communities in the litter horizon across an alpine treeline ecotone in the eastern Tibetan Plateau. Sci. Rep. 2018, 8, 10029. [Google Scholar] [CrossRef]
- Zhao, Y.; Wu, F.; Yang, W.; He, W.; Tan, B.; Xu, Z. Bacterial community changes during fir needle litter decomposition in an alpine forest in eastern Tibetan Plateau. Russ. J. Ecol. 2016, 47, 145–157. [Google Scholar] [CrossRef]
- Beth, F.T.B.; Cindy, E.P.; Sue, J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar]
- Geng, Y. Study on physcio-chemical property and health index of forest soil on Badaling Mountain in Beijing. Beijing For. Univ. 2006, 80–86. [Google Scholar]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [PubMed]
- Nagati, M.; Roy, M.; Manzi, S.; Richard, F.; Desrochers, A.; Gardes, M.; Bergeron, Y. Impact of local forest composition on soil fungal communities in a mixed boreal forest. Plant Soil 2018, 432, 345–357. [Google Scholar] [CrossRef]
- Coban, O.; De Deyn, G.B.; van der Ploeg, M. Soil microbiota as game-changers in restoration of degraded lands. Science 2022, 375, 990. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; Geng, Y.; Wang, Z.; Dai, R.; Tian, Q.; Ge, Y.; Chen, L. Effects of Forest Gaps on Forest Floor Microbial Community Composition in Pinus tabulaeformis Forests in a Rocky Mountain Area, Beijing, China. Forests 2023, 14, 1954. https://doi.org/10.3390/f14101954
Zhou H, Geng Y, Wang Z, Dai R, Tian Q, Ge Y, Chen L. Effects of Forest Gaps on Forest Floor Microbial Community Composition in Pinus tabulaeformis Forests in a Rocky Mountain Area, Beijing, China. Forests. 2023; 14(10):1954. https://doi.org/10.3390/f14101954
Chicago/Turabian StyleZhou, Hongjuan, Yuqing Geng, Zihan Wang, Ruihong Dai, Qinrui Tian, Yanling Ge, and Lixin Chen. 2023. "Effects of Forest Gaps on Forest Floor Microbial Community Composition in Pinus tabulaeformis Forests in a Rocky Mountain Area, Beijing, China" Forests 14, no. 10: 1954. https://doi.org/10.3390/f14101954