Soil Organic Carbon Distribution, Enzyme Activities, and the Temperature Sensitivity of a Tropical Rainforest in Wuzhishan, Hainan Island

Abstract
:1. Introduction
2. Materials and Methods
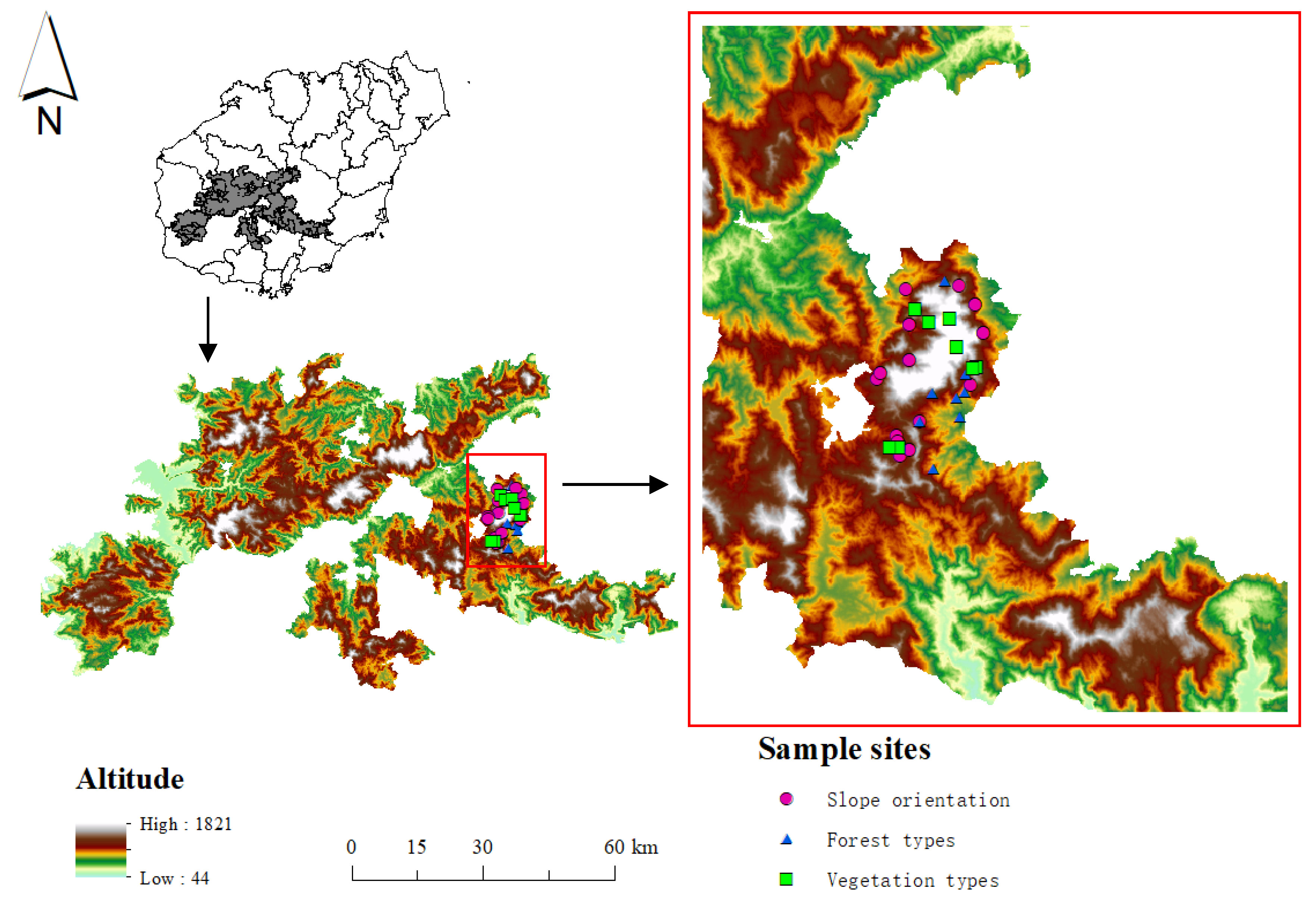
2.1. Study Area
2.2. Soil Sample Collection
2.3. Sample Analysis
2.4. Data Analysis
3. Results
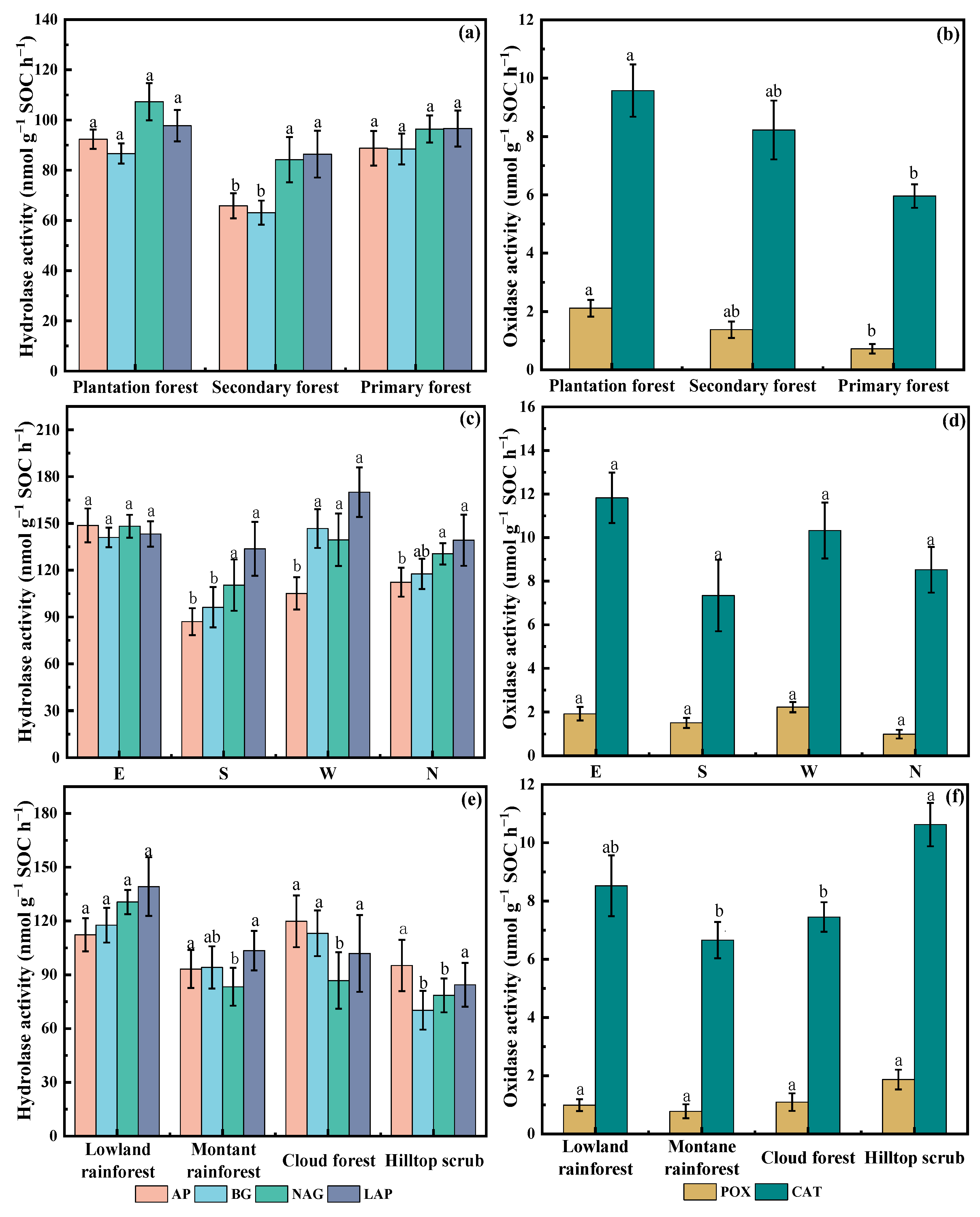
3.1. SOC of Different Forest Types, Slope Orientations, and Vegetation Types
3.2. Characteristics of Soil Enzyme Activities in Different Forest Types, Slope Orientations, and Vegetation Types
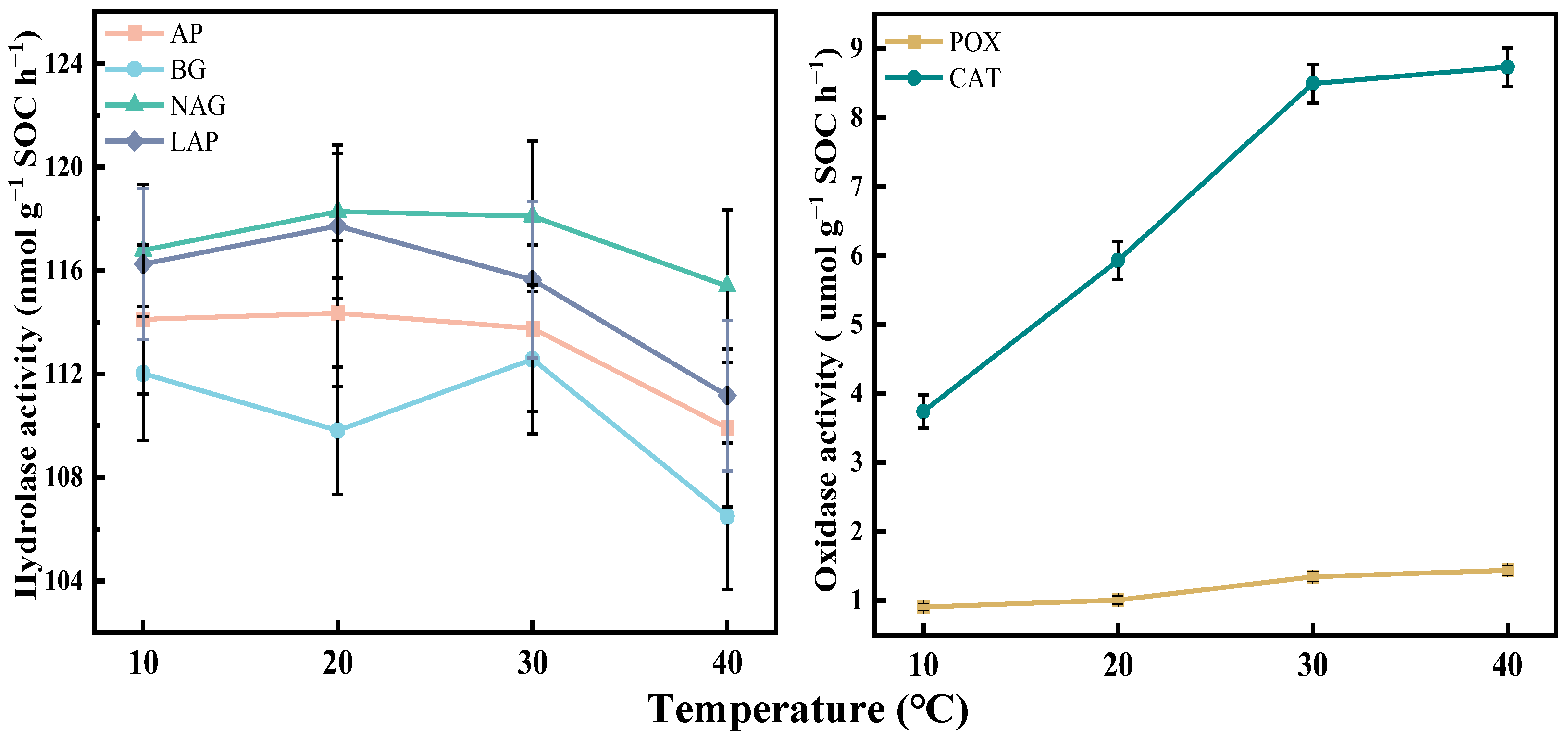
3.3. Temperature-Sensitive Characteristics of Soil Enzyme Activity
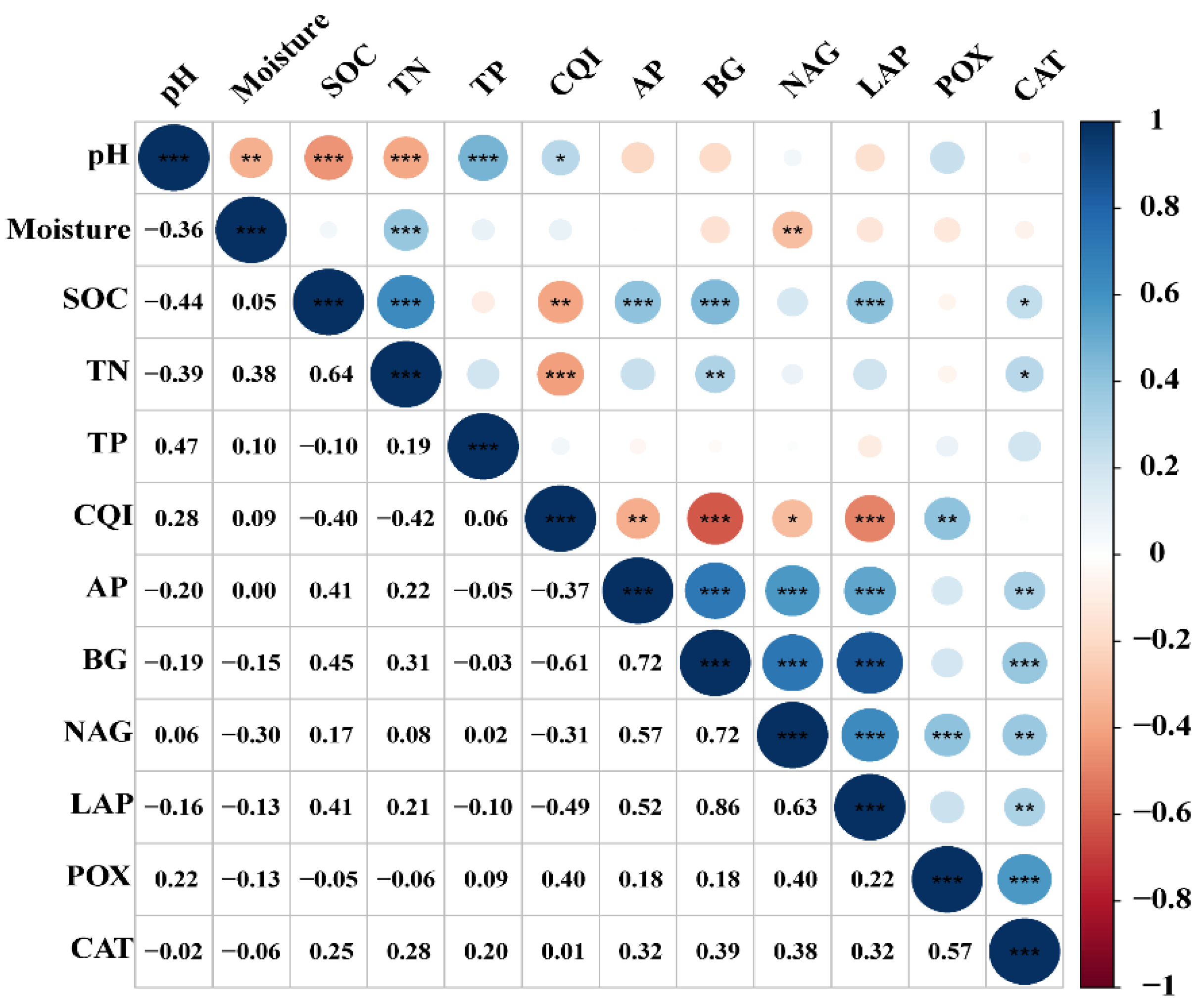
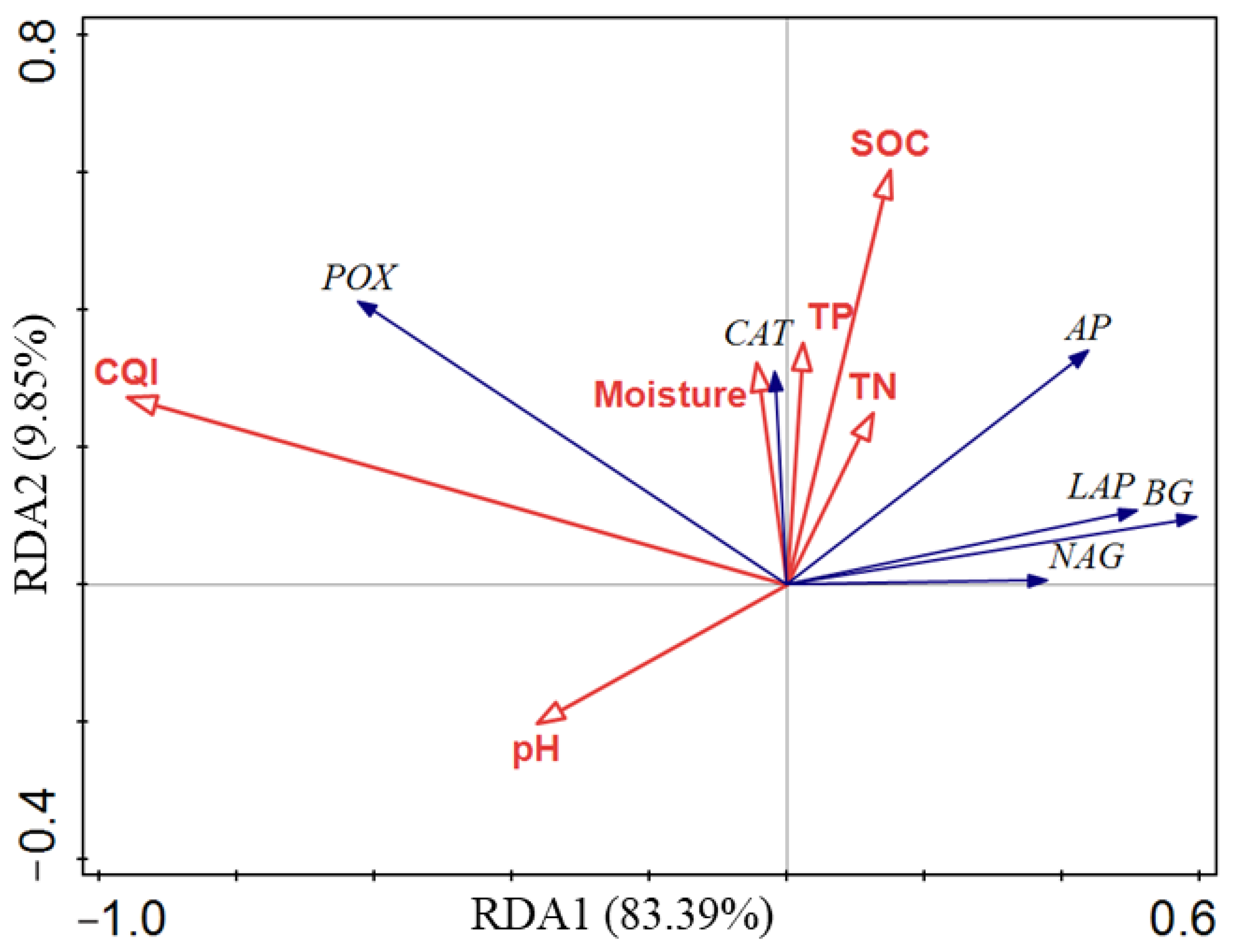
3.4. Relationship between Soil Enzyme Activity, SOC, and Other Soil Properties
4. Discussion
4.1. SOC and Enzyme Activity Distribution in Tropical Forests
4.2. Responses of Soil Enzyme Activities to Temperature Increase in the Tropical Rainforest
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Adhikari, K.; Owens, P.R.; Libohova, Z.; Miller, D.M.; Wills, S.A.; Nemecek, J. Assessing soil organic carbon stock of Wisconsin, USA and its fate under future land use and climate change. Sci. Total Environ. 2019, 667, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Tian, H.; Gu, X.; Li, N.; Zhao, X.; Lei, M.; Alharbi, H.; Megharaj, M.; He, W.; Kuzyakov, Y. Catalytic efficiency of soil enzymes explains temperature sensitivity: Insights from physiological theory. Sci. Total Environ. 2022, 822, 153365. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Li, N.; Leng, X.; Qiao, Y.; Cheng, X.; An, S. The impact of sea embankment reclamation on soil organic carbon and nitrogen pools in invasive Spartina alterniflora and native Suaeda salsa salt marshes in eastern China. Ecol. Eng. 2016, 97, 582–592. [Google Scholar] [CrossRef]
- Zhao, B.; Cao, J.; Geng, Y.; Zhao, X.; von Gadow, K. Inconsistent responses of soil respiration and its components to thinning intensity in a Pinus tabuliformis plantation in northern China. Agric. For. Meteorol. 2019, 265, 370–380. [Google Scholar] [CrossRef]
- Zhang, D.; Cai, X.; Diao, L.; Wang, Y.; Wang, J.; An, S.; Cheng, X.; Yang, W. Changes in soil organic carbon and nitrogen pool sizes, dynamics, and biochemical stability during ∼160 years natural vegetation restoration on the Loess Plateau, China. CATENA 2022, 211, 106014. [Google Scholar] [CrossRef]
- Wang, C.; Houlton, B.Z.; Liu, D.; Hou, J.; Cheng, W.; Bai, E. Stable isotopic constraints on global soil organic carbon turnover. Biogeosciences 2018, 15, 987–995. [Google Scholar] [CrossRef] [Green Version]
- Zeh, L.; Limpens, J.; Erhagen, B.; Bragazza, L.; Kalbitz, K. Plant functional types and temperature control carbon input via roots in peatland soils. Plant Soil 2019, 438, 19–38. [Google Scholar] [CrossRef]
- Conant, R.T.; Ryan, M.G.; Ågren, G.I.; Birge, H.E.; Davidson, E.A.; Eliasson, P.E.; Evans, S.E.; Frey, S.D.; Giardina, C.P.; Hopkins, F.M.; et al. Temperature and soil organic matter decomposition rates—Synthesis of current knowledge and a way forward. Glob. Chang. Biol. 2011, 17, 3392–3404. [Google Scholar] [CrossRef]
- Varik, M.; Aosaar, J.; Ostonen, I.; Lõhmus, K.; Uri, V. Carbon and nitrogen accumulation in belowground tree biomass in a chronosequence of silver birch stands. For. Ecol. Manag. 2013, 302, 62–70. [Google Scholar] [CrossRef]
- Ma, J.; Bu, R.; Liu, M.; Chang, Y.; Qin, Q.; Hu, Y. Ecosystem carbon storage distribution between plant and soil in different forest types in Northeastern China. Ecol. Eng. 2015, 81, 353–362. [Google Scholar] [CrossRef]
- Chaturvedi, R.K.; Raghubanshi, A.S. Aboveground biomass estimation of small diameter woody species of tropical dry forest. New For. 2013, 44, 509–519. [Google Scholar] [CrossRef]
- Su, F.; Xu, S.; Sayer, E.J.; Chen, W.; Du, Y.; Lu, X. Distinct storage mechanisms of soil organic carbon in coniferous forest and evergreen broadleaf forest in tropical China. J. Environ. Manag. 2021, 295, 113142. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Ding, Y.; Sayer, E.J.; Li, Q.; Zou, B.; Mo, Q.; Li, Y.; Lu, X.; Tang, J.; Zhu, W.; et al. Tropical forest restoration: Fast resilience of plant biomass contrasts with slow recovery of stable soil C stocks. Funct. Ecol. 2017, 31, 2344–2355. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Jiang, Y.; Yang, Q.; Yang, H.; Li, Y.; Li, Z.; Mao, W.; Luo, Y.; Wang, X.; Tan, Z. Spatial distribution and stability mechanisms of soil organic carbon in a tropical montane rainforest. Ecol. Indic. 2021, 129, 107965. [Google Scholar] [CrossRef]
- Martin, D.; Lal, T.; Sachdev, C.B.; Sharma, J.P. Soil organic carbon storage changes with climate change, landform and land use conditions in Garhwal hills of the Indian Himalayan mountains. Agric. Ecosyst. Environ. 2010, 138, 64–73. [Google Scholar] [CrossRef]
- Meng, C.; Tian, D.; Zeng, H.; Li, Z.; Chen, H.Y.H.; Niu, S. Global meta-analysis on the responses of soil extracellular enzyme activities to warming. Sci. Total Environ. 2020, 705, 135992. [Google Scholar] [CrossRef]
- Deng, Y.; Chen, X.; Yao, X.; Dong, L.; Zhang, H.; Zeng, H.; Wang, W. No thermal adaptation in soil extracellular enzymes across a temperate grassland region. Soil Biol. Biochem. 2022, 165, 108540. [Google Scholar] [CrossRef]
- He, Q.; Wu, Y.; Bing, H.; Zhou, J.; Wang, J. Vegetation type rather than climate modulates the variation in soil enzyme activities and stoichiometry in subalpine forests in the eastern Tibetan Plateau. Geoderma 2020, 374, 114424. [Google Scholar] [CrossRef]
- Boeddinghaus, R.S.; Nunan, N.; Berner, D.; Marhan, S.; Kandeler, E. Do general spatial relationships for microbial biomass and soil enzyme activities exist in temperate grassland soils? Soil Biol. Biochem. 2015, 88, 430–440. [Google Scholar] [CrossRef]
- Brockett, B.F.T.; Prescott, C.E.; Grayston, S.J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Brouns, K.; Eikelboom, T.; Jansen, P.C.; Janssen, R.; Kwakernaak, C.; van den Akker, J.J.H.; Verhoeven, J.T.A. Spatial Analysis of Soil Subsidence in Peat Meadow Areas in Friesland in Relation to Land and Water Management, Climate Change, and Adaptation. Environ. Manag. 2015, 55, 360–372. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.Y.; Liu, W.X.; Tian, R.; Yang, S.; Wang, J.; Huang, J.S.; Yang, Y.S.; Liu, L.L. Effects of altitude and aspect on soil extracellular enzyme activities in Tanggula Mountain. Acta Ecol. Sin. 2021, 41, 7659–7668. [Google Scholar]
- Zuo, Y.; Zhang, H.; Li, J.; Yao, X.; Chen, X.; Zeng, H.; Wang, W. The effect of soil depth on temperature sensitivity of extracellular enzyme activity decreased with elevation: Evidence from mountain grassland belts. Sci. Total Environ. 2021, 777, 146136. [Google Scholar] [CrossRef] [PubMed]
- Jyothi, A.; Rao, P.C.; Padmaja, G.; Madhavi, M. Effect of Temperature on Soil Enzyme Acid Phosphatase. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 15. [Google Scholar]
- McDaniel, M.D.; Kaye, J.P.; Kaye, M.W. Increased temperature and precipitation had limited effects on soil extracellular enzyme activities in a post-harvest forest. Soil Biol. Biochem. 2013, 56, 90–98. [Google Scholar] [CrossRef]
- Jing, X.; Wang, Y.; Chung, H.; Mi, Z.; Wang, S.; Zeng, H.; He, J.-S. No temperature acclimation of soil extracellular enzymes to experimental warming in an alpine grassland ecosystem on the Tibetan Plateau. Biogeochemistry 2014, 117, 39–54. [Google Scholar] [CrossRef]
- German, D.P.; Marcelo, K.R.B.; Stone, M.M.; Allison, S.D. The Michaelis-Menten kinetics of soil extracellular enzymes in response to temperature: A cross-latitudinal study. Glob. Chang. Biol. 2012, 18, 1468–1479. [Google Scholar] [CrossRef]
- Liu, Q.; Xu, X.; Wang, H.; Blagodatskaya, E.; Kuzyakov, Y. Dominant extracellular enzymes in priming of SOM decomposition depend on temperature. Geoderma 2019, 343, 187–195. [Google Scholar] [CrossRef]
- Ali, R.S.; Ingwersen, J.; Demyan, M.S.; Funkuin, Y.N.; Wizemann, H.-D.; Kandeler, E.; Poll, C. Modelling in situ activities of enzymes as a tool to explain seasonal variation of soil respiration from agro-ecosystems. Soil Biol. Biochem. 2015, 81, 291–303. [Google Scholar] [CrossRef]
- Yang, X.; Lin, Y.; Liang, S. Forest vegetation in Wuzhishan, Hainan Island Ⅰ. Types of forest vegetation in the Wuzhishan Mountains. J. Hainan Univ. (Nat. Sci.Ed.) 1994, 12, 220–236. [Google Scholar]
- Yin, W.; Li, J.; Huang, L.; Liu, W.; Li, D. Research on physicochemical properties of soil in thatch land of Wuzhishan National Nature Reserve. For. Surv. Plan. 2018, 43, 41–46+61. [Google Scholar]
- Zhuo, S.; Hu, N.; Chen, K.; Li, J.; Yin, W.; Huang, L.; Long, W. Species distribution patterns of tree communities on different slopes and elevations in Wuzhishan Nature Reserve. J. Trop. Biol. 2017, 8, 436–443. [Google Scholar]
- Li, J.; Liang, Y.; Liu, W.; Yang, Q.; Xu, W.; Tang, S.; Wang, J. Effect of organic fertilizer instead of chemical nitrogen fertilizer on the growth and soil environment of rubber seedlings. J. Appl. Ecol. 2022, 33, 431–438. [Google Scholar]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zakb, D.R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Wallenstein, M.D.; McMahon, S.K.; Schimel, J.P. Seasonal variation in enzyme activities and temperature sensitivities in Arctic tundra soils. Glob. Chang. Biol. 2009, 15, 1631–1639. [Google Scholar] [CrossRef]
- Hill, B.H.; Elonen, C.M.; Jicha, T.M.; Kolka, R.K.; Lehto, L.L.P.; Sebestyen, S.D.; Seifert-Monson, L.R. Ecoenzymatic stoichiometry and microbial processing of organic matter in northern bogs and fens reveals a common P-limitation between peatland types. Biogeochemistry 2014, 120, 203–224. [Google Scholar] [CrossRef]
- Peng, X.; Wang, W. Stoichiometry of soil extracellular enzyme activity along a climatic transect in temperate grasslands of northern China. Soil Biol. Biochem. 2016, 98, 74–84. [Google Scholar] [CrossRef]
- Moorhead, D.L.; Hill, B.H.; Weintraub, M.N.; Sinsabaugh, R.L. Vector analysis of ecoenzyme activities reveal constraints on coupled C, N and P dynamics. Soil Biol. Biochem. 2016, 93, 1–7. [Google Scholar] [CrossRef]
- Yu, H.; Gao, R.; Yang, W.; Yang, L.; Li, S.; Lin, Y.; Wang, D.; Li, K. Leaf, root and soil carbon, nitrogen and phosphorus contents and relationships of dominant herbaceous plants in a dry thermal valley. J. Appl. Environ. Biol. 2022, 28, 727–735. [Google Scholar]
- Lin, W.Q.; Zou, X.J.; She, H.J.; Xue, L. Vertical Distribution of Soil Organic Carbon and Nutrient Element Storages of An Elaeocarpus sylvestris Plantation. J. Northeast. For. Univ. 2019, 47, 55–59. [Google Scholar]
- Ding, Y.; Mao, Z.; Zhang, L.; Ding, L. Comparative study of soil organic carbon pools in primary broadleaf red pine and maple birch secondary forests in the Xiaoxingan Mountains. Plant Res. 2015, 35, 604–611. [Google Scholar]
- Zhou, L.; Liu, S.; Shen, H.; Zhao, M.; Xu, L.; Xing, A.; Fang, J. Soil extracellular enzyme activity and stoichiometry in China’s forests. Funct. Ecol. 2020, 34, 1461–1471. [Google Scholar] [CrossRef]
- Xu, G.; Long, Z.; Ren, P.; Ren, C.; Cao, Y.; Huang, Y.; Hu, S. Differential responses of soil hydrolytic and oxidative enzyme activities to the natural forest conversion. Sci. Total Environ. 2020, 716, 136414. [Google Scholar] [CrossRef] [PubMed]
- Liu, W. Carbon, Nitrogen and Phosphorus Content and Ecological Chemometrics of Jiaozhou Bay Wetland Ecosystem. Master’s Thesis, Qingdao University, Qingdao, China, 2014. [Google Scholar]
- Sinsabaugh, R.L.; Shah, J.J.F. Ecoenzymatic Stoichiometry and Ecological Theory. Annu. Rev.Ecol. Evol. Syst. 2012, 43, 313. [Google Scholar] [CrossRef] [Green Version]
- Weintraub, S.R.; Wieder, W.R.; Cleveland, C.C.; Townsend, A.R. Organic matter inputs shift soil enzyme activity and allocation patterns in a wet tropical forest. Biogeochemistry 2013, 114, 313–326. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, J.; Vogt, R.D.; Mulder, J.; Wang, Y.; Qian, C.; Wang, J.; Zhang, X. Soil acidification as an additional driver to organic carbon accumulation in major Chinese croplands. Geoderma 2020, 366, 114234. [Google Scholar] [CrossRef]
- Huang, X.; Cui, C.; Hou, E.; Li, F.; Liu, W.; Jiang, L.; Luo, Y.; Xu, X. Acidification of soil due to forestation at the global scale. For. Ecol. Manag. 2022, 505, 119951. [Google Scholar] [CrossRef]
- Le, V.S.; Herrmann, L.; Hudek, L.; Nguyen, T.B.; Bräu, L.; Lesueur, D. How application of agricultural waste can enhance soil health in soils acidified by tea cultivation: A review. Environ. Chem. Lett. 2022, 20, 813–839. [Google Scholar] [CrossRef]
- Ghiloufi, W.; Seo, J.; Kim, J.; Chaieb, M.; Kang, H. Effects of Biological Soil Crusts on Enzyme Activities and Microbial Community in Soils of an Arid Ecosystem. Microb. Ecol. 2019, 77, 201–216. [Google Scholar] [CrossRef]
- Wu, S.G.; Wang, W.P.; Li, B.; Liang, Y.L.; Liu, Y.Z. Altitudinal gradients of soil organic carbon in a central subtropical forest. Soil J. 2020, 57, 1539–1547. [Google Scholar]
- Angel, R.; Soares, M.I.M.; Ungar, E.D.; Gillor, O. Biogeography of soil archaea and bacteria along a steep precipitation gradient. ISME J. 2010, 4, 553–563. [Google Scholar] [CrossRef]
- Huang, Y.-M.; Liu, D.; An, S.-S. Effects of slope aspect on soil nitrogen and microbial properties in the Chinese Loess region. CATENA 2015, 125, 135–145. [Google Scholar] [CrossRef]
- Shen, J.; Li, X.; Yuan, Y.; Wen, J. Response of soil enzymes to different elevations and slope orientations in Napanhe Nature Reserve. Soil Water Conserv. Res. 2018, 25, 111–119+125. [Google Scholar]
- Zhou, P.; Wei, L.; Wei, X.; Zhu, Z.; Yuan, H.; Li, Q.; Wu, J. Characterization of β-1,4-glucosidase activity in paddy soils in response to temperature changes. Environ. Sci. Res. 2018, 3l, 1282–1288. [Google Scholar]
- Nottingham, A.T.; Turner, B.L.; Whitaker, J.; Ostle, N.; Bardgett, R.D.; McNamara, N.P.; Salinas, N.; Meir, P. Temperature sensitivity of soil enzymes along an elevation gradient in the Peruvian Andes. Biogeochemistry 2016, 127, 217–230. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Liu, G.; Li, P.; Xue, S. Dynamics of soil specific enzyme activities and temperature sensitivities during grassland succession after farmland abandonment. CATENA 2021, 199, 105081. [Google Scholar] [CrossRef]
- Razavia, B.S.; Blagodatskayab, E.; Kuzyakova, Y. Temperature selects for static soil enzyme systems to maintain high catalytic efficiency. Soil Biol. Biochem. 2016, 97, 15–22. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Allison, S.D.; Romero-Olivares, A.L.; Lu, Y.; Taylor, J.W.; Treseder, K.K. Temperature sensitivities of extracellular enzyme Vmax and Km across thermal environments. Glob. Chang. Biol. 2018, 24, 2884–2897. [Google Scholar] [CrossRef] [Green Version]
- Razavi, B.; Blagodatskaya, E.; Kuzyakov, Y. Nonlinear temperature sensitivity of enzyme kinetics explains canceling effect-A case study on loamy haplic Luvisol. Front. Microbiol. 2015, 6, 1126. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Elsgaard, L.; van Groenigen, K.J.; Olesen, J.E.; Liang, Z.; Jiang, Y.; Lærke, P.E.; Zhang, Y.; Luo, Y.; Hungate, B.A.; et al. Soil carbon loss with warming: New evidence from carbon-degrading enzymes. Glob. Chang. Biol. 2020, 26, 1944–1952. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | pH | Moisture % | SOC g/kg | TN g/kg | TP g/kg | SAP mg/kg | Vector L | Vector A | CQI | |
|---|---|---|---|---|---|---|---|---|---|---|
| Forest type | Plantation forest | 6.20 ± 0.03 a | 7.54 ± 0.41 c | 28.96 ± 1.56 ab | 2.02 ± 0.11 a | 0.15 ± 0.014 b | 1.85 ± 0.014 a | 1.30 ± 0.010 a | 40.39 ± 0.42 a | 0.63 ± 0.004 a |
| Secondary forest | 5.99 ± 0.05 ab | 15.15 ± 0.39 a | 24.82 ± 1.62 b | 2.08 ± 0.08 a | 0.26 ± 0.075 a | 1.66 ± 0.014 b | 1.29 ± 0.026 a | 39.62 ± 0.94 a | 0.63 ± 0.009 a | |
| Primary forest | 5.83 ± 0.09 b | 11.17 ± 0.37 b | 32.25 ± 1.84 a | 2.09 ± 0.15 a | 0.11 ± 0.028 b | 1.82 ± 0.052 a | 1.31 ± 0.011 a | 40.34 ± 0.36 a | 0.58 ± 0.011 b | |
| Vegetation type | Lowland rainforest | 5.86 ± 0.06 a | 14.69 ± 0.30 b | 36.91 ± 1.72 b | 2.46 ± 0.17 b | 0.31 ± 0.058 a | 1.59 ± 0.014 b | 1.32 ± 0.029 a | 40.12 ± 0.81 a | 0.59 ± 0.011 a |
| Montane rainforest | 5.35 ± 0.08 b | 15.64 ± 1.67 b | 33.49 ± 0.38 b | 2.10 ± 0.26 b | 0.15 ± 0.010 b | 1.98 ± 0.002 a | 1.34 ± 0.016 a | 41.90 ± 0.68 a | 0.59 ± 0.017 a | |
| Cloud forest | 5.50 ± 0.10 b | 30.17 ± 2.17 a | 87.72 ± 0.82 a | 5.92 ± 0.06 a | 0.18 ± 0.047 b | 1.70 ± 0.097 b | 1.34 ± 0.016 a | 42.25 ± 1.22 a | 0.59 ± 0.013 a | |
| Hilltop scrub | 5.45 ± 0.07 b | 34.89 ± 2.52 a | 86.07 ± 2.19 a | 4.97 ± 0.51 a | 0.14 ± 0.019 b | 1.65 ± 0.009 b | 1.25 ± 0.024 a | 41.59 ± 0.79 a | 0.63 ± 0.009 a | |
| Slope orientation | E | 5.57 ± 0.71 c | 14.16 ± 1.3 b | 44.59 ± 5.05 ab | 2.31 ± 0.13 a | 0.02 ± 0.002 c | 1.82 ± 0.026 a | 1.32 ± 0.011 a | 41.34 ± 0.26 a | 0.60 ± 0.007 a |
| S | 5.77 ± 0.05 c | 14.38 ± 0.49 b | 27.08 ± 0.29 b | 2.04 ± 0.08 a | 0.06 ± 0.013 bc | 1.66 ± 0.011 ab | 1.34 ± 0.023 a | 40.29 ± 1.10 a | 0.62 ± 0.010 a | |
| W | 5.79 ± 0.12 c | 14.09 ± 0.42 b | 35.89 ± 0.68 ab | 2.51 ± 0.11 a | 0.29 ± 0.131 ab | 1.55 ± 0.006 b | 1.38 ± 0.022 a | 39.02 ± 0.21 a | 0.61 ± 0.003 a | |
| N | 5.87 ± 0.06 bc | 14.69 ± 0.30 b | 36.93 ± 1.72 ab | 2.46 ± 0.17 a | 0.31 ± 0.058 a | 1.59 ± 0.014 b | 1.32 ± 0.029 a | 40.12 ± 0.81 a | 0.59 ± 0.011 a | |
| ES | 5.21 ± 0.03 d | 18.58 ± 1.77 a | 56.30 ± 14.99 a | 2.60 ± 0.44 a | 0.20 ± 0.068 abc | 1.72 ± 0.015 ab | 1.30 ± 0.011 a | 40.99 ± 0.51 a | 0.60 ± 0.015 a | |
| EN | 6.18 ± 0.17 b | 13.39 ± 0.63 b | 31.96 ± 3.60 b | 1.92 ± 0.17 a | 0.13 ± 0.013 abc | 1.84 ± 0.085 a | 1.34 ± 0.004 a | 42.08 ± 0.37 a | 0.59 ± 0.011 a | |
| WS | 5.68 ± 0.06 c | 13.23 ± 0.68 b | 31.03 ± 2.40 b | 1.95 ± 0.26 a | 0.18 ± 0.025 abc | 1.72 ± 0.048 ab | 1.36 ± 0.031 a | 42.13 ± 1.09 a | 0.60 ± 0.009 a | |
| WN | 6.58 ± 0.05 a | 16.32 ± 1.01 ab | 29.71 ± 4.61 b | 2.27 ± 0.35 a | 0.37 ± 0.058 a | 1.80 ± 0.078 a | 1.37 ± 0.028 a | 40.87 ± 0.94 a | 0.60 ± 0.007 a |
| Type | Temperature (°C) | AP (Q10) | BG (Q10) | NAG (Q10) | LAP (Q10) | POX (Q10) | CAT (Q10) | |
|---|---|---|---|---|---|---|---|---|
| Forest type | Plantation forest | 10–20 | 1.02 ± 0.05 a | 0.93 ± 0.06 a | 1.02 ± 0.06 a | 0.98 ± 0.07 a | 0.80 ± 0.12 a | 1.71 ± 0.08 a |
| 20–30 | 1.16 ± 0.08 a | 1.07 ± 0.07 a | 1.07 ± 0.04 a | 1.06 ± 0.06 a | 1.11 ± 0.06 ab | 1.92 ± 0.05 a | ||
| 30–40 | 0.87 ± 0.06 a | 0.87 ± 0.03 a | 0.93 ± 0.09 a | 0.77 ± 0.04 b | 1.00 ± 0.07 a | 1.16 ± 0.08 a | ||
| Secondary forest | 10–20 | 1.04 ± 0.10 a | 0.83 ± 0.07 a | 1.16 ± 0.07 a | 0.97 ± 0.05 a | 0.87 ± 0.06 a | 1.64 ± 0.17 a | |
| 20–30 | 1.12 ± 0.15 a | 1.08 ± 0.11 a | 1.11 ± 0.08 a | 0.98 ± 0.06 ab | 0.96 ± 0.05 b | 1.53 ± 0.15 ab | ||
| 30–40 | 0.78 ± 0.07 a | 0.75 ± 0.10 a | 0.78 ± 0.03 a | 0.88 ± 0.08 ab | 0.90 ± 0.04 a | 1.24 ± 0.09 a | ||
| Primary forest | 10–20 | 1.12 ± 0.12 a | 0.88 ± 0.06 a | 0.96 ± 0.07 a | 0.99 ± 0.08 a | 1.14 ± 0.08 a | 1.48 ± 0.10 a | |
| 20–30 | 0.99 ± 0.11 a | 1.06 ± 0.08 a | 0.92 ± 0.07 a | 0.97 ± 0.06 a | 1.25 ± 0.09 a | 1.21 ± 0.09 b | ||
| 30–40 | 1.09 ± 0.14 a | 0.88 ± 0.09 a | 0.94 ± 0.07 a | 1.04 ± 0.05 a | 1.03 ± 0.07 a | 1.01 ± 0.04 a | ||
| Vegetation type | Lowland rainforest | 10–20 | 1.10 ± 0.06 a | 1.05 ± 0.09 a | 0.89 ± 0.03 a | 1.10 ± 0.07 a | 0.81 ± 0.06 a | 1.10 ± 0.09 a |
| 20–30 | 0.95 ± 0.06 a | 1.02 ± 0.08 a | 1.05 ± 0.07 a | 0.97 ± 0.08 a | 0.92 ± 0.13 a | 0.97 ± 0.10 a | ||
| 30–40 | 1.01 ± 0.05 a | 1.01 ± 0.05 a | 0.94 ± 0.01 a | 0.99 ± 0.05 a | 1.14 ± 0.05 a | 0.99 ± 0.07 a | ||
| Montane rainforest | 10–20 | 1.11 ± 0.09 a | 1.04 ± 0.08 a | 1.00 ± 0.05 a | 1.04 ± 0.10 a | 0.62 ± 0.08 a | 1.04 ± 0.12 a | |
| 20–30 | 0.99 ± 0.05 a | 1.08 ± 0.06 a | 1.05 ± 0.04 a | 0.96 ± 0.04 a | 1.04 ± 0.09 a | 0.96 ± 0.06 a | ||
| 30–40 | 0.90 ± 0.07 a | 0.71 ± 0.07 b | 0.89 ± 0.08 a | 1.10 ± 0.05 a | 1.11 ± 0.09 a | 1.10 ± 0.07 a | ||
| Cloud forest | 10–20 | 1.14 ± 0.08 a | 1.11 ± 0.07 a | 1.05 ± 0.04 a | 1.01 ± 0.09 a | 0.87 ± 0.08 a | 1.01 ± 0.11 a | |
| 20–30 | 1.00 ± 0.09 a | 0.86 ± 0.07 a | 0.94 ± 0.05 a | 0.92 ± 0.05 a | 1.12 ± 0.02 a | 0.92 ± 0.07 a | ||
| 30–40 | 0.85 ± 0.05 a | 1.06 ± 0.06 a | 0.97 ± 0.07 a | 1.01 ± 0.08 a | 1.00 ± 0.09 a | 1.01 ± 0.10 a | ||
| Hilltop scrub | 10–20 | 0.96 ± 0.07 a | 1.10 ± 0.04 a | 0.88 ± 0.06 a | 0.98 ± 0.06 a | 0.80 ± 0.03 a | 0.98 ± 0.08 a | |
| 20–30 | 1.01 ± 0.04 a | 0.95 ± 0.02 a | 0.99 ± 0.07 a | 1.00 ± 0.07 a | 1.29 ± 0.06 a | 1.00 ± 0.07 a | ||
| 30–40 | 0.88 ± 0.09 a | 0.68 ± 0.05 b | 0.98 ± 0.07 a | 1.00 ± 0.09 a | 0.98 ± 0.04 a | 1.00 ± 0.09 a | ||
| Slope orientation | E | 10–20 | 1.00 ± 0.08 a | 1.05 ± 0.05 a | 1.01 ± 0.03 a | 0.85 ± 0.05 b | 1.05 ± 0.08 a | 1.59 ± 0.14 a |
| 20–30 | 0.94 ± 0.05 a | 0.93 ± 0.06 a | 0.98 ± 0.05 a | 0.96 ± 0.04 a | 0.98 ± 0.09 a | 1.23 ± 0.03 a | ||
| 30–40 | 1.12 ± 0.09 a | 1.08 ± 0.09 a | 1.05 ± 0.07 a | 1.01 ± 0.03 a | 1.00 ± 0.05 a | 0.95 ± 0.04 a | ||
| S | 10–20 | 1.09 ± 0.09 a | 0.99 ± 0.10 a | 1.08 ± 0.04 a | 1.04 ± 0.07 ab | 1.02 ± 0.04 a | 1.72 ± 0.08 a | |
| 20–30 | 1.11 ± 0.06 a | 1.03 ± 0.04 a | 1.03 ± 0.07 a | 1.04 ± 0.08 a | 1.01 ± 0.09 a | 1.23 ± 0.12 a | ||
| 30–40 | 1.02 ± 0.05 a | 1.00 ± 0.08 a | 1.03 ± 0.08 a | 0.89 ± 0.07 a | 1.02 ± 0.06 a | 1.02 ± 0.03 a | ||
| W | 10–20 | 0.94 ± 0.08 a | 0.92 ± 0.06 a | 0.97 ± 0.07 a | 0.94 ± 0.03 ab | 0.98 ± 0.06 a | 1.37 ± 0.12 a | |
| 20–30 | 1.02 ± 0.07 a | 0.87 ± 0.06 a | 1.03 ± 0.08 a | 0.90 ± 0.03 a | 0.86 ± 0.09 a | 1.19 ± 0.07 a | ||
| 30–40 | 0.97 ± 0.05 a | 1.04 ± 0.09 a | 1.09 ± 0.09 a | 0.98 ± 0.06 a | 1.07 ± 0.08 a | 0.94 ± 0.04 a | ||
| N | 10–20 | 1.10 ± 0.06 a | 1.05 ± 0.09 a | 0.89 ± 0.03 a | 1.10 ± 0.07 a | 0.81 ± 0.06 a | 1.58 ± 0.09 a | |
| 20–30 | 0.95 ± 0.06 a | 1.02 ± 0.08 a | 1.05 ± 0.07 a | 1.04 ± 0.06 a | 0.89 ± 0.09 a | 1.53 ± 0.06 a | ||
| 30–40 | 1.01 ± 0.05 a | 1.01 ± 0.05 a | 0.94 ± 0.01 a | 0.89 ± 0.07 a | 1.16 ± 0.03 a | 0.85 ± 0.08 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Li, J.; Xing, G.; Mai, S.; Liu, W.; Jiang, Y.; Xu, W.; Yang, Q.; Yang, H.; Lu, J.; et al. Soil Organic Carbon Distribution, Enzyme Activities, and the Temperature Sensitivity of a Tropical Rainforest in Wuzhishan, Hainan Island. Forests 2022, 13, 1943. https://doi.org/10.3390/f13111943
Wang X, Li J, Xing G, Mai S, Liu W, Jiang Y, Xu W, Yang Q, Yang H, Lu J, et al. Soil Organic Carbon Distribution, Enzyme Activities, and the Temperature Sensitivity of a Tropical Rainforest in Wuzhishan, Hainan Island. Forests. 2022; 13(11):1943. https://doi.org/10.3390/f13111943
Chicago/Turabian StyleWang, Xiaofang, Jialing Li, Guitong Xing, Siwei Mai, Wenjie Liu, Yamin Jiang, Wenxian Xu, Qiu Yang, Huai Yang, Jingli Lu, and et al. 2022. "Soil Organic Carbon Distribution, Enzyme Activities, and the Temperature Sensitivity of a Tropical Rainforest in Wuzhishan, Hainan Island" Forests 13, no. 11: 1943. https://doi.org/10.3390/f13111943