


The Effect of Quercetin on the Growth, Development, Nutrition Utilization, and Detoxification Enzymes in Hyphantria cunea Drury (Lepidoptera: Arctiidae)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect
2.2. Feeding Treatment
2.3. Growth and Development Assay
2.4. Mortality Assay
2.5. Nutritional Effect Assay
2.6. Detoxification Enzyme Activity or Concentration Assays
2.7. Statistical Analysis
3. Results
3.1. Effects of Quercetin on the Growth and Development of H. cunea
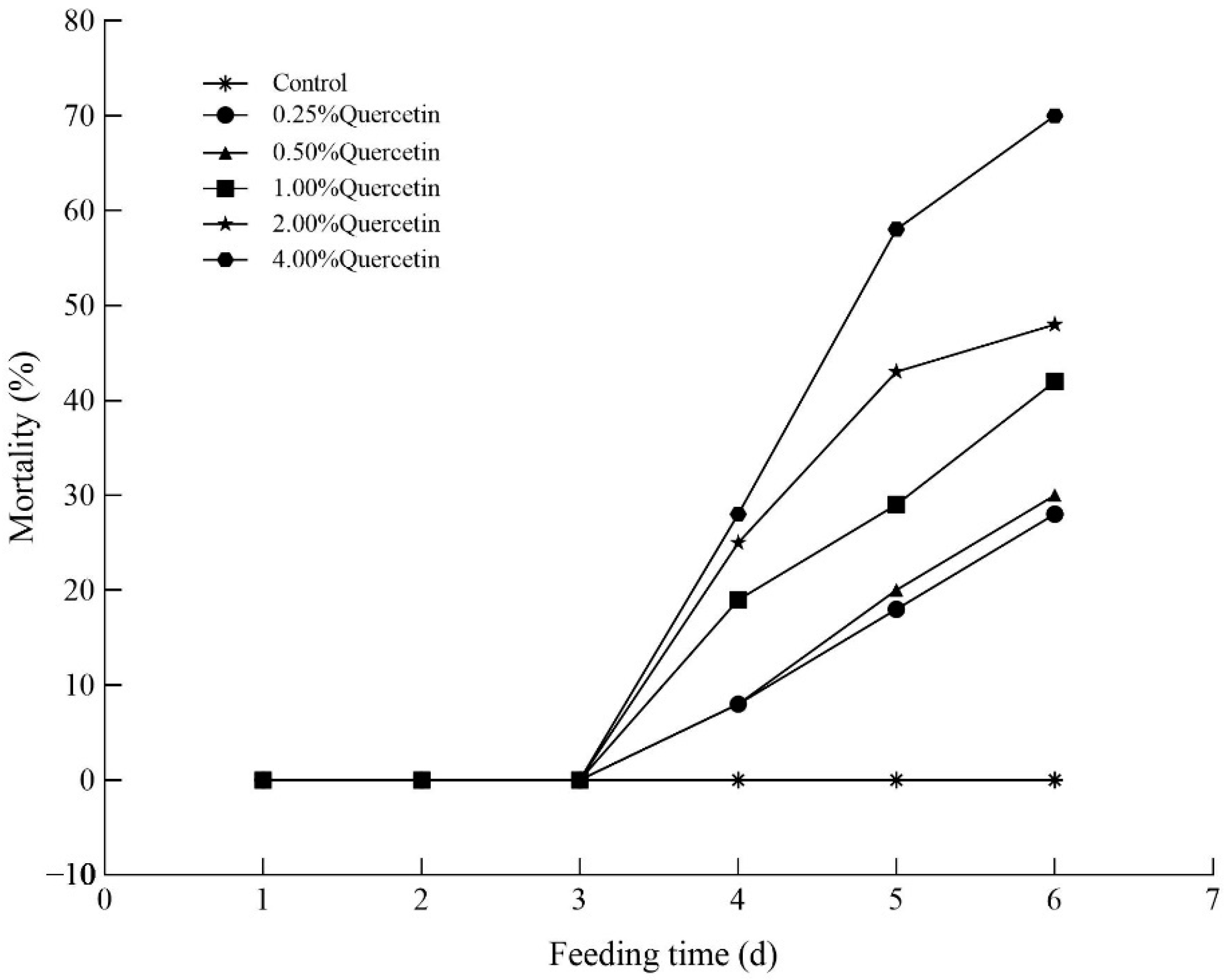
3.2. Effects of Quercetin on the Mortality of H. cunea
3.3. Effects of Quercetin on Nutrition in H. cunea Larvae
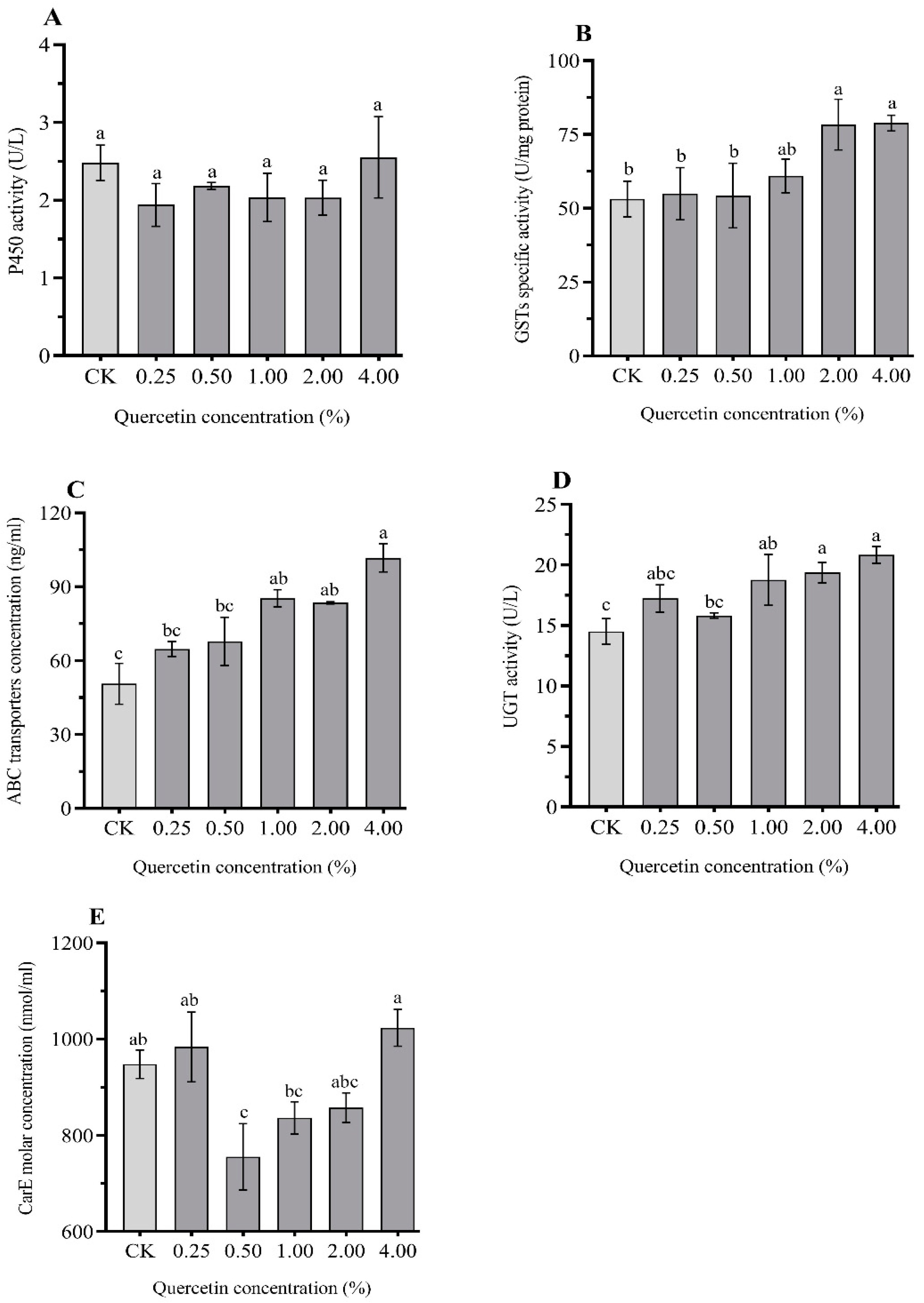
3.4. Effects of Quercetin on the Detoxification Enzymes in H. cunea Larvae
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [Green Version]
- Senthil-Nathan, S. Physiological and biochemical effect of neem and other Meliaceae plants secondary metabolites against Lepidopteran insects. Front. Physiol. 2013, 4, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatehouse, J.A. Plant resistance towards insect herbivores: A dynamic interaction. New Phytol. 2002, 156, 145–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, H.; Sujana, G.; Manohar Rao, D. Morphological and chemical components of resistance to pod borer, Helicoverpa armigera in wild relatives of pigeonpea. Arthropod-Plant Interact. 2009, 3, 151–161. [Google Scholar] [CrossRef] [Green Version]
- Kelly, M.; Curry, J. The influence of phenolic compounds on the suitability of three Salix species as hosts for the willow beetle Phratora vulgatissima. Entomol. Exp. Appl. 1991, 61, 25–32. [Google Scholar] [CrossRef]
- Aljbory, Z.; Chen, M.S. Indirect plant defense against insect herbivores: A review. Insect Sci. 2018, 25, 2–23. [Google Scholar] [CrossRef]
- Hafeez, M.; Liu, S.; Jan, S.; Ali, B.; Shahid, M.; Fernandez-Grandon, G.M.; Nawaz, M.; Ahmad, A.; Wang, M. Gossypol-induced fitness gain and increased resistance to deltamethrin in beet armyworm, Spodoptera exigua (Hubner). Pest Manag. Sci. 2019, 75, 683–693. [Google Scholar] [CrossRef]
- Caballero, C.; Lopez-Olguin, J.; Ruiz, M.; Ortego, F.; Castanera, P. Antifeedant activity and effects of terpenoids on detoxication enzymes of the beet armyworm, Spodoptera exigua (Hubner). Span. J. Agric. Res. 2008, 6, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Walker, W.B., III.; Vogel, H.; Chattington, S.; Larsson, M.C.; Anderson, P.; Heckel, D.G.; Schlyter, F. Diet dependent metabolic responses in three generalist insect herbivores Spodoptera spp. Insect Biochem. Mol. Biol. 2016, 71, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Termonia, A.; Hsiao, T.H.; Pasteels, J.M.; Milinkovitch, M.C. Feeding specialization and host-derived chemical defense in Chrysomeline leaf beetles did not lead to an evolutionary dead end. Proc. Natl. Acad. Sci. USA 2001, 98, 3909–3914. [Google Scholar] [CrossRef]
- Heidel-Fischer, H.M.; Vogel, H. Molecular mechanisms of insect adaptation to plant secondary compounds. Curr. Opin. Insect. Sci. 2015, 8, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Dobler, S.; Petschenka, G.; Pankoke, H. Coping with toxic plant compounds—The insect’s perspective on iridoid glycosides and cardenolides. Phytochemistry 2011, 72, 1593–1604. [Google Scholar] [CrossRef] [PubMed]
- Després, L.; David, J.-P.; Gallet, C. The evolutionary ecology of insect resistance to plant chemicals. Trends Ecol. Evol. 2007, 22, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Bozzolan, F.; Siaussat, D.; Maria, A.; Durand, N.; Pottier, M.A.; Chertemps, T.; Maïbèche-Coisne, M. Antennal uridine diphosphate (UDP)-glycosyltransferases in a pest insect: Diversity and putative function in odorant and xenobiotics clearance. Insect Mol. Biol. 2014, 23, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Ren, N.; Xie, M.; You, Y.; Li, J.; Chen, W.; Cheng, X.; You, M. Fipronil-resistance mediated by carboxylesterases in the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Acta Entomol. Sin. 2015, 58, 288–296. [Google Scholar]
- Feyereisen, R. Insect P450 enzymes. Annu. Rev. Entomol. 1999, 44, 507. [Google Scholar] [CrossRef]
- Ge, X.; He, S.; Zhu, C.; Wang, T.; Xu, Z.; Zong, S. Projecting the current and future potential global distribution of Hyphantria cunea (Lepidoptera: Arctiidae) using CLIMEX. Pest Manag. Sci. 2019, 75, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, G.; Ozman-Sullivan, S. Tachinid (Diptera) parasitoids of Hyphantria cunea (Lepidoptera: Arctiidae) in its native North America and in Europe and Asia–a literature review. Entomol. Fenn. 2012, 23, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Pan, L.; Zhao, Y.; Zhang, X.; Wang, F.; Yu, Y.; Fan, W.; Liu, Q.; Zhang, S.; Li, M. Chemical investigations of volatile kairomones produced by Hyphantria cunea (Drury), a host of the parasitoid Chouioia cunea Yang. Bull. Entomol. Res. 2017, 107, 234–240. [Google Scholar] [CrossRef]
- Chen, Q.; Zhao, H.; Wen, M.; Li, J.; Zhou, H.; Wang, J.; Zhou, Y.; Liu, Y.; Du, L.; Kang, H.; et al. Genome of the webworm Hyphantria cunea unveils genetic adaptations supporting its rapid invasion and spread. BMC Genom. 2020, 21, 242. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Huang, X.; Chang, B.H.; Zhang, Z. The survival, growth, and detoxifying enzyme activities of grasshoppers Oedaleus asiaticus (Orthoptera: Acrididae) exposed to toxic rutin. Appl. Entomol. Zool. 2020, 55, 385–393. [Google Scholar] [CrossRef]
- Harnly, J.M.; Doherty, R.F.; Beecher, G.R.; Holden, J.M.; Haytowitz, D.B.; Bhagwat, S.; Gebhardt, S. Flavonoid content of US fruits, vegetables, and nuts. J. Agric. Food Chem. 2006, 54, 9966–9977. [Google Scholar] [CrossRef] [PubMed]
- Harwood, M.; Danielewska-Nikiel, B.; Borzelleca, J.; Flamm, G.; Williams, G.; Lines, T. A critical review of the data related to the safety of quercetin and lack of evidence of in vivo toxicity, including lack of genotoxic/carcinogenic properties. Food Chem. Toxicol. 2007, 45, 2179–2205. [Google Scholar] [CrossRef] [PubMed]
- Aboshi, T.; Ishida, M.; Matsushita, K.; Hirano, Y.; Nishida, R.; Mori, N. Stage-specific quercetin sulfation in the gut of Mythimna separata larvae (Lepidoptera: Noctuidae). Biosci. Biotechnol. Biochem. 2014, 78, 38–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Yang, J.; Gu, S.; Wang, X.; Wang, J.; Zhou, C.; Lv, J.; Zeng, X. Study on antioxidant activity of quercetin extract from mulberry leaves. J. Silk 2018, 55, 15–20. [Google Scholar] [CrossRef]
- Hajhashemi, V.; Ghannadi, A.; Mousavi, S. Antinociceptive study of extracts of Platanus orientalis leaves in mice. Res. Pharm. Sci. 2011, 6, 123. [Google Scholar]
- Gao, M.; He, M.; Xing, R.; Wang, X.; Wang, Z. Borate-modified carbon dots as a probe for quercetin in plants. Analyst 2021, 146, 590–596. [Google Scholar] [CrossRef]
- Li, Z.; Guan, X.; Michaud, J.; Zhang, Q.; Liu, X. Quercetin interacts with Cry1Ac protein to affect larval growth and survival of Helicoverpa armigera. Pest Manag. Sci. 2016, 72, 1359–1365. [Google Scholar] [CrossRef]
- Hafeez, M.; Qasim, M.; Ali, S.; Yousaf, H.K.; Waqas, M.; Ali, E.; Ahmad, M.A.; Jan, S.; Bashir, M.A.; Noman, A. Expression and functional analysis of P450 gene induced tolerance/resistance to lambda-cyhalothrin in quercetin fed larvae of beet armyworm Spodoptera exigua (Hübner). Saudi J. Biol. Sci. 2020, 27, 77–87. [Google Scholar] [CrossRef]
- Liu, D.; Yuan, Y.; Li, M.; Qiu, X. Effects of dietary quercetin on performance and cytochrome P450 expression of the cotton bollworm, Helicoverpa armigera. Bull. Entomol. Res. 2015, 105, 771–777. [Google Scholar] [CrossRef]
- Selin-Rani, S.; Senthil-Nathan, S.; Thanigaivel, A.; Vasantha-Srinivasan, P.; Edwin, E.-S.; Ponsankar, A.; Lija-Escaline, J.; Kalaivani, K.; Abdel-Megeed, A.; Hunter, W.B. Toxicity and physiological effect of quercetin on generalist herbivore, Spodoptera litura Fab. and a non-target earthworm Eisenia fetida Savigny. Chemosphere 2016, 165, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Diaz Napal, G.N.; Palacios, S.M. Bioinsecticidal effect of the flavonoids pinocembrin and quercetin against Spodoptera frugiperda. J. Pest Sci. 2015, 88, 629–635. [Google Scholar] [CrossRef]
- Cui, B.; Huang, X.; Li, S.; Hao, K.; Chang, B.H.; Tu, X.; Pang, B.; Zhang, Z. Quercetin affects the growth and development of the grasshopper Oedaleus asiaticus (Orthoptera: Acrididae). J. Econ. Entomol. 2019, 112, 1175–1182. [Google Scholar] [CrossRef]
- Wang, Z.; Nur, F.A.; Ma, J.; Wang, J.; Cao, C. Effects of poplar secondary metabolites on performance and detoxification enzyme activity of Lymantria dispar. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2019, 225, 108587. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Dong, X.; Zhao, Y.; Zheng, B. Induction of carboxylesterase, glutathione S-transferase and acetylcholinesterase by quercetin in Helicoverpa armigera. Chin. J. Pestic. Sci. 1999, 1, 56–60. [Google Scholar]
- Chen, C.; Kang, Z.; Shi, X.; Gao, X. Metabolic adaptation mechanisms of insects to plant secondary metabolites and their implications for insecticide resistance of insects. J. Entomol. 2015, 58, 1126–1139. [Google Scholar] [CrossRef]
- Wang, R.; Sun, Y.; Liang, X.; Song, Y.; Yijuan, S.; Keyuan, Z. Effects of six plant secondary metabolites on activities of detoxification enzymes in Spodoptera litura. Acta Ecol. Sin. 2012, 32, 5191–5198. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Hussain, B.; Buhroo, A.A.; Ignacimuthu, S.; Sharma, H.C. Effect of plant secondary metabolites on legume pod borer, Helicoverpa armigera. J. Pest Sci. 2013, 86, 399–408. [Google Scholar] [CrossRef]
- Chen, C.; Han, P.; Yan, W.; Wang, S.; Shi, X.; Zhou, X.; Desneux, N.; Gao, X. Uptake of quercetin reduces larval sensitivity to lambda-cyhalothrin in Helicoverpa armigera. J. Pest Sci. 2018, 91, 919–926. [Google Scholar] [CrossRef]
- Tang, F.; Zhang, X.; Liu, Y.; Gao, X.; Ju, Y. Inhibition of glutathione S-transferase activity by twenty-one insecticides and three allelochemicals in various tissues of Clostera anachoreta (Fabricius)(Lepidoptera: Notodontidae). Acta Entomol. Sin. 2009, 52, 576–581. [Google Scholar] [CrossRef]
- Cao, L.; Yang, F.; Tang, S.; Chen, M. Development of an artificial diet for three lepidopteran insects. Chin. J. Appl. Entomol. 2014, 51, 1376–1386. [Google Scholar]
- Wu, H.; Sun, D.; Qian, W.; Lou, L.; Jiang, H. Acid hydrolysis followed by HPLC for simultaneous determination of quercetin and kaempferol in Folium Mori. Chin. J. Mod. Appl. Pharm. 2010, 27, 248–251. [Google Scholar] [CrossRef]
- Di, X.; Gu, L.; Wang, H.; Xu, L.; Bao, L. Simultaneous determination of chlorogenic acid, rutin, isoquercetin, astragalin and quercetin in mori folium by HPLC. Chin. J. Exp. Tradit. Med. Formulae 2014, 20, 92–95. [Google Scholar] [CrossRef]
- Pan, Z.; Mo, X.; Meng, X.; Chen, M. Effects of chlorogenic acid on the growth and development and detoxification-related protein activities in Hyphantria cunea (Lepidoptera: Arctiidae) larvae. Acta Entomol. Sin. 2020, 63, 1081–1090. [Google Scholar] [CrossRef]
- Waldbauer, G. The consumption and utilization of food by insects. In Advances in Insect Physiology; Elsevier: Amsterdam, The Netherlands, 1968; Volume 5, pp. 229–288. [Google Scholar]
- Raubenheimer, D.; Simpson, S. Analysis of covariance: An alternative to nutritional indices. Entomol. Exp. Appl. 1992, 62, 221–231. [Google Scholar] [CrossRef]
- Baldwin, I. Plant responses to insect herbivory: The emerging molecular analysis. Scavenging deleterious oxygen radicals. Annu. Rev. Plant Biol. 2002, 53, 299–328. [Google Scholar]
- Wink, M. Plant secondary metabolites modulate insect behavior-steps toward addiction? Front. Physiol. 2018, 9, 364. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Li, L.; Zhao, J.; Chen, M. Effect of tannic acid on nutrition and activities of detoxification enzymes and acetylcholinesterase of the fall webworm (Lepidoptera: Arctiidae). J. Insect Sci. 2020, 20, 8. [Google Scholar] [CrossRef]
- Riddick, E.W. Potential of quercetin to reduce herbivory without disrupting natural enemies and pollinators. Agriculture 2021, 11, 476. [Google Scholar] [CrossRef]
- Gikonyo, N.K.; Mwangi, R.W.; Midiwo, J.O. Toxicity and growth-inhibitory activity of Polygonum senegalense (Meissn.) surface exudate against Aedes aegypti larvae. Int. J. Trop. Insect Sci. 1998, 18, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Sohal, S.K. Bioefficacy of quercetin against melon fruit fly. Bull. Insectol. 2013, 66, 79–83. [Google Scholar]
- Shi, G.; Kang, Z.; Ren, F.; Zhou, Y.; Guo, P. Effects of quercetin on the growth and expression of immune-pathway-related genes in silkworm (Lepidoptera: Bombycidae). J. Insect Sci. 2020, 20, 23. [Google Scholar] [CrossRef] [PubMed]
- Mallikarjuna, N.; Kranthi, K.; Jadhav, D.; Kranthi, S.; Chandra, S. Influence of foliar chemical compounds on the development of Spodoptera litura (Fab.) in interspecific derivatives of groundnut. J. Appl. Entomol. 2004, 128, 321–328. [Google Scholar] [CrossRef]
- Ateyyat, M.; Abu-Romman, S.; Abu-Darwish, M.; Ghabeish, I. Impact of flavonoids against woolly apple aphid, Eriosoma lanigerum (Hausmann) and its sole parasitoid, Aphelinus mali (Hald.). J. Agric. Sci. 2012, 4, 227. [Google Scholar] [CrossRef]
- Scriber, J.; Slansky Jr, F. The nutritional ecology of immature insects. Annu. Rev. Entomol. 1981, 26, 183–211. [Google Scholar] [CrossRef]
- Zeng, X.; Yang, Y.; Chen, Z.; Tang, Q.; Shi, L. Contents Determination of Luteoloside, Quercetin and Hyperoside in the Extract of Lonicera japonica by HPCE. China Pharm. 2017, 28, 2543–2545. [Google Scholar] [CrossRef]
- Zhang, Y.; Tang, Q.; Dai, J.; Wei, X.; Yan, J.; Jiang, Y. Study on the preparation of flavonoids extracted from mulberry leaves and its anti-oxidative activity in vitro. China Food Addit. 2020, 31, 47–52. [Google Scholar] [CrossRef]
- Lei, W.; Lu-Sha, L.; Li-Ying, W.; Yu-Fei, Y.; Min, C. Effects of gallic acid on the nutritional efficiency and detoxificationenzymes in Hyphantria cunea larvae. J. Environ. Entomol. 2020, 42, 471–479. [Google Scholar] [CrossRef]
- Birnbaum, S.S.; Rinker, D.C.; Gerardo, N.M.; Abbot, P. Transcriptional profile and differential fitness in a specialist milkweed insect across host plants varying in toxicity. Mol. Ecol. 2017, 26, 6742–6761. [Google Scholar] [CrossRef]
- Barve, P.R.; Tellis, M.B.; Barvkar, V.T.; Joshi, R.S.; Giri, A.P.; Kotkar, H.M. Functional Diversity of the Lepidopteran ATP-Binding Cassette Transporters. J. Mol. Evol. 2022, 90, 258–270. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Enayati, A.A.; Ranson, H.; Hemingway, J. Insect glutathione transferases and insecticide resistance. Insect Mol. Biol. 2005, 14, 3–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Hu, S.; Huang, H.; Wei, Q.; Ren, M.; Huang, S.; Tian, X.; Su, J. Insecticides induce the co-expression of glutathione S-transferases through ROS/CncC pathway in Spodoptera exigua. Pestic. Biochem. Physiol. 2019, 155, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Xu, L.; Liu, H.; Chen, X.; Zhou, L. Metabolism and antioxidant activity of SlGSTD1 in Spodoptera litura as a detoxification enzyme to pyrethroids. Sci. Rep. 2022, 12, 10108. [Google Scholar] [CrossRef]
- Zhang, Y.-E.; Ma, H.-J.; Feng, D.-D.; Lai, X.-F.; Chen, Z.-M.; Xu, M.-Y.; Yu, Q.-Y.; Zhang, Z. Induction of detoxification enzymes by quercetin in the silkworm. J. Econ. Entomol. 2012, 105, 1034–1042. [Google Scholar] [CrossRef]
- Tangtrakulwanich, K.; Reddy, G.V.; Wu, S.; Miller, J.H.; Ophus, V.L.; Prewett, J. Developing nominal threshold levels for Phyllotreta cruciferae (Coleoptera: Chrysomelidae) damage on canola in Montana, USA. Crop Prot. 2014, 66, 8–13. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatment | Developmental Duration of Larval Instar (Day) | Larval Period from the 3rd Instar (Day) | ||||
|---|---|---|---|---|---|---|
| 3rd Instar | 4th Instar | 5th Instar | 6th Instar | 7th Instar | ||
| Control | 3.0 ± 0.00 b | 5.4 ± 0.09 b | 4.7 ± 0.05 b | 3.8 ± 0.07 a | 4.1 ± 0.11 b | 21.2 ± 0.13 b |
| Quercetin | 4.2 ± 0.06 a | 7.1 ± 0.09 a | 5.8 ± 0.09 a | 3.8 ± 0.12 a | 4.4 ± 0.11 a | 25.6 ± 0.11 a |
| Treatment | Total Survival Rate of Larvae (%) | Pupation Rate (%) | Emergence Rate (%) | Sex Ratio (m/f) | Number of Eggs Laid Per Female |
|---|---|---|---|---|---|
| Control | 74.44 ± 1.14 a | 100 ± 0.00 a | 92.54 ± 1.17 a | 0.72 ± 0.017 a | 673.39 ± 34.80 a |
| Quercetin | 60.00 ± 1.18 b | 94.44 ± 1.15 b | 94.12 ± 1.13 a | 0.46 ± 0.023 b | 592.80 ± 43.03 a |
| Quercetin Concentration (%) | AD (%) | ECD (%) | ECI (%) | GR (g/g·d) | CI (g/g·d) |
|---|---|---|---|---|---|
| Control | 0.885 ± 0.001 a | 0.113 ± 0.013 a | 0.100 ± 0.010 a | 0.200 ± 0.011 a | 1.992 ± 0.068 a |
| 0.25 | 0.848 ± 0.035 ab | 0.140 ± 0.004 a | 0.120 ± 0.006 ab | 0.207 ± 0.008 a | 1.721 ± 0.028 b |
| 0.50 | 0.757 ± 0.019 b | 0.227 ± 0.001 a | 0.172 ± 0.010 a | 0.260 ± 0.012 a | 1.513 ± 0.040 c |
| 1.00 | 0.646 ± 0.016 bc | 0.270 ± 0.031 a | 0.174 ± 0.017 a | 0.244 ± 0.025 a | 1.401 ± 0.014 cd |
| 2.00 | 0.602 ± 0.024 c | 0.301 ± 0.076 a | 0.182 ± 0.042 a | 0.235 ± 0.049 a | 1.300 ± 0.031 d |
| 4.00 | 0.326 ± 0.035 d | 0.183 ± 0.086 a | 0.054 ± 0.020 b | 0.051 ± 0.020 b | 0.912 ± 0.040 e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.-L.; Pan, Z.-Y.; Meng, X.; Yuan, Y.-F.; Li, H.-Y.; Chen, M. The Effect of Quercetin on the Growth, Development, Nutrition Utilization, and Detoxification Enzymes in Hyphantria cunea Drury (Lepidoptera: Arctiidae). Forests 2022, 13, 1945. https://doi.org/10.3390/f13111945
Gao Y-L, Pan Z-Y, Meng X, Yuan Y-F, Li H-Y, Chen M. The Effect of Quercetin on the Growth, Development, Nutrition Utilization, and Detoxification Enzymes in Hyphantria cunea Drury (Lepidoptera: Arctiidae). Forests. 2022; 13(11):1945. https://doi.org/10.3390/f13111945
Chicago/Turabian StyleGao, Yi-Lin, Zhong-Yu Pan, Xiang Meng, Yu-Fei Yuan, Hong-Yun Li, and Min Chen. 2022. "The Effect of Quercetin on the Growth, Development, Nutrition Utilization, and Detoxification Enzymes in Hyphantria cunea Drury (Lepidoptera: Arctiidae)" Forests 13, no. 11: 1945. https://doi.org/10.3390/f13111945