
Nitrogen Enrichment Regulates the Changes in Soil Aggregate-Associated Bacterial Community: Evidence from a Typical Temperate Forest
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
2.3. Sample Collection
2.4. Physical and Chemical Analyses
2.5. DNA Extraction, Sequencing, and Data Processing
2.6. Statistical Analyses
3. Results
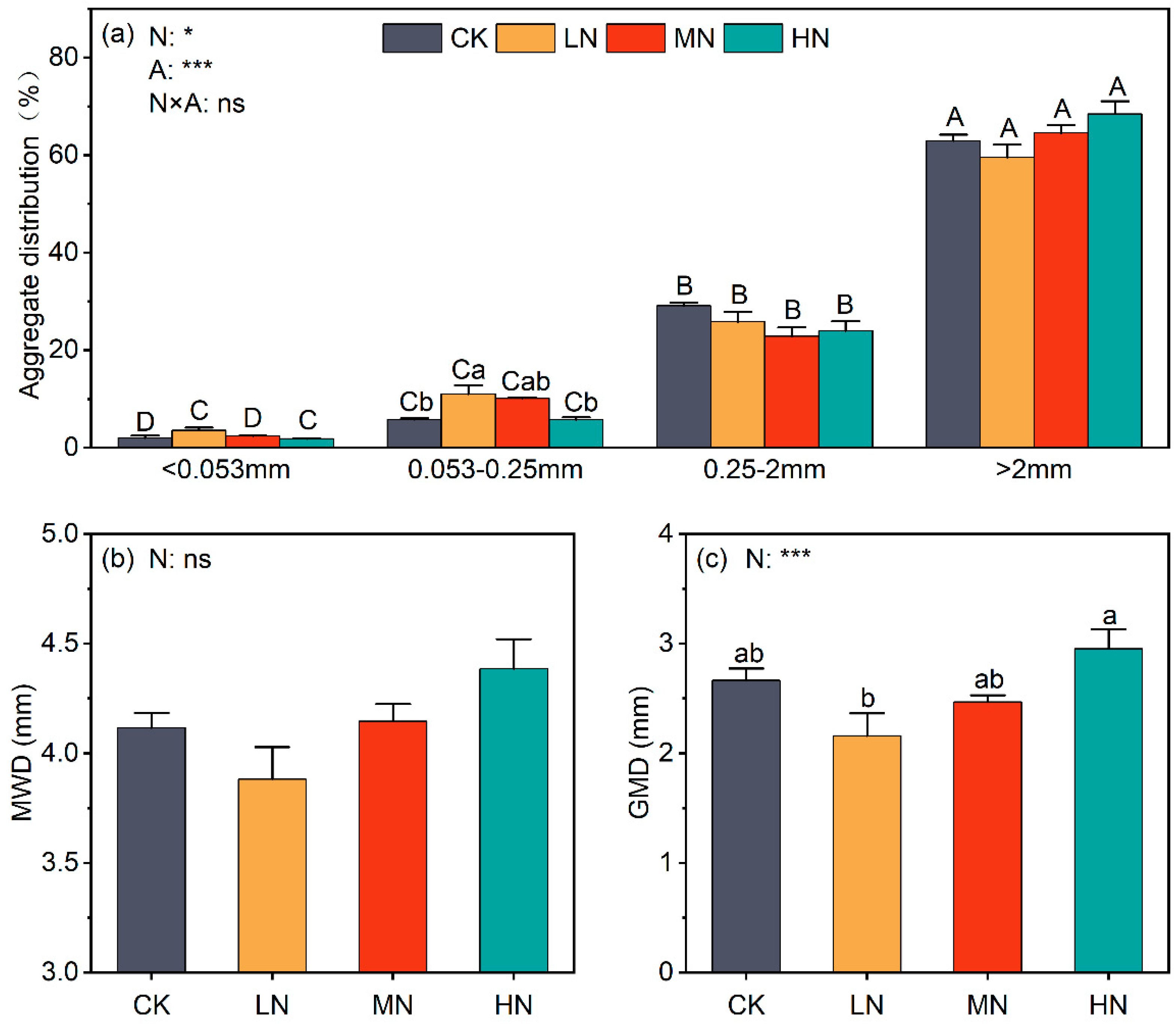
3.1. Effects of N Enrichment on Soil Aggregate Distribution and Nutrient Content
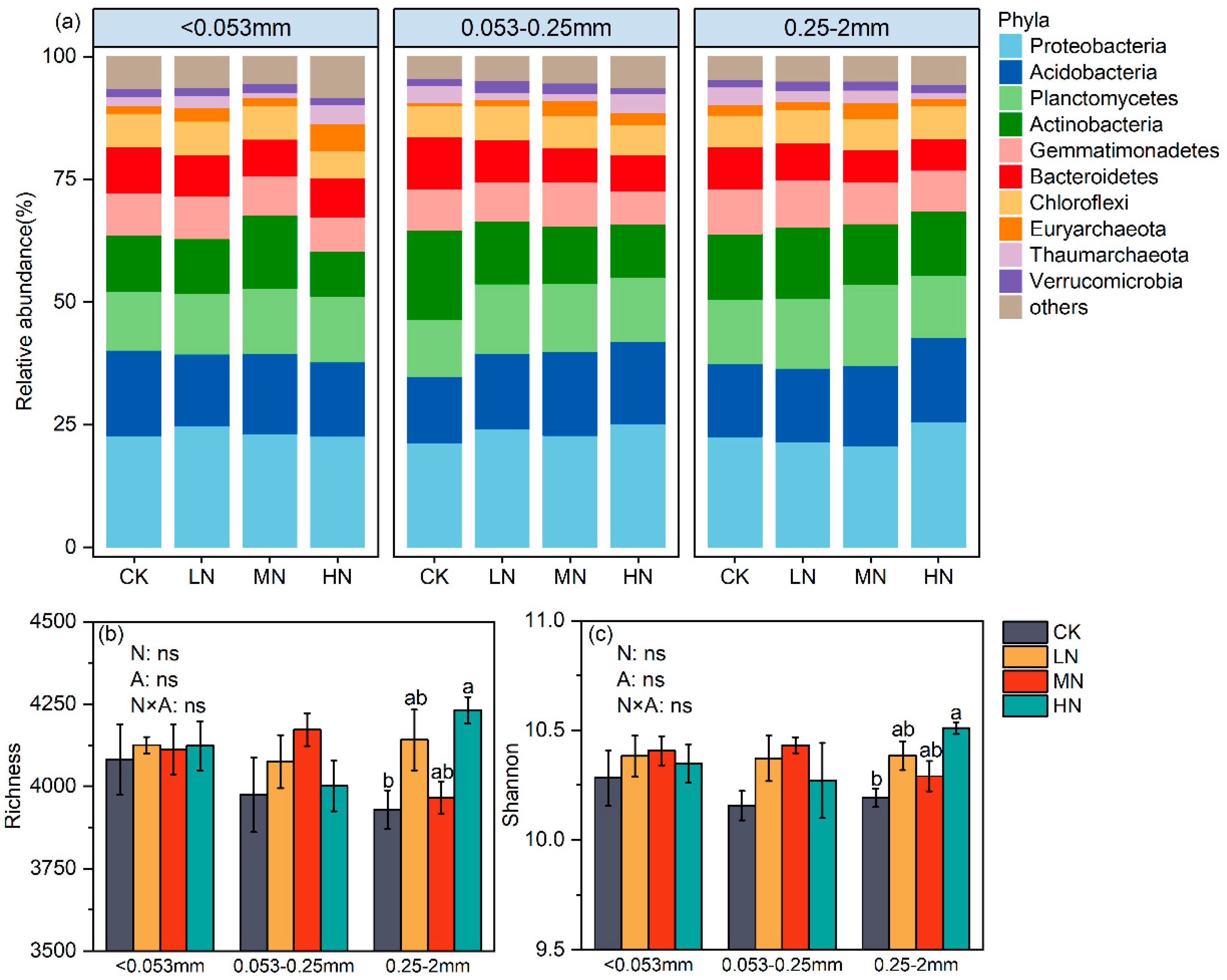
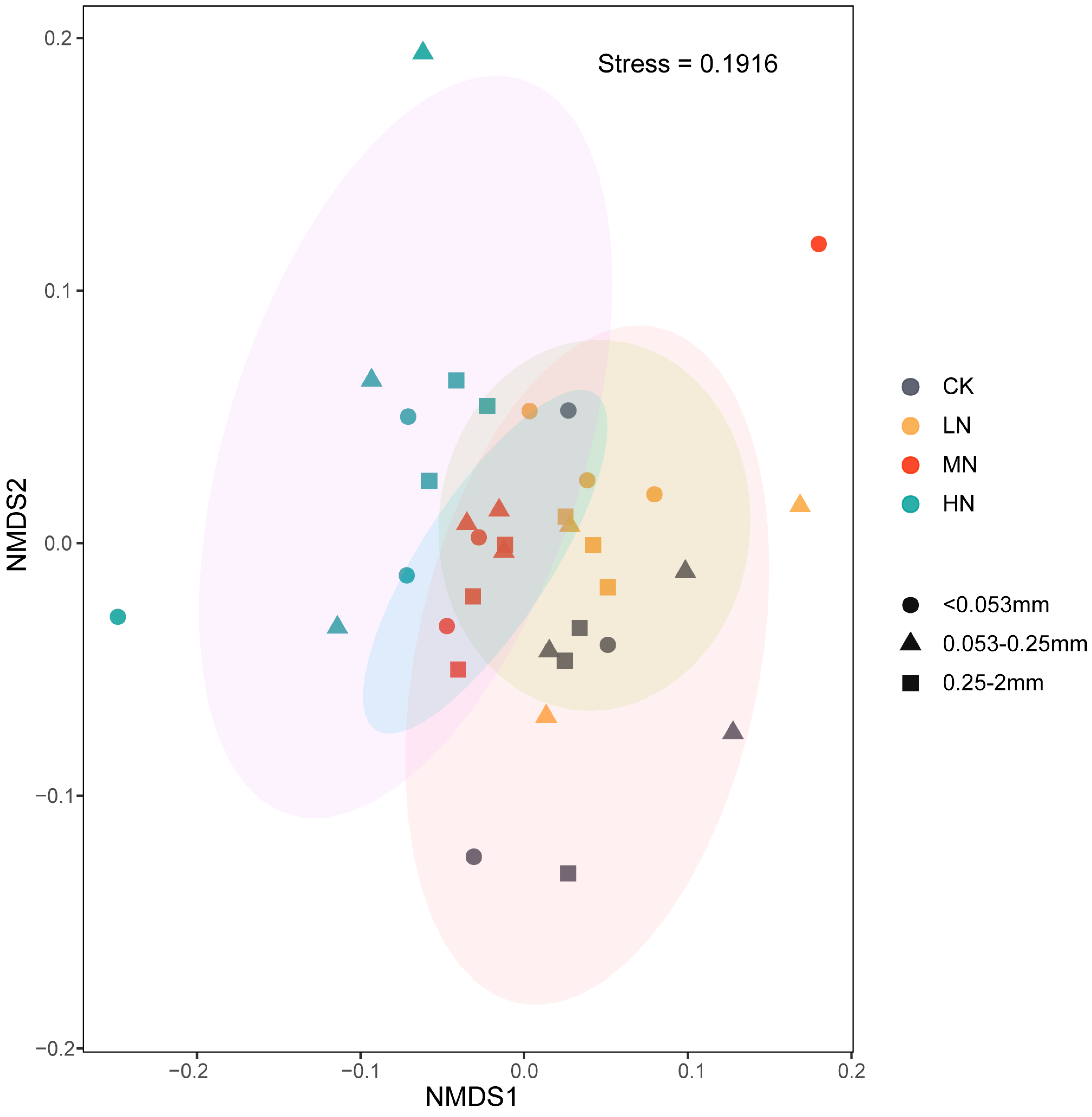
3.2. Effects of N Enrichment on Soil Bacterial Communities across Aggregate Size Fractions
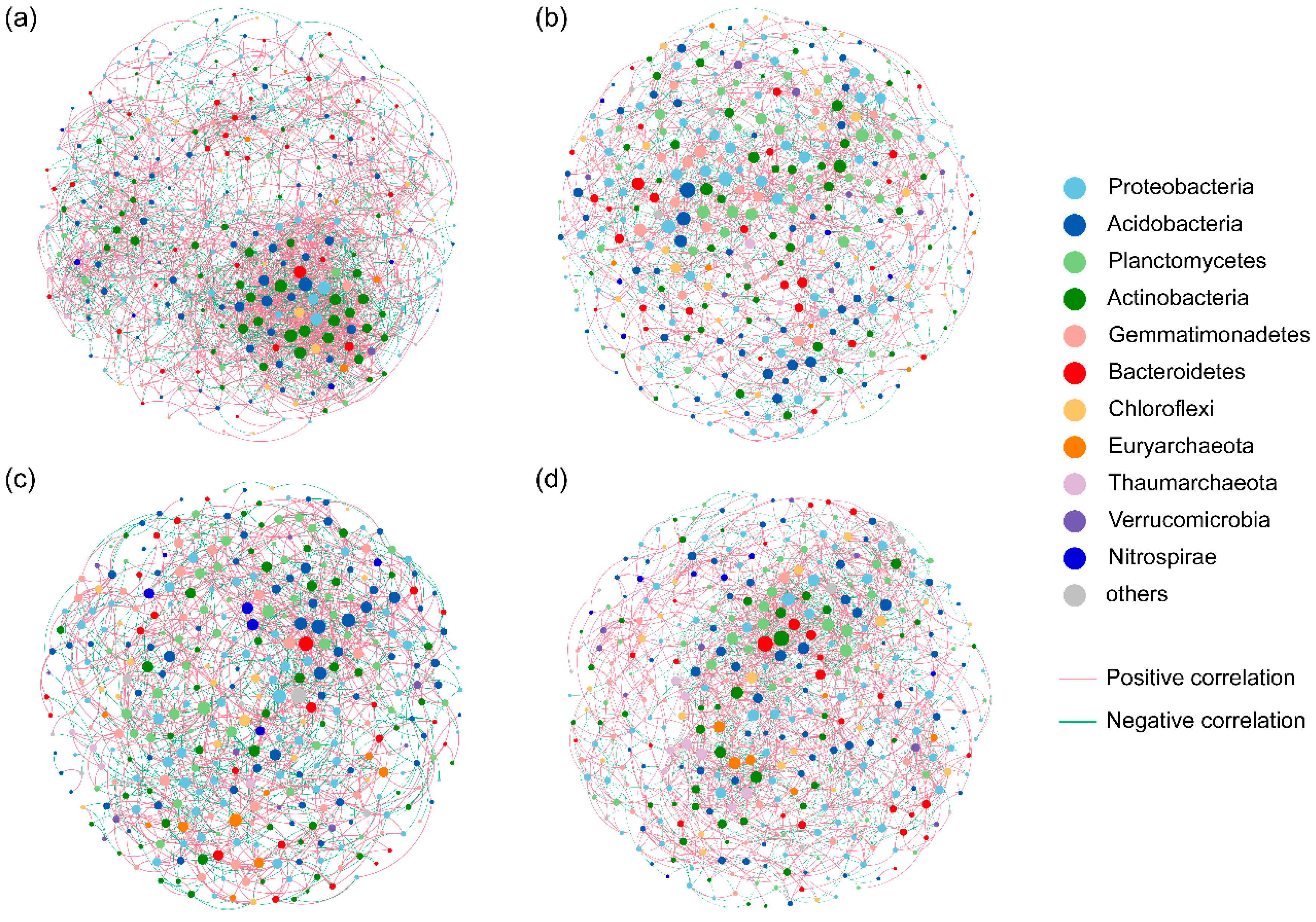
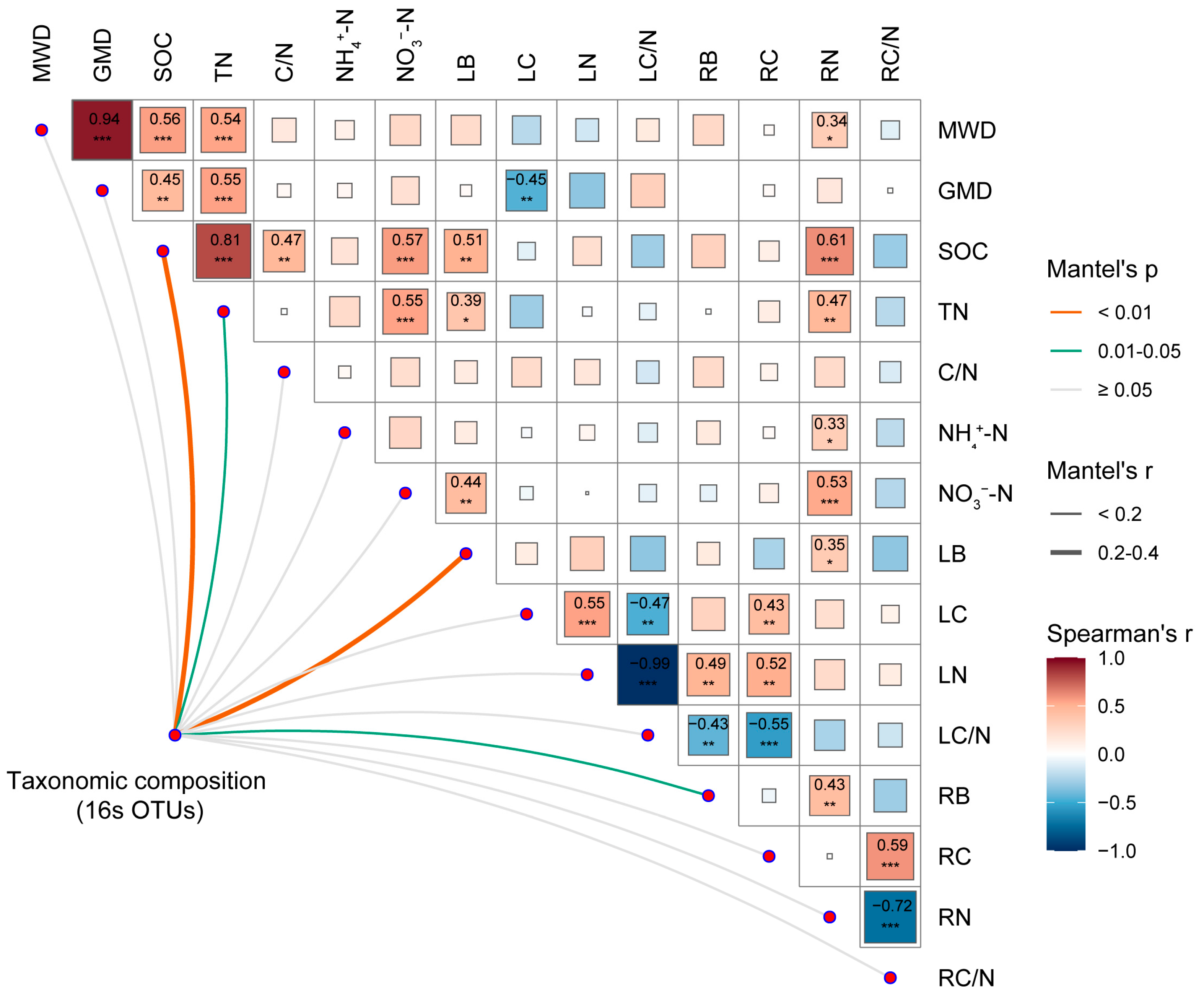
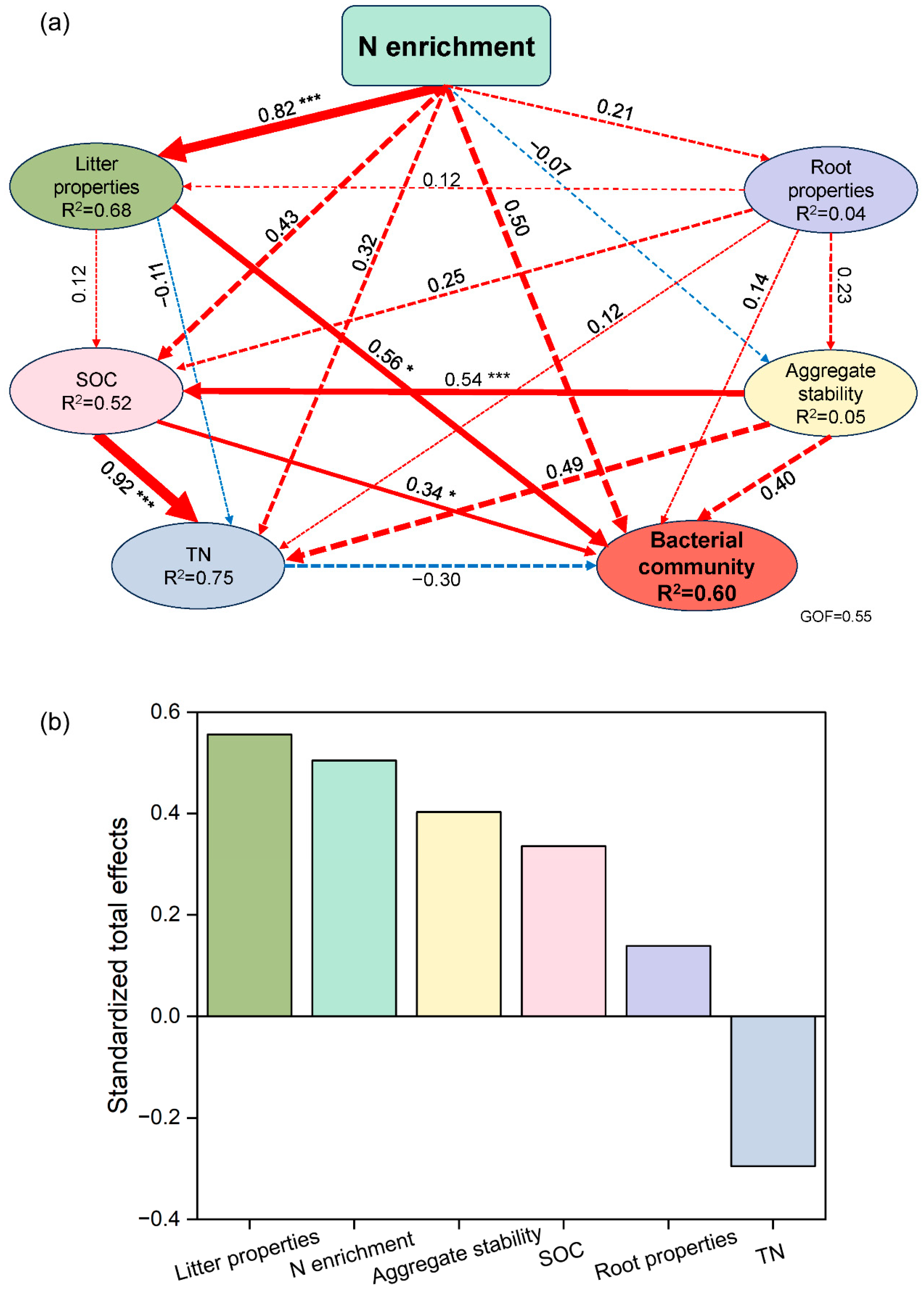
3.3. Relationship between Soil aggregate-Associated Bacteria and Environmental Factors under N Enrichment
4. Discussion
4.1. Effects of N Enrichment on Soil Aggregate Stability and Soil Properties
4.2. Effects of N Enrichment on Soil aggregate-Associated Bacterial Communities
4.3. Major Environmental Factors Affecting Soil aggregate-Associated Bacterial Communities under N Enrichment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Galloway, J.N.; Bleeker, A.; Erisman, J.W. The human creation and use of reactive nitrogen: A global and regional perspective. Annu. Rev. Environ. Resour. 2021, 46, 255–288. [Google Scholar] [CrossRef]
- Lu, X.; Jiang, H.; Zhang, X.; Liu, J.; Zhang, Z.; Jin, J.; Wang, Y.; Xu, J.; Cheng, M. Estimated global nitrogen deposition using NO2 column density. Int. J. Remote Sens. 2013, 34, 8893–8906. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Han, W.; Tang, A.; Shen, J.; Cui, Z.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Van Der Heijden, M.G.A.; Bardgett, R.D.; Van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, A.; Dubey, P.K.; Giri, B.; Moulick, D.; Srivastava, A.K.; Roychowdhury, T.; Bose, S.; Jaiswal, M.K. Combined effects of dry-wet irrigation, redox changes and microbial diversity on soil nutrient bioavailability in the rice field. Soil Tillage Res. 2023, 232, 105752. [Google Scholar] [CrossRef]
- Baldrian, P.; Kolařík, M.; Štursová, M.; Kopecký, J.; Valášková, V.; Větrovský, T.; Žifčáková, L.; Šnajdr, J.; Rídl, J.; Vlček, Č.; et al. Active and total microbial communities in forest soil are largely different and highly stratified during decomposition. ISME J. 2012, 6, 248–258. [Google Scholar] [CrossRef]
- López-Mondéjar, R.; Voříšková, J.; Větrovský, T.; Baldrian, P. The bacterial community inhabiting temperate deciduous forests is vertically stratified and undergoes seasonal dynamics. Soil Biol. Biochem. 2015, 87, 43–50. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, J.; Chang, S.X.; Cai, Z.; Müller, C.; Zhang, J. Nitrogen deposition affects both net and gross soil nitrogen transformations in forest ecosystems: A review. Environ. Pollut. 2019, 244, 608–616. [Google Scholar] [CrossRef]
- Wang, C.; Liu, D.; Bai, E. Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition. Soil Biol. Biochem. 2018, 120, 126–133. [Google Scholar] [CrossRef]
- Zhou, J.; Liu, X.; Xie, J.; Lyu, M.; Zheng, Y.; You, Z.; Fan, Y.; Lin, C.; Chen, G.; Chen, Y.; et al. Nitrogen addition affects soil respiration primarily through changes in microbial community structure and biomass in a subtropical natural forest. Forests 2019, 10, 435. [Google Scholar] [CrossRef]
- Liu, W.; Jiang, L.; Yang, S.; Wang, Z.; Tian, R.; Peng, Z.; Chen, Y.; Zhang, X.; Kuang, J.; Ling, N.; et al. Critical transition of soil bacterial diversity and composition triggered by nitrogen enrichment. Ecology 2020, 101, e03053. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Lauber, C.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shi, X.; Zheng, C.; Suter, H.; Huang, Z. Different responses of soil bacterial and fungal communities to nitrogen deposition in a subtropical forest. Sci. Total. Environ. 2021, 755, 142449. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Zhong, X.; Xia, Y.; Xu, L.; Zeng, Q.; Yang, L.; Fan, Y. Long-term nitrogen addition exerts minor effects on microbial community but alters sensitive microbial species in a subtropical natural forest. Forests 2023, 14, 928. [Google Scholar] [CrossRef]
- Lu, J.; Li, S.; Liang, G.; Wu, X.; Zhang, Q.; Gao, C.; Li, J.; Jin, D.; Zheng, F.; Zhang, M.; et al. The contribution of microorganisms to soil organic carbon accumulation under fertilization varies among aggregate size classes. Agronomy 2021, 11, 2126. [Google Scholar] [CrossRef]
- Chen, X.; Li, Z.; Liu, M.; Jiang, C.; Che, Y. Microbial community and functional diversity associated with different aggregate fractions of a paddy soil fertilized with organic manure and/or NPK fertilizer for 20 years. J. Soils Sediments 2015, 15, 292–301. [Google Scholar] [CrossRef]
- Lu, X.; Hou, E.; Guo, J.; Gilliam, F.S.; Li, J.; Tang, S.; Kuang, Y. Nitrogen addition stimulates soil aggregation and enhances carbon storage in terrestrial ecosystems of China: A meta-analysis. Glob. Chang. Biol. 2021, 27, 2780–2792. [Google Scholar] [CrossRef]
- Liao, H.; Zhang, Y.; Zuo, Q.; Du, B.; Chen, W.; Wei, D.; Huang, Q. Contrasting responses of bacterial and fungal communities to aggregate-size fractions and long-term fertilizations in soils of northeastern China. Sci. Total Environ. 2018, 635, 784–792. [Google Scholar] [CrossRef]
- Biesgen, D.; Frindte, K.; Maarastawi, S.; Knief, C. Clay content modulates differences in bacterial community structure in soil aggregates of different size. Geoderma 2020, 376, 114544. [Google Scholar] [CrossRef]
- Yang, C.; Liu, N.; Zhang, Y. Soil aggregates regulate the impact of soil bacterial and fungal communities on soil respiration. Geoderma 2019, 337, 444–452. [Google Scholar] [CrossRef]
- Kong, A.Y.Y.; Scow, K.M.; Córdova-Kreylos, A.L.; Holmes, W.E.; Six, J. Microbial community composition and carbon cycling within soil microenvironments of conventional, low-input, and organic cropping systems. Soil Biol. Biochem. 2011, 43, 20–30. [Google Scholar] [CrossRef]
- Wang, X.; Bian, Q.; Jiang, Y.; Zhu, L.; Chen, Y.; Liang, Y.; Sun, B. Organic amendments drive shifts in microbial community structure and keystone taxa which increase C mineralization across aggregate size classes. Soil Biol. Biochem. 2021, 153, 108062. [Google Scholar] [CrossRef]
- Zhang, Y.; Shangguan, Z. Long-term N addition accelerated organic carbon mineralization in aggregates by shifting microbial community composition. Agric. Ecosyst. Environ. 2023, 342, 108249. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Z.; Li, X.; Ping, Q.; Yuan, X.; Agathokleous, E.; Feng, Z. Interactive effects of ozone exposure and nitrogen addition on the rhizosphere bacterial community of poplar saplings. Sci. Total Environ. 2021, 754, 142134. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Dorodnikov, M.; Yang, S.; Zhang, Y.; Filley, T.R.; Turco, R.F.; Zhang, Y.; Xu, Z.; Li, H.; Jiang, Y. Responses of enzymatic activities within soil aggregates to 9-year nitrogen and water addition in a semi-arid grassland. Soil Biol. Biochem. 2015, 81, 159–167. [Google Scholar] [CrossRef]
- Huang, L.; Shao, M.a. Advances and perspectives on soil water research in China’s Loess Plateau. Earth-Sci. Rev. 2019, 199, 102962. [Google Scholar] [CrossRef]
- Sun, J.-J.; Zhou, T.; Zhang, R.-T.; Jia, Y.; Zhao, Y.-M.; Yang, J.; Zhao, G.-F. Comparative transcriptomes and development of expressed sequence tag-simple sequence repeat markers for two closely related oak species. J. Syst. Evol. 2019, 57, 440–450. [Google Scholar] [CrossRef]
- Deng, L.; Wang, K.-B.; Chen, M.-L.; Shangguan, Z.-P.; Sweeney, S. Soil organic carbon storage capacity positively related to forest succession on the Loess Plateau, China. Catena 2013, 110, 1–7. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, Y.; Tang, Z.; Shangguan, Z.; Chang, F.; Jia, F.A.; Chen, Y.; He, X.; Shi, W.; Deng, L. Effects of grassland afforestation on structure and function of soil bacterial and fungal communities. Sci. Total Environ. 2019, 676, 396–406. [Google Scholar] [CrossRef]
- Deng, L.; Wang, K.; Zhu, G.; Liu, Y.; Chen, L.; Shangguan, Z. Changes of soil carbon in five land use stages following 10 years of vegetation succession on the Loess Plateau, China. Catena 2018, 171, 185–192. [Google Scholar] [CrossRef]
- Wang, J.; Liu, G.; Zhang, C.; Wang, G.; Fang, L.; Cui, Y. Higher temporal turnover of soil fungi than bacteria during long-term secondary succession in a semiarid abandoned farmland. Soil Tillage Res. 2019, 194, 104305. [Google Scholar] [CrossRef]
- Fu, Y.; Xu, W.; Wen, Z.; Han, M.; Sun, J.; Tang, A.; Liu, X. Enhanced atmospheric nitrogen deposition at a rural site in northwest China from 2011 to 2018. Atmos. Res. 2020, 245, 105071. [Google Scholar] [CrossRef]
- Li, H.; Yang, S.; Xu, Z.; Yan, Q.; Li, X.; van Nostrand, J.D.; He, Z.; Yao, F.; Han, X.; Zhou, J.; et al. Responses of soil microbial functional genes to global changes are indirectly influenced by aboveground plant biomass variation. Soil Biol. Biochem. 2017, 104, 18–29. [Google Scholar] [CrossRef]
- Tian, P.; Zhang, J.; Müller, C.; Cai, Z.; Jin, G. Effects of six years of simulated N deposition on gross soil N transformation rates in an old-growth temperate forest. J. For. Res. 2018, 29, 647–656. [Google Scholar] [CrossRef]
- Wang, Q.; Tian, P.; Liu, S.; Sun, T. Inhibition effects of N deposition on soil organic carbon decomposition was mediated by N types and soil nematode in a temperate forest. Appl. Soil Ecol. 2017, 120, 105–110. [Google Scholar] [CrossRef]
- Sainju, U.M. Carbon and nitrogen pools in soil aggregates separated by dry and wet sieving methods. Soil Sci. 2006, 171, 937–949. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis; SSSA Book Series; Wiley: Hoboken, NJ, USA, 1996; pp. 961–1010. [Google Scholar]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Obalum, S.E.; Uteau-Puschmann, D.; Peth, S. Reduced tillage and compost effects on soil aggregate stability of a silt-loam Luvisol using different aggregate stability tests. Soil Tillage Res. 2019, 189, 217–228. [Google Scholar] [CrossRef]
- Xia, R.; Shi, D.; Ni, S.; Wang, R.; Zhang, J.; Song, G. Effects of soil erosion and soil amendment on soil aggregate stability in the cultivated-layer of sloping farmland in the Three Gorges Reservoir area. Soil Tillage Res. 2022, 223, 105447. [Google Scholar] [CrossRef]
- Chang, R.; Zhou, W.; Fang, Y.; Bing, H.; Sun, X.; Wang, G. Anthropogenic nitrogen deposition increases soil carbon by enhancing new carbon of the soil aggregate formation. J. Geophys. Res. Biogeosci. 2019, 124, 572–584. [Google Scholar] [CrossRef]
- Sun, L.; Jing, H.; Wang, G.; Liu, G. Nitrogen addition increases the contents of glomalin-related soil protein and soil organic carbon but retains aggregate stability in a Pinus tabulaeformis forest. PeerJ 2018, 6, e5039. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Geng, S.; Zhou, X.; Gui, H.; Zhang, L.; Huang, Z.; Wang, M.; Zhang, J.; Han, S. Nitrogen addition decreases soil aggregation but enhances soil organic carbon stability in a temperate forest. Geoderma 2022, 426, 116112. [Google Scholar] [CrossRef]
- Bai, T.; Wang, P.; Ye, C.; Hu, S. Form of nitrogen input dominates N effects on root growth and soil aggregation: A meta-analysis. Soil Biol. Biochem. 2021, 157, 108251. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, L.; Shangguan, Z. Appropriate N addition improves soil aggregate stability through AMF and glomalin-related soil proteins in a semiarid agroecosystem. Land Degrad. Dev. 2023, 34, 710–722. [Google Scholar] [CrossRef]
- von Lützow, M.; Kögel-Knabner, I. Temperature sensitivity of soil organic matter decomposition—What do we know? Biol. Fertil. Soils 2009, 46, 1–15. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, H.; Wang, J.; Tian, D.; Li, Y.; He, N.; Niu, S. Soil and climate determine differential responses of soil respiration to nitrogen and acid deposition along a forest transect. Eur. J. Soil Biol. 2019, 93, 103097. [Google Scholar] [CrossRef]
- Huang, J.Y.; Yu, H.L.; Zhang, F.J.; Li, M.; Lin, H. Water- and N-induced changes in soil C:N:P stoichiometry and its implications for N limitation of a desert steppe species, Glycyrrhiza uralensis. Pol. J. Ecol. 2016, 64, 241–254. [Google Scholar] [CrossRef]
- Lu, G.; Xie, B.; Cagle, G.A.; Wang, X.; Han, G.; Wang, X.; Hou, A.; Guan, B. Effects of simulated nitrogen deposition on soil microbial community diversity in coastal wetland of the Yellow River Delta. Sci. Total Environ. 2021, 757, 143825. [Google Scholar] [CrossRef]
- Liu, C.; Dong, Y.; Sun, Q.; Jiao, R. Soil bacterial community response to short-term manipulation of the nitrogen deposition form and dose in a Chinese fir plantation in southern china. Water Air Soil Pollut. 2016, 227, 447. [Google Scholar] [CrossRef]
- Yang, Y.; Cheng, H.; Gao, H.; An, S. Response and driving factors of soil microbial diversity related to global nitrogen addition. Land Degrad. Dev. 2020, 31, 190–204. [Google Scholar] [CrossRef]
- Dove, N.C.; Stark, J.M.; Newman, G.S.; Hart, S.C. Carbon control on terrestrial ecosystem function across contrasting site productivities: The carbon connection revisited. Ecology 2019, 100, e02695. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Sun, X.; Sun, F.; Zhao, Y.; Sun, W.; Guo, J.; Zhang, T. Sensitivity of soil fungal and bacterial community compositions to nitrogen and phosphorus additions in a temperate meadow. Plant Soil 2022, 471, 477–490. [Google Scholar] [CrossRef]
- Giovannoni, S.J.; Cameron Thrash, J.; Temperton, B. Implications of streamlining theory for microbial ecology. ISME J. 2014, 8, 1553–1565. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Knelman, J.E.; Gasarch, E.; Wang, D.; Nemergut, D.R.; Seastedt, T.R. Plant community and soil chemistry responses to long-term nitrogen inputs drive changes in alpine bacterial communities. Ecology 2016, 97, 1543–1554. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Bezemer, T.M.; Yang, J.; Lü, X.; Li, X.; Liang, W.; Han, X.; Li, Q. Changes in litter quality induced by N deposition alter soil microbial communities. Soil Biol. Biochem. 2019, 130, 33–42. [Google Scholar] [CrossRef]
- Wang, C.; Wang, G.; Wang, Y.; Rafique, R.; Ma, L.; Hu, L.; Luo, Y. Urea addition and litter manipulation alter plant community and soil microbial community composition in a Kobresia humilis meadow. Eur. J. Soil Biol. 2015, 70, 7–14. [Google Scholar] [CrossRef]
- Hossain, M.Z.; Sugiyama, S.-I. Relative importance of soil microbes and litter quality on decomposition and nitrogen cycling in grasslands. Environ. Res. 2020, 35, 912–924. [Google Scholar] [CrossRef]
- Liu, R.; Zhang, Y.; Hu, X.-F.; Wan, S.; Wang, H.; Liang, C.; Chen, F.-S. Litter manipulation effects on microbial communities and enzymatic activities vary with soil depth in a subtropical Chinese fir plantation. Forest Ecol. Manag. 2021, 480, 118641. [Google Scholar] [CrossRef]
- Li, Z.; Peng, Q.; Dong, Y.; Guo, Y. The influence of increased precipitation and nitrogen deposition on the litter decomposition and soil microbial community structure in a semiarid grassland. Sci. Total Environ. 2022, 844, 157115. [Google Scholar] [CrossRef]
- Zhao, F.Z.; Fan, X.D.; Ren, C.J.; Zhang, L.; Han, X.H.; Yang, G.H.; Wang, J.; Doughty, R. Changes of the organic carbon content and stability of soil aggregates affected by soil bacterial community after afforestation. Catena 2018, 171, 622–631. [Google Scholar] [CrossRef]
- Rahman, M.T.; Zhu, Q.H.; Zhang, Z.B.; Zhou, H.; Peng, X. The roles of organic amendments and microbial community in the improvement of soil structure of a Vertisol. Appl. Soil Ecol. 2017, 111, 84–93. [Google Scholar] [CrossRef]
- Blaud, A.; Chevallier, T.; Virto, I.; Pablo, A.-L.; Chenu, C.; Brauman, A. Bacterial community structure in soil microaggregates and on particulate organic matter fractions located outside or inside soil macroaggregates. Pedobiologia 2014, 57, 191–194. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, X.; Liu, L.; Li, T.; Dou, Y.; Qiao, J.; Wang, Y.; An, S.; Chang, S.X. Nitrogen fertilization weakens the linkage between soil carbon and microbial diversity: A global meta-analysis. Glob. Chang. Biol. 2022, 28, 6446–6461. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Xu, X.; Ju, C.; Chen, H.Y.H.; Wilsey, B.J.; Luo, Y.; Fan, W. Long-term, amplified responses of soil organic carbon to nitrogen addition worldwide. Glob. Chang. Biol. 2021, 27, 1170–1180. [Google Scholar] [CrossRef] [PubMed]
- Duchicela, J.; Vogelsang, K.M.; Schultz, P.A.; Kaonongbua, W.; Middleton, E.L.; Bever, J.D. Non-native plants and soil microbes: Potential contributors to the consistent reduction in soil aggregate stability caused by the disturbance of North American grasslands. New Phytol. 2012, 196, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wu, X.; Li, H.; Alharbi, H.; Wang, J.; Dang, P.; Chen, X.; Kuzyakov, Y.; Yan, W. Soil organic matter, nitrogen and pH driven change in bacterial community following forest conversion. Forest Ecol. Manag. 2020, 477, 118473. [Google Scholar] [CrossRef]
- Wei, M.; Hu, G.; Wang, H.; Bai, E.; Lou, Y.; Zhang, A.; Zhuge, Y. 35 years of manure and chemical fertilizer application alters soil microbial community composition in a Fluvo-aquic soil in Northern China. Eur. J. Soil Biol. 2017, 82, 27–34. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef]
- McKee, L.S.; La Rosa, S.L.; Westereng, B.; Eijsink, V.G.; Pope, P.B.; Larsbrink, J. Polysaccharide degradation by the Bacteroidetes: Mechanisms and nomenclature. Environ. Microbiol. Rep. 2021, 13, 559–581. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dependent Varibles | Regression Equation | R2 | p | n |
|---|---|---|---|---|
| Acidobacteria | Y = 0.113 + 0.029RN | 0.121 | <0.05 | 36 |
| Planctomycetes | Y = 0.120 − 0.019GMD + 0.040RN | 0.350 | <0.01 | 36 |
| Actinobacteria | Y = 0.174 − 0.003SOC | 0.176 | <0.05 | 36 |
| Bacteroidetes | Y = 0.019 + 0.00022LC/N | 0.308 | <0.001 | 36 |
| Chloroflexi | Y = 0.079 − 0.008TN | 0.237 | <0.05 | 36 |
| Verrucomicrobia | Y = 0.032 − 0.006GMD | 0.143 | <0.05 | 36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, W.; Liu, Y.; Hai, X.; Liao, Y.; Li, J.; Dong, L.; Shangguan, Z.; Deng, L. Nitrogen Enrichment Regulates the Changes in Soil Aggregate-Associated Bacterial Community: Evidence from a Typical Temperate Forest. Forests 2024, 15, 77. https://doi.org/10.3390/f15010077
Lv W, Liu Y, Hai X, Liao Y, Li J, Dong L, Shangguan Z, Deng L. Nitrogen Enrichment Regulates the Changes in Soil Aggregate-Associated Bacterial Community: Evidence from a Typical Temperate Forest. Forests. 2024; 15(1):77. https://doi.org/10.3390/f15010077
Chicago/Turabian StyleLv, Wenwen, Yulin Liu, Xuying Hai, Yang Liao, Jiwei Li, Lingbo Dong, Zhouping Shangguan, and Lei Deng. 2024. "Nitrogen Enrichment Regulates the Changes in Soil Aggregate-Associated Bacterial Community: Evidence from a Typical Temperate Forest" Forests 15, no. 1: 77. https://doi.org/10.3390/f15010077