


Drought Offsets the Potential Effects of Nitrogen Addition on Soil Respiration and Organic Carbon in Model Subtropical Forests
 and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Sign
2.2. Soil Respiration Measurements
2.3. Aboveground Biomass and Belowground Biomass
2.4. Soil Organic Carbon, Microbial Carbon, and Dissolved Organic Carbon
2.5. Data Analysis
3. Results
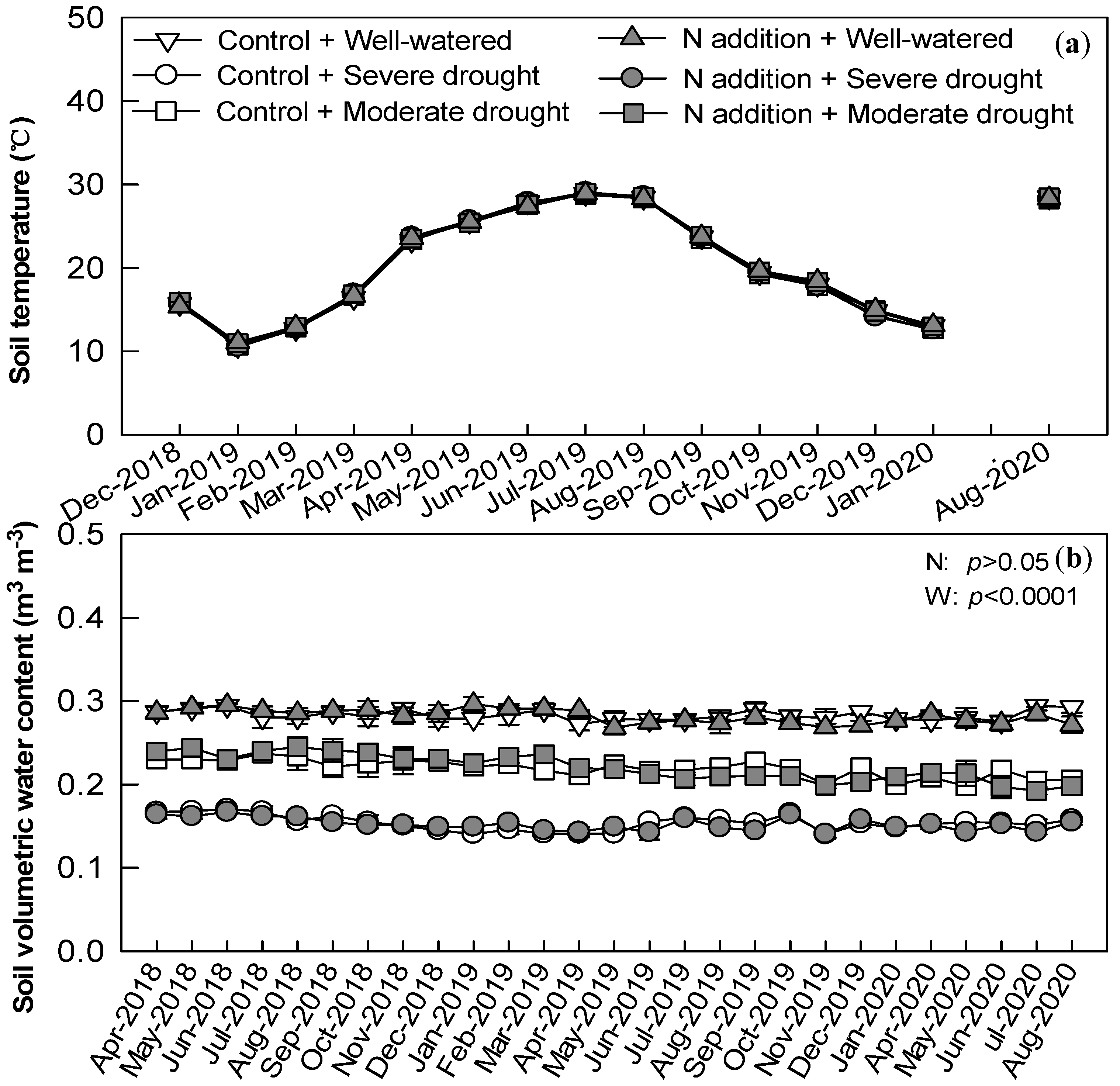
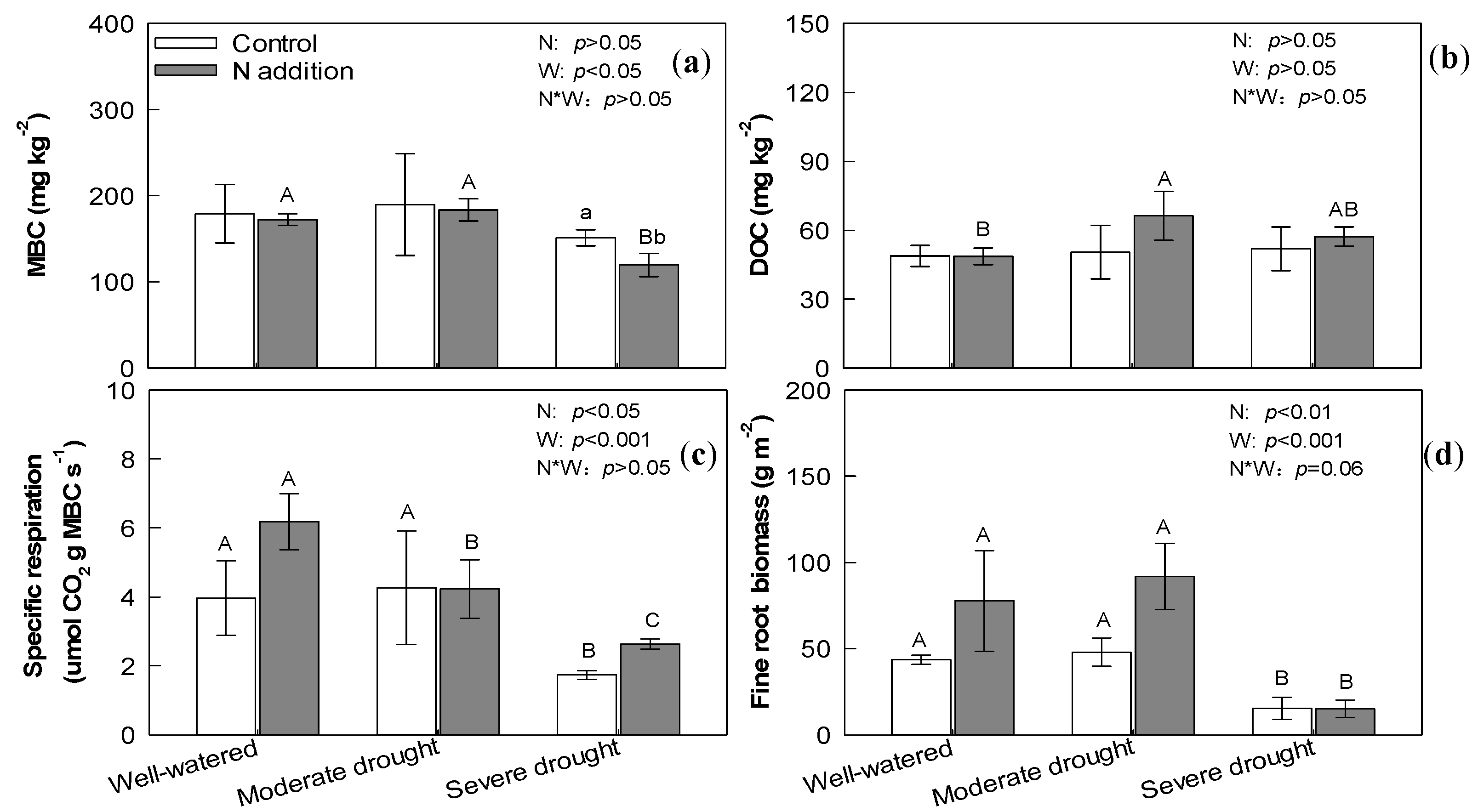
3.1. Soil Properties
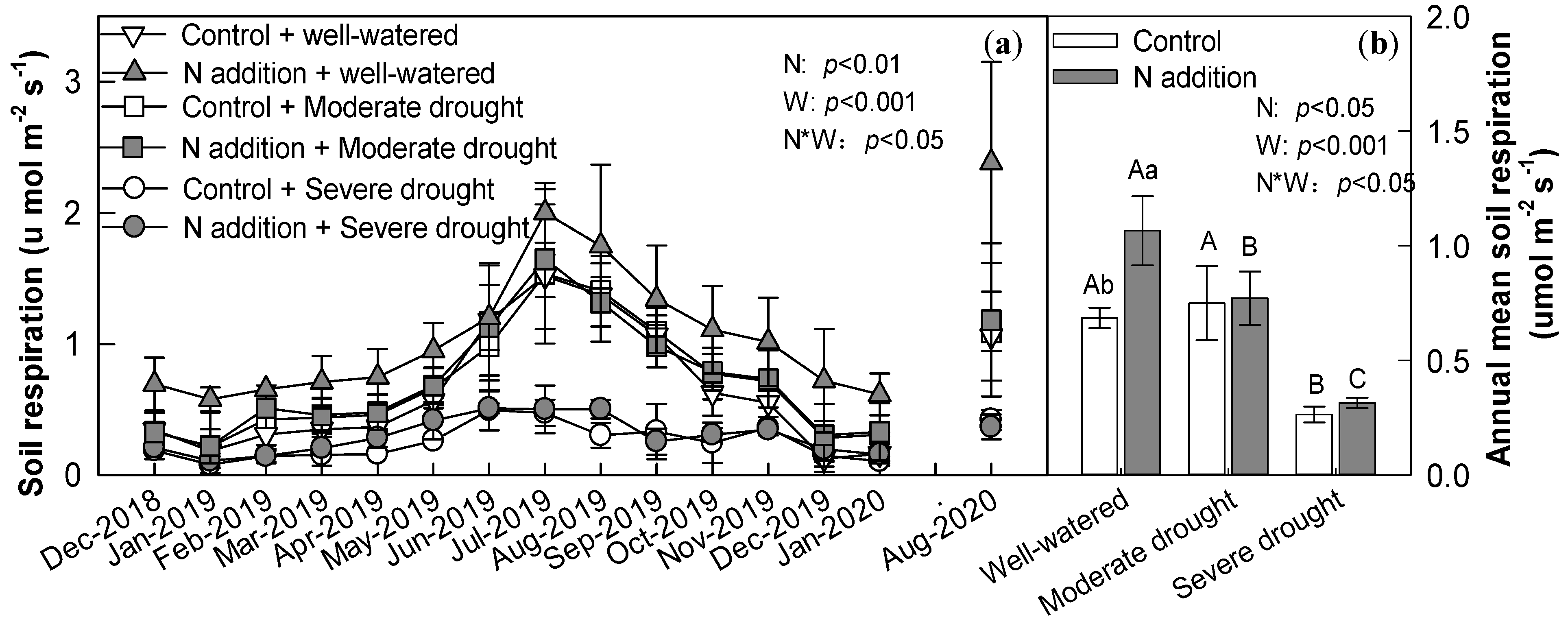
3.2. Soil Respiration
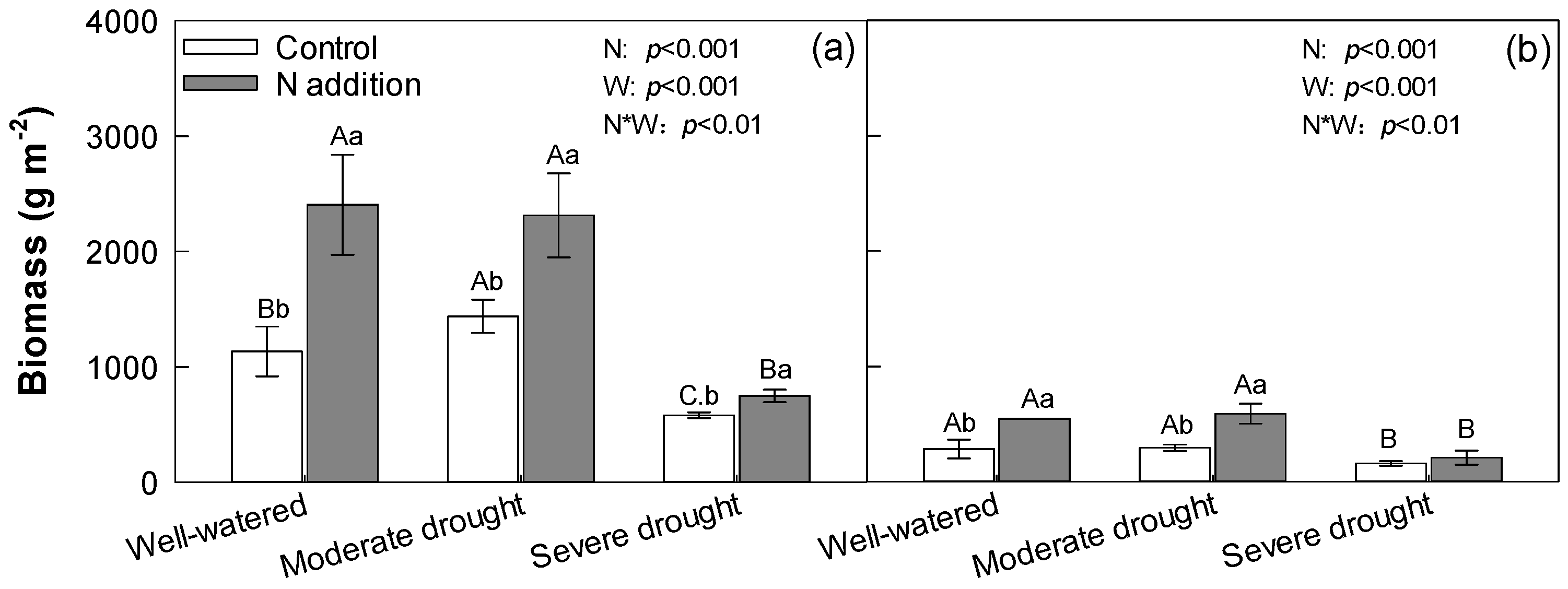
3.3. Aboveground Biomass and Belowground Biomass
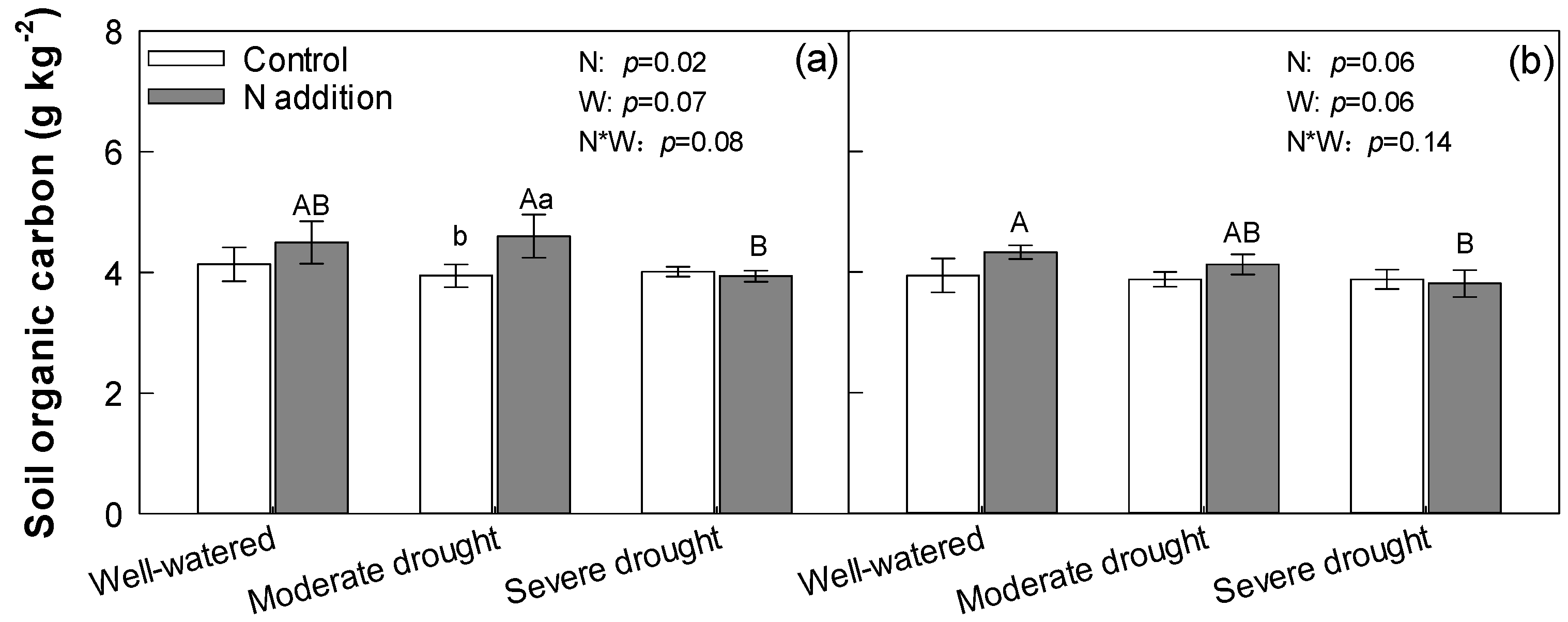
3.4. Soil Organic Carbon, Microbial Carbon, and Dissolved Organic Carbon
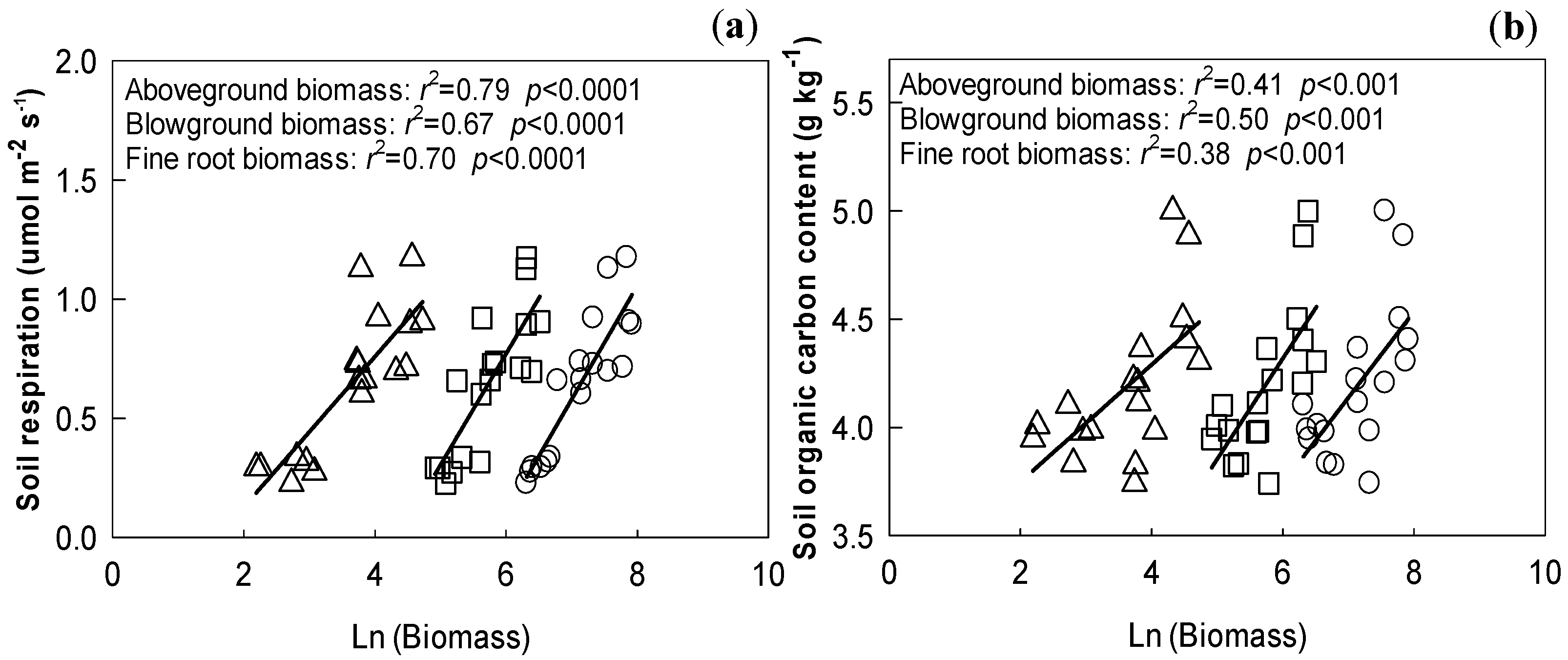
3.5. Relationships
4. Discussion
4.1. Effect of N Addition, Drought, and Their Interaction on Soil Respiration
4.2. Effect of N Addition, Drought, and Their Interaction on Soil Organic Carbon
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Satakhun, D.; Gay, F.; Chairungsee, N.; Kasemsap, P.; Chantuma, P.; Thanisawanyangkura, S.; Thaler, P.; Epron, D. Soil CO2 efflux and soil carbon balance of a tropical rubber plantation. Ecol. Res. 2013, 28, 969–979. [Google Scholar] [CrossRef]
- Sun, W.L.; Liu, X.H. Review on carbon storage estimation of forest ecosystem and applications in China. For. Ecosyst. 2020, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Schlesinger, W.H.; Andrews, J.A. Soil respiration and the global carbon cycle. Biogeochemistry 2000, 48, 7–20. [Google Scholar] [CrossRef]
- Davidson, E.A. The contribution of manure and fertilizer nitrogen to atmospheric nitrous oxide since 1860. Nat. Geosci. 2009, 2, 659–662. [Google Scholar] [CrossRef]
- Gruber, N.; Galloway, J.N. An earth-system perspective of the global nitrogen cycle. Nature 2008, 451, 293–296. [Google Scholar] [CrossRef] [Green Version]
- Pussinen, A.; Karjalainen, T.; Mäkipää, R.; Valsta, L.; Kellomäki, L. Forest carbon sequestration and harvests in Scots pine stand under different climate and nitrogen deposition scenarios. For. Ecol. Manag. 2002, 158, 103–115. [Google Scholar] [CrossRef]
- Liu, X.; Yang, T.; Wang, Q.; Huang, F.R.; Li, L.H. Dynamics of soil carbon and nitrogen stocks after afforestation in arid and semi-arid regions: A meta-analysis. Sci. Total Environ. 2018, 618, 1658–1664. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Balogh, J.; Papp, M.; Pintér, K.; Fóti, S.; Posta, K.; Eugster, W.; Nagy, Z. Autotrophic component of soil respiration is repressed by drought more than the heterotrophic one in dry grasslands. Biogeosciences 2016, 13, 5171–5182. [Google Scholar] [CrossRef] [Green Version]
- Deng, L.; Peng, C.H.; Kim, D.G.; Li, J.W.; Liu, Y.L.; Hai, X.Y.; Liu, Q.Y.; Huang, C.B.; Shangguan, Z.P.; Kuzyakov, Y. Drought effects on soil carbon and nitrogen dynamics in global natural ecosystems. Earth-Sci. Rev. 2021, 214, 103501. [Google Scholar] [CrossRef]
- Cook, F.J.; Orchard, V.A. Relationships between soil respiration and soil moisture. Soil Biol. Biochem. 2008, 40, 1013–1018. [Google Scholar] [CrossRef]
- Liu, L.L.; Greaver, T.L. A global perspective on belowground carbon dynamics under nitrogen enrichment. Ecol. Lett. 2010, 13, 819–828. [Google Scholar] [CrossRef]
- Huang, X.M.; Lakso, A.N.; Eissenstat, D.M. Interactive effects of soil temperature and moisture on Concord grape root respiration. J. Exp. Bot. 2005, 56, 2651–2660. [Google Scholar] [CrossRef] [Green Version]
- Jing, H.; Liu, Y.; Wang, G.L.; Liu, G.B. Effects of nitrogen addition on root respiration of trees and understory herbs at different temperatures in Pinus tabulaeformis forest. Plant Soil 2021, 463, 447–459. [Google Scholar] [CrossRef]
- Bowden, R.D.; Davidson, E.; Savage, K.; Arabia, K.; Steudler, P. Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest. For. Ecol. Manag. 2004, 196, 43–56. [Google Scholar] [CrossRef]
- Yan, Z.F.; Bond-Lamberty, B.; Todd-Brown, K.E.; Bailey, V.L.; Li, S.L.; Liu, C.Q.; Liu, C.X. A moisture function of soil heterotrophic respiration that incorporates microscale processes. Nat. Commun. 2018, 9, 2562. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.Y.; Zhou, X.H.; Shao, J.J.; Nie, Y.Y.; He, Y.H.; Jiang, L.L.; Wu, Z.T.; Hosseini Bai, S. Interactive effects of global change factors on soil respiration and its components: A meta-analysis. Glob. Chang. Biol. 2016, 22, 3157–3169. [Google Scholar] [CrossRef]
- Lu, X.K.; Vitousek, P.M.; Mao, Q.G.; Gilliam, F.S.; Luo, Y.Q.; Turner, B.L.; Zhou, G.Y.; Mo, J.M. Nitrogen deposition accelerates soil carbon sequestration in tropical forests. Proc. Natl. Acad. Sci. USA 2021, 118, e2020790118. [Google Scholar] [CrossRef]
- Janssens, I.A.; Dieleman, W.; Luyssaert, S.; Subke, J.A.; Reichstein, M.; Ceulemans, R.; Ciais, P.; Dolman, A.J.; Grace, J.; Matteucci, G.; et al. Reduction of forest soil respiration in response to nitrogen deposition. Nat. Geosci. 2010, 3, 315–322. [Google Scholar] [CrossRef]
- Sun, Z.Z.; Liu, L.L.; Ma, Y.C.; Yin, G.D.; Zhao, C.; Zhang, Y.; Piao, S.L. The effect of nitrogen addition on soil respiration from a nitrogen-limited forest soil. Agric. For. Meteorol. 2014, 197, 103–110. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Burton, A.J.; Zak, D.R.; Talhelm, A.F. Simulated chronic nitrogen deposition increases carbon storage in northern temperate forests. Glob. Chang. Biol. 2008, 14, 142–153. [Google Scholar] [CrossRef]
- Xu, W.H.; Wan, S.Q. Water- and plant-mediated responses of soil respiration to topography, fire, and nitrogen fertilization in a semiarid grassland in northern China. Soil Biol. Biochem. 2008, 40, 679–687. [Google Scholar] [CrossRef]
- Mack, M.C.; Schuur, E.A.; Bret-Harte, M.S.; Shaver, G.R.; Chapin, F.S. Ecosystem carbon storage in arctic tundra reduced by long-term nutrient fertilization. Nature 2004, 431, 440–443. [Google Scholar] [CrossRef] [PubMed]
- Everard, K.; Seabloom, E.W.; Harpole, W.S.; de Mazancourt, C. Plant water use affects competition for nitrogen: Why drought favors invasive species in California. Am. Nat. 2010, 175, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Yan, G.Y.; Xing, Y.J.; Zhang, J.H.; Wang, Q.G.; Wang, H.L.; Huang, B.B.; Hong, Z.M.; Dai, G.H.; Zheng, X.B.; et al. Effects of N addition and precipitation reduction on soil respiration and its components in a temperate forest. Agric. For. Meteorol. 2019, 271, 336–345. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, S.R.; Luan, J.W.; Chen, C.; Cai, C.J.; Zhou, F.; Di, Y.P.; Gao, X.M. Nitrogen Addition Exacerbates the Negative Effect of Throughfall Reduction on Soil Respiration in a Bamboo Forest. Forests 2021, 12, 724. [Google Scholar] [CrossRef]
- Harpole, W.S.; Potts, D.I.; Suding, K.N. Ecosystem responses to water and nitrogen amendment in a California grassland. Glob. Chang. Biol. 2007, 13, 2341–2348. [Google Scholar] [CrossRef]
- Valliere, J.M.; Allen, E.B. Interactive effects of nitrogen deposition and drought-stress on plant-soil feedbacks of Artemisia californica seedlings. Plant Soil 2016, 403, 277–290. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.M.; Chen, S.P.; Huang, J.H.; Lin, G.H. Differential responses of auto-and hetero-trophic soil respiration to water and nitrogen addition in a semiarid temperate steppe. Glob. Chang. Biol. 2010, 16, 2345–2357. [Google Scholar] [CrossRef]
- McCulley, R.L.; Boutton, T.W.; Archer, S.R. Soil respiration in a subtropical savanna parkland: Response to water additions. Soil Sci. Soc. Am. J. 2007, 71, 820–828. [Google Scholar] [CrossRef]
- Xu, X.K.; Yin, L.; Duan, C.T.; Jing, Y.S. Effect of N addition, moisture, and temperature on soil microbial respiration and microbial biomass in forest soil at different stages of litter decomposition. J. Soils Sediments 2016, 16, 1421–1439. [Google Scholar] [CrossRef]
- Zhang, X.L.; Tan, Y.L.; Zhang, B.W.; Li, A.; Daryanto, S.; Wang, L.X.; Huang, J.H. The impacts of precipitation increase and nitrogen addition on soil respiration in a semiarid temperate steppe. Ecosphere 2017, 8, e01655. [Google Scholar] [CrossRef]
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, W.; Fu, W.J.; Jiang, P.K.; Zhao, K.L.; Li, Y.H.; Tao, J.X. Spatial pattern of carbon stocks in forest ecosystems of a typical subtropical region of southeastern China. For. Ecol. Manag. 2018, 409, 288–297. [Google Scholar] [CrossRef]
- Mo, J.M.; Brown, S.; Xue, J.H.; Fang, Y.T.; Li, Z.A. Response of litter decomposition to simulated N deposition in disturbed, rehabilitated and mature forests in subtropical China. Plant Soil 2006, 282, 135–151. [Google Scholar] [CrossRef]
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.C.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.Y.; Wang, Z.C.; Liu, H.H.; Zhang, C.; Fu, S.L.; Fang, X. Responses in growth and anatomical traits of two subtropical tree species to nitrogen addition, drought, and their Interactions. Front. Plant Sci. 2021, 12, 1606. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, J.; Ostle, N.; McNamara, N.P.; Nottingham, A.T.; Stott, A.W.; Bardgett, R.D.; Salinas, N.; Ccahuana, A.J.Q.; Meir, P. Microbial carbon mineralization intropical lowland and montane forest soils of Peru. Front. Microbiol. 2014, 5, 720. [Google Scholar] [CrossRef]
- Xing, G.X.; Shi, S.L.; Shen, G.Y.; Du, L.J.; Xiong, Z.Q. Nitrous oxide emissions from paddy soil in three rice-based cropping systems in China. Nutr. Cycl. Agroecosystems 2002, 64, 135–143. [Google Scholar] [CrossRef]
- Fang, X.; Zhu, Y.L.; Liu, J.D.; Lin, X.P.; Sun, H.Z.; Tang, X.H.; Hu, Y.L.; Huang, Y.P.; Yi, Z.G. Effects of moisture and temperature on soil organic carbon decomposition along a vegetation restoration gradient of subtropical China. Forests 2022, 13, 578. [Google Scholar] [CrossRef]
- Liu, J.X.; Liu, S.G.; Li, Y.Y.; Liu, S.Z.; Yin, G.C.; Huang, J.; Xu, Y.; Zhou, G.Y. Warming effects on the decomposition of two litter species in model subtropical forests. Plant Soil 2017, 409, 277–287. [Google Scholar] [CrossRef]
- Wu, J.; Joergensen, R.G.; Pommerening, B.; Chaussod, R.; Brookes, P.C. Measurement of soil microbial biomass C by fumigation-extraction-an automated procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, Q.M.; Wang, Z.X.; Zheng, H.F.; Chen, Y.M.; Chen, X.; Wang, L.F.; Li, H.J.; Zhang, J. Nitrogen addition alleviates microbial nitrogen limitations and promotes soil respiration in a Subalpine Coniferous forest. Forests 2019, 10, 1038. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.P.; Niu, D.C.; Hall, S.J.; Wen, H.Y.; Li, X.D.; Fu, H.; Wan, C.G.; Elser, J.J. Effects of simulated nitrogen deposition on soil respiration components and their temperature sensitivities in a semiarid grassland. Soil Biol. Biochem. 2014, 75, 113–123. [Google Scholar] [CrossRef]
- Sun, S.; Lei, H.; Chang, S.X. Drought differentially affects autotrophic and heterotrophic soil respiration rates and their temperature sensitivity. Biol. Fertil. Soils 2019, 55, 275–283. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. N. Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Karst, J.; Gaster, J.; Wiley, E.; Landhäusser, S.M. Stress differentially causes roots of tree seedlings to exude carbon. Tree Physiol. 2017, 37, 154–164. [Google Scholar] [CrossRef] [Green Version]
- Preece, C.; Farré-Armengol, G.; Llusià, J.; Peñuelas, J. Thirsty tree roots exude more carbon. Tree Physiol. 2018, 38, 690–695. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.B.; Wang, D.L.; Shi, B.K.; Sun, W. Differential effects of grazing, water, and nitrogen addition on soil respiration and its components in a meadow steppe. Plant Soil 2020, 447, 581–598. [Google Scholar] [CrossRef]
- Bi, J.; Zhang, N.L.; Liang, Y.; Yang, H.J.; Ma, K.P. Interactive effects of water and nitrogen addition on soil microbial communities in a semiarid steppe. J. Plant Ecol. 2011, 5, 320–329. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, U.; von Oheimb, G.; Kriebitzsch, W.U.; Schlesselmann, K.; Weber, M.S.; Hardtle, W. Nitrogen deposition increases susceptibility to drought-experimental evidence with the perennial grass Molinia caerulea (L) Moench. Plant Soil 2012, 353, 59–71. [Google Scholar] [CrossRef]
- Zak, D.R.; Holmes, W.E.; Burton, A.J.; Pregitzer, K.S.; Talhelm, A.F. Simulated atmospheric NO−3 deposition increases soil organic matter by slowing decomposition. Ecol. Appl. 2008, 18, 2016–2027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, C.D.; Harrison, R.B. The Case for Digging Deeper: Soil Organic Carbon Storage, Dynamics, and Controls in Our Changing World. Soil Syst. 2019, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Zhang, Y.S.; Guan, D.S.; Xiao, L.; Singh, M. The potential of mature Sonneratia apetala plantations to enhance carbon stocks in the Zhanjiang Mangrove National Nature Reserve. Ecol. Indic. 2021, 133, 108415. [Google Scholar] [CrossRef]
- Jaman, M.S.; Wu, H.; Yu, Q.; Tan, Q.; Zhang, Y.; Dam, Q.K.; Muraina, T.O.; Xu, C.; Jing, M.; Jia, X.; et al. Contrasting responses of plant above and belowground biomass carbon pools to extreme drought in six grasslands spanning an aridity gradient. Plant Soil 2022, 473, 167–180. [Google Scholar] [CrossRef]
- de Vries, F.T.; Brown, C.; Stevens, C.J. Grassland species root response to drought: Consequences for soil carbon and nitrogen availability. Plant Soil 2016, 409, 297–312. [Google Scholar] [CrossRef]
- Knapp, A.K.; Carroll, C.J.; Denton, E.M.; La Pierre, K.J.; Collins, S.L.; Smith, M.D. Differential sensitivity to regional-scale drought in six central US grasslands. Oecologia 2015, 177, 949–957. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Y.B.; Huang, C.D.; Hu, T.X.; Tu, L.H.; Zhou, S.X.; Xiao, Y.X.; Gao, B.D. Effects of simulated nitrogen deposition and precipitation changes on soil respiration in an evergreen broad-leaved forest in the Rainy Area of Western China. Acta Ecol. Sin. 2017, 36, 5235. [Google Scholar]
- Chen, Z.J.; Zhou, X.Y.; Geng, S.C.; Miao, Y.; Cao, Y.H.; Chen, Z.; Zhang, J.H.; Han, S.J. Interactive effect of nitrogen addition and throughfall reduction decreases soil aggregate stability through reducing biological binding agents. For. Ecol. Manag. 2019, 445, 13–19. [Google Scholar] [CrossRef]
- Högberg, P.; Johannisson, C.; Hällgren, J.E. Studies of 13C in the foliage reveal interactions between nutrients and water in forest fertilization experiments. Plant Soil 1993, 152, 207–221. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | pH | Total N (g kg−1) | NO−3-N (mg kg−1) | NH+4-N (mg kg−1) | AP (mg kg−1) |
|---|---|---|---|---|---|
| Well-watered | |||||
| Control | 6.18 ± 0.14 a | 0.37 ± 0.02 | 1.99 ± 0.23 | 2.20 ± 0.57 | 5.42 ± 0.55 A |
| N addition | 6.02 ± 0.16 a | 0.39 ± 0.03 | 2.55 ± 0.84 b | 1.74 ± 0.50 c | 3.48 ± 0.80 B |
| Moderate drought | |||||
| Control | 5.98 ± 0.04 b | 0.37 ± 0.01 B | 2.56 ± 1.99 | 1.90 ± 0.42 | 4.60 ± 0.66 |
| N addition | 5.86 ± 1.18 a | 0.42 ± 0.03 A | 4.20 ± 0.40 b | 2.57 ± 0.43 b | 3.70 ± 0.06 |
| Severe drought | |||||
| Control | 5.98 ± 0.11 bA | 0.38 ± 0.04 | 3.92 ± 1.42 B | 2.28 ± 0.86 B | 5.79 ± 1.31 |
| N addition | 5.44 ± 0.03 bB | 0.41 ± 0.04 | 12.17 ± 5.27 aA | 3.96 ± 0.51 aA | 4.41 ± 0.31 |
| Analysis of variance (p values) | |||||
| N | <0.001 | 0.034 | <0.001 | 0.035 | 0.002 |
| W | <0.001 | 0.800 | 0.003 | 0.011 | 0.109 |
| N × W | 0.213 | 0.621 | 0.035 | 0.317 | 0.220 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.-l.; Lin, X.-p.; Huang, Y.-p.; Tang, X.-h.; Fang, X.; Yi, Z.-g. Drought Offsets the Potential Effects of Nitrogen Addition on Soil Respiration and Organic Carbon in Model Subtropical Forests. Forests 2022, 13, 1615. https://doi.org/10.3390/f13101615
Zhu Y-l, Lin X-p, Huang Y-p, Tang X-h, Fang X, Yi Z-g. Drought Offsets the Potential Effects of Nitrogen Addition on Soil Respiration and Organic Carbon in Model Subtropical Forests. Forests. 2022; 13(10):1615. https://doi.org/10.3390/f13101615
Chicago/Turabian StyleZhu, Yu-lin, Xue-ping Lin, Yun-peng Huang, Xing-hao Tang, Xiong Fang, and Zhi-gang Yi. 2022. "Drought Offsets the Potential Effects of Nitrogen Addition on Soil Respiration and Organic Carbon in Model Subtropical Forests" Forests 13, no. 10: 1615. https://doi.org/10.3390/f13101615