
Divergent Effects of Fertilizer Regimes on Taxonomic and Functional Compositions of Rhizosphere Bacteria and Fungi in Phoebe bournei Young Plantations Are Associated with Root Exudates
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Sampling Design
2.2. Soil and Root Sampling
2.3. Soil Physicochemical Properties Measurement
2.4. DNA Extraction and PCR Amplification
2.5. Collection, Separation, Extraction, and Measurement of Root Exudates
2.6. Statistical Analysis
3. Results
3.1. Soil Properties and Root Exudates
3.2. Summary of Sequencing Information
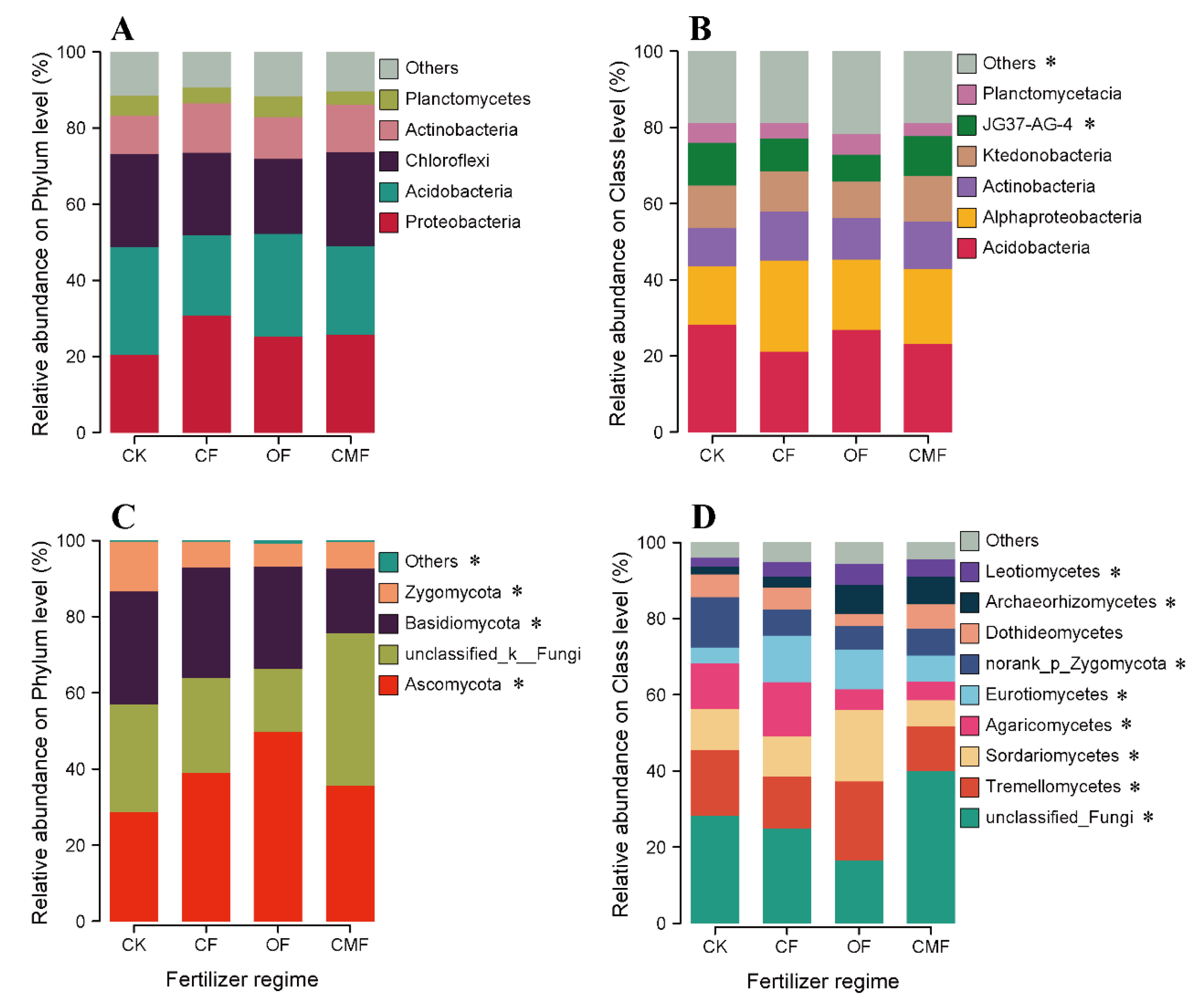
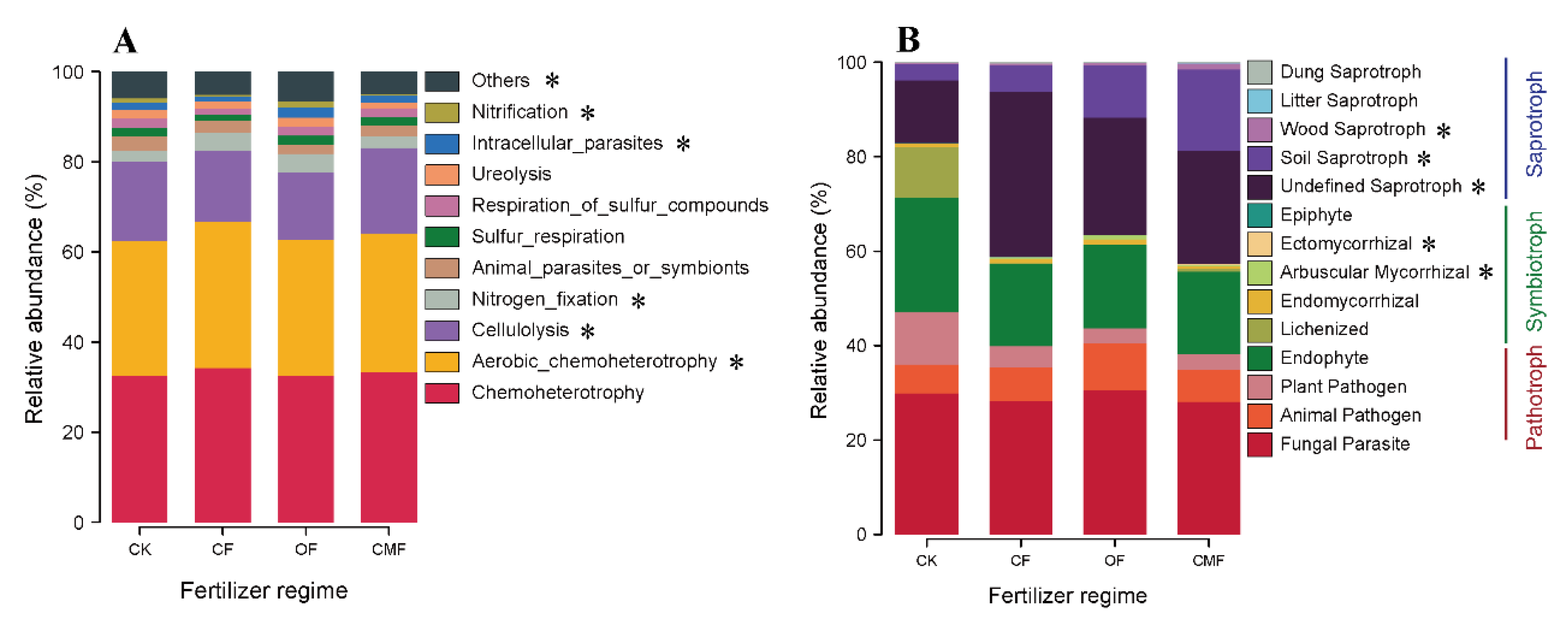
3.3. Relative Abundance of Taxonomic and Functional Compositions of Soil Bacteria and Fungi
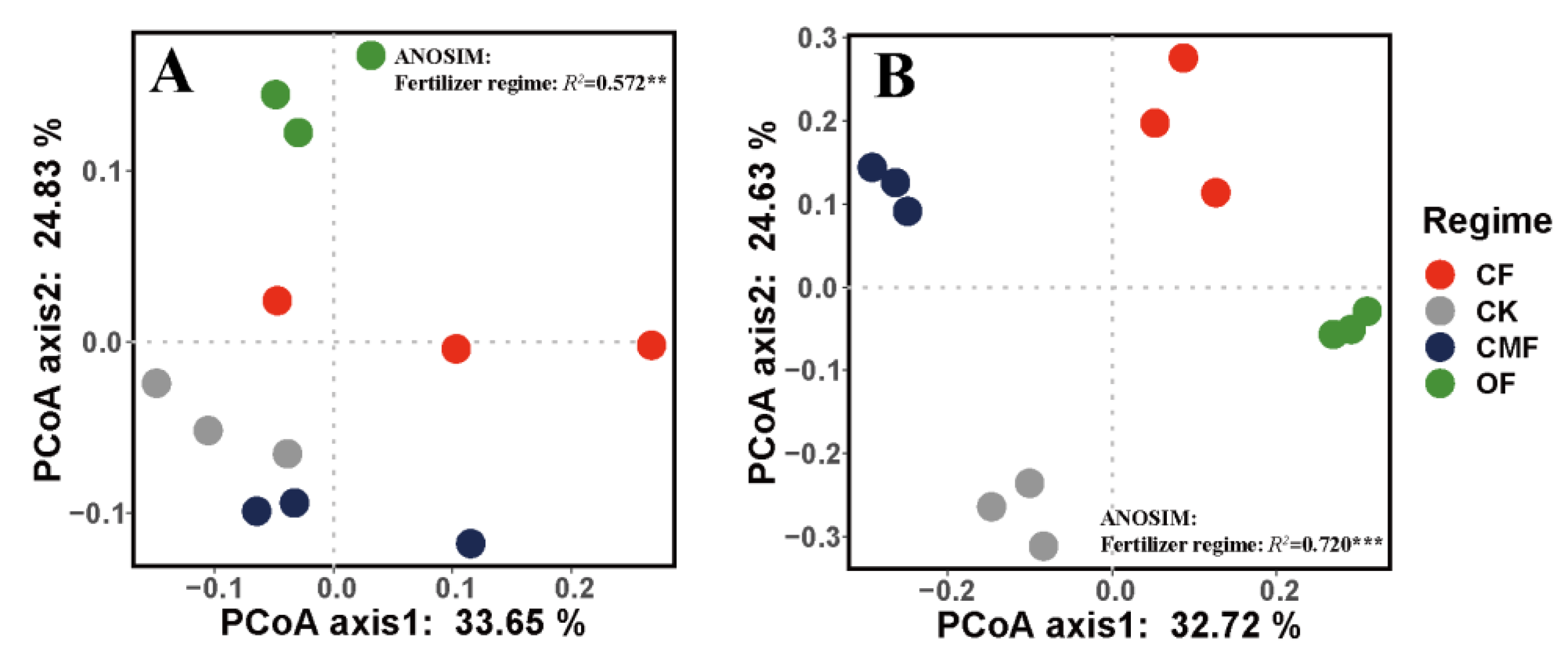
3.4. Soil Microbial α and β Diversity under Different Fertilizer Regimes
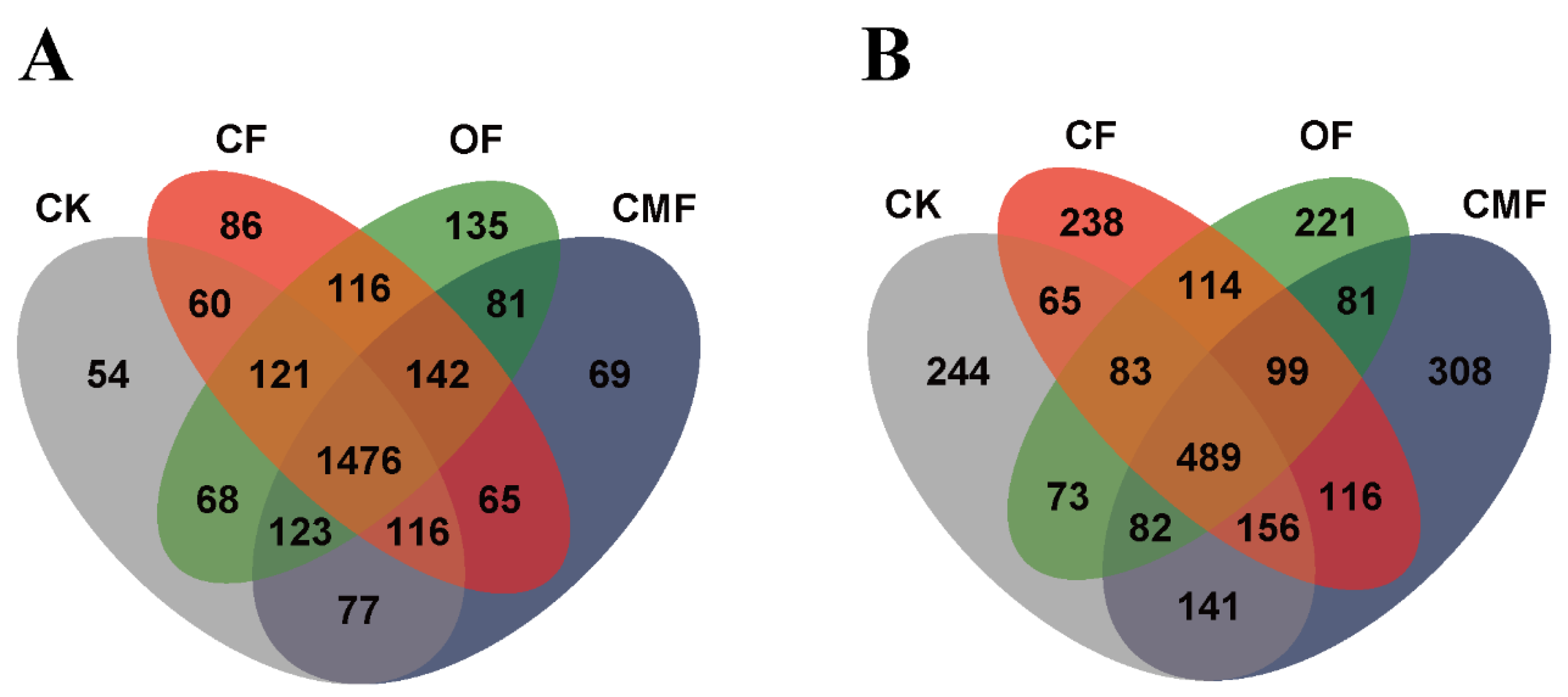
3.5. Shared and Unique OTUs
3.6. Main Driving Factors of Taxonomic and Functional Compositions of Soil Microbial Community
4. Discussion
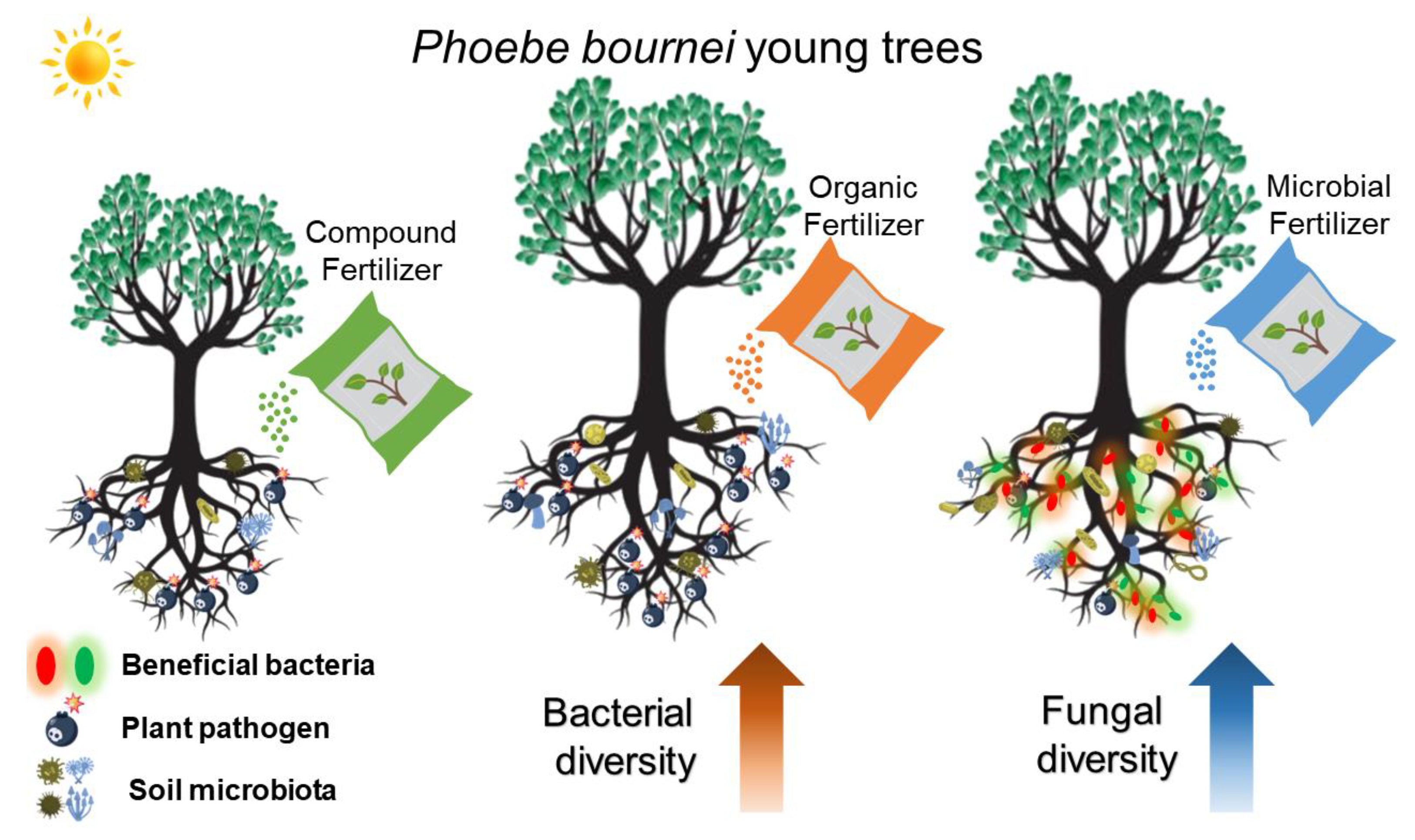
4.1. Soil Bacteria Were More Responsive to Organic Fertilizer, While Fungi Were More Sensitive to Compound Microbial Fertilizer
4.2. Root Exudates Had a Pronounced Association with Soil Microbial Community Than Soil Physiochemical Properties
4.3. Functional Composition of Soil Bacteria and Fungi Exhibited Higher Resolution Information Than the Taxonomic Level
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Albaugh, T.J.; Kiser, L.C.; Fox, T.R.; Allen, H.L.; Rubilar, R.A.; Stape, J.L. Ecosystem nutrient retention after fertilization of Pinus taeda. For. Sci. 2014, 60, 1131–1139. [Google Scholar] [CrossRef] [Green Version]
- Consalter, R.; Barbosa, J.Z.; Prior, S.A.; Vezzani, F.M.; Bassaco, M.V.M.; Pedreira, G.Q.; Motta, A.C.V. Mid-rotation fertilization and liming effects on nutrient dynamics of Pinus taedal in subtropical brazil. Eur. J. For. Res. 2021, 140, 19–35. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen–phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.H.; Liu, X.J.; Zhang, Y.; Shen, J.; Han, W.; Zhang, W.; Christie, P.; Goulding, K.; Vitousek, P.; Zhang, F. Significant acidification in major Chinese croplands. Science 2010, 327, 1008–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, L.E.; Siciliano, G. A comprehensive review of constraints to improved management of fertilizers in China and mitigation of diffuse water pollution from agriculture. Agric. Ecosyst. Environ. 2015, 209, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.; Dsouza, M.; Gilbert, J.A.; Guo, X.; Wang, D.; Guo, Z.; Ni, Y.; Chu, H. Fungal community composition in soils subjected to long-term chemical fertilization is most influenced by the type of organic matter. Environ. Microbiol. 2016, 18, 5137–5150. [Google Scholar] [CrossRef]
- Sradnick, A.; Murugan, R.; Oltmanns, M.; Raupp, J.; Joergensen, R.G. Changes in functional diversity of the soil microbial community in a heterogeneous sandy soil after long-term fertilization with cattle manure and mineral fertilizer. Appl. Soil Ecol. 2013, 63, 23–28. [Google Scholar] [CrossRef]
- Jannoura, R.; Joergensen, R.G.; Bruns, C. Organic fertilizer effects on growth, crop yield, and soil microbial biomass indices in sole and intercropped peas and oats under organic farming conditions. Eur. J. Agron. 2014, 52, 259–270. [Google Scholar] [CrossRef]
- Ren, J.; Liu, X.; Yang, W.; Yang, X.; Li, W.; Xia, Q.; Li, J.; Gao, Z.; Yang, Z. Rhizosphere soil properties, microbial community, and enzyme activities: Short-term responses to partial substitution of chemical fertilizer with organic manure. J. Environ. Manag. 2021, 299, 113650. [Google Scholar] [CrossRef]
- Zhong, W.; Gu, T.; Wang, W.; Zhang, B.; Lin, X.; Huang, Q.; Shen, W. The effects of mineral fertilizer and organic manure on soil microbial community and diversity. Plant Soil 2010, 326, 511–522. [Google Scholar] [CrossRef]
- Mahanty, T.; Bhattacharjee, S.; Goswami, M.; Bhattacharyya, P.; Das, B.; Ghosh, A.; Tribedi, P. Biofertilizers: A potential approach for sustainable agriculture development. Environ. Sci. Pollut. Res. 2017, 24, 3315–3335. [Google Scholar] [CrossRef] [PubMed]
- Van Der Heijden, M.G.; Bardgett, R.D.; Van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.K.; Trivedi, P.; Egidi, E.; Macdonald, C.A.; Delgado-Baquerizo, M. Crop microbiome and sustainable agriculture. Nat. Rev. Microbiol. 2021, 19, 72. [Google Scholar] [CrossRef]
- Feng, M.; Adams, J.M.; Fan, K.; Shi, Y.; Sun, R.; Wang, D.; Guo, X.; Chu, H. Long-term fertilization influences community assembly processes of soil diazotrophs. Soil Biol. Biochem. 2018, 126, 151–158. [Google Scholar] [CrossRef]
- Hu, J.; Lin, X.; Wang, J.; Dai, J.; Chen, R.; Zhang, J.; Wong, M.H. Microbial functional diversity, metabolic quotient, and invertase activity of a sandy loam soil as affected by long-term application of organic amendment and mineral fertilizer. J. Soils Sed. 2011, 11, 271–280. [Google Scholar] [CrossRef]
- Suzuki, C.; Nagaoka, K.; Shimada, A.; Takenaka, M. Bacterial communities are more dependent on soil type than fertilizer type, but the reverse is true for fungal communities. Soil Sci. Plant Nutr. 2009, 55, 80–90. [Google Scholar] [CrossRef] [Green Version]
- Lazcano, C.; Gómez-Brandón, M.; Revilla, P.; Domínguez, J. Short-term effects of organic and inorganic fertilizers on soil microbial community structure and function. Biol. Fertil. Soils 2013, 49, 723–733. [Google Scholar] [CrossRef]
- Pan, H.; Chen, M.; Feng, H.; Wei, M.; Song, F.; Lou, Y.; Cui, X.; Wang, H.; Zhuge, Y. Organic and inorganic fertilizers respectively drive bacterial and fungal community compositions in a fluvo-aquic soil in northern China. Soil Till. Res. 2020, 198, 104540. [Google Scholar] [CrossRef]
- Grosso, F.; Bååth, E.; De Nicola, F. Bacterial and fungal growth on different plant litter in mediterranean soils: Effects of C/N ratio and soil ph. Appl. Soil Ecol. 2016, 108, 1–7. [Google Scholar] [CrossRef]
- Mead, D.J. Opportunities for improving plantation productivity. How much? How quickly? How realistic? Biomass Bioenergy 2005, 28, 249–266. [Google Scholar] [CrossRef]
- Meidute, S.; Demoling, F.; Bååth, E. Antagonistic and synergistic effects of fungal and bacterial growth in soil after adding different carbon and nitrogen sources. Soil Biol. Biochem. 2008, 40, 2334–2343. [Google Scholar] [CrossRef]
- Zhao, S.; Li, K.; Zhou, W.; Qiu, S.; Huang, S.; He, P. Changes in soil microbial community, enzyme activities and organic matter fractions under long-term straw return in North-Central China. Agric. Ecosyst. Environ. 2016, 216, 82–88. [Google Scholar] [CrossRef]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef] [Green Version]
- Marschner, P.; Kandeler, E.; Marschner, B. Structure and function of the soil microbial community in a long-term fertilizer experiment. Soil Biol. Biochem. 2003, 35, 453–461. [Google Scholar] [CrossRef]
- Crecchio, C.; Curci, M.; Mininni, R.; Ricciuti, P.; Ruggiero, P. Short-term effects of municipal solid waste compost amendments on soil carbon and nitrogen content, some enzyme activities and genetic diversity. Biol. Fertil. Soils 2001, 34, 311–318. [Google Scholar] [CrossRef]
- Bittman, S.; Forge, T.; Kowalenko, C. Responses of the bacterial and fungal biomass in a grassland soil to multi-year applications of dairy manure slurry and fertilizer. Soil Biol. Biochem. 2005, 37, 613–623. [Google Scholar] [CrossRef]
- Banerjee, S.; Kirkby, C.A.; Schmutter, D.; Bissett, A.; Kirkegaard, J.A.; Richardson, A.E. Network analysis reveals functional redundancy and keystone taxa amongst bacterial and fungal communities during organic matter decomposition in an arable soil. Soil Biol. Biochem. 2016, 97, 188–198. [Google Scholar] [CrossRef]
- Li, H.; Bi, Q.; Yang, K.; bo Lasson, S.; Zheng, B.; Cui, L.; Zhu, Y.; Ding, K. High starter phosphorus fertilization facilitates soil phosphorus turnover by promoting microbial functional interaction in an arable soil. J. Environ. Sci. 2020, 94, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. Funguild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Tanunchai, B.; Ji, L.; Schroeter, S.A.; Wahdan, S.F.M.; Hossen, S.; Delelegn, Y.; Buscot, F.; Lehnert, A.-S.; Alves, E.G.; Hilke, I. Fungaltraits vs. Funguild: Comparison of ecological functional assignments of leaf-and needle-associated fungi across 12 temperate tree species. Microb. Ecol. 2022, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sansupa, C.; Wahdan, S.F.M.; Hossen, S.; Disayathanoowat, T.; Wubet, T.; Purahong, W. Can we use functional annotation of prokaryotic taxa (faprotax) to assign the ecological functions of soil bacteria? Appl. Sci. 2021, 11, 688. [Google Scholar] [CrossRef]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.; Wei, Z.; Wang, X.; Friman, V.-P.; Huang, J.; Wang, X.; Mei, X.; Xu, Y.; Shen, Q.; Jousset, A. Pathogen invasion indirectly changes the composition of soil microbiome via shifts in root exudation profile. Biol. Fertil. Soils 2016, 52, 997–1005. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Bakker, M.G.; Manter, D.K.; Sheflin, A.M.; Weir, T.L.; Vivanco, J.M. Harnessing the rhizosphere microbiome through plant breeding and agricultural management. Plant Soil 2012, 360, 1–13. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Sheflin, A.M.; Manter, D.K.; Vivanco, J.M. Manipulating the soil microbiome to increase soil health and plant fertility. Biol. Fertil. Soils 2012, 48, 489–499. [Google Scholar] [CrossRef]
- Zhao, M.; Zhao, J.; Yuan, J.; Hale, L.; Wen, T.; Huang, Q.; Vivanco, J.M.; Zhou, J.; Kowalchuk, G.A.; Shen, Q. Root exudates drive soil-microbe-nutrient feedbacks in response to plant growth. Plant Cell Environ. 2021, 44, 613–628. [Google Scholar] [CrossRef]
- Zahran, H.H. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiol. Mol. Biol. Rev. 1999, 63, 968–989. [Google Scholar] [CrossRef] [Green Version]
- Davidson, I.; Robson, M. Effect of contrasting patterns of nitrate application on the nitrate uptake, n2-fixation, nodulation and growth of white clover. Ann. Bot. 1986, 57, 331–338. [Google Scholar] [CrossRef]
- Wang, Z.; He, G.; Hou, Z.; Luo, Z.; Chen, S.; Lu, J.; Zhao, J. Soil c: N: P stoichiometry of typical coniferous (Cunninghamia Lanceolata) and/or evergreen broadleaved (Phoebe Bournei) plantations in south china. For. Ecol. Manag. 2021, 486, 118974. [Google Scholar] [CrossRef]
- Li, Y.; Liu, X.; Ma, J.; Zhang, X.; Xu, L. Phenotypic variation in Phoebe bournei populations preserved in the primary distribution area. J. For. Res. 2018, 29, 35–44. [Google Scholar] [CrossRef]
- Ji, L.; Tanunchai, B.; Wahdan, S.F.M.; Schaedler, M.; Purahong, W. Future climate change enhances the complexity of plastisphere microbial co-occurrence networks, but does not significantly affect the community assembly. Sci. Total Environ. 2022, 844, 157016. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magoč, T.; Salzberg, S.L. Flash: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Ji, L.; Shen, F.; Liu, Y.; Yang, Y.; Wang, J.; Purahong, W.; Yang, L. Contrasting altitudinal patterns and co-occurrence networks of soil bacterial and fungal communities along soil depths in the cold-temperate montane forests of China. Catena 2022, 209, 105844. [Google Scholar] [CrossRef]
- Li, T.; Wang, H.; Xia, X.; Cao, S.; Yao, J.; Zhang, L. Inhibitory effects of components from root exudates of welsh onion against root knot nematodes. PLoS ONE 2018, 13, e0201471. [Google Scholar] [CrossRef] [Green Version]
- Dai, Z.; Su, W.; Chen, H.; Barberán, A.; Zhao, H.; Yu, M.; Yu, L.; Brookes, P.C.; Schadt, C.W.; Chang, S.X.; et al. Long-term nitrogen fertilization decreases bacterial diversity and favors the growth of actinobacteria and proteobacteria in agro-ecosystems across the globe. Glob. Change Biol. 2018, 24, 3452–3461. [Google Scholar] [CrossRef]
- Acuna, J.J.; Marileo, L.G.; Araya, M.A.; Rilling, J.I.; Larama, G.A.; Mora, M.L.; Epstein, S.; Jorquera, M.A.J. In situ cultivation approach to increase the culturable bacterial diversity in the rhizobiome of plants. J. Soil Sci. Plant Nutr. 2020, 20, 1411–1426. [Google Scholar] [CrossRef]
- Trivedi, P.; Anderson, I.C.; Singh, B.K. Microbial modulators of soil carbon storage: Integrating genomic and metabolic knowledge for global prediction. Trends Microbiol. 2013, 21, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, Y.; Zhang, S.; Wei, W.; Kuzyakov, Y.; Ding, X. Fertilization effects on microbial community composition and aggregate formation in saline-alkaline soil. Plant Soil 2021, 463, 523–535. [Google Scholar] [CrossRef]
- Yao, T.; Chen, R.; Zhang, J.; Feng, Y.; Huang, M.; Lin, X. Divergent patterns of microbial community composition shift under two fertilization regimes revealed by responding species. Appl. Soil Ecol. 2020, 154, 103590. [Google Scholar] [CrossRef]
- Allison, S.D.; Martiny, J.B. Resistance, resilience, and redundancy in microbial communities. Proc. Natl. Acad. Sci. USA 2008, 105, 11512–11519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naveed, M.; Herath, L.; Moldrup, P.; Arthur, E.; Nicolaisen, M.; Norgaard, T.; Ferré, T.P.; de Jonge, L.W. Spatial variability of microbial richness and diversity and relationships with soil organic carbon, texture and structure across an agricultural field. Appl. Soil Ecol. 2016, 103, 44–55. [Google Scholar] [CrossRef]
- Wohl, D.L.; Arora, S.; Gladstone, J.R. Functional redundancy supports biodiversity and ecosystem function in a closed and constant environment. Ecology 2004, 85, 1534–1540. [Google Scholar] [CrossRef]
- Yang, W.; Guo, Y.; Wang, X.; Chen, C.; Hu, Y.; Cheng, L.; Gu, S.; Xu, X. Temporal variations of soil microbial community under compost addition in black soil of northeast China. Appl. Soil Ecol. 2017, 121, 214–222. [Google Scholar] [CrossRef]
- Högberg, M.N.; Högberg, P.; Myrold, D.D. Is microbial community composition in boreal forest soils determined by pH, C-to-N ratio, the trees, or all three? Oecologia 2007, 150, 590–601. [Google Scholar] [CrossRef]
- Ji, L.; Yang, Y.; Yang, L. Seasonal variations in soil fungal communities and co-occurrence networks along an altitudinal gradient in the cold temperate zone of China: A case study on Oakley Mountain. Catena 2021, 204, 105448. [Google Scholar] [CrossRef]
- Robson, A.; Abbott, L. The effect of soil acidity on microbial activity in soils. In Soil Acidity and Plant Growth; Elsevier: Amsterdam, The Netherlands, 1989; pp. 139–165. [Google Scholar]
- Brar, B.S.; Singh, J.; Singh, G.; Kaur, G. Effects of long term application of inorganic and organic fertilizers on soil organic carbon and physical properties in maize–wheat rotation. Agronomy 2015, 5, 220–238. [Google Scholar] [CrossRef] [Green Version]
- Stamford, N.P.; Felix, F.; Oliveira, W.; Silva, E.; Carolina, S.; Arnaud, T.; Freitas, A.D. Interactive effectiveness of microbial fertilizer enriched in n on lettuce growth and on characteristics of an ultisol of the rainforest region. Sci. Hortic. 2019, 247, 242–246. [Google Scholar] [CrossRef]
- Fasusi, O.A.; Cruz, C.; Babalola, O.O. Agricultural sustainability: Microbial biofertilizers in rhizosphere management. Agriculture 2021, 11, 163. [Google Scholar] [CrossRef]
- Kuhns, E.H.; Martini, X.; Tribuiani, Y.; Coy, M.; Gibbard, C.; Peña, J.; Hulcr, J.; Stelinski, L.L. Eucalyptol is an attractant of the redbay ambrosia beetle, Xyleborus glabratus. J. Chem. Ecol. 2014, 40, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Kendra, P.E.; Montgomery, W.S.; Deyrup, M.A.; Wakarchuk, D. Improved lure for redbay ambrosia beetle developed by enrichment of α-copaene content. J. Pest Sci. 2016, 89, 427–438. [Google Scholar] [CrossRef]
- Chuankun, X.; Minghe, M.; Leming, Z.; Keqin, Z. Soil volatile fungistasis and volatile fungistatic compounds. Soil Biol. Biochem. 2004, 36, 1997–2004. [Google Scholar] [CrossRef]
- Lin, C.; Owen, S.M.; Peñuelas, J. Volatile organic compounds in the roots and rhizosphere of Pinus spp. Soil Biol. Biochem. 2007, 39, 951–960. [Google Scholar] [CrossRef]
- Smolander, A.; Kanerva, S.; Adamczyk, B.; Kitunen, V. Nitrogen transformations in boreal forest soils—Does composition of plant secondary compounds give any explanations? Plant Soil 2012, 350, 1–26. [Google Scholar] [CrossRef]
- Chen, C.J.; Li, Q.Q.; Ma, Y.N.; Wang, W.; Cheng, Y.X.; Xu, F.R.; Dong, X. Antifungal effect of essential oils from five kinds of rutaceae plants–avoiding pesticide residue and resistance. Chem. Biodivers. 2019, 16, e1800688. [Google Scholar] [CrossRef]
- Santonja, M.; Bousquet-Mélou, A.; Greff, S.; Ormeño, E.; Fernandez, C. Allelopathic effects of volatile organic compounds released from pinus halepensis needles and roots. Ecol. Evol. 2019, 9, 8201–8213. [Google Scholar] [CrossRef]
- Sahebi, M.; Hanafi, M.M.; van Wijnen, A.J.; Akmar, A.S.N.; Azizi, P.; Idris, A.S.; Taheri, S.; Foroughi, M. Profiling secondary metabolites of plant defence mechanisms and oil palm in response to ganoderma boninense attack. Int. Biodeterior. Biodegrad. 2017, 122, 151–164. [Google Scholar] [CrossRef]
- Zhang, H.; Zheng, X.; Wang, X.; Xiang, W.; Xiao, M.; Wei, L.; Zhang, Y.; Song, K.; Zhao, Z.; Lv, W.; et al. Effect of fertilization regimes on continuous cropping growth constraints in watermelon is associated with abundance of key ecological clusters in the rhizosphere. Agric. Ecosyst. Environ. 2022, 339, 108135. [Google Scholar] [CrossRef]
- Chen, D.; Wang, X.; Zhang, W.; Zhou, Z.; Ding, C.; Liao, Y.; Li, X. Persistent organic fertilization reinforces soil-borne disease suppressiveness of rhizosphere bacterial community. Plant Soil 2020, 452, 313–328. [Google Scholar] [CrossRef]
- Zhao, S.; Liu, D.; Ling, N.; Chen, F.; Fang, W.; Shen, Q. Bio-organic fertilizer application significantly reduces the Fusarium oxysporum population and alters the composition of fungi communities of watermelon Fusarium wilt rhizosphere soil. Biol. Fertil. Soils 2014, 50, 765–774. [Google Scholar] [CrossRef]
- Liu, Z.; Ma, X.; He, N.; Zhang, J.; Wu, J.; Liu, C. Shifts in microbial communities and networks are correlated with the soil ionome in a kiwifruit orchard under different fertilization regimes. Appl. Soil Ecol. 2020, 149, 103517. [Google Scholar] [CrossRef]
- Raza, W.; Yuan, J.; Ling, N.; Huang, Q.; Shen, Q. Production of volatile organic compounds by an antagonistic strain paenibacillus polymyxa wr-2 in the presence of root exudates and organic fertilizer and their antifungal activity against Fusarium oxysporum f. sp. Niveum. Biol. Control 2015, 80, 89–95. [Google Scholar] [CrossRef]
- Sharma, P.; Thakur, D. Antimicrobial biosynthetic potential and diversity of culturable soil actinobacteria from forest ecosystems of northeast India. Sci. Rep. 2020, 10, 4104. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, J.; Rhodes, G.; He, J.-Z.; Ge, Y. Adaptive responses of comammox nitrospira and canonical ammonia oxidizers to long-term fertilizations: Implications for the relative contributions of different ammonia oxidizers to soil nitrogen cycling. Sci. Total Environ. 2019, 668, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Hu, H.-W.; Wang, J.; He, J.-Z.; Zheng, C.; Wan, X.; Huang, Z. Niche separation of comammox nitrospira and canonical ammonia oxidizers in an acidic subtropical forest soil under long-term nitrogen deposition. Soil Biol. Biochem. 2018, 126, 114–122. [Google Scholar] [CrossRef]
- Han, S.; Huang, Q.; Chen, W. Partitioning nitrospira community structure and co-occurrence patterns in a long-term inorganic and organic fertilization soil. J. Soils Sed. 2021, 21, 1099–1108. [Google Scholar] [CrossRef]
- Baldi, E.; Marcolini, G.; Quartieri, M.; Sorrenti, G.; Toselli, M. Effect of organic fertilization on nutrient concentration and accumulation in nectarine (Prunus persica var. Nucipersica) trees: The effect of rate of application. Sci. Hortic. 2014, 179, 174–179. [Google Scholar]
- Berthrong, S.T.; Yeager, C.M.; Gallegos-Graves, L.; Steven, B.; Eichorst, S.A.; Jackson, R.B.; Kuske, C.R. Nitrogen fertilization has a stronger effect on soil nitrogen-fixing bacterial communities than elevated atmospheric CO2. Appl. Environ. Microbiol. 2014, 80, 3103–3112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahapatra, S.; Yadav, R.; Ramakrishna, W. Bacillus subtilis impact on plant growth, soil health and environment: Dr. Jekyll and Mr. Hyde. J. Appl. Microbiol. 2022, 132, 3543–3562. [Google Scholar] [CrossRef] [PubMed]
- Seshadri, S.; Ignacimuthu, S.; Vadivelu, M.; Lakshminarasimhan, C. Inorganic Phosphate Solubilization by Two Insect Pathogenic Bacillus sp. In Proceedings of the First International Meeting on Microbial Phosphate Solubilization, Salamanca, Spain, 16–19 July 2002; Springer: Berlin/Heidelberg, Germany, 2007; pp. 351–355. [Google Scholar]
- Santoro, M.V.; Bogino, P.C.; Nocelli, N.; Cappellari, L.d.R.; Giordano, W.F.; Banchio, E. Analysis of plant growth-promoting effects of fluorescent pseudomonas strains isolated from mentha piperita rhizosphere and effects of their volatile organic compounds on essential oil composition. Front. Microbiol. 2016, 7, 1085. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Variables | CK | CF | OF | CMF | |
|---|---|---|---|---|---|
| Soil physiochemical property | pH | 4.09 ± 0.01 a | 4.04 ± 0.01 a | 4.06 ± 0.03 a | 4.06 ± 0.02 a |
| SOC (g·kg−1) | 12.52 ± 0.34 c | 20.96 ± 0.39 ab | 21.17 ± 0.54 a | 18.88 ± 0.54 b | |
| TN (g·kg−1) | 1.52 ± 0.01 b | 1.87 ± 0.02 b | 1.87 ± 0.02 b | 2.42 ± 0.18 a | |
| TP (g·kg−1) | 0.16 ± 0.01 c | 0.21 ± 0.01 ab | 0.20 ± 0.01 b | 0.23 ± 0.01 a | |
| TK (g·kg−1) | 6.50 ± 0.13 b | 8.23 ± 0.03 a | 8.23 ± 0.09 a | 8.53 ± 0.11 a | |
| AN (mg·kg−1) | 4.24 ± 0.08 c | 5.62 ± 0.04 ab | 5.51 ± 0.04 b | 5.78 ± 0.06 a | |
| AP (mg·kg−1) | 0.71 ± 0.01 c | 0.89 ± 0.02 b | 1.12 ± 0.04 a | 0.93 ± 0.01 b | |
| Root exudate | αCop (%) | 4.85 ± 0.16 b | 3.51 ± 0.09 c | 6.74 ± 0.28 a | 3.95 ± 0.06 c |
| Cal (%) | 22.60 ± 0.57 b | 40.34 ± 2.50 a | 29.45 ± 2.07 b | 42.15 ± 1.66 a | |
| dLim (%) | 5.41 ± 0.56 a | 1.77 ± 0.13 bc | 2.97 ± 0.15 b | 0.97 ± 0.17 c | |
| αFen (%) | 24.20 ± 0.03 a | 11.53 ± 0.60 c | 14.38 ± 0.06 b | 11.07 ± 0.79 c | |
| Gua (%) | 1.41 ± 0.14 c | 3.72 ± 0.21 a | 2.74 ± 0.15 b | 4.10 ± 0.11 a | |
| αSan (%) | 7.29 ± 0.82 b | 14.24 ± 0.77 a | 13.57 ± 1.04 a | 13.63 ± 1.20 a | |
| Ses (%) | 54.38 ± 3.85 b | 74.84 ± 0.55 a | 67.18 ± 1.84 a | 76.29 ± 0.68 a | |
| Mon (%) | 37.56 ± 2.98 a | 15.38 ± 0.70 bc | 22.06 ± 0.37 b | 14.86 ± 0.46 c | |
| Fur (%) | —— | 0.57 ± 0.32 a | 0.16 ± 0.16 a | 0.50 ± 0.30 a | |
| Alc (%) | 4.57 ± 0.85 a | 6.08 ± 0.16 a | 6.31 ± 0.59 a | 6.77 ± 0.50 a | |
| Est (%) | 3.40 ± 0.26 a | 2.21 ± 0.55 a | 4.29 ± 1.05 a | 1.40 ± 0.75 a | |
| Taxonomy | Parameter | CK | CF | OF | CMF |
|---|---|---|---|---|---|
| Bacteria | Richness | 1589.33 ± 16.59 b | 1646.33 ± 31.52 b | 1772 ± 24.95 a | 1605.33 ± 30.82 b |
| Shannon | 5.81 ± 0.02 b | 5.88 ± 0.09 ab | 6.09 ± 0.02 a | 5.83 ± 0.04 b | |
| Chao 1 | 2002.65 ± 66.87 a | 2084.03 ± 37.32 a | 2198.76 ± 51.27 a | 2065.84 ± 49.32 a | |
| Fungi | Richness | 851.33 ± 11.62 a | 864.33 ± 73.95 a | 819.67 ± 44.21 a | 949.33 ± 12.25 a |
| Shannon | 4.28 ± 0.14 a | 4.18 ± 0.26 a | 3.99 ± 0.19 a | 4.62 ± 0.03 a | |
| Chao 1 | 922.16 ± 15.06 a | 944.2 ± 79.97 a | 959.73 ± 53.92 a | 1020.2 ± 18.92 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Z.; Yang, X.; Li, J.; Wen, S.; Yang, L.; Ji, L.; He, G. Divergent Effects of Fertilizer Regimes on Taxonomic and Functional Compositions of Rhizosphere Bacteria and Fungi in Phoebe bournei Young Plantations Are Associated with Root Exudates. Forests 2023, 14, 126. https://doi.org/10.3390/f14010126
Luo Z, Yang X, Li J, Wen S, Yang L, Ji L, He G. Divergent Effects of Fertilizer Regimes on Taxonomic and Functional Compositions of Rhizosphere Bacteria and Fungi in Phoebe bournei Young Plantations Are Associated with Root Exudates. Forests. 2023; 14(1):126. https://doi.org/10.3390/f14010126
Chicago/Turabian StyleLuo, Zhong, Xisha Yang, Jing Li, Shizhi Wen, Lili Yang, Li Ji, and Gongxiu He. 2023. "Divergent Effects of Fertilizer Regimes on Taxonomic and Functional Compositions of Rhizosphere Bacteria and Fungi in Phoebe bournei Young Plantations Are Associated with Root Exudates" Forests 14, no. 1: 126. https://doi.org/10.3390/f14010126