
Pollen Morphological Inter- and Intraspecific Variability in Selected Species of Rubus L. (Rosaceae)

Abstract
:1. Introduction
2. Material and Methods
3. Results
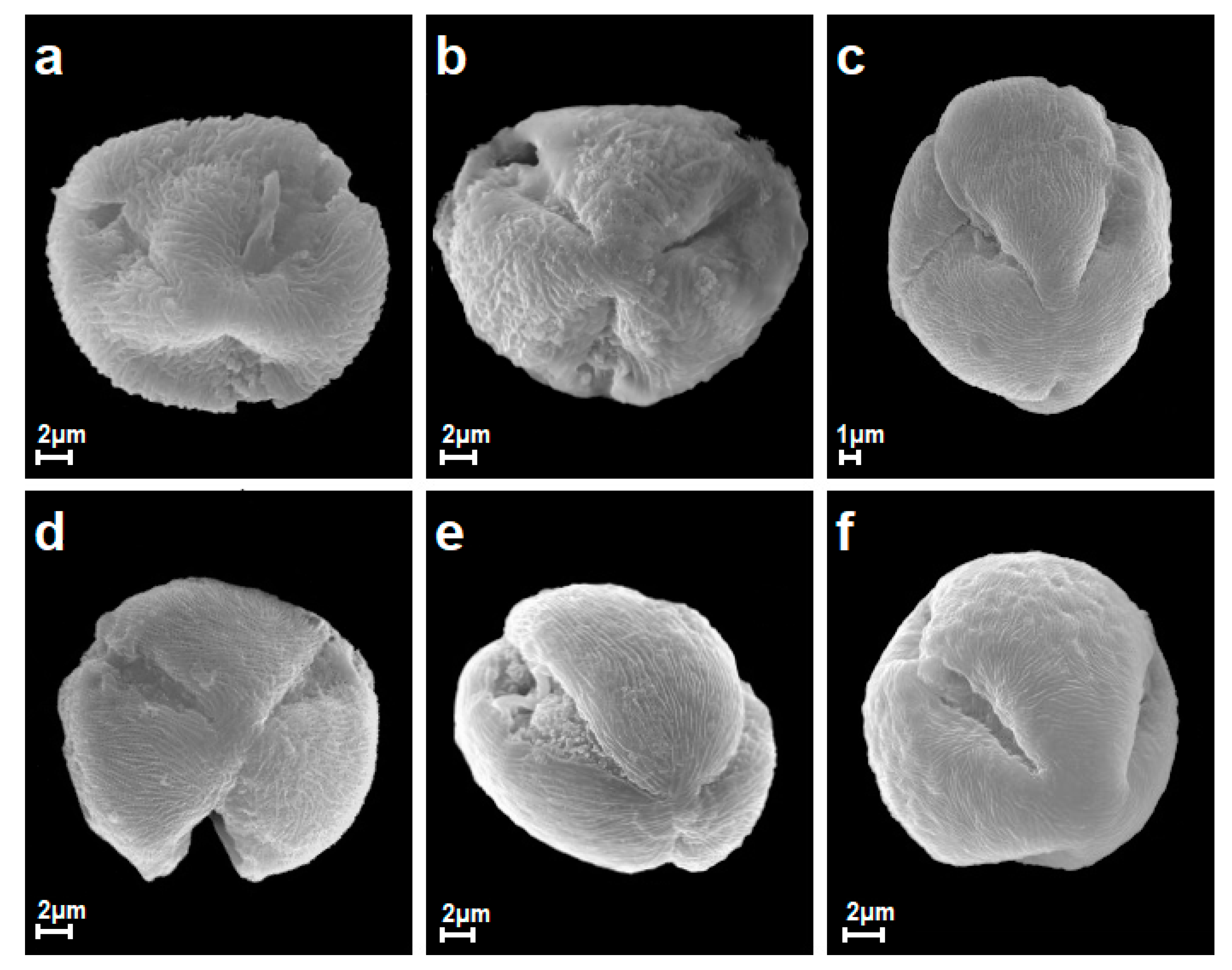
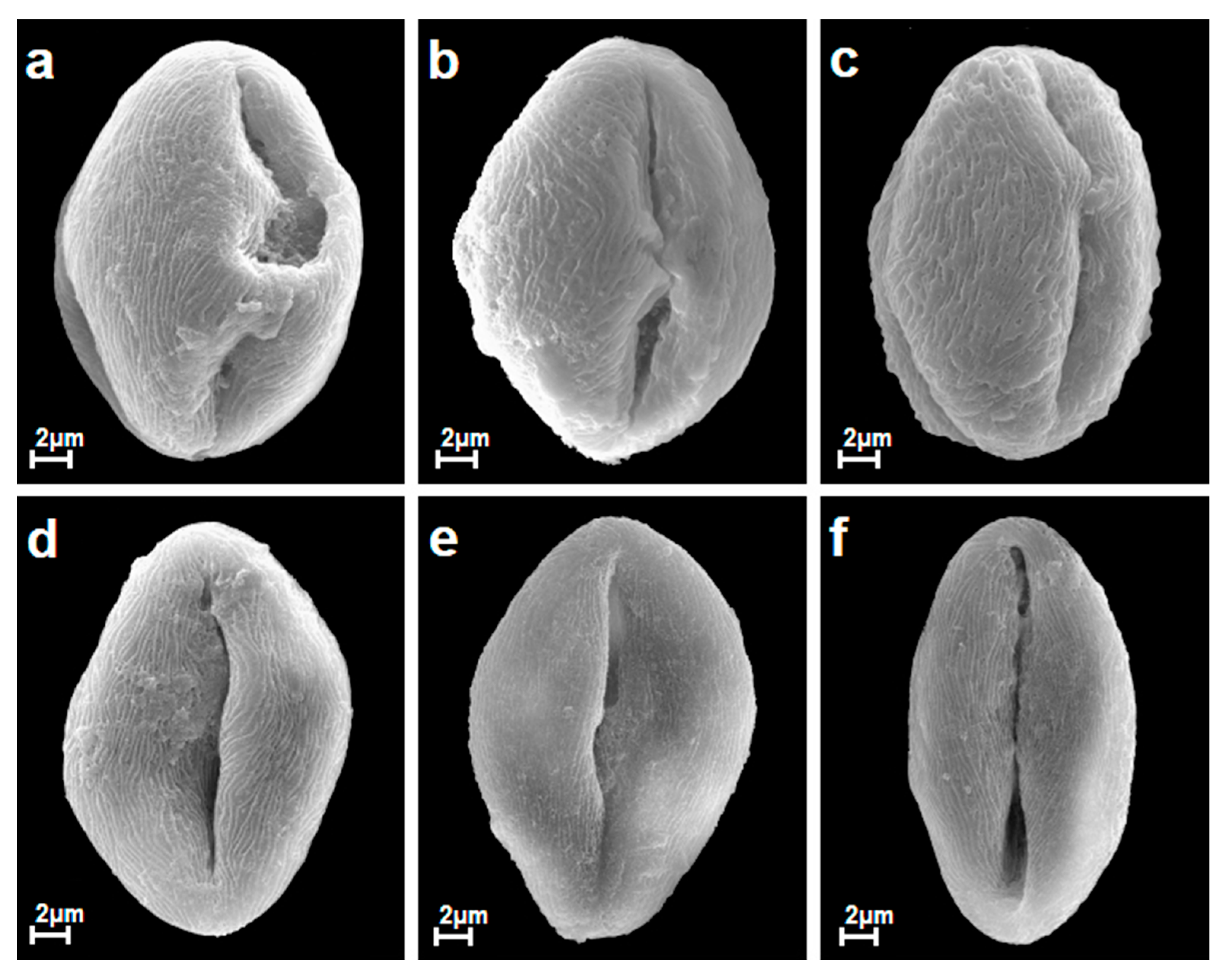
3.1. General Morphological Description of Pollen
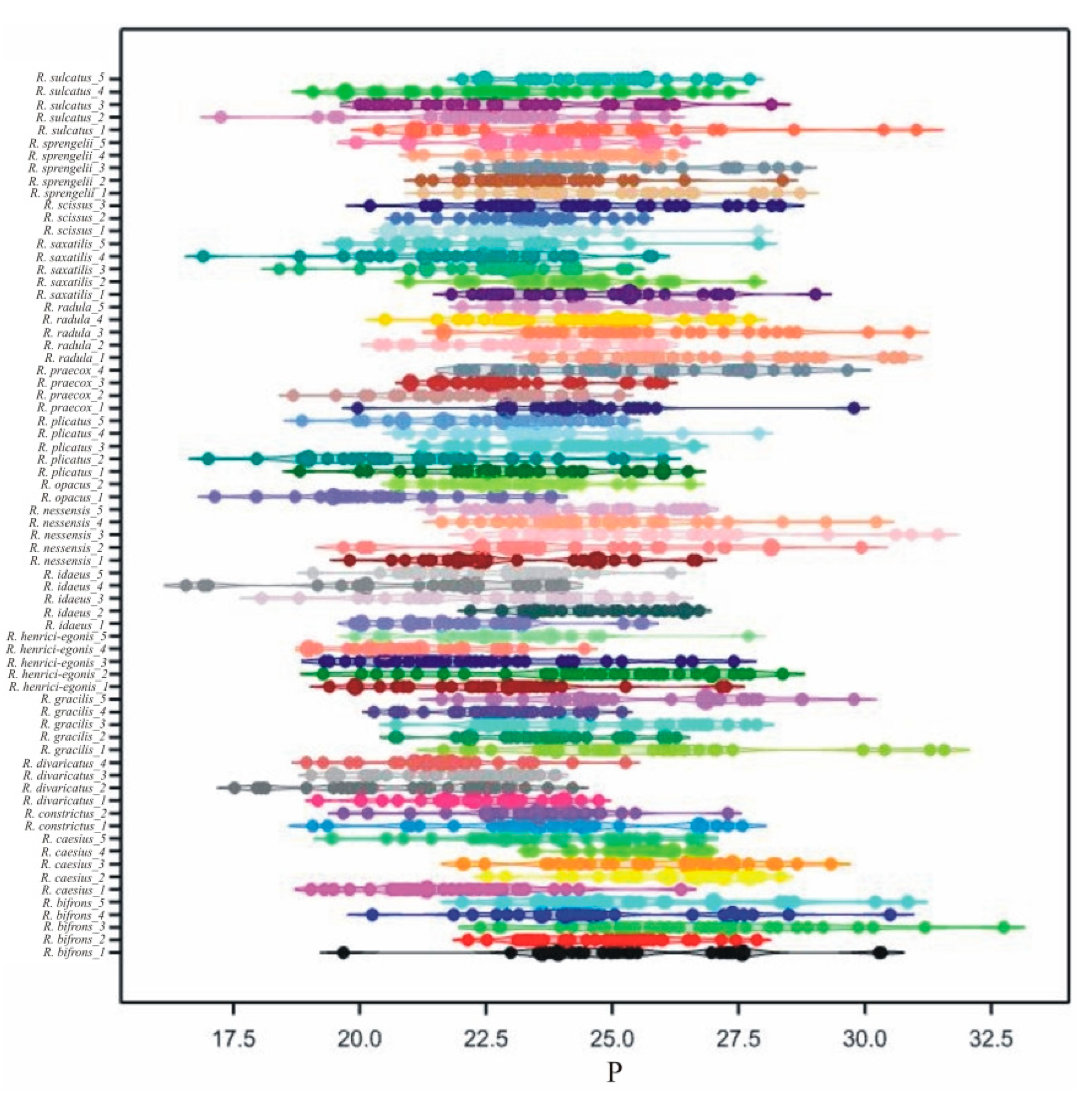
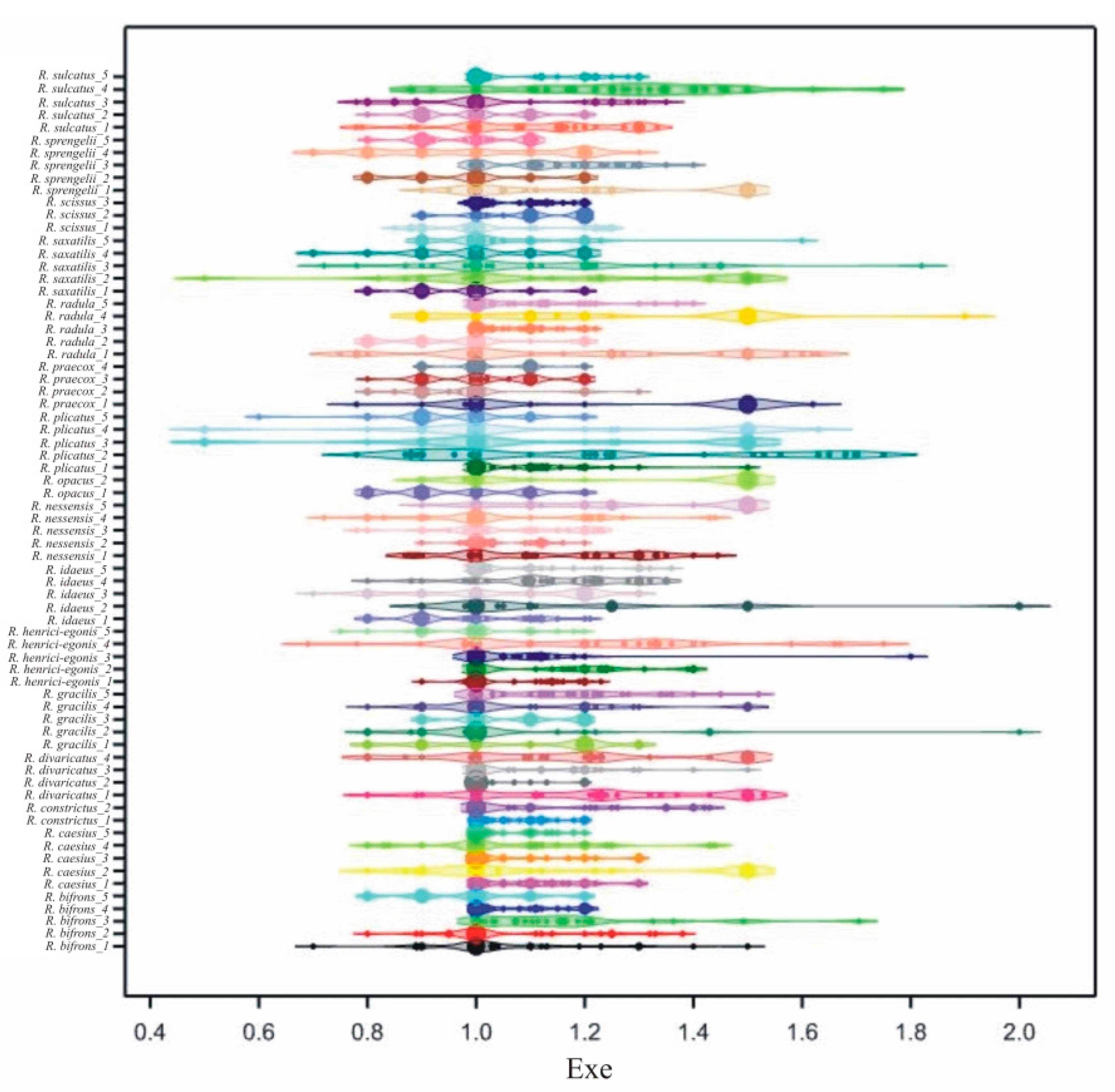
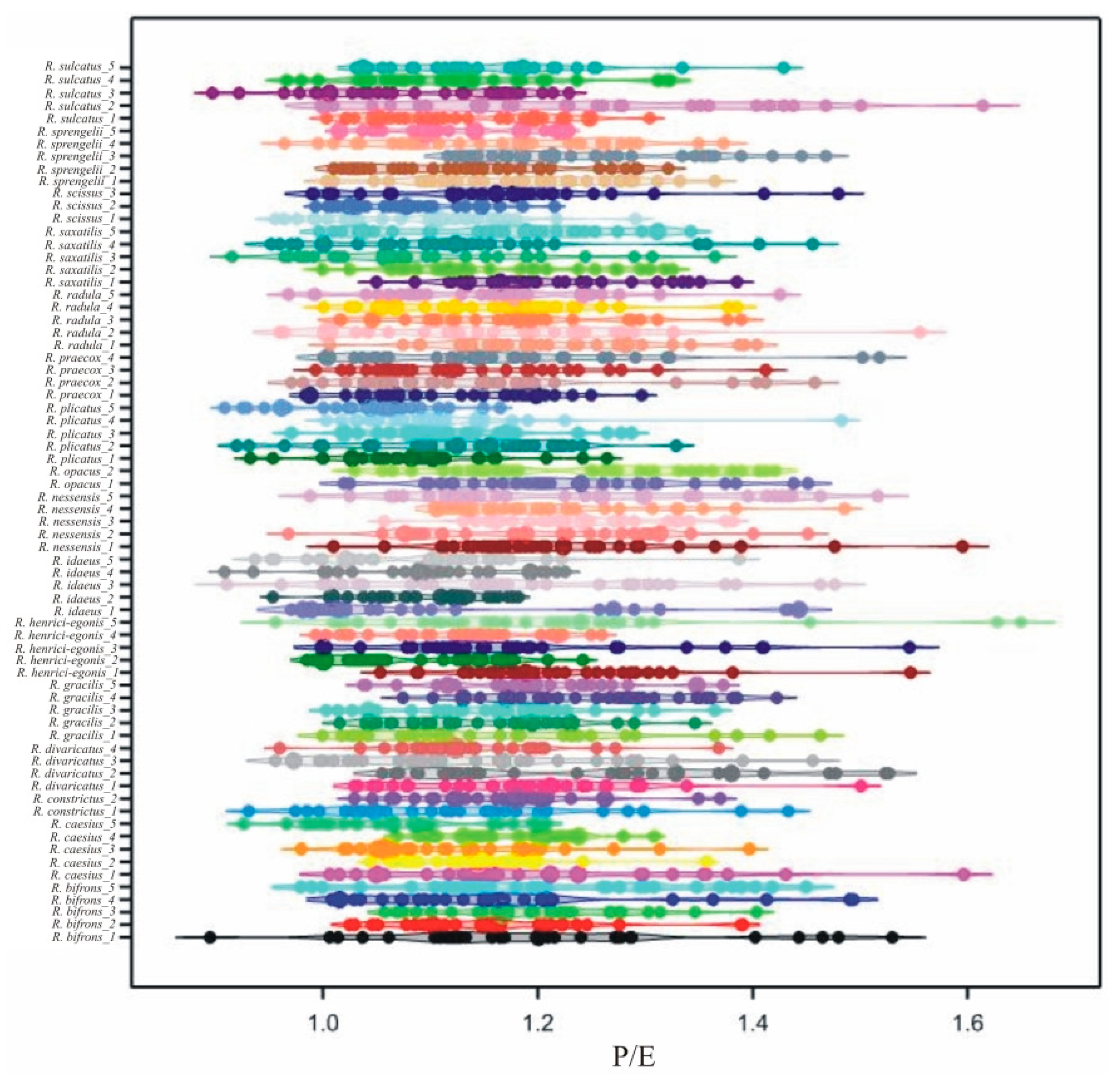
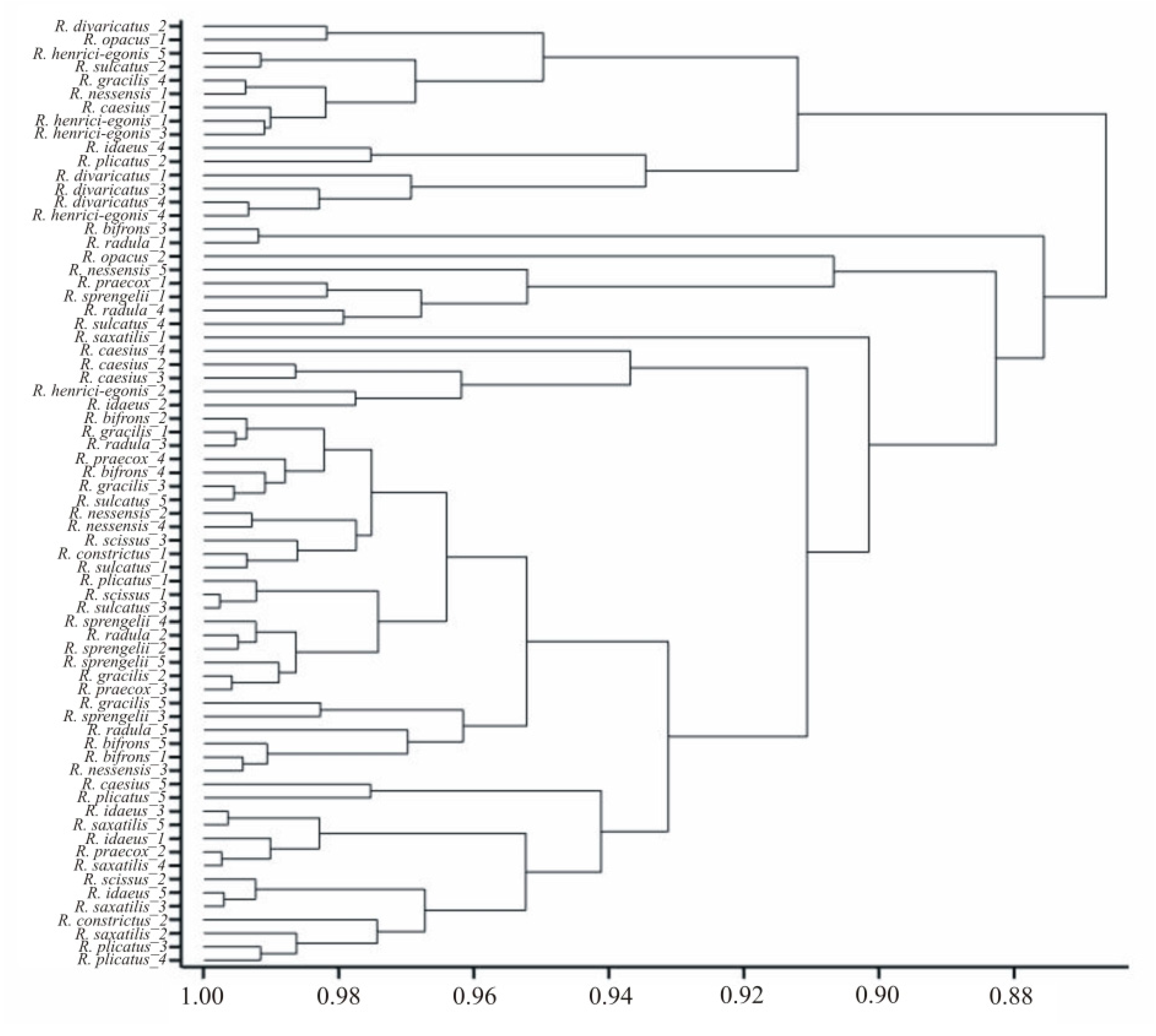
3.2. Interspecific Variability of Pollen Grains
3.3. Intraspecific Variability of Pollen Grains
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gustafsson, A. The genesis of the European blackberry flora. Acta Univ. Lund. 1943, 239, 1–200. [Google Scholar]
- Kurtto, A.; Weber, H.E.; Lampinen, R.; Sennikov, A.N. Atlas Florae Europaeae: Distribution of Vascular Plants in Europe: Rosaceae (Rubus); The Committee for Mapping the Flora of Europe & Societas Biologica Fennica Vanamo: Helskinki, Finland, 2010; 362p. [Google Scholar]
- Govaerts, R.; Nic Lughadha, E.; Black, N.; Turner, R.; Paton, A. The World Checklist of Vascular Plants, a continuously updated resource for exploring global plant diversity. Sci. Data 2021, 8, 215. [Google Scholar] [CrossRef] [PubMed]
- Focke, W.O. Synopsis Ruborum Germaniae: Die Deutschen Brombeerarten Ausführlich Beschrieben und Erläutert; CE Müllers’s Verlagsbuchhandlung: Bremen, Germany, 1877; 434p. [Google Scholar]
- Focke, W.O. Rosaceae. Die Natürlichen Pflanzenfamilien nebst ihren Gattungen und wichtigeren Arten insbesondere den Nutzpflanzen unter Mitwirkung zahlreicher hervorragender Fachgelehrten; Engler, A., Prantl, K., Eds.; Verlag von Wilhelm Engelmann: Leipzig, Germany, 1894; Volume 3, pp. 1–61. [Google Scholar]
- Jennings, D.L. Raspberries and Blackberries. Their Breeding, Diseases and Growth; Academic Press: London, UK, 1988; 230p. [Google Scholar]
- Robertson, K.R. The genera of Rosaceae in the southeastern United States. J. Arnold Arbor. 1974, 55, 352–360. [Google Scholar]
- Weber, H.E. Rubus L. Illustrierte Flora von Mitteleuropa IV/2a; Blackwell Wissenschafts-Verlag: Berlin, Germany, 1995; 595p. [Google Scholar]
- Alice, L.A.; Goldman, D.H.; Macklin, J.A.; Moore, G. Rubus Linnaeus. In Flora of North America North of Mexico; Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, UK; Oxford, UK, 2014; Volume 9, pp. 28–56. [Google Scholar]
- Zieliński, J. The genus Rubus (Rosaceae) in Poland. Pol. Bot. Stud. 2004, 16, 1–300. [Google Scholar]
- Kosiński, P.; Maliński, T.; Śliwińska, E.; Zieliński, J. Rubus prissanicus (Rosaceae), a new bramble species from North West Poland. Phytotaxa 2018, 344, 239–247. [Google Scholar] [CrossRef]
- Király, G. Alien Rubus species in Hungary: Distribution, habitats and threats. Dendrobiology 2018, 80, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Weber, H.E. Former and modern taxonomic treatment of the apomictic Rubus complex. Folia Geobot. Phytotax. 1996, 31, 373–380. [Google Scholar] [CrossRef]
- Alice, L.A.; Campbell, C.S. Phylogeny of Rubus (Rosaceae) based on nuclear ribosomal DNA internal transcribed spacer region sequences. Am. J. Bot. 1999, 86, 81–97. [Google Scholar] [CrossRef]
- Erdtman, G.; Berglund, B.; Praglowski, J. An Introduction to a Scandinavian Pollen Flora. Grana 1961, 2, 3–86. [Google Scholar] [CrossRef]
- Reitsma, T. Pollen morphology of some European Rosaceae. Acta Bot. Neerl. 1966, 15, 290–307. [Google Scholar] [CrossRef] [Green Version]
- Teppner, H. Zur Kenntnis der Gattung Waldsteinia L.—Schlüssel zum Bestimmen von Rosaceen Polleeinschliesslich ählicher Pollen—Formen aus andere Familien. Phyton 1966, 11, 224–238. [Google Scholar]
- Eide, F. Key for Northwest European Rosaceae pollen. Grana 1981, 20, 101–118. [Google Scholar] [CrossRef]
- Eide, F. On the pollen morphology of Rubus chamaemorus L. (Rosaceae). Grana 1981, 20, 25–27. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez Romano, M.L.; Candau, P.A. Contribution to palynological studies in the Rosaceae. Acta Bot. Malac. 1989, 14, 105–116. [Google Scholar] [CrossRef]
- Monasterio-Huelin, E.; Pardo, C. Pollen morphology and wall stratification in Rubus L. (Rosaceae) in the Iberian Peninsula. Grana 1995, 34, 229–236. [Google Scholar] [CrossRef]
- Tomlik-Wyremblewska, A. Pollen morphology of genus Rubus L. Part I. Introductory studies of the European representatives of the subgenus Rubus L. Acta Soc. Bot. Pol. 1995, 64, 187–203. [Google Scholar] [CrossRef] [Green Version]
- Wrońska-Pilarek, D.; Jagodziński, A.M.; Maliński, T. Morphological studies of pollen grains of the Polish endemic species of the genus Rubus L. (Rosaceae). Biologia 2012, 67, 87–96. [Google Scholar] [CrossRef]
- Jagodzinski, A.M.; Maciejewska-Rutkowska, I.; Wronska-Pilarek, D.; Bocianowski, J. Taxonomic significance of achene morphology of selected Rosa taxa (Rosaceae) occurring in Poland. Acta Soc. Bot. Pol. 2016, 85, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Xiong, X.; Zhou, X.; Li, M.; Xul, B.; Dengl, H.; Yul, Q.; Gao, X. Pollen morphology in Rubus (Rosaceae) and its taxonomic implications. Plant Syst. Evol. 2019, 305, 705–716. [Google Scholar] [CrossRef]
- Lechowicz, K.; Wrońska-Pilarek, D.; Bocianowski, J.; Maliński, T. Systematic importance of pollen morphological features of selected species from the genus Rubus (Rosaceae). PLoS ONE 2020, 15, e0221607. [Google Scholar] [CrossRef] [Green Version]
- Naruhashi, N.; Takano, H. Size variation of pollen grains in some Rubus species. J. Phytogeogr. Taxon. 1980, 28, 27–32. [Google Scholar]
- Kosenko, V.N.; Nguen, T.H.; Jacovlev, G.P. Palynomorphological study of the representatives of the genus Rubus (Rosaceae) in the flora of Vietnam. Bot. Z. 1982, 69, 497–503. [Google Scholar]
- Hebda, R.J.; Chinnappa, C.C. Studies on pollen morphology of Rosaceae in Canada. Rev. Palaeobot. Palynol. 1990, 64, 103–108. [Google Scholar] [CrossRef]
- Hebda, R.J.; Chinnappa, C.C. Studies on pollen morphology of Rosaceae. Bot. Lett. 1994, 141, 183–193. [Google Scholar]
- Ueda, Y. Pollen surface morphology in the genus Rosa, related genera. Jpn. J. Palynol. 1992, 38, 94–105. [Google Scholar]
- Tomlik-Wyremblewska, A. Pollen morphology of genus Rubus L., Part II. Introductory studies on the Malesian species of subgenus Micranthobatus. Acta Soc. Bot. Pol. 2000, 69, 31–40. [Google Scholar] [CrossRef]
- Li, W.L.; He, S.A.; Gu, Y.; Shu, P.; Pu, Z.M. Pollen morphology of the genus Rubus from China. Acta Phytotax. Sin. 2021, 39, 234–247. [Google Scholar]
- Tomlik-Wyremblewska, A.; Van der Ham, R.W.J.M.; Kosiński, P. Pollen morphology of genus Rubus L. Part III. Studies on the Malesian species of subgenera Chamaebatus L. and Idaeobatus L. Acta Soc. Bot. Pol. 2004, 73, 207–227. [Google Scholar] [CrossRef]
- Wrońska Pilarek, D.; Maliński, T.; Lira, J. Pollen morphology of Polish species of genus Rubus L.—Rubus gracilis J. Presl & C. Presl. Dendrobiology 2006, 56, 69–77. [Google Scholar]
- Wang, X.; Tang, H.; Huang, L.; He, Z.; Dong, X.; Fu, H.; Deng, Q. Comparative studies on pollen submicroscopic morphology of some wild species and cultivars of bramble (Rubus L.). Acta Hortic. Sin. 2007, 34, 1395–1404. [Google Scholar]
- Kasalkheh, R.; Jorjani, E.; Sabouri, H.; Habibi, M.; Sattarian, A. Pollen morphology of the genus Rubus L. subgenus Rubus (Rosaceae) in Iran. Nova Biol. Reper. 2017, 4, 9–18. [Google Scholar] [CrossRef]
- Gupta, C.; Dash, S.S. A new species of Rubus (Rosaceae) from Arunachal Pradesh, India. Blumea 2018, 63, 26–30. [Google Scholar] [CrossRef] [Green Version]
- Erdtman, G. The acetolysis method. A revised description. Sven Bot. Tidskr. 1960, 54, 561–564. [Google Scholar]
- Ueda, Y.; Tomita, H. Morphometric analysis of pollen patterns in Roses. Hort. J. 1989, 58, 211–220. [Google Scholar]
- Punt, W.; Hoen, P.P.; Blackmore, S.; Nilsson, S.; Le Thomas, A. Glossary of pollen and spore terminology. Rev. Palaeobot. Palynol. 2007, 1431, 1–81. [Google Scholar] [CrossRef]
- Halbritter, H.; Hess Ulrich, S.; Grímssonm, F.; Weber, M.; Zetter, R.; Hesse, M.; Buchner, R.; Svojtka, M.; Frosch-Radivo, A. Illustrated Pollen Terminology, 2nd ed.; Springer: Vienna, Austria, 2018; p. 483. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Seidler-Łożykowska, K.; Bocianowski, J. Evaluation of variability of morphological traits of selected caraway (Carum carvi L.) genotypes. Ind. Crops Prod. 2012, 35, 140–145. [Google Scholar] [CrossRef]
- Mahalanobis, P.C. On the generalized distance in statistics. Proc. Natl. Acad. Sci. India A 1936, 12, 49–55. [Google Scholar]
- Erdtman, G. Pollen Morphology and Plant Taxonomy. Angiosperms. An Introduction to Palynology; Almquist and Wiksell: Stockholm, Sweden, 1952; p. 365. [Google Scholar]
- APG IV (The Angiosperm Phylogeny Group); Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Potter, D.; Eriksson, T.; Evans, R.C.; Oh, S.; Smedmark, J.E.E.; Morgan, D.R.; Kerr, M.; Robertson, K.R.; Arsenault, M.; Dickinson, T.A.; et al. Phylogeny and classification of Rosaceae. Plant Syst. Evol. 2007, 266, 5–43. [Google Scholar] [CrossRef]
- Plants of the World Online. Available online: https://powo.science.kew.org (accessed on 10 October 2022).
- Ueda, Y.; Okada, Y. Discrimination of rose cultivar groups by pollen surface structure. J. Hortic. Sci. 1994, 69, 601–607. [Google Scholar] [CrossRef]
- Ghosh, A.; Saha, I. Pollen morphological study of some selected Indian taxa of Rosaceae. Indian J. Appl. Pure Bio 2017, 32, 121–130. [Google Scholar]
- Candau, P.; Romanos, L.G. Rosaceae. In Atlas Polinico de Andalucla Occidental; Valdis, B., Diez, M.J., Fernin-Dez, I., Eds.; Instituto de Desarrollo Regional, Universidad de Sevilla, Excma. Diputacion de Cadiz: Cidiz, Spain, 1987; pp. 179–184. [Google Scholar]
- Wronska-Pilarek, D.; Jagodzinski, A.M. Pollen morphological variability of Polish native species of Rosa L. (Rosaceae). Dendrobiology 2009, 62, 71–82. [Google Scholar]
- Singh, K.; Sharma, Y.P.; Sharma, P.R.; Gairola, S. Pollen morphology and variability of the Rosa L. species of Western Himalaya in India. Genet. Resour. Crop Evol. 2020, 67, 2129–2148. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Species | Subgenus | Section | Subsection | Series |
|---|---|---|---|---|---|
| 1 | R. bifrons Vest | Rubus | Rubus | Hiemales | Discolores |
| 2 | R. caesius L. | Rubus | Caesii | - | - |
| 3 | R. constrictus P. J. Müll. & Lefévre | Rubus | Rubus | Rubus | Rubus |
| 4 | R. divaricatus P. J. Müll. | Rubus | Rubus | Rubus | Rubus |
| 5 | R. gracilis J. Presl & C. Presl | Rubus | Rubus | Hiemales | Rhamnifolii |
| 6 | R. henrici-egonis Holub | Rubus | Rubus | Hiemales | Discolores |
| 7 | R. idaeus L. | Idaeobatus | - | - | - |
| 8 | R. nessensis W. Hall | Rubus | Rubus | Rubus | Nessenses |
| 9 | R. opacus Focke | Rubus | Rubus | Rubus | Rubus |
| 10 | R. plicatus Weihe & Nees | Rubus | Rubus | Rubus | Rubus |
| 11 | R. praecox Bertol. | Rubus | Rubus | Hiemales | Discolores |
| 12 | R. radula Weihe | Rubus | Rubus | Hiemales | Radulae |
| 13 | R. saxatilis L. | Cylactis | - | - | Saxatiles |
| 14 | R. scissus W. C. R. Watson | Rubus | Rubus | Rubus | Nessenses |
| 15 | R. sprengelii Weihe | Rubus | Rubus | Hiemales | Sprengeliani |
| 16 | R. sulcatus Vest | Rubus | Rubus | Rubus | Rubus |
| No. | Species | Localities | Geographical Coordinates | Collector, Herbarium |
|---|---|---|---|---|
| 1 | R. bifrons 1 | Poland, Kietrz | 50°04′46.7″ N, 18°00′07.6″ E | Kosiński, Tomaszewski, Zieliński; KOR |
| 2 | R. bifrons 2 | Poland, Mszana Dolna | 49°40′51.9″ N, 20°04′35.4″ E | Białobok; KOR |
| 3 | R. bifrons 3 | Poland, Braszowice | 50°33′15.3″ N, 16°47′53.2″ E | Kosiński; KOR |
| 4 | R. bifrons 4 | Poland, Faliszówka | 49°38′39.5″ N, 21°36′30.1″ E | Oklejewicz; KOR |
| 5 | R. bifrons 5 | Poland, Pomorzowice | 50°17′02.1″ N, 17°45′33.4″ E | Tomlik, Zieliński; KOR |
| 6 | R. caesius 1 | Poland, Skomętno Wielkie | 53°51′00.0″ N, 22°37′29.9″ E | Tomlik; KOR |
| 7 | R. caesius 2 | Poland, Wiewiórczyn | 51°36′24.9″ N, 19°07′23.9″ E | Stefanek; KOR |
| 8 | R. caesius 3 | Poland, Pilchowice | 50°12′57.8″ N, 18°33′49.3″ E | Tomlik, Zieliński; KOR |
| 9 | R. caesius 4 | Poland, Piła-Młyn | 53°30′52.0″ N, 17°53′17.0″ E | Stefanek; KOR |
| 10 | R. caesius 5 | Poland, Dzierżysław | 50°02′51.6″ N, 17°58′09.1″ E | Tomlik, Zieliński; KOR |
| 11 | R. constrictus 1 | Poland, Ząbkowice Śląskie | 50°35′21.4″ N, 16°48′40.2″ E | Kosiński; KOR |
| 12 | R. constrictus 2 | Poland, Staniszów | 50°50′24.0″ N, 15°44′13.1″ E | Oklejewicz; KOR |
| 13 | R. divaricatus 1 | Poland, Lubin | 51°23′49.1″ N, 16°12′20.0″ E | Zieliński; KOR |
| 14 | R. divaricatus 2 | Poland, Moryń | 52°48′40.0″ N, 14°21′10.0″ E | Maliński; KOR |
| 15 | R. divaricatus 3 | Poland, Antonin | 51°30′59.1″ N, 17°51′16.9″ E | Zieliński; KOR |
| 16 | R. divaricatus 4 | Germany, Wiednitz | 51°23′14.1″ N, 14°01′45.6″ E | Hans-Werner; KOR |
| 17 | R. gracilis 1 | Poland, Pątnów | 51°08′47.3″ N, 18°37′29.3″ E | Zieliński; KOR |
| 18 | R. gracilis 2 | Poland, Świdwin | 53°46′28.1″ N, 15°46′38.8″ E | Kosiński; KOR |
| 19 | R. gracilis 3 | Poland, Kutyły | 50°34′51.5″ N, 22°15′23.5″ E | Oklejewicz; KOR |
| 20 | R. gracilis 4 | Poland, Marcinów | 50°23′06.7″ N, 16°41′19.9″ E | Kosiński; KOR |
| 21 | R. gracilis 5 | Poland, Starkowo | 51°58′37.8″ N, 16°18′35.7″ E | Zieliński; KOR |
| 22 | R. henrici-egonis 1 | Poland, Ząbkowice Śląskie | 50°35′21.4″ N, 16°48′40.2″ E | Kosiński; KOR |
| 23 | R. henrici-egonis 2 | Poland, Kłodzko | 50°26′17.8″ N, 16°39′16.7″ E | Kosiński; KOR |
| 24 | R. henrici-egonis 3 | Poland, Bardo | 50°30′24.3″ N, 16°44′24.9″ E | Kosiński; KOR |
| 25 | R. henrici-egonis 4 | Poland, Gorzyce | 49°57′30.6″ N, 18°23′45.4″ E | Kosiński, Tomaszewski, Zieliński; KOR |
| 26 | R. henrici-egonis 5 | Poland, Szklary | 49°27′58.7″ N, 21°50′28.2″ E | Kosiński, Zieliński; KOR |
| 27 | R. idaeus 1 | Poland, Pamiątkowo | 52°33′15.1″ N, 16°40′56.9″ E | Tomlik, KOR |
| 28 | R. idaeus 2 | Poland, Kórnik | 52°14′57.4″ N, 17°05′24.5″ E | Zieliński, KOR |
| 29 | R. idaeus 3 | Poland, Lubowidz | 53°07′18.0″ N, 19°50′26.9″ E | Anisimowicz; KOR |
| 30 | R. idaeus 4 | Poland, Pakosław | 51°37′00.0″ N, 17°03′26.8″ E | Kortus; KOR |
| 31 | R. idaeus 5 | Poland, Wąbrzeźno | 53°16′46.7″ N, 18°56′52.0″ E | Stempola; KOR |
| 32 | R. nessensis 1 | Poland, Kozielec | 53°15′09.3″ N, 18°13′51.2″ E | Krawiec; KOR |
| 33 | R. nessensis 2 | Poland, Wolsztyn | 52°07′02.1″ N, 16°06′45.6″ E | Paczoski; KOR |
| 34 | R. nessensis 3 | Poland, Prabuty | 53°45′21.6″ N, 19°12′16.0″ E | Boratyński, Zieliński; KOR |
| 35 | R. nessensis 4 | Poland, Wyspowo | 54°33′51.0″ N, 18°18′15.9″ E | Markowski; KOR |
| 36 | R. nessensis 5 | Poland, Brodnica | 53°15′28.9″ N, 19°24′19.6″ E | Boratyński, Zieliński; KOR |
| 37 | R. opacus 1 | Poland, Poźrzadło | 52°18′30.0″ N, 15°15′30.0″ E | Zieliński; KOR |
| 38 | R. opacus 2 | Poland, Starkowo | 51°58′37.8″ N, 16°18′35.7″ E | Zieliński; KOR |
| 39 | R. plicatus 1 | Poland, Karnocice | 53°51′34.5″ N, 14°31′04.3″ E | Piotrowska; KOR |
| 40 | R. plicatus 2 | Poland, Sarbsk | 54°45′07.2″ N, 17°40′02.7″ E | Zieliński; KOR |
| 41 | R. plicatus 3 | Poland, Karkonosze | 50°46′35.2″ N, 15°36′18.9″ E | Boratyńscy; KOR |
| 42 | R. plicatus 4 | Poland, Jastrzębia Góra | 54°49′49.6″ N, 18°18′15.9″ E | Piotrowska; KOR |
| 43 | R. plicatus 5 | Poland, Kamienna Góra | 50°47′00.3″ N, 16°01′59.2″ E | Boratyński, Zieliński; KOR |
| 44 | R. praecox 1 | Austria, Klausen-Leopoldsdorf | 48°06′27.0″ N, 16°01′52.7″ E | Kosiński; KOR |
| 45 | R. praecox 2 | Germany, Berlin | 52°29′03.0″ N, 13°14′08.0″ E | Mackiewicz; KOR |
| 46 | R. praecox 3 | Poland, Burkatów | 50°47′42.1″ N, 16°27′59.6″ E | Boratyński; KOR |
| 47 | R. praecox 4 | Bulgaria, Blagoevgrad | 42°00′29.9″ N, 23°05′28.5″ E | Boratyński, Browicz, Zieliński; KOR |
| 48 | R. radula 1 | Poland, Potakówka | 49°42′47.2″ N, 21°36′06.7″ E | Oklejewicz; KOR |
| 49 | R. radula 2 | Poland, Boszkowo | 51°58′28.1″ N, 16°19′41.9″ E | Hantz; KOR |
| 50 | R. radula 3 | Poland, Psary | 51°33′44.2″ N, 21°37′09.9″ E | Danielewicz, Maliński, Zieliński; KOR |
| 51 | R. radula 4 | Poland, Boguszyn | 51°56′36.5″ N, 16°28′16.4″ E | Danielewicz, Maliński; KOR |
| 52 | R. radula 5 | Poland, Postomino | 51°58′28.1″ N, 16°19′41.9″ E | Zieliński; KOR |
| 53 | R. saxatilis 1 | Germany, Wald | 47°43′20.9″ N, 10°33′24.8″ E | Hoffmann, KOR |
| 54 | R. saxatilis 2 | Poland, Promno | 52°27′03.0″ N, 17°14′42.9″ E | Browicz; KOR |
| 55 | R. saxatilis 3 | Poland, Lublin | 51°15′00.8″ N, 22°34′19.3″ E | Fijałkowski; KOR |
| 56 | R. saxatilis 4 | Poland, Błażejewo | 54°13′45.6″ N, 22°18′19.5″ E | Bugała; KOR |
| 57 | R. saxatilis 5 | Poland, Bodzewko | 51°49′56.0″ N, 17°06′19.9″ E | Kaczmarek; KOR |
| 58 | R. scissus 1 | Poland, Zielin | 54°14′47.9″ N, 17°05′58.9″ E | Zieliński, KOR |
| 59 | R. scissus 2 | Poland, Słowiński National Park | 54°41′57.3″ N, 17°19′10.9″ E | Zieliński; KOR |
| 60 | R. scissus 3 | Poland, Rudniki | 51°02′13.0″ N, 18°36′01.6″ E | Zieliński; KOR |
| 61 | R. sprengelii 1 | Poland, Wolin | 53°50′36.0″ N, 14°36′57.1″ E | Zieliński; KOR |
| 62 | R. sprengelii 2 | Poland, Ciechnowo | 53°51′34.1″ N, 15°44′20.9″ E | Zieliński; KOR |
| 63 | R. sprengelii 3 | Poland, Słowieńsko | 53°51′56.9″ N, 15°38′24.7″ E | Zieliński; KOR |
| 64 | R. sprengelii 4 | Poland, Kłoczewo | 53°57′56.2″ N, 14°36′57.1″ E | Boratyńska, Dolatowska, Zieliński; KOR |
| 65 | R. sprengelii 5 | Poland, Dobiegniew | 52°58′05.5″ N, 15°45′17.4″ E | Boratyńska, Dolatowska, Zieliński; KOR |
| 66 | R. sulcatus 1 | Poland, Bystrzyca Kłodzka | 50°17′48.7″ N, 16°39′07.3″ E | Kosiński; KOR |
| 67 | R. sulcatus 2 | Poland, Dydnia | 49°41′15.6″ N, 22°10′16.8″ E | Oklejewicz; KOR |
| 68 | R. sulcatus 3 | Poland, Poraż | 49°29′05.5″ N, 22°12′55.5″ E | Oklejewicz; KOR |
| 69 | R. sulcatus 4 | Poland, Jaśliska | 49°26′29.9″ N, 21°48′08.5″ E | Kaczmarek; KOR |
| 70 | R. sulcatus 5 | Poland, Słowik | 50°49′54.4″ N, 20°32′20.3″ E | Kaznowski; KOR |
| Trait | P | Le | d | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. | Species | Mean | Homogeneous Groups | Min | Max | s.d. | Mean | Homogeneous Groups | Min | Max | s.d. | Mean | Homogeneous Groups | Min | Max | s.d. |
| 1 | R. bifrons_1 | 25.53 | a--e | 19.69 | 30.33 | 2.206 | 21.22 | c-j | 18.28 | 25.97 | 2.039 | 2.36 | t-zA | 1 | 4 | 0.679 |
| 2 | R. bifrons_2 | 24.84 | b-i | 22.14 | 27.86 | 1.474 | 20.89 | c-m | 17.93 | 24.58 | 1.709 | 2.968 | f-w | 2 | 5.2 | 0.807 |
| 3 | R. bifrons_3 | 26.69 | a | 22.39 | 32.75 | 2.495 | 22.98 | ab | 18.8 | 29.08 | 2.511 | 2.842 | h-x | 1.5 | 4.13 | 0.702 |
| 4 | R. bifrons_4 | 24.78 | b-i | 20.25 | 30.5 | 2.196 | 21.09 | c-k | 17.47 | 26.25 | 2.083 | 2.58 | p-z | 1.4 | 5.2 | 0.923 |
| 5 | R. bifrons_5 | 25.33 | a-f | 22.01 | 30.85 | 2.185 | 21.31 | c-j | 16.67 | 26.83 | 2.235 | 2.531 | q-zA | 1.7 | 3.84 | 0.605 |
| 6 | R. caesius_1 | 21.89 | w-zABC | 19.03 | 26.36 | 1.643 | 18.26 | v-y | 14.63 | 22.67 | 1.736 | 2.347 | u-zA | 1.17 | 3.8 | 0.672 |
| 7 | R. caesius_2 | 26.15 | ab | 22.53 | 28.32 | 1.465 | 22.38 | a-c | 19.07 | 24.27 | 1.429 | 3.85 | bc | 2.35 | 5.8 | 0.964 |
| 8 | R. caesius_3 | 25.72 | a-c | 22.02 | 29.33 | 1.932 | 21.75 | a-d | 17.18 | 26.52 | 2.346 | 3.77 | b-d | 2 | 6.3 | 1.102 |
| 9 | R. caesius_4 | 25.58 | a-e | 23.32 | 26.91 | 0.945 | 21.03 | c-k | 18.07 | 24.19 | 1.474 | 4.553 | a | 3.04 | 6.22 | 0.829 |
| 10 | R. caesius_5 | 23.68 | g-v | 19.45 | 26.75 | 1.85 | 19.83 | g-v | 15.87 | 23.15 | 2.047 | 4.075 | ab | 2.8 | 5.35 | 0.664 |
| 11 | R. constrictus_1 | 23.89 | f-u | 19.08 | 27.58 | 2.264 | 20.1 | e-s | 15.19 | 24.23 | 2.401 | 3.283 | c-o | 1.9 | 6 | 1.079 |
| 12 | R. constrictus_2 | 23.46 | h-w | 19.67 | 27.28 | 1.517 | 19.71 | j-v | 14.73 | 22.95 | 1.657 | 2.734 | k-z | 1.4 | 4.94 | 0.886 |
| 13 | R. divaricatus_1 | 22.34 | u-zAB | 19.17 | 24.74 | 1.361 | 19.15 | p-y | 15.4 | 22.08 | 1.513 | 1.834 | A | 1 | 3 | 0.464 |
| 14 | R. divaricatus_2 | 21.01 | BC | 17.52 | 24.21 | 1.84 | 17.7 | Y | 14 | 20.36 | 1.803 | 2.066 | zA | 1.24 | 3.6 | 0.573 |
| 15 | R. divaricatus_3 | 21.76 | x-zABC | 19.04 | 23.89 | 1.445 | 18.17 | v-y | 13.88 | 20.5 | 1.787 | 2.088 | yzA | 1.07 | 3.39 | 0.594 |
| 16 | R. divaricatus_4 | 21.72 | x-zABC | 18.96 | 25.26 | 1.349 | 18.19 | v-y | 14.96 | 21.59 | 1.49 | 2.047 | zA | 1 | 4.02 | 0.678 |
| 17 | R. gracilis_1 | 25.63 | a-d | 21.65 | 31.57 | 2.563 | 21.65 | a-e | 17.47 | 27.66 | 2.636 | 3.098 | d-s | 1.96 | 6.3 | 1.064 |
| 18 | R. gracilis_2 | 23.7 | g-v | 20.69 | 26.27 | 1.431 | 20.33 | d-s | 17.45 | 23.08 | 1.587 | 2.581 | o-z | 1.55 | 5 | 0.834 |
| 19 | R. gracilis_3 | 24.69 | b-j | 20.74 | 27.86 | 1.945 | 21.41 | b-g | 17.62 | 25.49 | 2.228 | 3 | f-u | 1.5 | 5.35 | 0.950 |
| 20 | R. gracilis_4 | 22.74 | o-z | 20.28 | 25.19 | 1.221 | 19.29 | m-y | 16.37 | 21.85 | 1.265 | 2.442 | r-zA | 1.45 | 5 | 0.731 |
| 21 | R. gracilis_5 | 25.46 | a-f | 21.63 | 29.79 | 2.163 | 21.23 | c-j | 16.47 | 25.93 | 2.301 | 2.72 | k-z | 1.6 | 4.9 | 0.859 |
| 22 | R. henrici-egonis_1 | 22.67 | r-A | 19.41 | 27.25 | 1.846 | 18.84 | r-y | 14.75 | 23.28 | 1.788 | 2.266 | w-zA | 1.29 | 4.6 | 0.642 |
| 23 | R. henrici-egonis_2 | 24.42 | c-m | 19.27 | 28.37 | 2.488 | 20.87 | c-n | 15.19 | 24.51 | 2.448 | 3.722 | b-e | 2.3 | 5.5 | 0.930 |
| 24 | R. henrici-egonis_3 | 22.42 | t-zAB | 19.31 | 27.41 | 2.162 | 18.91 | q-y | 15.06 | 26.36 | 2.442 | 2.151 | x-zA | 1.3 | 3.2 | 0.406 |
| 25 | R. henrici-egonis_4 | 21.23 | zABC | 18.97 | 24.47 | 1.356 | 17.92 | w-y | 15.22 | 20.85 | 1.332 | 2.288 | v-zA | 0.94 | 4.8 | 0.864 |
| 26 | R. henrici-egonis_5 | 22.7 | q-zA | 19.91 | 27.71 | 1.614 | 19.26 | n-y | 15.8 | 23.93 | 1.596 | 2.614 | o-z | 1.22 | 4.02 | 0.658 |
| 27 | R. idaeus_1 | 22.16 | v-zAB | 19.89 | 25.61 | 1.518 | 19.23 | o-y | 16.34 | 23.22 | 1.866 | 3.444 | b-j | 2 | 5.7 | 1.062 |
| 28 | R. idaeus_2 | 24.62 | b-l | 22.19 | 26.72 | 1.288 | 20.32 | d-s | 16.14 | 22.88 | 1.504 | 4.053 | ab | 3.08 | 5.06 | 0.534 |
| 29 | R. idaeus_3 | 22.81 | n-z | 18.07 | 26.18 | 2.123 | 19.15 | p-y | 12.85 | 21.78 | 1.992 | 3.076 | d-s | 2.05 | 4.6 | 0.671 |
| 30 | R. idaeus_4 | 21.39 | yzABC | 16.56 | 24 | 2.293 | 18.39 | t-y | 14.02 | 21.23 | 2.121 | 3.321 | c-n | 2.03 | 4.21 | 0.669 |
| 31 | R. idaeus_5 | 22.52 | t-zAB | 19.07 | 26.17 | 1.588 | 19.12 | p-y | 14.63 | 22.89 | 1.673 | 3.542 | b-h | 2.5 | 4.75 | 0.569 |
| 32 | R. nessensis_1 | 23.1 | k-x | 19.8 | 26.69 | 1.827 | 19.6 | k-v | 16.47 | 23.85 | 2.135 | 2.549 | p-z | 1 | 4.98 | 0.828 |
| 33 | R. nessensis_2 | 24 | e-t | 19.67 | 29.93 | 2.651 | 20.26 | d-s | 16.4 | 25.82 | 2.546 | 3.07 | d-s | 1.6 | 5.51 | 1.055 |
| 34 | R. nessensis_3 | 25.63 | a-d | 22.19 | 31.45 | 2.488 | 21.34 | c-i | 17.74 | 26.26 | 2.168 | 2.597 | o-z | 1.3 | 6.13 | 0.970 |
| 35 | R. nessensis_4 | 24.9 | b-h | 21.62 | 30.23 | 2.076 | 20.82 | c-o | 16.72 | 25.64 | 2.014 | 2.987 | f-v | 1.29 | 5.56 | 1.064 |
| 36 | R. nessensis_5 | 24.31 | c-o | 21.41 | 26.82 | 1.539 | 20.34 | d-s | 17.8 | 24.28 | 1.769 | 2.44 | r-zA | 1.5 | 4.03 | 0.632 |
| 37 | R. opacus_1 | 20.54 | C | 17.13 | 23.8 | 1.697 | 17.73 | xy | 15.58 | 20.9 | 1.371 | 2.147 | x-zA | 1.5 | 2.86 | 0.335 |
| 38 | R. opacus_2 | 22.96 | m-y | 20.73 | 26.54 | 1.393 | 18.46 | t-y | 13.93 | 22.66 | 1.866 | 3.061 | e-t | 1.8 | 5.51 | 0.915 |
| 39 | R. plicatus_1 | 23.11 | j-x | 18.84 | 26.5 | 1.913 | 19.75 | i-v | 16.05 | 23.34 | 1.837 | 2.638 | m-z | 1.56 | 4.24 | 0.652 |
| 40 | R. plicatus_2 | 21.15 | ABC | 17 | 26 | 2.173 | 18.36 | u-y | 14.23 | 23.12 | 2.2 | 2.833 | i-x | 1.77 | 4.54 | 0.758 |
| 41 | R. plicatus_3 | 24.29 | c-p | 21.25 | 26.62 | 1.425 | 19.74 | i-v | 17.11 | 21.98 | 1.23 | 3.138 | d-r | 1.5 | 6 | 1.206 |
| 42 | R. plicatus_4 | 23.46 | h-w | 20.75 | 27.9 | 1.587 | 19.33 | l-x | 16.05 | 22.24 | 1.392 | 3.333 | c-m | 2 | 6.2 | 0.931 |
| 43 | R. plicatus_5 | 22.7 | q-zA | 18.85 | 25.21 | 1.666 | 19.3 | l-y | 16.52 | 21.58 | 1.56 | 3.653 | b-f | 2 | 5.65 | 0.975 |
| 44 | R. praecox_1 | 24.29 | c-p | 19.98 | 29.78 | 1.533 | 20.7 | d-p | 16.53 | 24.14 | 1.459 | 2.811 | j-x | 1.24 | 5.73 | 1.002 |
| 45 | R. praecox_2 | 22.27 | v-zAB | 18.68 | 25.14 | 1.576 | 18.87 | r-y | 16.11 | 21.4 | 1.439 | 3.384 | b-l | 2 | 5.52 | 0.734 |
| 46 | R. praecox_3 | 23.04 | l-x | 21 | 26.01 | 1.453 | 19.78 | h-v | 17.23 | 23.14 | 1.427 | 2.7 | l-z | 1.87 | 4.97 | 0.673 |
| 47 | R. praecox_4 | 25.36 | a-f | 21.97 | 29.66 | 2.275 | 21.77 | a-d | 18.56 | 26.1 | 2.129 | 2.979 | f-v | 2 | 5.09 | 0.694 |
| 48 | R. radula_1 | 26.76 | a | 23.43 | 30.75 | 2.226 | 23.15 | a | 20.4 | 28.63 | 2.154 | 2.428 | s-zA | 1.32 | 3.66 | 0.639 |
| 49 | R. radula_2 | 23.63 | g-v | 20.36 | 25.98 | 1.743 | 20.09 | e-s | 14.52 | 22.91 | 1.943 | 2.62 | n-z | 1.94 | 3.7 | 0.425 |
| 50 | R. radula_3 | 25.63 | a-d | 21.66 | 30.87 | 2.267 | 21.29 | c-j | 15.15 | 26.36 | 2.525 | 3.153 | c-q | 1.5 | 6.25 | 1.208 |
| 51 | R. radula_4 | 24.27 | c-q | 20.47 | 27.72 | 1.825 | 19.98 | f-u | 16.91 | 22.94 | 1.586 | 3.324 | c-m | 2.05 | 5.7 | 0.922 |
| 52 | R. radula_5 | 25.05 | b-g | 22.04 | 27.21 | 1.436 | 21.25 | c-j | 17.95 | 24.22 | 1.838 | 2.153 | x-zA | 1.4 | 5.5 | 0.844 |
| 53 | R. saxatilis_1 | 24.48 | c-m | 21.81 | 29.01 | 1.811 | 21.57 | a-f | 18.46 | 25.6 | 1.695 | 3.669 | b-f | 2.09 | 5.81 | 0.913 |
| 54 | R. saxatilis_2 | 24.12 | d-s | 20.96 | 27.8 | 1.487 | 20 | f-t | 15.46 | 23.13 | 1.917 | 3.421 | b-k | 2.04 | 5.44 | 0.892 |
| 55 | R. saxatilis_3 | 22.63 | s-zA | 18.41 | 25.3 | 1.643 | 19.24 | o-y | 14.58 | 21.83 | 1.659 | 3.345 | c-l | 1.99 | 5.54 | 0.865 |
| 56 | R. saxatilis_4 | 22.22 | v-zAB | 16.9 | 25.8 | 1.942 | 18.8 | s-y | 15.04 | 21.8 | 1.664 | 3.544 | b-h | 2 | 5.29 | 0.954 |
| 57 | R. saxatilis_5 | 22.54 | s-zAB | 19.63 | 27.91 | 1.732 | 19.15 | p-y | 15.86 | 23.4 | 1.749 | 3.239 | c-p | 2.29 | 4.53 | 0.590 |
| 58 | R. scissus_1 | 22.73 | p-zA | 20.53 | 27.89 | 1.625 | 19.18 | p-y | 14.81 | 22.42 | 1.695 | 2.597 | o-z | 1.6 | 3.75 | 0.607 |
| 59 | R. scissus_2 | 23.29 | i-x | 20.72 | 25.61 | 1.112 | 19.8 | g-v | 17.23 | 24.11 | 1.39 | 3.529 | b-i | 1.8 | 6.4 | 1.190 |
| 60 | R. scissus_3 | 24.36 | c-n | 20.2 | 28.34 | 2.183 | 20.64 | d-p | 16.73 | 25.14 | 1.995 | 3.576 | b-g | 2 | 5.8 | 1.044 |
| 61 | R. sprengelii_1 | 24.8 | b-i | 21.27 | 28.72 | 1.919 | 21.15 | c-k | 18.24 | 25.31 | 1.961 | 3.222 | c-q | 2 | 5.61 | 0.867 |
| 62 | R. sprengelii_2 | 23.5 | g-v | 21.2 | 28.36 | 1.509 | 19.75 | i-v | 16.64 | 24.38 | 1.745 | 2.774 | j-y | 1.7 | 5 | 0.683 |
| 63 | R. sprengelii_3 | 24.74 | b-i | 22 | 28.65 | 1.975 | 20.91 | c-l | 17.65 | 25.83 | 2.192 | 2.707 | l-z | 1.76 | 5.1 | 0.843 |
| 64 | R. sprengelii_4 | 24.22 | c-r | 21.06 | 26.2 | 1.457 | 20.52 | d-q | 15.45 | 23.02 | 1.58 | 2.997 | f-u | 2 | 4.4 | 0.652 |
| 65 | R. sprengelii_5 | 23.58 | g-v | 19.9 | 26.43 | 1.69 | 20.45 | d-r | 16.9 | 23.55 | 1.83 | 2.9 | g-w | 2 | 5.5 | 0.863 |
| 66 | R. sulcatus_1 | 24.34 | c-n | 20.38 | 31.02 | 2.734 | 20.41 | d-s | 16.8 | 25.21 | 2.285 | 3.011 | f-u | 2 | 5.73 | 0.911 |
| 67 | R. sulcatus_2 | 22.46 | t-zAB | 17.25 | 26.04 | 1.859 | 19.31 | l-y | 14.38 | 22.76 | 1.931 | 2.696 | l-z | 2 | 3.99 | 0.598 |
| 68 | R. sulcatus_3 | 23.1 | k-x | 20 | 28.15 | 2.2 | 19.55 | k-v | 15 | 25.37 | 2.161 | 2.78 | j-y | 1.5 | 4.33 | 0.799 |
| 69 | R. sulcatus_4 | 23.09 | k-x | 19.07 | 27.31 | 2.234 | 19.38 | l-w | 15.8 | 23.36 | 2.148 | 2.927 | g-w | 1.51 | 4.8 | 0.979 |
| 70 | R. sulcatus_5 | 24.66 | b-k | 22.02 | 27.72 | 1.45 | 21.38 | b-h | 17.99 | 25.2 | 1.668 | 2.734 | k-z | 1.45 | 4.72 | 0.786 |
| LSD0.001 | 1.581 | 1.618 | 0.702 | |||||||||||||
| F-ANOVA | 17.62 *** | 12.47 *** | 13.24 *** | |||||||||||||
| Trait | E | Exp | P/E | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. | Species | Mean | Homogeneous Groups | Min | Max | s.d. | Mean | Homogeneous Groups | Min | Max | s.d. | Mean | Homogeneous Groups | Min | Max | s.d. |
| 1 | R. bifrons_1 | 21.4 | a-m | 16.21 | 24.92 | 2.167 | 1.058 | g-q | 0.7 | 1.5 | 0.163 | 1.204 | a-l | 0.895 | 1.530 | 0.150 |
| 2 | R. bifrons_2 | 21.54 | a-k | 18.44 | 25.2 | 1.771 | 1.06 | g-q | 0.8 | 1.38 | 0.141 | 1.159 | d-s | 1.025 | 1.390 | 0.091 |
| 3 | R. bifrons_3 | 22.36 | a-e | 18.47 | 25.84 | 1.956 | 1.165 | a-i | 1 | 1.705 | 0.167 | 1.196 | a-m | 1.058 | 1.404 | 0.087 |
| 4 | R. bifrons_4 | 21.35 | b-n | 16.37 | 26.53 | 2.419 | 1.073 | f-q | 1 | 1.21 | 0.082 | 1.171 | a-r | 1.009 | 1.494 | 0.132 |
| 5 | R. bifrons_5 | 21.01 | c-q | 17 | 24.23 | 1.775 | 0.99 | l-q | 0.8 | 1.2 | 0.109 | 1.213 | a-i | 0.979 | 1.450 | 0.134 |
| 6 | R. caesius_1 | 18.39 | x-z | 14 | 20.82 | 1.498 | 1.096 | e-q | 1 | 1.3 | 0.093 | 1.198 | a-m | 1.006 | 1.596 | 0.136 |
| 7 | R. caesius_2 | 22.92 | ab | 19.08 | 25.05 | 1.471 | 1.15 | a-j | 0.8 | 1.5 | 0.250 | 1.143 | g-t | 1.044 | 1.357 | 0.063 |
| 8 | R. caesius_3 | 22.97 | a | 18.63 | 27.13 | 2.007 | 1.071 | f-q | 1 | 1.3 | 0.105 | 1.124 | i-u | 0.980 | 1.397 | 0.094 |
| 9 | R. caesius_4 | 21.77 | a-j | 19.52 | 23.93 | 1.201 | 1.067 | g-q | 0.8 | 1.44 | 0.152 | 1.177 | a-q | 1.068 | 1.308 | 0.061 |
| 10 | R. caesius_5 | 22.22 | a-f | 17.8 | 24.55 | 1.469 | 1.048 | h-q | 1 | 1.2 | 0.064 | 1.068 | tu | 0.926 | 1.208 | 0.079 |
| 11 | R. constrictus_1 | 21.32 | b-n | 17.2 | 25.57 | 2.31 | 1.068 | g-q | 1 | 1.2 | 0.071 | 1.128 | i-t | 0.931 | 1.433 | 0.121 |
| 12 | R. constrictus_2 | 20.18 | j-w | 15.98 | 24.34 | 1.983 | 1.131 | b-l | 1 | 1.43 | 0.157 | 1.168 | b-r | 1.029 | 1.370 | 0.079 |
| 13 | R. divaricatus_1 | 18.93 | v-y | 14.99 | 22.3 | 1.674 | 1.225 | a-e | 0.8 | 1.53 | 0.214 | 1.187 | a-n | 1.029 | 1.500 | 0.104 |
| 14 | R. divaricatus_2 | 16.81 | z | 12.81 | 21.19 | 1.768 | 1.032 | i-q | 1 | 1.2 | 0.065 | 1.26 | a | 1.055 | 1.527 | 0.144 |
| 15 | R. divaricatus_3 | 19.27 | s-y | 15.45 | 24.57 | 1.9 | 1.109 | c-p | 1 | 1.5 | 0.138 | 1.139 | g-t | 0.955 | 1.456 | 0.130 |
| 16 | R. divaricatus_4 | 19.15 | t-y | 17.08 | 23.03 | 1.546 | 1.185 | a-h | 0.8 | 1.5 | 0.227 | 1.138 | g-t | 0.959 | 1.369 | 0.078 |
| 17 | R. gracilis_1 | 22.08 | a-g | 16.57 | 25.72 | 2.003 | 1.08 | e-q | 0.8 | 1.3 | 0.161 | 1.167 | b-s | 1.000 | 1.462 | 0.127 |
| 18 | R. gracilis_2 | 20.7 | f-t | 18.7 | 22.81 | 1.186 | 1.053 | h-q | 0.8 | 2 | 0.229 | 1.147 | f-t | 1.016 | 1.346 | 0.084 |
| 19 | R. gracilis_3 | 21.43 | a-l | 17.79 | 24.5 | 2.043 | 1.063 | g-q | 0.9 | 1.2 | 0.103 | 1.157 | d-t | 1.003 | 1.366 | 0.089 |
| 20 | R. gracilis_4 | 18.23 | yz | 16.39 | 20.62 | 1.157 | 1.098 | e-q | 0.8 | 1.5 | 0.187 | 1.252 | ab | 1.074 | 1.422 | 0.093 |
| 21 | R. gracilis_5 | 21.3 | c-n | 19.38 | 25.09 | 1.34 | 1.169 | a-i | 0.99 | 1.52 | 0.151 | 1.196 | a-m | 1.038 | 1.372 | 0.084 |
| 22 | R. henrici-egonis_1 | 18.64 | w-y | 16.33 | 22.3 | 1.414 | 1.057 | g-q | 0.9 | 1.23 | 0.086 | 1.219 | a-h | 1.054 | 1.547 | 0.098 |
| 23 | R. henrici-egonis_2 | 22.53 | a-c | 16.36 | 25.71 | 2.088 | 1.135 | a-l | 1 | 1.4 | 0.137 | 1.086 | r-u | 0.985 | 1.241 | 0.074 |
| 24 | R. henrici-egonis_3 | 19.13 | t-y | 14.88 | 22.56 | 1.89 | 1.083 | e-q | 0.99 | 1.8 | 0.150 | 1.179 | a-p | 1.001 | 1.546 | 0.130 |
| 25 | R. henrici-egonis_4 | 18.91 | v-y | 15.21 | 23.09 | 1.832 | 1.202 | a-g | 0.69 | 1.75 | 0.260 | 1.128 | i-t | 0.993 | 1.260 | 0.075 |
| 26 | R. henrici-egonis_5 | 19.12 | t-y | 13.8 | 22.42 | 1.977 | 0.993 | l-q | 0.75 | 1.2 | 0.099 | 1.201 | a-l | 0.956 | 1.650 | 0.161 |
| 27 | R. idaeus_1 | 20.13 | k-w | 13.88 | 23.47 | 2.83 | 0.982 | n-q | 0.8 | 1.21 | 0.118 | 1.121 | k-u | 0.970 | 1.443 | 0.163 |
| 28 | R. idaeus_2 | 22.46 | a-d | 21.08 | 24.1 | 0.86 | 1.181 | a-h | 0.9 | 2 | 0.287 | 1.097 | n-u | 0.953 | 1.182 | 0.058 |
| 29 | R. idaeus_3 | 19.43 | q-y | 14.35 | 22.18 | 1.847 | 1.05 | h-q | 0.7 | 1.3 | 0.168 | 1.183 | a-o | 0.910 | 1.477 | 0.146 |
| 30 | R. idaeus_4 | 19.51 | p-y | 13.87 | 25.1 | 2.786 | 1.141 | a-k | 0.8 | 1.35 | 0.143 | 1.105 | n-u | 0.909 | 1.225 | 0.080 |
| 31 | R. idaeus_5 | 20.77 | e-s | 16.98 | 25.1 | 1.543 | 1.088 | e-q | 1 | 1.36 | 0.116 | 1.088 | q-u | 0.937 | 1.387 | 0.095 |
| 32 | R. nessensis_1 | 19.00 | u-y | 14.48 | 22.35 | 1.833 | 1.135 | a-l | 0.87 | 1.444 | 0.171 | 1.223 | a-g | 1.010 | 1.595 | 0.121 |
| 33 | R. nessensis_2 | 20.11 | k-w | 16.75 | 25.18 | 2.066 | 1.042 | h-q | 0.9 | 1.2 | 0.068 | 1.198 | a-m | 0.968 | 1.451 | 0.117 |
| 34 | R. nessensis_3 | 20.8 | e-s | 17.39 | 27.07 | 2.134 | 1.052 | h-q | 0.78 | 1.23 | 0.124 | 1.235 | a-f | 1.058 | 1.381 | 0.076 |
| 35 | R. nessensis_4 | 20.63 | f-t | 16.36 | 23.66 | 1.854 | 1.046 | h-q | 0.72 | 1.44 | 0.175 | 1.211 | a-k | 1.104 | 1.485 | 0.094 |
| 36 | R. nessensis_5 | 19.79 | m-y | 15.51 | 23.64 | 1.93 | 1.277 | a | 0.9 | 1.5 | 0.210 | 1.241 | a-d | 0.988 | 1.516 | 0.150 |
| 37 | R. opacus_1 | 16.88 | z | 14.16 | 19.63 | 1.697 | 0.966 | pq | 0.8 | 1.2 | 0.124 | 1.223 | a-g | 1.019 | 1.452 | 0.111 |
| 38 | R. opacus_2 | 18.61 | w-y | 15.48 | 21.56 | 1.489 | 1.269 | ab | 0.9 | 1.5 | 0.256 | 1.24 | a-e | 1.030 | 1.421 | 0.118 |
| 39 | R. plicatus_1 | 21.3 | c-n | 19.13 | 24.98 | 1.639 | 1.094 | e-q | 1 | 1.5 | 0.116 | 1.087 | q-u | 0.932 | 1.264 | 0.075 |
| 40 | R. plicatus_2 | 18.97 | u-y | 15.12 | 26 | 2.786 | 1.222 | a-e | 0.78 | 1.75 | 0.313 | 1.125 | i-u | 0.920 | 1.329 | 0.099 |
| 41 | R. plicatus_3 | 21.79 | a-j | 18.29 | 24.5 | 1.737 | 1.124 | b-n | 0.5 | 1.5 | 0.295 | 1.118 | l-u | 0.970 | 1.287 | 0.076 |
| 42 | R. plicatus_4 | 21.25 | c-n | 17.03 | 25.24 | 1.973 | 1.128 | b-m | 0.5 | 1.63 | 0.302 | 1.109 | m-u | 1.002 | 1.483 | 0.087 |
| 43 | R. plicatus_5 | 21.95 | a-h | 17.02 | 25.61 | 1.762 | 0.963 | q | 0.6 | 1.2 | 0.122 | 1.036 | u | 0.908 | 1.165 | 0.064 |
| 44 | R. praecox_1 | 21.67 | a-k | 19.49 | 27.44 | 1.74 | 1.236 | a-d | 0.78 | 1.62 | 0.267 | 1.125 | i-u | 0.984 | 1.296 | 0.085 |
| 45 | R. praecox_2 | 19.53 | o-y | 15.56 | 23.03 | 1.973 | 0.982 | n-q | 0.8 | 1.3 | 0.116 | 1.15 | e-t | 0.971 | 1.458 | 0.131 |
| 46 | R. praecox_3 | 20.44 | h-v | 15.91 | 26.05 | 2.414 | 1.029 | i-q | 0.8 | 1.2 | 0.111 | 1.137 | g-t | 0.994 | 1.412 | 0.099 |
| 47 | R. praecox_4 | 21.9 | a-i | 16.07 | 28.32 | 2.514 | 1.027 | i-q | 0.9 | 1.2 | 0.079 | 1.168 | b-r | 1.001 | 1.518 | 0.132 |
| 48 | R. radula_1 | 22.14 | a-g | 19.19 | 25.07 | 1.708 | 1.202 | a-g | 0.75 | 1.63 | 0.281 | 1.212 | a-j | 1.006 | 1.404 | 0.101 |
| 49 | R. radula_2 | 20.58 | g-u | 16.54 | 25.56 | 1.753 | 0.963 | q | 0.8 | 1.2 | 0.127 | 1.156 | d-t | 0.960 | 1.556 | 0.133 |
| 50 | R. radula_3 | 21.84 | a-i | 18.16 | 26.07 | 1.839 | 1.071 | f-q | 1 | 1.22 | 0.073 | 1.177 | a-q | 1.016 | 1.390 | 0.099 |
| 51 | R. radula_4 | 21.11 | c-p | 16.84 | 24.45 | 2.024 | 1.247 | a-c | 0.9 | 1.9 | 0.265 | 1.155 | d-t | 1.001 | 1.385 | 0.097 |
| 52 | R. radula_5 | 21.82 | a-i | 18.65 | 25.25 | 1.834 | 1.1 | d-q | 1 | 1.4 | 0.124 | 1.154 | d-t | 0.968 | 1.425 | 0.103 |
| 53 | R. saxatilis_1 | 20.31 | i-v | 17.11 | 22.31 | 1.162 | 0.977 | o-q | 0.8 | 1.2 | 0.107 | 1.207 | a-l | 1.049 | 1.385 | 0.085 |
| 54 | R. saxatilis_2 | 20.86 | d-s | 18.41 | 23.34 | 1.297 | 1.153 | a-i | 0.5 | 1.52 | 0.266 | 1.16 | c-s | 0.998 | 1.325 | 0.091 |
| 55 | R. saxatilis_3 | 20.83 | e-s | 16.08 | 23.15 | 1.914 | 1.12 | c-o | 0.72 | 1.82 | 0.228 | 1.093 | o-u | 0.915 | 1.365 | 0.111 |
| 56 | R. saxatilis_4 | 19.77 | n-y | 16.15 | 23.44 | 1.757 | 1.007 | j-q | 0.7 | 1.2 | 0.148 | 1.13 | h-t | 0.952 | 1.456 | 0.127 |
| 57 | R. saxatilis_5 | 19.37 | r-y | 16.54 | 21.7 | 1.427 | 1.047 | h-q | 0.9 | 1.6 | 0.144 | 1.168 | b-r | 0.997 | 1.343 | 0.104 |
| 58 | R. scissus_1 | 20.94 | c-r | 17.99 | 25.58 | 2.15 | 1.033 | i-q | 0.85 | 1.25 | 0.125 | 1.091 | p-u | 0.955 | 1.291 | 0.081 |
| 59 | R. scissus_2 | 21.68 | a-k | 19.03 | 25.13 | 1.49 | 1.096 | e-q | 0.9 | 1.2 | 0.100 | 1.077 | s-u | 0.993 | 1.215 | 0.057 |
| 60 | R. scissus_3 | 21.13 | c-o | 17.15 | 25.9 | 1.929 | 1.068 | g-q | 0.98 | 1.2 | 0.074 | 1.159 | d-s | 0.989 | 1.480 | 0.116 |
| 61 | R. sprengelii_1 | 21.08 | c-p | 17.86 | 24.54 | 1.707 | 1.216 | a-f | 0.9 | 1.5 | 0.223 | 1.181 | a-p | 1.003 | 1.365 | 0.096 |
| 62 | R. sprengelii_2 | 20.72 | f-t | 16.73 | 26.34 | 2.091 | 0.993 | l-q | 0.8 | 1.2 | 0.129 | 1.141 | g-t | 1.009 | 1.322 | 0.094 |
| 63 | R. sprengelii_3 | 19.92 | l-x | 16.3 | 25.34 | 2.227 | 1.149 | a-j | 0.99 | 1.4 | 0.119 | 1.25 | a-c | 1.116 | 1.468 | 0.104 |
| 64 | R. sprengelii_4 | 21 | c-q | 16.84 | 25.42 | 2.344 | 0.997 | k-q | 0.7 | 1.3 | 0.177 | 1.163 | b-s | 0.965 | 1.373 | 0.108 |
| 65 | R. sprengelii_5 | 20.93 | c-r | 19.1 | 23.76 | 1.287 | 0.985 | m-q | 0.8 | 1.11 | 0.095 | 1.127 | i-t | 1.015 | 1.227 | 0.066 |
| 66 | R. sulcatus_1 | 21.65 | a-k | 17.38 | 25.91 | 2.348 | 1.104 | c-q | 0.78 | 1.333 | 0.155 | 1.127 | i-t | 1.002 | 1.304 | 0.082 |
| 67 | R. sulcatus_2 | 18.37 | x-z | 14.57 | 25.47 | 2.524 | 1 | k-q | 0.8 | 1.2 | 0.105 | 1.239 | a-e | 1.000 | 1.614 | 0.164 |
| 68 | R. sulcatus_3 | 21.33 | b-n | 18.46 | 24.1 | 1.474 | 1.044 | h-q | 0.78 | 1.35 | 0.166 | 1.085 | r-u | 0.897 | 1.229 | 0.091 |
| 69 | R. sulcatus_4 | 20.66 | f-t | 16.72 | 24.29 | 2.096 | 1.26 | ab | 0.88 | 1.75 | 0.215 | 1.122 | j-u | 0.966 | 1.324 | 0.094 |
| 70 | R. sulcatus_5 | 21.42 | a-l | 17.79 | 24.77 | 1.855 | 1.098 | e-q | 1 | 1.3 | 0.111 | 1.157 | d-t | 1.031 | 1.429 | 0.091 |
| LSD0.001 | 1.61 | 0.145 | 0.09 | |||||||||||||
| F-ANOVA | 15.41 *** | 6.79 *** | 6.70 *** | |||||||||||||
| Trait | Le/P | d/E | Exp/P | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. | Species | Mean | Homogeneous Groups | Min | Max | s.d. | Mean | Homogeneous Groups | in | Max | s.d. | Mean | Homogeneous Groups | Min | Max | s.d. |
| 1 | R. bifrons_1 | 0.832 | c-k | 0.748 | 0.928 | 0.043 | 0.111 | t-x | 0.044 | 0.164 | 0.031 | 0.042 | n-r | 0.030 | 0.063 | 0.007 |
| 2 | R. bifrons_2 | 0.841 | b-j | 0.762 | 0.922 | 0.042 | 0.137 | h-w | 0.091 | 0.206 | 0.031 | 0.043 | l-r | 0.031 | 0.058 | 0.007 |
| 3 | R. bifrons_3 | 0.860 | a-e | 0.811 | 0.941 | 0.031 | 0.127 | n-x | 0.076 | 0.181 | 0.028 | 0.044 | i-r | 0.033 | 0.066 | 0.009 |
| 4 | R. bifrons_4 | 0.851 | a-i | 0.698 | 0.949 | 0.049 | 0.120 | r-x | 0.068 | 0.221 | 0.037 | 0.044 | k-r | 0.035 | 0.053 | 0.005 |
| 5 | R. bifrons_5 | 0.841 | b-j | 0.731 | 0.933 | 0.043 | 0.121 | r-x | 0.073 | 0.195 | 0.030 | 0.039 | r | 0.032 | 0.051 | 0.005 |
| 6 | R. caesius_1 | 0.834 | b-k | 0.743 | 0.920 | 0.044 | 0.129 | n-x | 0.065 | 0.234 | 0.039 | 0.050 | b-k | 0.040 | 0.067 | 0.006 |
| 7 | R. caesius_2 | 0.856 | a-i | 0.806 | 0.909 | 0.026 | 0.168 | b-h | 0.098 | 0.247 | 0.043 | 0.044 | j-r | 0.031 | 0.067 | 0.010 |
| 8 | R. caesius_3 | 0.845 | b-j | 0.718 | 0.919 | 0.049 | 0.165 | b-k | 0.100 | 0.329 | 0.051 | 0.042 | n-r | 0.034 | 0.053 | 0.005 |
| 9 | R. caesius_4 | 0.822 | i-k | 0.704 | 0.901 | 0.045 | 0.210 | A | 0.148 | 0.275 | 0.039 | 0.042 | n-r | 0.031 | 0.062 | 0.007 |
| 10 | R. caesius_5 | 0.836 | b-k | 0.758 | 0.899 | 0.035 | 0.184 | ab | 0.121 | 0.278 | 0.033 | 0.045 | i-r | 0.037 | 0.058 | 0.005 |
| 11 | R. constrictus_1 | 0.841 | b-j | 0.751 | 0.955 | 0.050 | 0.155 | b-q | 0.084 | 0.265 | 0.050 | 0.045 | h-r | 0.037 | 0.058 | 0.005 |
| 12 | R. constrictus_2 | 0.840 | b-j | 0.701 | 0.907 | 0.043 | 0.136 | i-w | 0.068 | 0.210 | 0.041 | 0.048 | d-n | 0.040 | 0.063 | 0.007 |
| 13 | R. divaricatus_1 | 0.857 | a-h | 0.685 | 0.916 | 0.046 | 0.097 | x | 0.056 | 0.151 | 0.024 | 0.055 | a-d | 0.037 | 0.074 | 0.011 |
| 14 | R. divaricatus_2 | 0.843 | b-j | 0.692 | 0.918 | 0.044 | 0.123 | q-x | 0.077 | 0.229 | 0.034 | 0.049 | c-l | 0.041 | 0.057 | 0.005 |
| 15 | R. divaricatus_3 | 0.834 | b-k | 0.713 | 0.909 | 0.045 | 0.109 | v-x | 0.061 | 0.210 | 0.033 | 0.051 | b-i | 0.043 | 0.077 | 0.007 |
| 16 | R. divaricatus_4 | 0.837 | b-k | 0.789 | 0.902 | 0.029 | 0.107 | wx | 0.055 | 0.179 | 0.032 | 0.055 | a-e | 0.036 | 0.070 | 0.010 |
| 17 | R. gracilis_1 | 0.844 | b-j | 0.764 | 0.909 | 0.041 | 0.140 | f-v | 0.079 | 0.271 | 0.045 | 0.042 | m-r | 0.030 | 0.055 | 0.007 |
| 18 | R. gracilis_2 | 0.858 | a-g | 0.769 | 1.008 | 0.049 | 0.125 | o-x | 0.079 | 0.237 | 0.041 | 0.044 | i-r | 0.033 | 0.077 | 0.009 |
| 19 | R. gracilis_3 | 0.866 | ab | 0.797 | 0.947 | 0.042 | 0.141 | f-v | 0.062 | 0.233 | 0.043 | 0.043 | l-r | 0.033 | 0.053 | 0.005 |
| 20 | R. gracilis_4 | 0.848 | a-i | 0.807 | 0.909 | 0.027 | 0.134 | k-w | 0.077 | 0.267 | 0.039 | 0.049 | c-n | 0.035 | 0.073 | 0.009 |
| 21 | R. gracilis_5 | 0.833 | b-k | 0.742 | 0.905 | 0.045 | 0.128 | n-x | 0.077 | 0.234 | 0.039 | 0.046 | g-r | 0.035 | 0.059 | 0.006 |
| 22 | R. henrici-egonis_1 | 0.831 | d-k | 0.738 | 0.895 | 0.039 | 0.121 | r-x | 0.069 | 0.206 | 0.029 | 0.047 | f-q | 0.037 | 0.058 | 0.006 |
| 23 | R. henrici-egonis_2 | 0.854 | a-i | 0.770 | 0.929 | 0.037 | 0.167 | b-j | 0.102 | 0.245 | 0.045 | 0.047 | g-q | 0.037 | 0.068 | 0.007 |
| 24 | R. henrici-egonis_3 | 0.842 | b-j | 0.771 | 0.991 | 0.048 | 0.113 | s-x | 0.071 | 0.156 | 0.019 | 0.049 | c-n | 0.036 | 0.078 | 0.008 |
| 25 | R. henrici-egonis_4 | 0.844 | b-j | 0.766 | 0.898 | 0.033 | 0.121 | r-x | 0.053 | 0.233 | 0.045 | 0.057 | ab | 0.033 | 0.080 | 0.012 |
| 26 | R. henrici-egonis_5 | 0.848 | a-i | 0.772 | 0.914 | 0.034 | 0.137 | h-w | 0.068 | 0.200 | 0.031 | 0.044 | j-r | 0.032 | 0.055 | 0.006 |
| 27 | R. idaeus_1 | 0.866 | ab | 0.796 | 0.927 | 0.039 | 0.170 | b-g | 0.112 | 0.247 | 0.038 | 0.044 | i-r | 0.035 | 0.058 | 0.005 |
| 28 | R. idaeus_2 | 0.825 | f-k | 0.727 | 0.886 | 0.037 | 0.181 | a-c | 0.139 | 0.228 | 0.023 | 0.048 | e-o | 0.037 | 0.088 | 0.012 |
| 29 | R. idaeus_3 | 0.840 | b-j | 0.711 | 0.912 | 0.038 | 0.158 | b-n | 0.105 | 0.242 | 0.031 | 0.046 | g-r | 0.033 | 0.057 | 0.007 |
| 30 | R. idaeus_4 | 0.860 | a-e | 0.750 | 0.931 | 0.041 | 0.171 | b-f | 0.111 | 0.221 | 0.028 | 0.054 | a-f | 0.037 | 0.069 | 0.009 |
| 31 | R. idaeus_5 | 0.849 | a-i | 0.766 | 0.931 | 0.040 | 0.171 | b-f | 0.127 | 0.226 | 0.027 | 0.049 | c-n | 0.038 | 0.069 | 0.007 |
| 32 | R. nessensis_1 | 0.847 | b-j | 0.744 | 0.930 | 0.046 | 0.134 | k-w | 0.057 | 0.268 | 0.042 | 0.049 | c-m | 0.034 | 0.068 | 0.007 |
| 33 | R. nessensis_2 | 0.844 | b-j | 0.748 | 0.919 | 0.043 | 0.152 | c-r | 0.093 | 0.258 | 0.046 | 0.044 | k-r | 0.033 | 0.056 | 0.005 |
| 34 | R. nessensis_3 | 0.833 | b-k | 0.679 | 0.924 | 0.046 | 0.124 | p-x | 0.058 | 0.226 | 0.039 | 0.041 | o-r | 0.028 | 0.054 | 0.005 |
| 35 | R. nessensis_4 | 0.836 | b-k | 0.759 | 0.917 | 0.035 | 0.144 | e-s | 0.062 | 0.244 | 0.047 | 0.042 | m-r | 0.027 | 0.060 | 0.008 |
| 36 | R. nessensis_5 | 0.836 | b-k | 0.754 | 0.922 | 0.038 | 0.125 | o-x | 0.067 | 0.207 | 0.036 | 0.053 | a-g | 0.034 | 0.070 | 0.010 |
| 37 | R. opacus_1 | 0.864 | a-d | 0.800 | 0.931 | 0.030 | 0.128 | n-x | 0.097 | 0.163 | 0.018 | 0.047 | f-p | 0.034 | 0.061 | 0.007 |
| 38 | R. opacus_2 | 0.803 | k | 0.671 | 0.878 | 0.047 | 0.164 | b-l | 0.101 | 0.277 | 0.045 | 0.055 | a-c | 0.037 | 0.072 | 0.011 |
| 39 | R. plicatus_1 | 0.855 | a-i | 0.774 | 0.897 | 0.030 | 0.123 | q-x | 0.081 | 0.193 | 0.026 | 0.048 | e-p | 0.038 | 0.064 | 0.007 |
| 40 | R. plicatus_2 | 0.867 | ab | 0.783 | 0.946 | 0.032 | 0.150 | c-r | 0.081 | 0.250 | 0.038 | 0.059 | a | 0.036 | 0.099 | 0.019 |
| 41 | R. plicatus_3 | 0.814 | jk | 0.700 | 0.906 | 0.046 | 0.145 | e-r | 0.082 | 0.298 | 0.059 | 0.046 | g-q | 0.021 | 0.071 | 0.013 |
| 42 | R. plicatus_4 | 0.825 | g-k | 0.755 | 0.889 | 0.033 | 0.157 | b-o | 0.106 | 0.255 | 0.039 | 0.048 | d-n | 0.021 | 0.072 | 0.013 |
| 43 | R. plicatus_5 | 0.851 | a-i | 0.771 | 0.918 | 0.039 | 0.167 | b-j | 0.082 | 0.263 | 0.043 | 0.043 | l-r | 0.027 | 0.055 | 0.006 |
| 44 | R. praecox_1 | 0.853 | a-i | 0.784 | 0.938 | 0.041 | 0.128 | n-x | 0.057 | 0.210 | 0.039 | 0.051 | b-j | 0.033 | 0.066 | 0.011 |
| 45 | R. praecox_2 | 0.848 | a-i | 0.778 | 0.935 | 0.039 | 0.173 | b-e | 0.106 | 0.240 | 0.033 | 0.044 | i-r | 0.033 | 0.067 | 0.006 |
| 46 | R. praecox_3 | 0.859 | a-g | 0.751 | 0.905 | 0.034 | 0.132 | l-w | 0.093 | 0.218 | 0.028 | 0.045 | h-r | 0.035 | 0.057 | 0.005 |
| 47 | R. praecox_4 | 0.859 | a-f | 0.753 | 0.918 | 0.035 | 0.136 | h-w | 0.087 | 0.205 | 0.028 | 0.041 | p-r | 0.034 | 0.053 | 0.005 |
| 48 | R. radula_1 | 0.865 | a-c | 0.788 | 0.935 | 0.031 | 0.110 | u-x | 0.058 | 0.170 | 0.028 | 0.045 | h-r | 0.027 | 0.066 | 0.011 |
| 49 | R. radula_2 | 0.850 | a-i | 0.696 | 0.925 | 0.047 | 0.128 | n-x | 0.099 | 0.178 | 0.021 | 0.041 | p-r | 0.032 | 0.052 | 0.006 |
| 50 | R. radula_3 | 0.830 | e-k | 0.699 | 0.912 | 0.048 | 0.143 | e-s | 0.083 | 0.283 | 0.052 | 0.042 | n-r | 0.035 | 0.051 | 0.004 |
| 51 | R. radula_4 | 0.824 | h-k | 0.760 | 0.886 | 0.026 | 0.156 | b-p | 0.102 | 0.233 | 0.033 | 0.052 | b-h | 0.033 | 0.082 | 0.011 |
| 52 | R. radula_5 | 0.848 | a-j | 0.750 | 0.908 | 0.039 | 0.098 | x | 0.061 | 0.223 | 0.034 | 0.044 | j-r | 0.037 | 0.059 | 0.006 |
| 53 | R. saxatilis_1 | 0.881 | a | 0.831 | 0.939 | 0.029 | 0.181 | a-c | 0.119 | 0.290 | 0.044 | 0.040 | qr | 0.031 | 0.048 | 0.005 |
| 54 | R. saxatilis_2 | 0.829 | e-k | 0.704 | 0.998 | 0.057 | 0.164 | b-l | 0.087 | 0.247 | 0.040 | 0.048 | e-o | 0.018 | 0.068 | 0.012 |
| 55 | R. saxatilis_3 | 0.850 | a-i | 0.775 | 0.923 | 0.034 | 0.161 | b-m | 0.105 | 0.250 | 0.042 | 0.050 | c-l | 0.031 | 0.077 | 0.010 |
| 56 | R. saxatilis_4 | 0.847 | b-j | 0.712 | 0.902 | 0.040 | 0.179 | a-d | 0.093 | 0.279 | 0.044 | 0.045 | h-r | 0.035 | 0.058 | 0.005 |
| 57 | R. saxatilis_5 | 0.849 | a-i | 0.754 | 0.914 | 0.037 | 0.168 | b-i | 0.118 | 0.231 | 0.030 | 0.047 | g-q | 0.036 | 0.069 | 0.007 |
| 58 | R. scissus_1 | 0.844 | b-j | 0.721 | 0.960 | 0.045 | 0.125 | o-x | 0.080 | 0.175 | 0.029 | 0.046 | h-r | 0.032 | 0.059 | 0.007 |
| 59 | R. scissus_2 | 0.850 | a-i | 0.753 | 0.941 | 0.039 | 0.164 | b-l | 0.077 | 0.305 | 0.056 | 0.047 | f-p | 0.038 | 0.057 | 0.005 |
| 60 | R. scissus_3 | 0.847 | b-j | 0.774 | 0.894 | 0.032 | 0.169 | b-g | 0.089 | 0.250 | 0.047 | 0.044 | i-r | 0.035 | 0.059 | 0.005 |
| 61 | R. sprengelii_1 | 0.853 | a-i | 0.780 | 0.940 | 0.035 | 0.152 | b-r | 0.093 | 0.250 | 0.036 | 0.049 | c-m | 0.034 | 0.064 | 0.009 |
| 62 | R. sprengelii_2 | 0.840 | b-j | 0.695 | 0.928 | 0.048 | 0.135 | j-w | 0.084 | 0.230 | 0.035 | 0.042 | m-r | 0.031 | 0.052 | 0.005 |
| 63 | R. sprengelii_3 | 0.844 | b-j | 0.765 | 0.917 | 0.038 | 0.136 | i-w | 0.084 | 0.253 | 0.039 | 0.047 | g-q | 0.037 | 0.059 | 0.006 |
| 64 | R. sprengelii_4 | 0.847 | a-j | 0.734 | 0.931 | 0.038 | 0.143 | e-t | 0.100 | 0.220 | 0.029 | 0.041 | o-r | 0.028 | 0.058 | 0.007 |
| 65 | R. sprengelii_5 | 0.867 | ab | 0.806 | 0.915 | 0.030 | 0.138 | g-w | 0.095 | 0.255 | 0.038 | 0.042 | n-r | 0.033 | 0.055 | 0.005 |
| 66 | R. sulcatus_1 | 0.840 | b-j | 0.751 | 0.917 | 0.041 | 0.139 | g-w | 0.085 | 0.227 | 0.035 | 0.046 | h-r | 0.031 | 0.064 | 0.008 |
| 67 | R. sulcatus_2 | 0.859 | a-f | 0.798 | 0.916 | 0.031 | 0.148 | d-r | 0.104 | 0.222 | 0.030 | 0.045 | h-r | 0.038 | 0.056 | 0.005 |
| 68 | R. sulcatus_3 | 0.846 | b-j | 0.731 | 0.917 | 0.043 | 0.131 | m-w | 0.068 | 0.220 | 0.037 | 0.046 | h-r | 0.032 | 0.065 | 0.009 |
| 69 | R. sulcatus_4 | 0.839 | b-j | 0.769 | 0.905 | 0.037 | 0.141 | f-u | 0.077 | 0.242 | 0.043 | 0.055 | a-d | 0.038 | 0.092 | 0.012 |
| 70 | R. sulcatus_5 | 0.867 | ab | 0.786 | 0.935 | 0.034 | 0.128 | n-x | 0.072 | 0.210 | 0.036 | 0.045 | h-r | 0.036 | 0.056 | 0.006 |
| LSD0.001 | 0.034 | 0.032 | 0.007 | |||||||||||||
| F-ANOVA | 3.51 *** | 10.96 *** | 8.64 *** | |||||||||||||
| Striate Exine Ornamentation Type or Subtype | Species |
|---|---|
| IIA | R. divaricatus, R. gracilis, R. henrici-egonis, R. plicatus, R. praecox, R. radula, R. scissus, R. sprengelii |
| IIB | R. caesius, R. idaeus |
| IIIA | R. bifrons, R. nessensis, R. opacus, R. saxatilis |
| IIIB | R. constrictus, R. sulcatus |
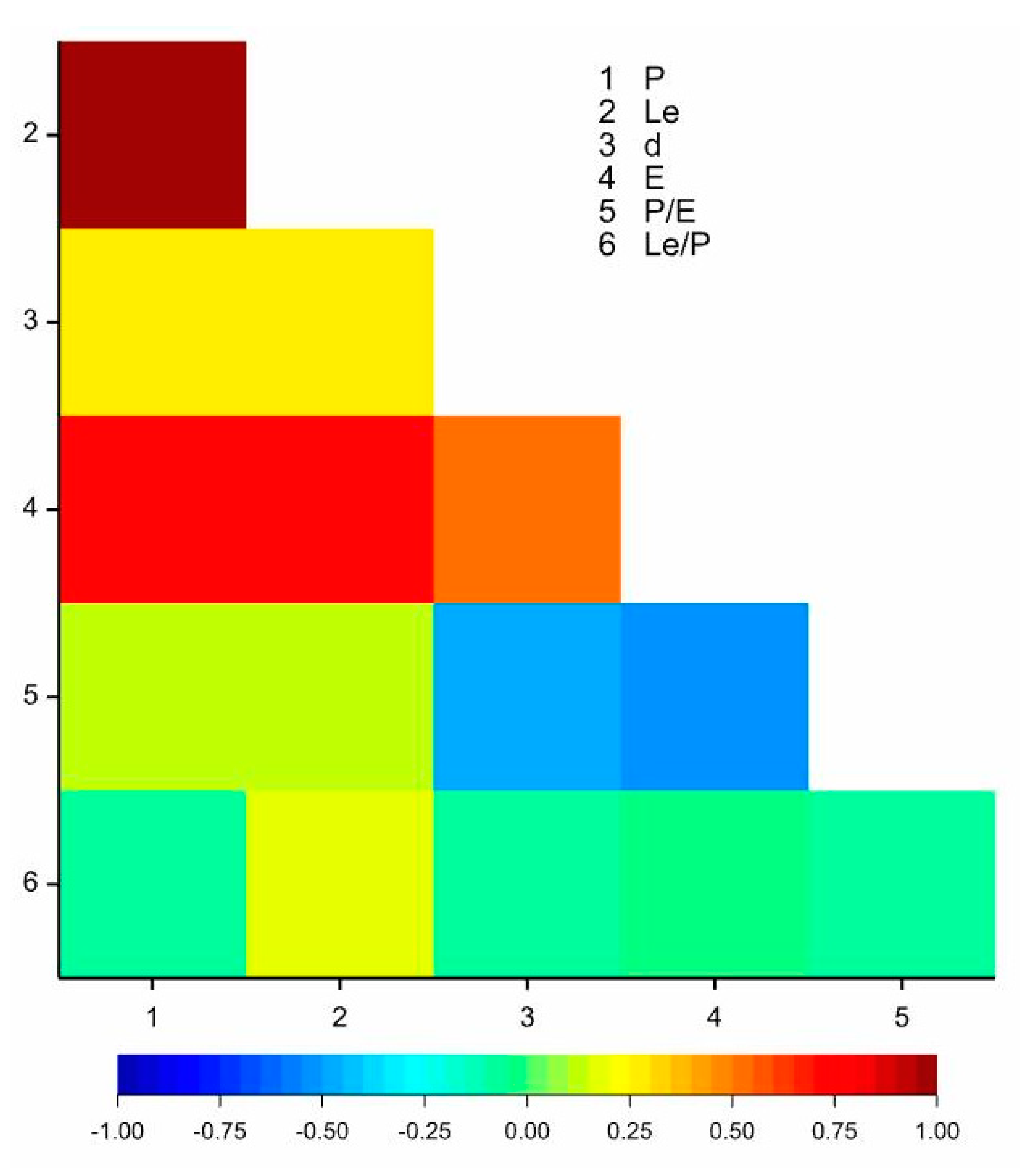
| Trait | First Canonical Variate | Second Canonical Variate |
|---|---|---|
| P | −0.951 *** | −0.249 * |
| Le | −0.914 *** | −0.206 |
| d | −0.485 *** | 0.804 *** |
| E | −0.918 *** | 0.189 |
| Exp | −0.062 | −0.384 ** |
| P/E | 0.141 | −0.625 *** |
| Le/P | 0.086 | 0.162 |
| d/E | −0.201 | 0.861 *** |
| Exp/P | 0.585 *** | −0.161 |
| Percentage of explained multivariate variability | 33.98 | 24.42 |
| Species | Ranking | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 (the Largest) | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 (the Smallest) | |
| R. bifrons | Exp *** | Le *** | P ** | d ** | d/E * | Exp/P * | Le/P | E | P/E |
| R. caesius | E *** | P *** | d *** | Le *** | d/E *** | P/E *** | Exp/P *** | Le/P * | Exp |
| R. constrictus | d * | Exp/P * | E * | Exp * | d/E | P/E | P | Le | Le/P |
| R. divaricatus | E *** | Exp *** | P/E *** | Le ** | P * | d/E * | Exp/P * | Le/P | d |
| R. gracilis | E *** | P *** | Le *** | P/E *** | Exp/P * | d * | Le/P * | Exp | d/E |
| R. henrici-egonis | d *** | E *** | Exp/P *** | d/E *** | P *** | Le *** | Exp *** | P/E *** | Le/P |
| R. idaeus | P *** | E *** | d *** | Exp *** | Exp/P *** | Le/P *** | Le ** | P/E * | d/E |
| R. nessensis | Exp/P *** | Exp *** | P *** | E ** | Le * | d * | d/E | P/E | Le/P |
| R. opacus | P *** | Le/P *** | Exp *** | d *** | E *** | d/E *** | Exp/P ** | Le | P/E |
| R. plicatus | P *** | Le/P *** | E *** | Exp/P *** | P/E *** | d *** | d/E ** | Exp ** | Le * |
| R. praecox | P *** | Le *** | Exp *** | d/E *** | Exp/P *** | E *** | d ** | P/E | Le/P |
| R. radula | d/E *** | P *** | Le *** | Exp *** | d *** | Exp/P *** | Le/P *** | E ** | P/E |
| R. saxatilis | Le *** | P *** | Le/P *** | Exp/P *** | E *** | P/E *** | Exp *** | d/E | d |
| R. scissus | d *** | d/E *** | P/E *** | P *** | Le ** | Exp | Exp/P | E | Le/P |
| R. sprengelii | Exp *** | Exp/P *** | P/E *** | P ** | Le * | Le/P | d | E | d/E |
| R. sulcatus | E *** | Exp *** | Exp/P *** | P/E *** | P *** | Le *** | Le/P * | d/E | d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lechowicz, K.; Bocianowski, J.; Wrońska-Pilarek, D. Pollen Morphological Inter- and Intraspecific Variability in Selected Species of Rubus L. (Rosaceae). Forests 2022, 13, 1946. https://doi.org/10.3390/f13111946
Lechowicz K, Bocianowski J, Wrońska-Pilarek D. Pollen Morphological Inter- and Intraspecific Variability in Selected Species of Rubus L. (Rosaceae). Forests. 2022; 13(11):1946. https://doi.org/10.3390/f13111946
Chicago/Turabian StyleLechowicz, Kacper, Jan Bocianowski, and Dorota Wrońska-Pilarek. 2022. "Pollen Morphological Inter- and Intraspecific Variability in Selected Species of Rubus L. (Rosaceae)" Forests 13, no. 11: 1946. https://doi.org/10.3390/f13111946