
Identification and Characterization of a Phosphate-Solubilizing Bacterium and Its Growth-Promoting Effect on Moso Bamboo Seedlings

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Isolation of PSB
2.3. Identification of PSB
2.4. Effects of Environmental Factors on the Phosphate-Solubilizing Activity of PSB
2.5. Characterization of Indole Acetic Acid (IAA) Production
2.6. Inoculation on Moso Bamboo Seedlings
2.7. Chemical Analysis of Soil and Plant Samples
2.8. Statistical Analysis
3. Results
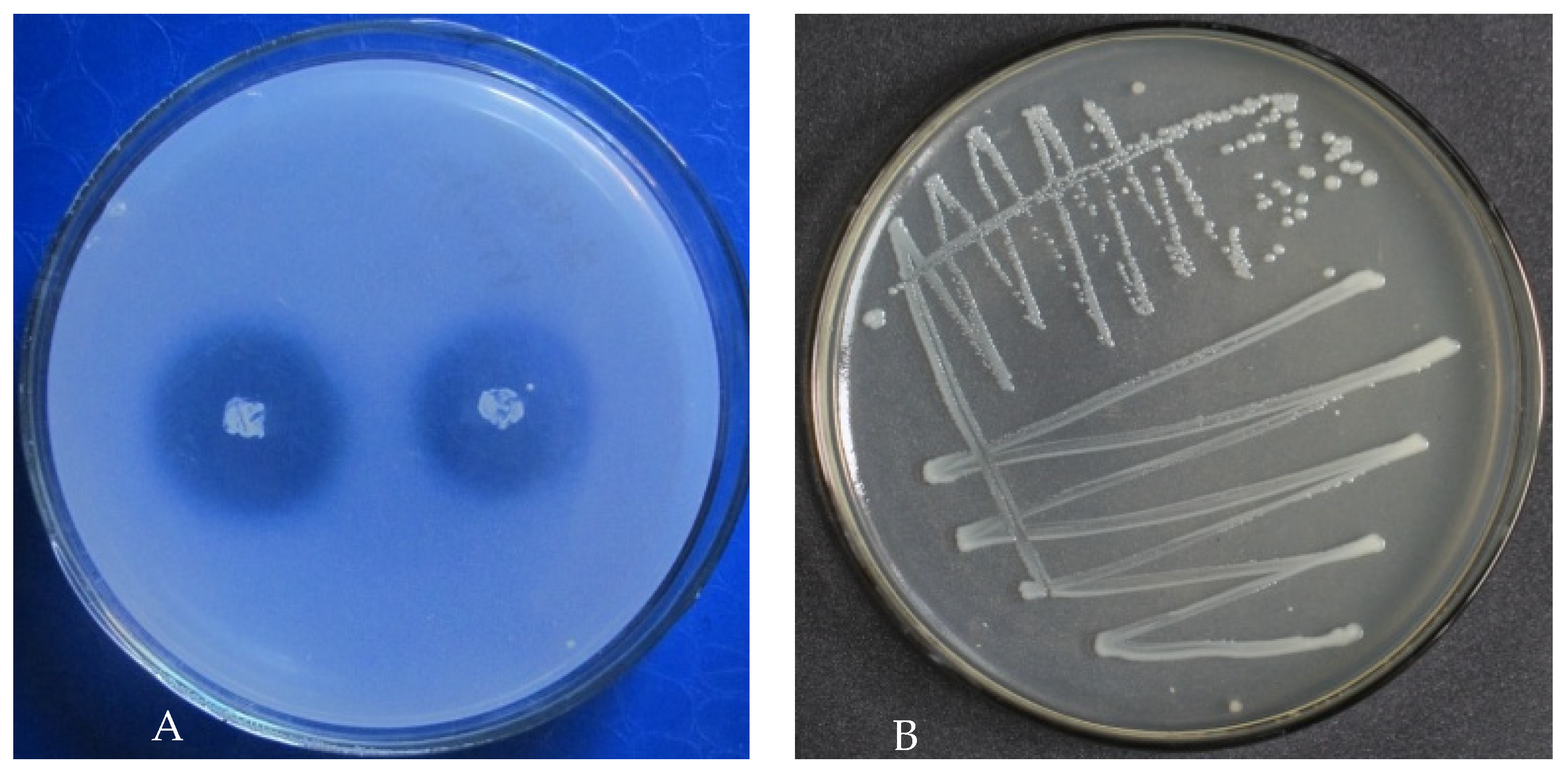
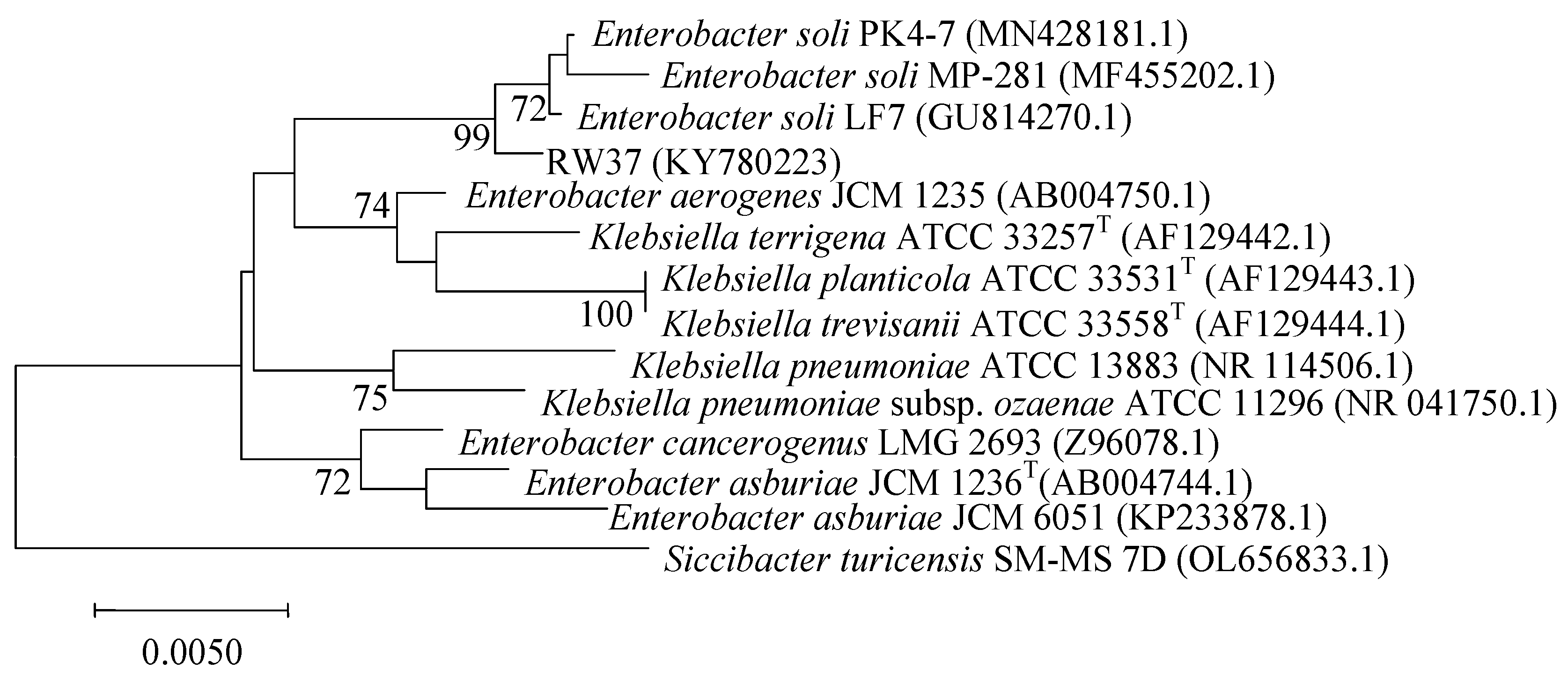
3.1. Isolation and Identification of PSB
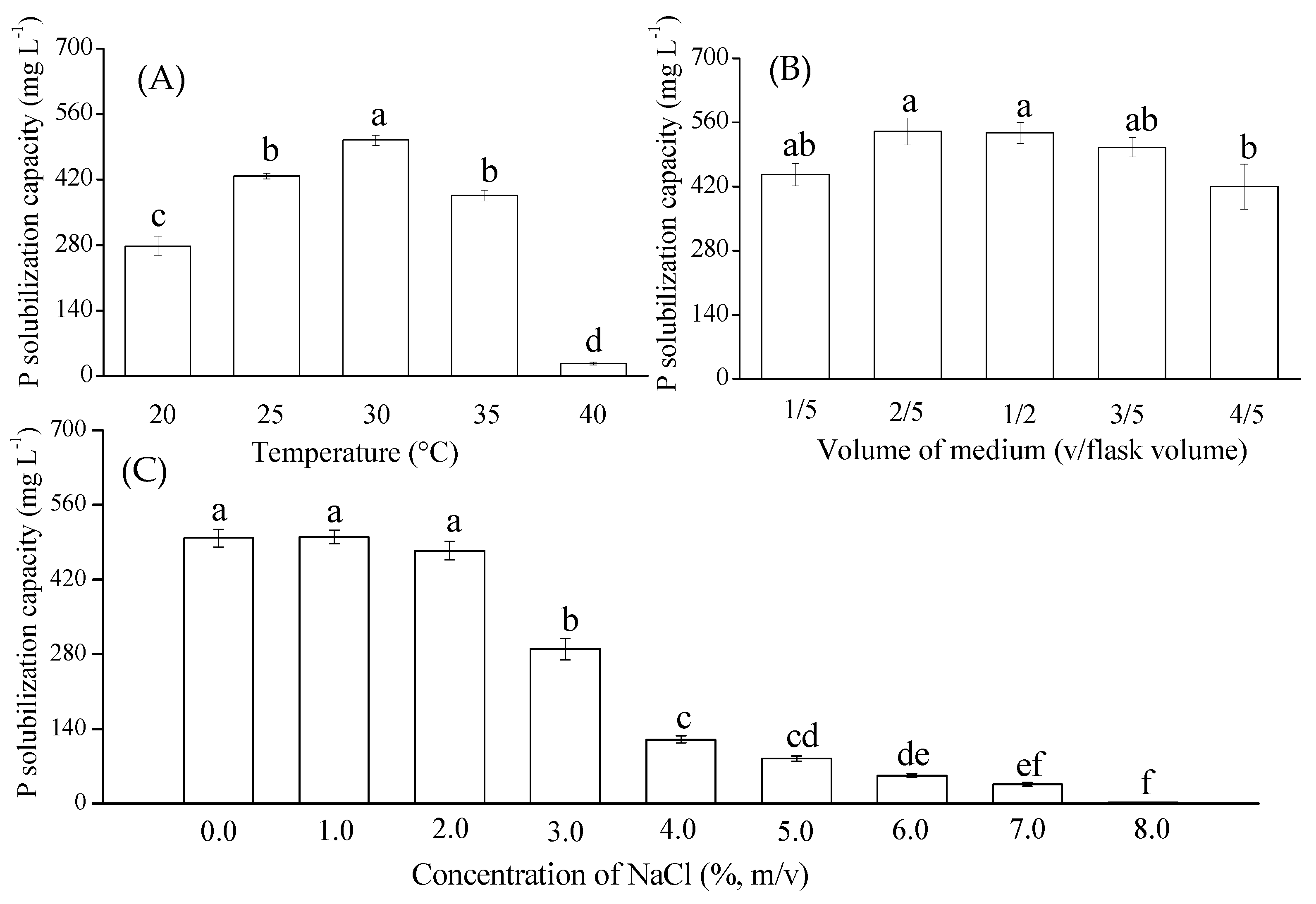
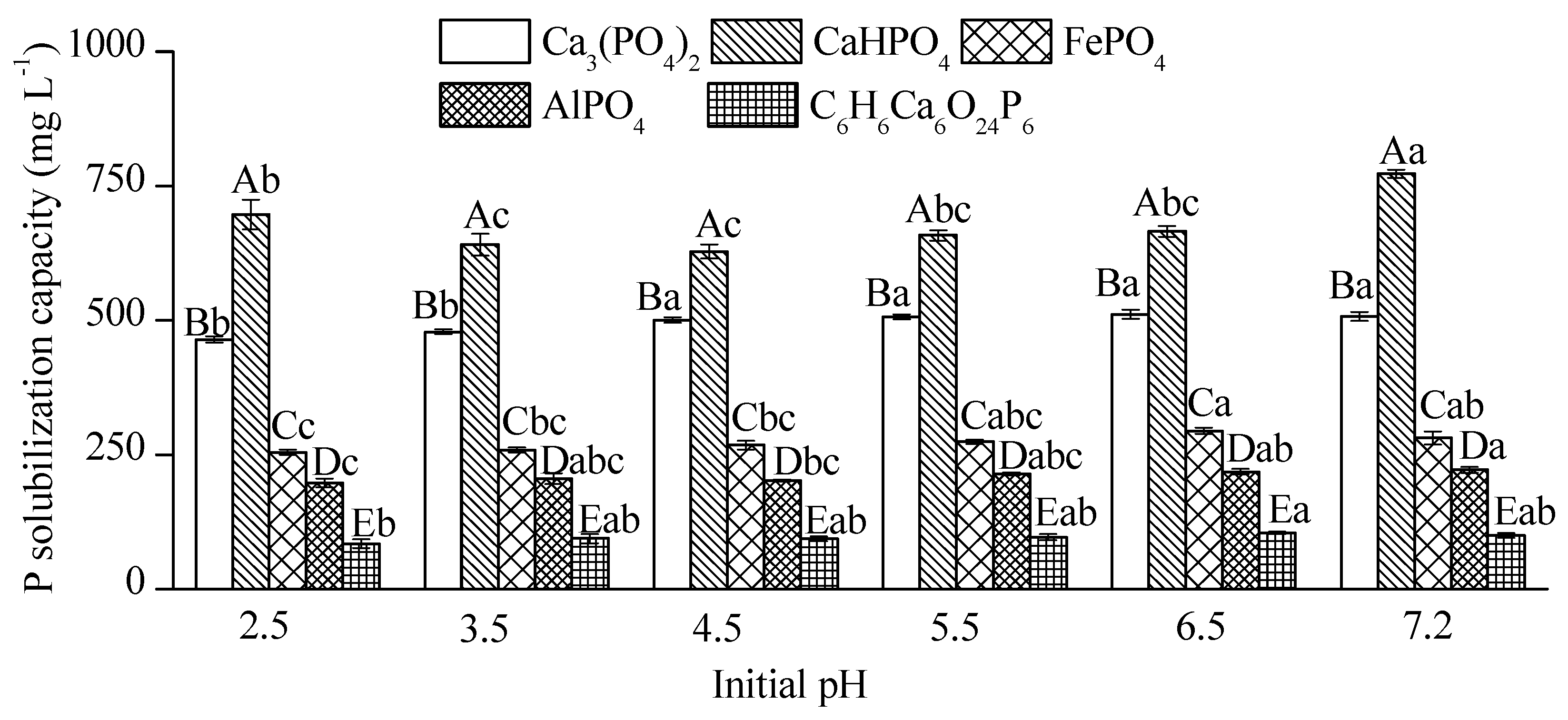
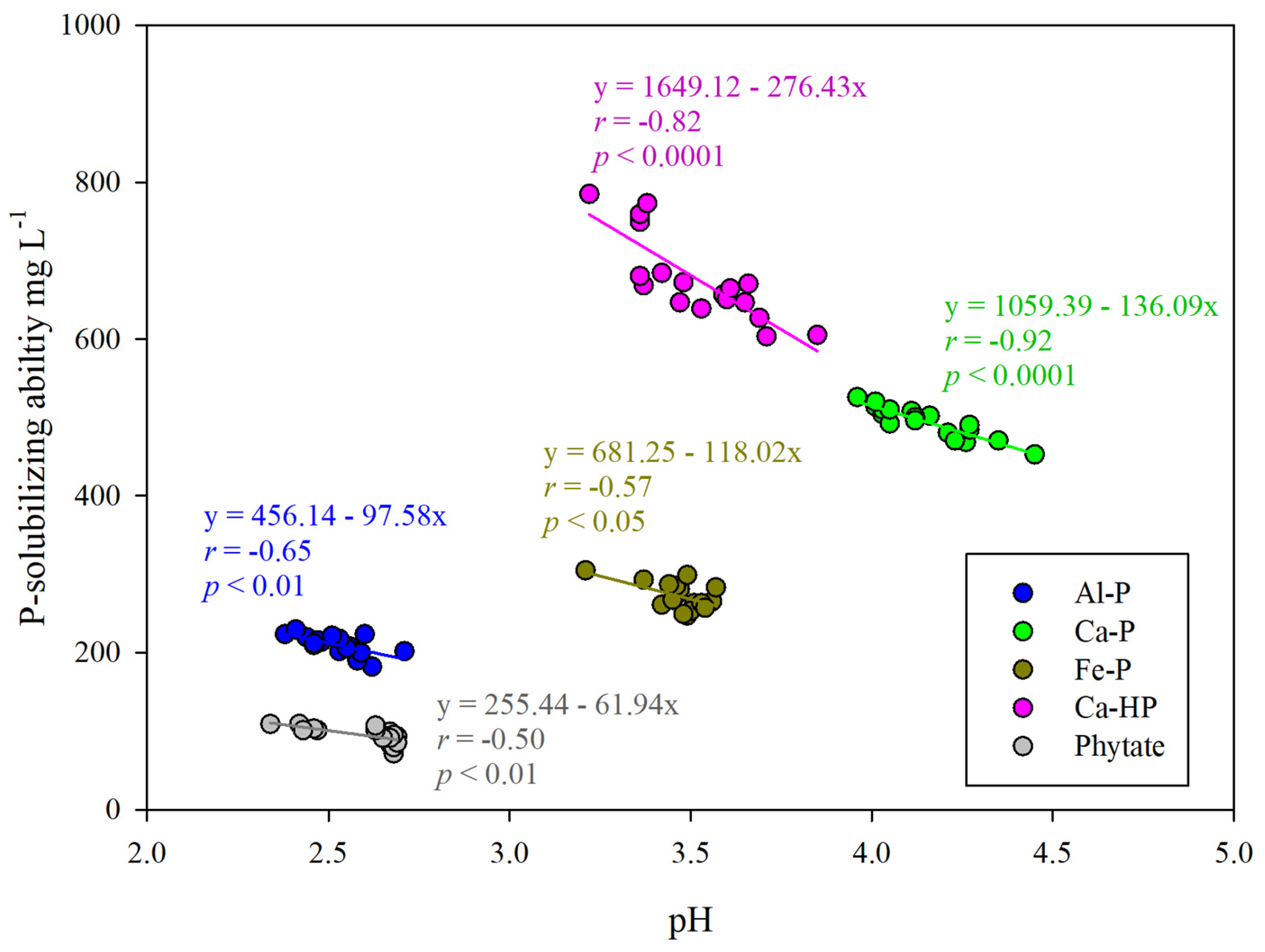
3.2. Effects of Environmental Factors on Phosphate-Solubilizing Activity of RW37
3.3. Characterization of IAA Production
3.4. Effect of Inoculation on Growth and Nutrient Content of Moso Bamboo Seedlings
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, F.; Chen, F.S.; Wang, G.G.; Mao, R.; Fang, X.M.; Wang, H.M.; Bu, W.S. Effects of experimental nitrogen addition on nutrients and nonstructural carbohydrates of dominant understory plants in a Chinese fir plantation. Forests 2019, 10, 155. [Google Scholar] [CrossRef]
- Shen, J.B.; Yuan, L.X.; Zhang, J.L.; Li, H.G.; Bai, Z.H.; Chen, X.P.; Zhang, W.F.; Zhang, F.S. Phosphorus dynamics: From soil to plant. Plant Physiol. 2011, 156, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, F.S.; Wu, X.Q.; Luan, F.G.; Zhang, L.P.; Fang, X.M.; Wan, S.Z.; Hu, X.F.; Ye, J.R. Isolation and characterization of two phosphate-solubilizing fungi from rhizosphere soil of moso bamboo and their functional capacities when exposed to different phosphorus sources and pH environments. PLoS ONE 2018, 13, e0199625. [Google Scholar] [CrossRef] [PubMed]
- Rani, R.; Usmani, Z.; Gupta, P.; Chandra, A.; Das, A.; Kumar, V. Effects of organochlorine pesticides on plant growth-promoting traits of phosphate-solubilizing rhizobacterium, Paenibacillus sp. IITISM08. Environ. Sci. Pollut. Res. 2018, 25, 5668–5680. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Xu, Z.; Guo, Z.; Jia, Z.; Wang, S.; Kai, D. Response of soil microbes to a reduction in phosphorus fertilizer in rice-wheat rotation paddy soils with varying soil P levels. Soil Till Res. 2018, 181, 127–135. [Google Scholar]
- Bornø, M.L.; Eduah, J.O.; Müller-Stöver, D.S.; Liu, F. Effect of different biochars on phosphorus (P) dynamics in the rhizosphere of Zea mays L. (maize). Plant Soil 2018, 431, 257–272. [Google Scholar] [CrossRef]
- Li, H.B.; Singh, R.K.; Singh, P.; Song, Q.Q.; Xing, Y.X.; Yang, L.T.; Li, Y.R. Genetic diversity of nitrogen-fixing and plant growth promoting Pseudomonas species isolated from Sugarcane Rhizosphere. Front. Microbiol. 2017, 1, 1268. [Google Scholar] [CrossRef]
- Meyer, G.; Maurhofer, M.; Frossard, E.; Gamper, H.A.; Mäder, P.; Mészáros, É.; Schönholzer-Mauclaire, L.; Symanczik, S.; Oberson, A. Pseudomonas protegens CHA0 does not increase phosphorus uptake from 33P labeled synthetic hydroxyapatite by wheat grown on calcareous soil. Soil Biol. Biochem. 2019, 131, 217–228. [Google Scholar] [CrossRef]
- Richardson, A.E.; Hocking, P.J.; Simpson, R.J.; George, T.S.; Cornish, P.S. Plant mechanisms to optimise access to soil phosphorus. Crop Pasture Sci. 2009, 60, 124–143. [Google Scholar] [CrossRef]
- Whitelaw, M.A. Growth promotion of plants inoculated with phosphate-solubilizing fungi. Adv. Agron. 1999, 69, 99–151. [Google Scholar]
- Shahid, M.; Hameed, S.; Imran, A.; Ali, S.; van Elsas, J. Root colonization and growth promotion of sunflower (Helianthus annuus L.) by phosphate solubilizing Enterobacter sp. Fs-11. World J. Microb. Biot. 2012, 28, 2749–2758. [Google Scholar] [CrossRef] [PubMed]
- Shankar, M.; Ponraj, P.; Ilakkiam, D.; Gunasekaran, P. Root colonization of a rice growth promoting strain of Enterobacter cloacae. J. Basic Microb. 2011, 51, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Xia, M.; Guan, F.; Fan, S. Spatial distribution of soil nitrogen, phosphorus and potassium stocks in moso bamboo forests in subtropical China. Forests 2016, 7, 267. [Google Scholar] [CrossRef]
- Zhang, X.; Zhong, Z.; Gai, X.; Ying, J.; Li, W.; Du, X.; Bian, F.; Yang, C. Leaf-associated shifts in bacterial and fungal communities in response to chicken rearing under moso bamboo forests in subtropical China. Forests 2019, 10, 216. [Google Scholar] [CrossRef]
- Hogarth, N.J.; Belcher, B. The contribution of bamboo to household income and rural livelihoods in a poor and mountainous county in Guangxi, China. Int. Forest Rev. 2013, 15, 71–81. [Google Scholar] [CrossRef]
- Kant, S.; Chiu, M. Bamboo sector reforms and the local economy of Linan county, Zhejiang province, People’s Republic of China. Forest Policy Econ. 2000, 1, 283–299. [Google Scholar] [CrossRef]
- Guo, X.M.; Niu, D.K.; Chen, F.; Zhang, W.Y. Balanced Fertilization and Nutrient Management for Bamboo Forest; Science Press: Beijing, China, 2013; pp. 27–36. [Google Scholar]
- Pikovskaya, R.I. Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Microbiologiya 1948, 17, 362–370. [Google Scholar]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef]
- Krieg, N.R. Bergey’s Manual of Systematic Bacteriology. In Bergey’s Manual of Systematic Bacteriology; Springer: Berlin/Heidelberg, Germany, 1986. [Google Scholar]
- Biswas, J.K.; Banerjee, A.; Rai, M.; Naidu, R.; Biswas, B.; Vithanage, M.; Dash, M.C.; Sarkar, S.K.; Meers, E. Potential application of selected metal resistant phosphate solubilizing bacteria isolated from the gut of earthworm (Metaphire posthuma) in plant growth promotion. Geoderma 2018, 330, 117–124. [Google Scholar] [CrossRef]
- Byers, H.K.; Stackebrandt, E.; Hayward, C.; Blackall, L.L. Molecular investigation of a microbial mat associated with the Great Artesian Basin. FEMS Microbiol. Ecol. 1998, 25, 391–403. [Google Scholar] [CrossRef]
- Liu, H.; Wu, X.Q.; Ren, J.H.; Ye, J.R. Isolation and identification of phosphobacteria in poplar rhizosphere from different regions of China. Pedosphere 2011, 21, 90–97. [Google Scholar] [CrossRef]
- Tamura, K.; Filipski, A.; Peterson, D.; Stecher, G.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Olsen, S.R.; Sommers, L.E. Phosphorus. In Methods of Soil Analysis; Page, A.L., Miller, R.H., Dennis, R.K., Eds.; American Society of Agronomy: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- Glickmann, E.; Dessaux, Y. A critical examination of the specificity of the salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [CrossRef]
- García-López, A.M.; Recena, R.; Avilés, M.; Delgado, A. Effect of Bacillus subtilis QST713 and Trichoderma asperellum T34 on P uptake by wheat and how it is modulated by soil properties. J. Soils Sediments 2018, 18, 727–738. [Google Scholar] [CrossRef]
- Allen, S.E. Chemical Analysis of Ecological Materials; Blackwell Scientific Publications: Oxford, UK, 1989. [Google Scholar]
- Li, F.; Zi, Q.; Sonne, H.Y.C.; Li, X.G. Microbiome sustains forest ecosystem functions across hierarchical scales. EEH 2023, 2, 24–31. [Google Scholar] [CrossRef]
- Zhu, J.; Li, M.; Whelan, M. Phosphorus activators contribute to legacy phosphorus availability in agricultural soils: A review. Sci. Total Environ. 2018, 612, 522–537. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Sheng, K.Y.; Zheng, Q.Y.; Hu, D.N.; Zhang, L.; Wang, J.W.; Zhang, W.Y. Inoculation with phosphate-solubilizing bacteria alters microbial community and activates soil phosphorus supply to promote maize growth. Land Degrad Dev. 2022, 34, 777–788. [Google Scholar] [CrossRef]
- Adhikari, A.; Lee, K.E.; Khan, M.A.; Kang, S.M.; Adhikari, B.; Imran, M.; Jan, R.; Kim, K.M.; Lee, I.J. Effect of silicate and phosphate solubilizing rhizobacterium Enterobacter ludwigii GAK2 on Oryza sativa L. under cadmium stress. J. Microbiol. Biotech. 2020, 30, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Barra, P.J.; Pontigo, S.; Delgado, M.; Parra–Almuna, L.; Duran, P.; Valentine, A.J.; Jorquera, M.A.; Mora, M.L. Phosphobacteria inoculation enhances the benefit of P-fertilization on Lolium perenne in soils contrasting in P-availabilit. Soil Biol. Biochem. 2019, 136, 107516. [Google Scholar] [CrossRef]
- Liu, F.P.; Liu, H.Q.; Zhou, H.L.; Dong, Z.G.; Bai, X.H.; Bai, P.; Qiao, J.J. Isolation and characterization of phosphate-solubilizing bacteria from betel nut (Areca catechu) and their effects on plant growth and phosphorus mobilization in tropical soils. Biol. Fert. Soils 2014, 50, 927–937. [Google Scholar] [CrossRef]
- Albdaiwi, R.N.; Khyami-Horani, H.; Ayad, J.Y.; Alananbeh, K.M.; Al-Sayaydeh, R. Isolation and characterization of halotolerant plant growth promoting rhizobacteria from durum wheat (Triticum turgidum subsp. durum) cultivated in saline areas of the dead sea region. Front. Microbiol. 2019, 10, 1639. [Google Scholar] [CrossRef]
- Yang, L.; Du, L.L.; Li, W.J.; Wang, R.; Guo, S.L. Divergent responses of phoD- and pqqC-harbouring bacterial communitie across soil aggregates to long fertilization practices. Soil Till. Res. 2023, 228, 105634. [Google Scholar] [CrossRef]
- Mendes, G.D.O.; Freitas, A.L.M.; Pereira, O.L.; Silva, I.R.; Vassilev, N.B.; Costa, M.D. Mechanisms of phosphate solubilization by fungal isolates when exposed to different P sources. Ann. Microbiol. 2014, 64, 239–249. [Google Scholar] [CrossRef]
- Amarasinghe, T.; Munaweera, I.; Kottegoda, N. Review on mechanisms of phosphate solubilization in rock phosphate fertilizer. Commun. Soil Sci. Plant Anal. 2022, 53, 944–960. [Google Scholar] [CrossRef]
- Li, X.; Luo, L.; Yang, J.; Li, B.; Yuan, H. Mechanisms for solubilization of various insoluble phosphates and activation of immobilized phosphates in different soils by an efficient and salinity-tolerant Aspergillus niger strain An2. Appl. Biochem. Biotech. 2015, 175, 2755–2768. [Google Scholar] [CrossRef]
- Kang, X.; Csetenyi, L.; Gadd, G.M. Colonization and bioweathering of monazite by Aspergillus niger: Solubilization and precipitation of rare earth elements. Environ. Microbiol. 2021, 23, 3970–3986. [Google Scholar] [CrossRef]
- Song, X.; Chen, X.; Zhou, G.; Jiang, H.; Peng, C. Observed high and persistent carbon uptake by moso bamboo forests and its response to environmental drivers. Agr. Forest Meteoro. 2017, 247, 467–475. [Google Scholar] [CrossRef]
- Yu, L.Y.; Huang, H.B.; Wang, X.H.; Li, S.; Feng, N.X.; Zhao, H.M.; Huang, X.P.; Li, Y.W.; Li, H.; Cai, Q.Y.; et al. Novel phosphate-solubilising bacteria isolated from sewage sludge and the mechanism of phosphate solubilisation. Sci. Total Environ. 2019, 658, 474–484. [Google Scholar] [CrossRef]
- Wan, W.; He, D.; Xue, Z. Removal of nitrogen and phosphorus by heterotrophic nitrification-aerobic denitrification of a denitrifying phosphorus-accumulating bacterium Enterobacter cloacae HW-15. Ecol. Eng. 2017, 99, 199–208. [Google Scholar] [CrossRef]
- Gómez-Muñoz, B.; Pittroff, S.M.; Neergaard, A.D.; Jensen, L.S.; Nicolaisen, M.H.; Magid, J. Penicillium bilaii effects on maize growth and P uptake from soil and localized sewage sludge in a rhizobox experiment. Biol. Fert. Soils 2017, 53, 23–35. [Google Scholar] [CrossRef]
- Moore, J.A.M.; Abraham, P.E.; Michener, J.K.; Muchero, W.; Cregger, M.A. Ecosystem consequences of introducing plant growth promoting rhizobacteria to managed systems and potential legacy effects. New Phytol. 2022, 234, 1914–1918. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.N.; Shi, L.; Wang, N.; Li, Q.N.; Zhang, L.S.; Han, N.; Yan, T.; Ren, D.Y.; Zhang, B.; Zhao, Y.; et al. Gut bacterial indole-3-acetic acid induced immune promotion mediates preventive effects of Fu Brick tea polyphenols on experimental colitis. J. Agr. Food Chem. 2023, 71, 1201–1213. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Sites | Elevation (m) | pH | Total N (g·kg−1) | Total P (g·kg−1) |
|---|---|---|---|---|
| Dagang | 381.0 | 5.16 ± 0.09 b | 4.65 ± 0.10 c | 0.27 ± 0.03 a |
| Guanshan | 548.0 | 4.37 ± 0.07 a | 2.51 ± 0.09 a | 0.33 ± 0.04 a |
| Dajing | 1105.0 | 4.39 ± 0.06 a | 3.44 ± 0.08 b | 0.54 ± 0.07 b |
| Biochemical Reactions | RW37 |
|---|---|
| 3% KOH solubility | + |
| Contact enzyme | + |
| Oxidase | − |
| Gelatin hydrolysis | + |
| Starch hydrolysis | + |
| Citrate | + |
| Indole production | − |
| Methyl red | + |
| Nitrate reduction | + |
| Malonate utilize | + |
| V-P determination | − |
| Gram staining | G− |
| Aerobism | aerobic |
| Bacterial form | rod shaped |
| Factors | RW37 | |
|---|---|---|
| Soluble P | Liquid pH | |
| Initial pH | 14.74 *** | 4.15 ** |
| P sources | 3900.47 *** | 2296.88 *** |
| Initial pH * P sources | 6.64 *** | 5.28 *** |
| Organism | Tryptophan (mg L−1) | IAA Production (µg mL−1) | |
|---|---|---|---|
| 6 Days | 12 Days | ||
| RW37 | 2 | 14.87 ± 1.15 b | 26.39 ± 2.44 a |
| 5 | 19.18 ± 1.56 b | 38.64 ± 3.19 a | |
| Item | Observed Variable | Control | Adding Strain RW37 | Increase Rate (%) |
|---|---|---|---|---|
| Plant growth | Seedling height/cm | 23.95 ± 0.71 b | 37.48 ± 1.06 a | 56.49% |
| Ground diameter/cm | 1.57 ± 0.10 b | 2.24 ± 0.08 a | 42.67% | |
| Aboveground biomass/g | 2.48 ± 0.13 b | 4.22 ± 0.16 a | 70.16% | |
| Underground biomass/g | 1.25 ± 0.06 b | 1.83 ± 0.09 a | 46.4% | |
| Total biomass/g | 3.73 ± 0.21 b | 6.05 ± 0.27 a | 62.19% | |
| Plant nutrients | Leaf P/g kg−1 | 2.01 ± 0.11 b | 2.89 ± 0.13 a | 43.78% |
| Stem P/g kg−1 | 0.92 ± 0.06 b | 1.61 ± 0.22 a | 75% | |
| Root P/g kg−1 | 0.86 ± 0.06 b | 1.74 ± 0.05 a | 102.33% | |
| Rhizosphere soil | Available P/mg kg−1 | 2.29 ± 0.25 b | 6.24 ± 0.08 a | 172.48% |
| NH4+-N/mg kg−1 | 0.96 ± 0.09 a | 0.81 ± 0.10 a | - | |
| NO3−-N/mg kg−1 | 2.11 ± 0.02 b | 4.11 ± 0.05 a | 94.79% | |
| Mineral N/mg kg−1 | 3.20 ± 0.06 b | 4.93 ± 0.13 a | 54.06% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Wan, S.; Shi, F.; Fang, X.; Huang, C. Identification and Characterization of a Phosphate-Solubilizing Bacterium and Its Growth-Promoting Effect on Moso Bamboo Seedlings. Forests 2024, 15, 364. https://doi.org/10.3390/f15020364
Zhang Y, Wan S, Shi F, Fang X, Huang C. Identification and Characterization of a Phosphate-Solubilizing Bacterium and Its Growth-Promoting Effect on Moso Bamboo Seedlings. Forests. 2024; 15(2):364. https://doi.org/10.3390/f15020364
Chicago/Turabian StyleZhang, Yang, Songze Wan, Fuxi Shi, Xiangmin Fang, and Chao Huang. 2024. "Identification and Characterization of a Phosphate-Solubilizing Bacterium and Its Growth-Promoting Effect on Moso Bamboo Seedlings" Forests 15, no. 2: 364. https://doi.org/10.3390/f15020364