


Dynamics of Enzyme Activities during the Decomposition of Castanopsis carlesii Leaf Litter in the Forest Canopy and Forest Floor in a Mid-Subtropical Area
 , and
, and 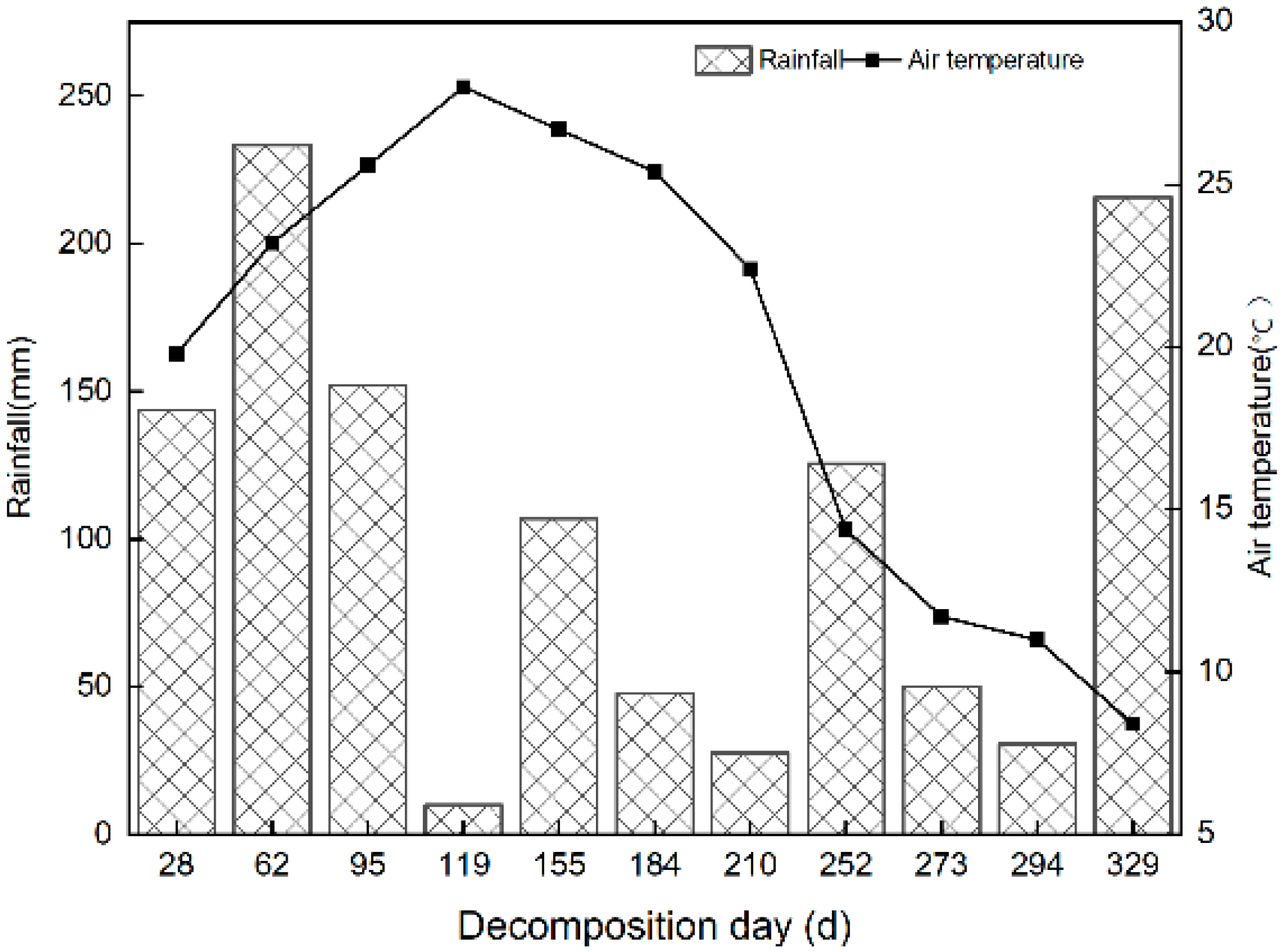{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Region
2.2. Experimental Design and Field Sampling
2.3. Measurements of Enzyme Activities
2.4. Analytical Methods and Calculations
2.5. Data Analysis
3. Results
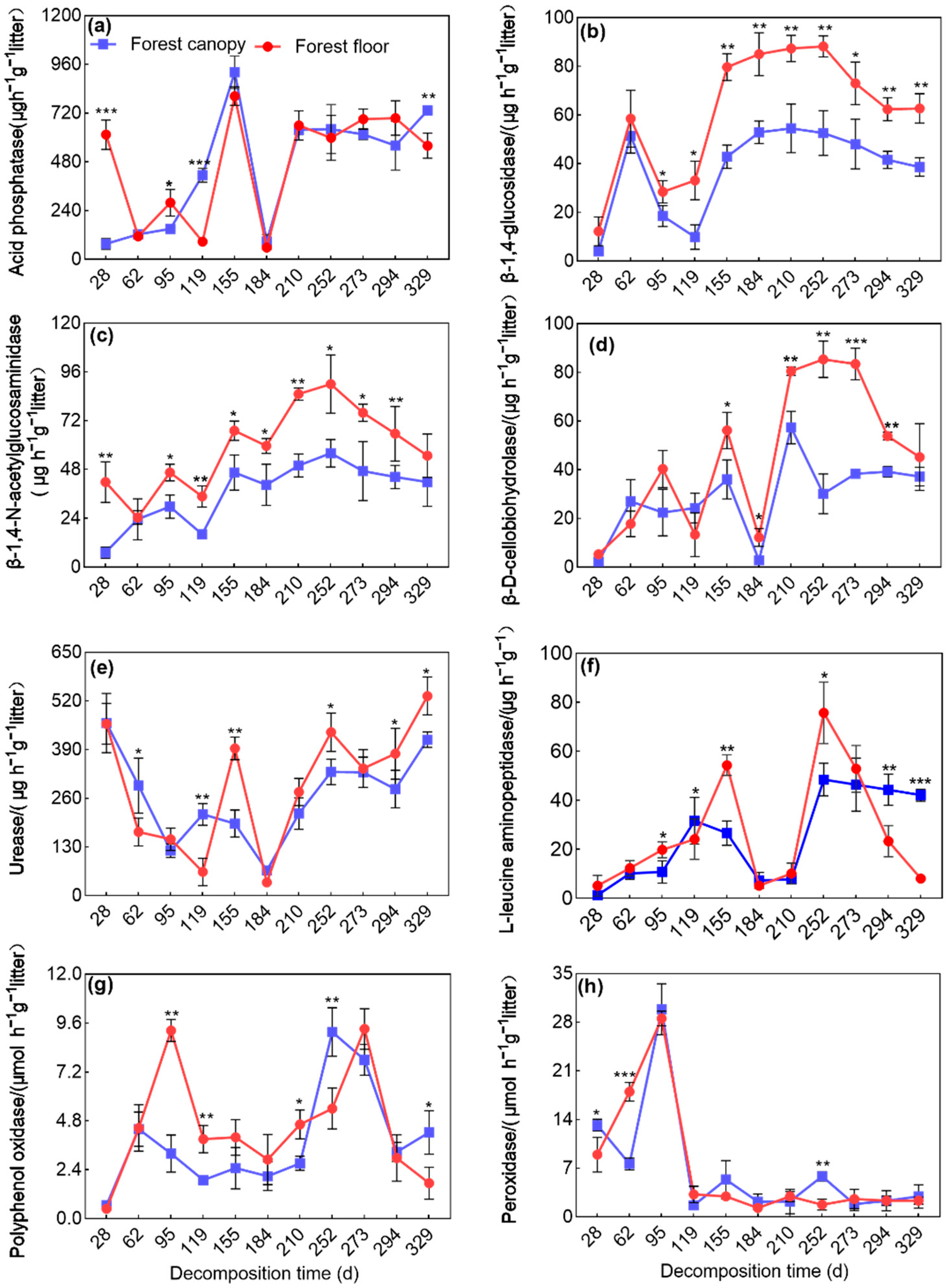
3.1. Dynamics of Enzyme Activities during Litter Decomposition
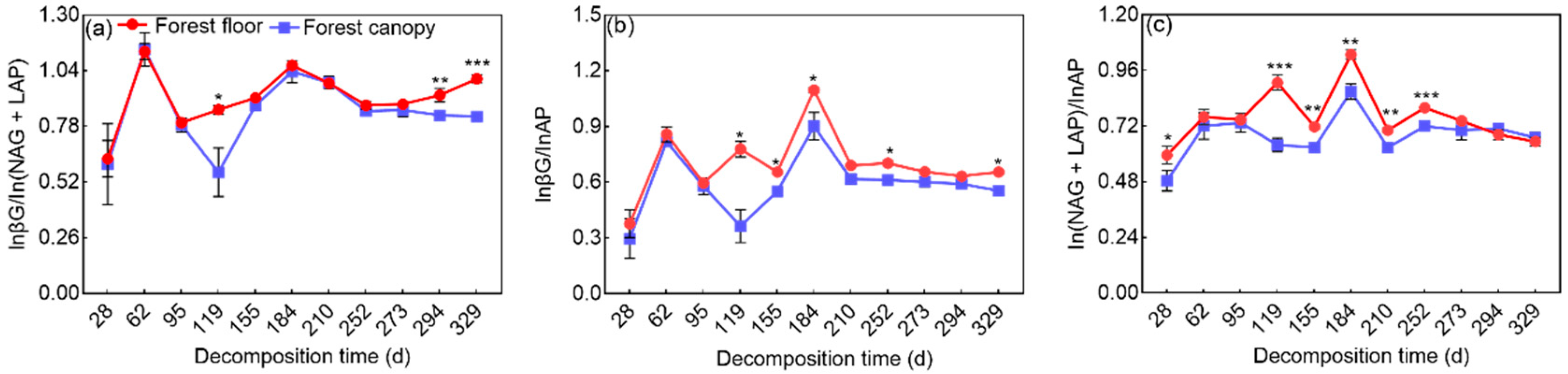
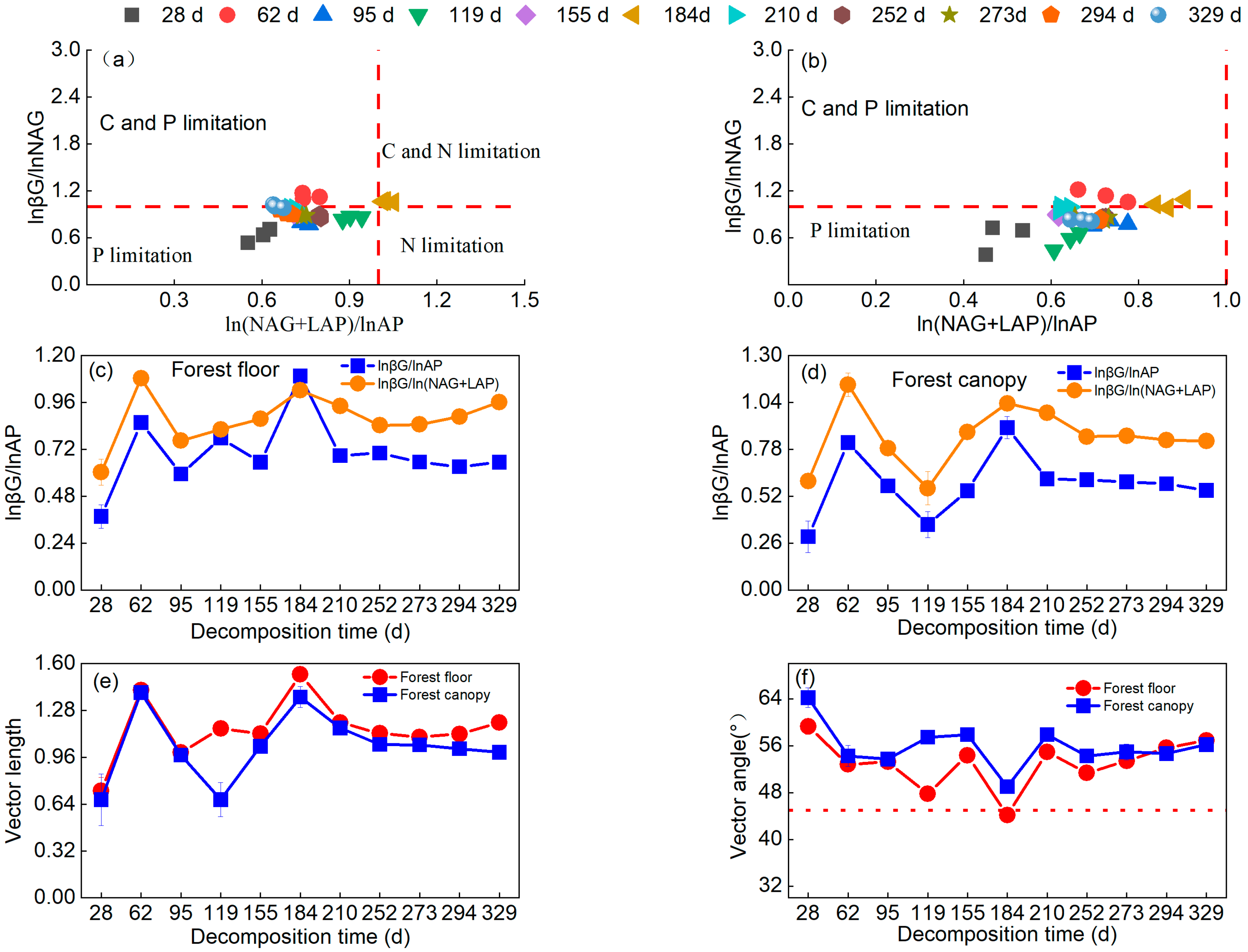
3.2. Indicators of Microbial Resource Limitation during Litter Decomposition
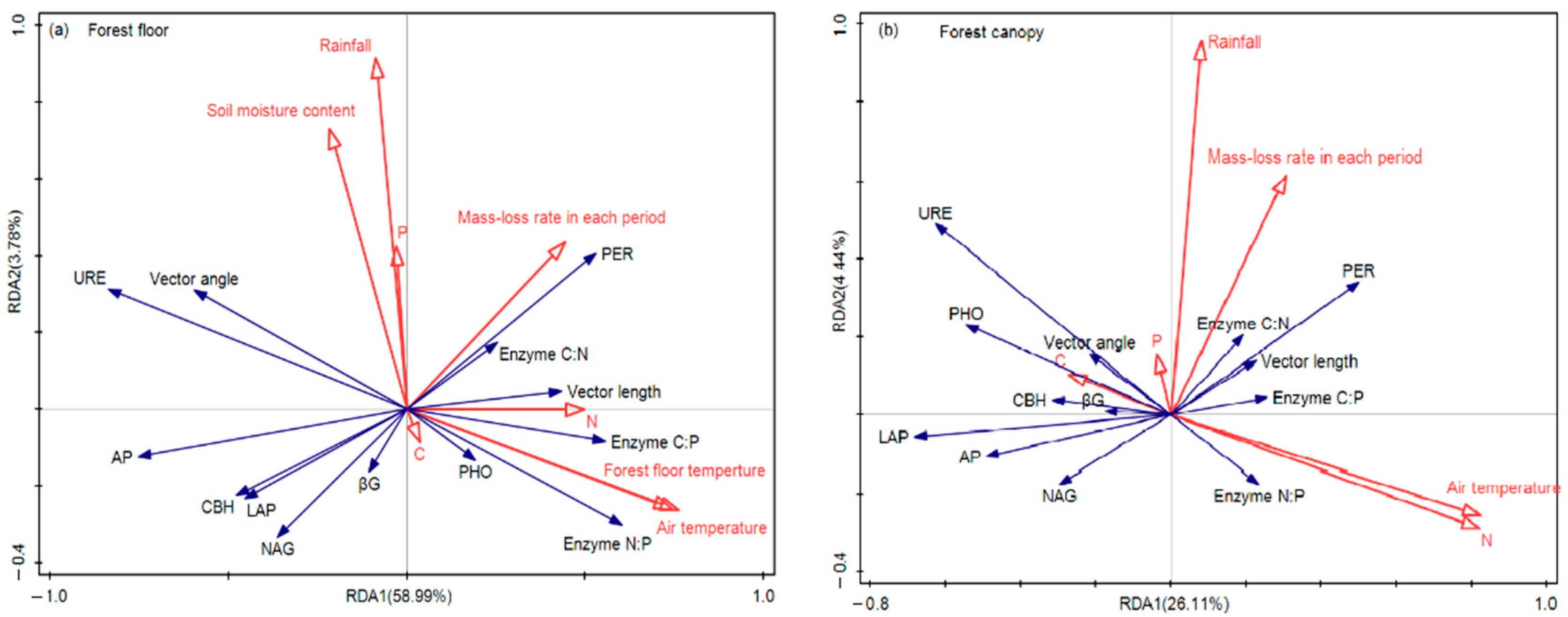
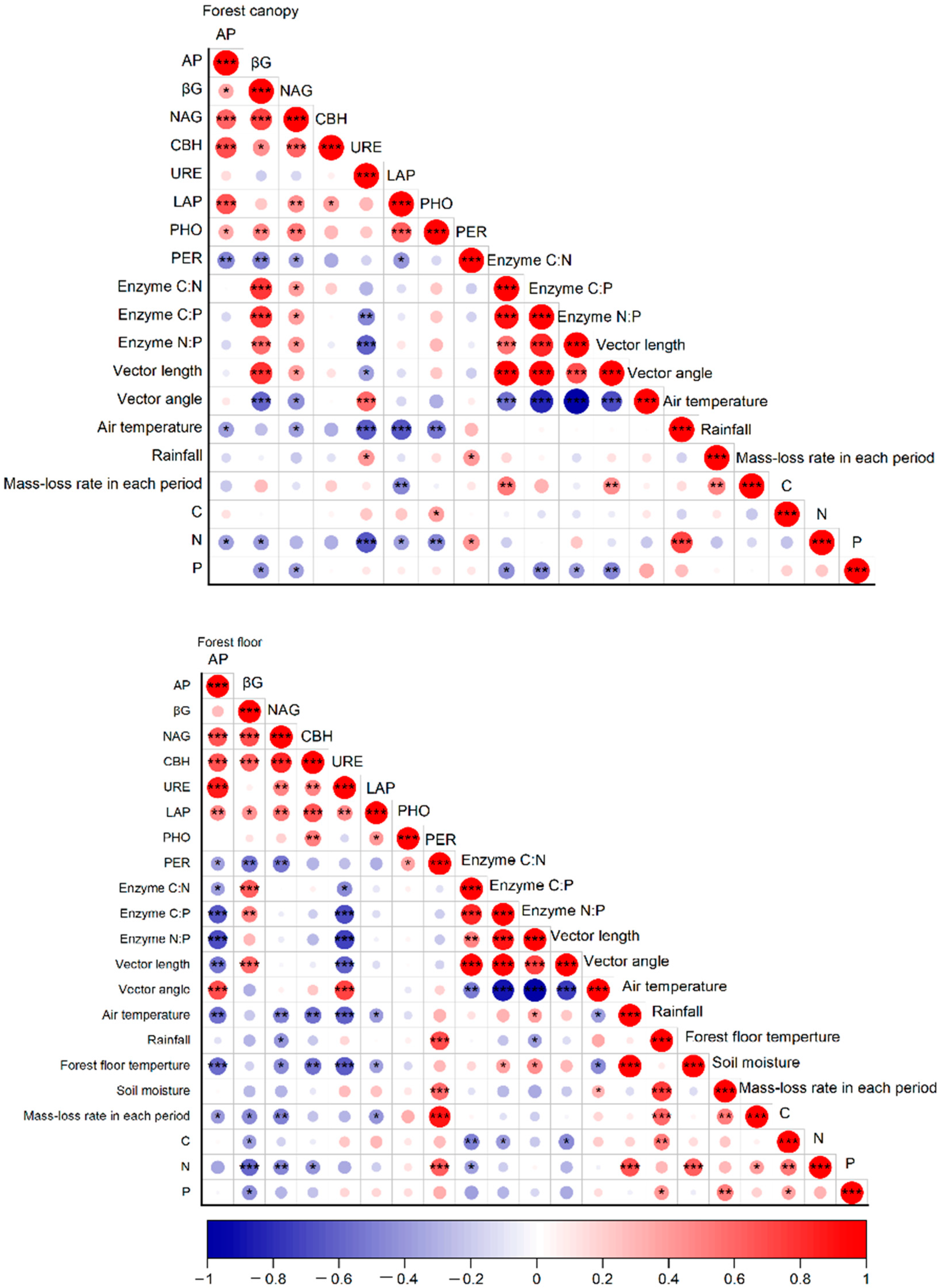
3.3. Driving Factors for Enzyme Activities during the Decomposition of Leaf Litter
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kourtev, P.S.; Ehrenfeld, J.G.; Huang, W.Z. Enzyme activities during litter decomposition of two exotic and two native plant species in hardwood forests of New Jersey. Soil Biol. Biochem. 2002, 34, 1207–1218. [Google Scholar] [CrossRef]
- Li, S.W.; Liu, D.; Li, Z.; Li, L.; Song, K.; Chen, Y.; Fu, Y. Slower rates of litter decomposition of dominant epiphytes in the canopy than on the forest floor in a subtropical montane forest, southwest China. Soil Biol. Biochem. 2014, 70, 211–220. [Google Scholar] [CrossRef]
- Ekschmitt, K.; Kandeler, E.; Poll, C.; Brune, A.; Buscot, F.; Friedrich, M.; Gleixner, G.; Hartmann, A.; Kästner, M.; Marhan, S.; et al. Soil-carbon preservation through habitat constraints and biological limitations on decomposer activity. J. Plant Nutr. Soil Sci. 2008, 171, 27–35. [Google Scholar] [CrossRef]
- Cui, Y.; Fang, L.; Deng, L.; Guo, X.; Zhang, X. Patterns of soil microbial nutrient limitations and their roles in the variation of soil organic carbon across a precipitation gradient in an arid and semi-arid region. Sci. Total Environ. 2019, 658, 1440–1451. [Google Scholar] [CrossRef]
- Duan, C.; Fang, L.; Yang, C.; Chen, W. Reveal the response of enzyme activities to heavy metals through in situ zymography. Ecotoxicol. Environ. Saf. 2018, 156, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Sinsabaugh, R.L.; Hill, B.H.; Shah, J.J.F. Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment. Nature 2009, 462, 795–798. [Google Scholar] [CrossRef]
- Fioretto, A.; Papa, S.; Curcio, E.; Sorrentino, G.; Fuggi, A. Enzyme dynamics on decomposing leaf litter of Cistus incanus and Myrtus communis in a Mediterranean ecosystem. Soil Biol. Biochem. 2000, 32, 1847–1855. [Google Scholar] [CrossRef]
- Qasemian, L.; Guiral, D.; Ziarelli, F.; Dang, T.K.V.; Farnet, A.M. Effects of anthracene on microbial activities and organic matter decomposition in a Pinus halepensis litter from a Mediterranean coastal area. Soil Biol. Biochem. 2012, 46, 148–154. [Google Scholar] [CrossRef]
- Bohlman, S.A.; Matelson, T.J.; Nadkarni, N.M. Moisture and temperature patterns of canopy humus and forest floor soil of a montane cloud forest, costa rica. Biotropica 1995, 27, 13–19. [Google Scholar] [CrossRef]
- Cardelús, C.L.; Chazdon, R.L. Inner-crown microenvironments of two emergent tree species in a lowland wet forest. Biotropica 2005, 37, 238–244. [Google Scholar] [CrossRef]
- Zotz, G.; Leja, M.; Cruz, Y.A.; Einzmann, H.J.R. How much water is in the tank? An allometric analysis with 205 bromeliad species. Flora 2020, 264, 151557. [Google Scholar] [CrossRef]
- Paoletti, M.G.; Taylor, R.A.J.; Stinner, D.H.; Benzing, D.H. Diversity of soil fauna in the canopy and forest floor of a venezuelan cloud forest. J. Trop. Ecol. 1991, 7, 373–383. [Google Scholar] [CrossRef]
- Pittl, E.; Innerebner, G.; Wanek, W.; Insam, H. Microbial communities of arboreal and ground soilsin the Esquinas rainforest, Costa Rica. Plant Soil 2009, 329, 65–74. [Google Scholar] [CrossRef]
- Güsewell, S.; Gessner, M.O. N:P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms. Funct. Ecol. 2009, 23, 211–219. [Google Scholar] [CrossRef]
- Clark, K.L.; Nadkarni, N.M.; Gholz, H.L. Growth, net production, litter decomposition, and net nitrogen accumulation by epiphytic bryophytes in a tropical montane forest. Biotropica 1998, 30, 12–23. [Google Scholar] [CrossRef]
- Cardelús, C.L.; Mack, M.C. The nutrient status of epiphytes and their host trees along an elevational gradient in Costa Rica. Plant Ecol. 2010, 207, 25–37. [Google Scholar] [CrossRef]
- Cardelús, C.L. Litter decomposition within the canopy and forest floor of three tree species in a tropical lowland rain forest, Costa Rica. Biotropica 2010, 42, 300–308. [Google Scholar] [CrossRef]
- Rousk, J.; Nadkarni, N.M. Growth measurements of saprotrophic fungi and bacteria reveal differences between canopy and forest floor soils. Soil Biol. Biochem. 2009, 41, 862–865. [Google Scholar] [CrossRef]
- Allen, A.P.; Gillooly, J.F. Towards an integration of ecological stoichiometry and the metabolic theory of ecology to better understand nutrient cycling. Ecol. Lett. 2009, 12, 369–384. [Google Scholar] [CrossRef]
- Moorhead, D.; Lashermes, G.; Recous, S.; Bertrand, I. Interacting microbe and litter quality controls on litter decomposition: A modeling analysis. PLoS ONE 2016, 9, 108769. [Google Scholar] [CrossRef]
- Cui, Y.X.; Bing, H.J.; Fang, L.C.; Jiang, M.; Zhang, X.C. Extracellular enzyme stoichiometry reveals the carbon and phosphorus limitations of microbial metabolisms in the rhizosphere and bulk soils in alpine ecosystems. Plant Soil 2021, 458, 7–20. [Google Scholar] [CrossRef]
- Zhou, L.H.; Liu, S.S.; Shen, H.H.; Zhao, M.Y.; Xu, L.C.; Xing, A.J.; Fang, J.Y. Soil extracellular enzyme activity and stoichiometry in China’s forests. Funct. Ecol. 2020, 34, 1461–1471. [Google Scholar] [CrossRef]
- Gao, W.; Reed, S.C.; Munson, S.M.; Rui, Y.; Hao, Y. Responses of soil extracellular enzyme activities and bacterial community composition to seasonal stages of drought in a semiarid grassland. Geoderma 2021, 401, 115327. [Google Scholar] [CrossRef]
- Fioretto, A.; Papa, S.; Sorrentino, G.; Fuggi, A. Decomposition of Cistus incanus leaf litter in a Mediterranean maquis ecosystem: Mass loss, microbial enzyme activities and nutrient changes. Soil Biol. Biochem. 2001, 33, 311–321. [Google Scholar] [CrossRef]
- Fanin, N.; Moorhead, D.M.; Bertrand, I. Eco-enzymatic stoichiometry and enzymatic vectors reveal differential C, N, P dynamics in decaying litter along a land-use gradient. Biogeochemistry 2016, 129, 21–36. [Google Scholar] [CrossRef]
- Wang, Q.W.; Robson, T.M.; Pieristè, M.; Kenta, T.; Zhou, W.M.; Kurokawa, H. Canopy structure and phenology modulate the impacts of solar radiation on C and N dynamics during litter decomposition in a temperate forest. Sci. Total Environ. 2022, 820, 153185. [Google Scholar] [CrossRef]
- Wu, B.; Guo, J.; Wu, J.; Ren, W.; Liu, X.; Yang, Y. Effects of logging residues on surface soil biochemical properties and enzymatic activity. Acta Ecol. Sinica 2014, 34, 1645–1653. [Google Scholar]
- Dearden, F.M.; Wardle, D.A. The potential for forest canopy litterfall interception by a dense fern understorey, and the consequences for litter decomposition. Oikos 2007, 117, 83–92. [Google Scholar] [CrossRef]
- Tan, Z.; Zhang, Y.; Schaefer, D.; Yu, G.R.; Liang, N.; Song, Q. An old-growth subtropical Asian evergreen forest as a large carbon sink. Atmos. Environ. 2011, 45, 1548–1554. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, L.; Yang, Z.; Xu, C.; Xie, J.; Chen, G.; Lin, C.; Guo, J.; Liu, X.; Xiong, D.; et al. Large ecosystem service benefits of assisted natural regeneration. J. Geophys. Res. 2018, 123, 676–687. [Google Scholar] [CrossRef]
- Ni, X.; Lin, C.; Chen, G.; Xie, J.; Yang, Z.; Liu, X.; Xiong, D.; Xu, C.; Yue, K.; Wu, F.; et al. Decline in Nutrient Inputs from Litterfall Following Forest Plantation in Subtropical China. For. Ecol. Manag. 2021, 496, 119445. [Google Scholar] [CrossRef]
- Lü, M.; Xie, J.; Wang, C.; Guo, J.; Wang, M.; Liu, X.; Chen, Y.; Chen, G.; Yang, Y. Forest Conversion Stimulated Deep Soil C. Losses and Decreased C Recalcitrance through Priming Effect in Subtropical China. Biol. Fertil. Soils 2015, 51, 857–867. [Google Scholar] [CrossRef]
- Li, S.; Liu, W.; Li, D. Epiphytic lichens in subtropical forest ecosystems in southwest China: Species diversity and implications for conservation. Biol. Conserv. 2013, 159, 88–95. [Google Scholar] [CrossRef]
- Fanin, N.; Bertrand, I. Aboveground litter quality is a better predictor than belowground microbial communities when estimating carbon mineralization along a land-use gradient. Soil Biol. Biochem. 2016, 94, 48–60. [Google Scholar] [CrossRef]
- Feng, J.; Wei, K.; Chen, Z.; Lü, X.; Tian, J.; Wang, C.; Chen, L. Coupling and decoupling of soil carbon and nutrient cycles across an aridity gradient in the drylands of northern china: Evidence from ecoenzymatic stoichiometry. Glob. Biogeochem. Cycles 2019, 33, 559–569. [Google Scholar] [CrossRef]
- Ai, C.; Liang, G.; Sun, J.; Wang, X.; Zhou, W. Responses of extracellular enzyme activities and microbial community in both the rhizosphere and bulk soil to long-term fertilization practices in a fluvo-aquic soil. Geoderma 2012, 173, 330–338. [Google Scholar] [CrossRef]
- Jin, K.; Sleutel, S.; Buchan, D.; Neve, S.D.; Cai, D.; Gabriels, D.; Jin, J. Changes of soil enzyme activities under different tillage practices in the Chinese Loess Plateau. Soil Tillage Res. 2009, 104, 115–120. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M.A. Glucosidases and galactosidases in soils. Biochemistry 1988, 20, 601–606. [Google Scholar] [CrossRef]
- Tabatabai, M.J.; Bremener, J.M. Use of p-nitrophenyl phosphate for assay of soil phosp hatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Šnajdr, J.; Valášková, V.; Merhautová, V.; Herinková, J.; Cajthaml, T.; Baldrian, P. Spatial variability of enzyme activities and microbial biomass in the upper layers of Quercus petraea forest soil. Soil Biol. Biochem. 2008, 40, 2068–2075. [Google Scholar] [CrossRef]
- Valášková, V.; Šnajdr, J.; Bittner, B.; Cajthaml, T.; Merhautová, V.; Hofrichter, M.; Baldrian, P. Production of lignocellulose-degrading enzymes and degradation of leaf litter by saprotrophic basidiomycetes isolated from a Quercus petraea forest. Soil Biol. Biochem. 2007, 39, 2651–2660. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Klug, M.J.; Collins, H.P.; Yeager, P.E.; Petersen, S.O.; Robertson, G.P.; Coleman, D.C.; Bledsoe, C.S.; Sollins, P. Characterizing soil microbial. In Standard Soil Methods for Long-Term Ecological Research; Oxford University Press: Oxford, UK, 1999; Volume 318, pp. 476–525. [Google Scholar]
- Hill, B.H.; Elonen, C.M.; Seifert, L.R.; May, A.A.; Tarquinio, E. Microbial enzyme stoichiometry and nutrient limitation in US streams and rivers. Ecol. Indic. 2012, 18, 540–551. [Google Scholar] [CrossRef]
- Schmidt, S.K.; Porazinska, D.; Concienne, B.L.; Darcy, J.L.; King, A.J.; Nemergut, D.R. Biogeochemical stoichiometry reveals P and N limitation across the post-glacial landscape of denali national park, alaska. Ecosystems 2016, 19, 1164–1177. [Google Scholar] [CrossRef]
- Waring, B.G.; Weintraub, S.R.; Sinsabaugh, R.L. Ecoenzymatic stoichiometry of microbial nutrient acquisition in tropical soils. Biogeochemistry 2014, 117, 101–113. [Google Scholar] [CrossRef]
- Moorhead, D.L.; Rinkes, Z.L.; Sinsabaugh, R.L.; Weintraub, M.N. Dynamic relationships between microbial biomass, respiration, inorganic nutrients and enzyme activities: Informing enzyme-based decomposition models. Front. Microbiol. 2013, 4, 223. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Chen, H.; Wang, Y.; Mao, Q.; Zheng, M.; Su, Y.; Xiao, K.; Wang, K.; Li, D. Responses of soil microbial resource limitation to multiple fertilization strategies. Soil Till Res. 2020, 196, 104474. [Google Scholar] [CrossRef]
- Porder, S.; Clark, D.A.; Vitousek, P.M. Persistence of rock-derived nutrients in the wet tropical forests of la La Selva, Costa Rica. Ecology 2006, 87, 594–602. [Google Scholar] [CrossRef]
- Grandy, S.A.; Neff, J.C.; Weintrau, M.N. Carbon structure and enzyme activities in alpine and forest ecosystems. Soil Biol. Biochem. 2007, 39, 2701–2711. [Google Scholar] [CrossRef]
- Mccain, C.M. Could temperature and water availability drive elevational species richness patterns? A global case study for bats. Glob. Ecol. Biogeogr. 2007, 16, 1–13. [Google Scholar] [CrossRef]
- Melillo, J.M.; Steudler, P.A.; Aber, J.D.; Newkirk, K.; Lux, H.; Bowles, F.P.; Catricala, C.; Magill, A.; Ahrens, T.; Morrisseau, S. Soil warming and carbon-cycle feedbacks to the climate system. Science 2002, 298, 2173–2176. [Google Scholar] [CrossRef]
- Ouyang, S.; Tian, Y.; Liu, Q.; Zhang, L.; Wang, R.; Xu, X. Nitrogen competition between three dominant plant species and microbes in a temperate grassland. Plant Soil 2016, 408, 121–132. [Google Scholar] [CrossRef]
- Peng, X.; Wang, W. Stoichiometry of soil extracellular enzyme activity along a climatic transect in temperate grasslands of northern China. Soil Biol. Biochem. 2016, 98, 74–84. [Google Scholar] [CrossRef]
- Xu, M.; Li, W.; Wang, J.; Zhu, Y.; Feng, Y.; Yang, G.; Zhang, W.; Han, X. Soil ecoenzymatic stoichiometry reveals microbial phosphorus limitation after vegetation restoration on the Loess Plateau, China. Sci. Total Environ. 2022, 815, 152918. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Wang, X.; Zhang, X.; Ju, W.; Duan, C.; Guo, X.; Wang, Y.; Fang, L. Soil moisture mediates microbial carbon and phosphorus metabolism during vegetation succession in a semiarid region. Biochemistry 2020, 147, 107814. [Google Scholar] [CrossRef]
- Duan, C.; Wang, Y.; Wang, Q.; Ju, W.; Zhang, Z.; Cui, Y.; Yuan, J.; Fan, Q.; Wei, S.; Li, S.; et al. Microbial metabolic limitation of rhizosphere under heavy metal stress: Evidence from soil ecoenzymatic stoichiometry. Environ. Pollut. 2022, 300, 118978. [Google Scholar] [CrossRef]
- Allison, S.D.; Vitousek, P.M. Extracellular enzyme activities and carbon chemistry as drivers of tropical plant litter decomposition. Biotropica 2004, 36, 285–296. [Google Scholar] [CrossRef]
- Schofield, J.A.; Hagerman, A.E.; Harold, A. Loss of tannins and other phenolics from willow leaf litter. J. Chem. Ecol. 1998, 24, 1409–1421. [Google Scholar] [CrossRef]
- Burns, R.G. Enzyme activity in soil: Location and a possible role in microbial ecology. Soil Biol. Biochem. 1982, 14, 423–427. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L. Enzymic analysis of microbial pattern and process. Biol. Fertil. Soils. 1994, 17, 69–74. [Google Scholar] [CrossRef]
- Appel, H.M. Phenolics in ecological interactions: The importance of oxidation. J. Chem. Ecol. 1993, 19, 1521–1552. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, L.; Wei, W.; Wu, R.; Zhang, X.; Guo, H.; Wang, D.; Wu, F. Dynamics of Enzyme Activities during the Decomposition of Castanopsis carlesii Leaf Litter in the Forest Canopy and Forest Floor in a Mid-Subtropical Area. Forests 2022, 13, 1944. https://doi.org/10.3390/f13111944
Zhu L, Wei W, Wu R, Zhang X, Guo H, Wang D, Wu F. Dynamics of Enzyme Activities during the Decomposition of Castanopsis carlesii Leaf Litter in the Forest Canopy and Forest Floor in a Mid-Subtropical Area. Forests. 2022; 13(11):1944. https://doi.org/10.3390/f13111944
Chicago/Turabian StyleZhu, Ling, Wentao Wei, Ruobing Wu, Xiaoyue Zhang, Hongrong Guo, Dingyi Wang, and Fuzhong Wu. 2022. "Dynamics of Enzyme Activities during the Decomposition of Castanopsis carlesii Leaf Litter in the Forest Canopy and Forest Floor in a Mid-Subtropical Area" Forests 13, no. 11: 1944. https://doi.org/10.3390/f13111944