
The Biological Origins of Soil Organic Matter in Different Land-Uses in the Highlands of Ethiopia



Abstract
:1. Introduction
2. Materials and Methods
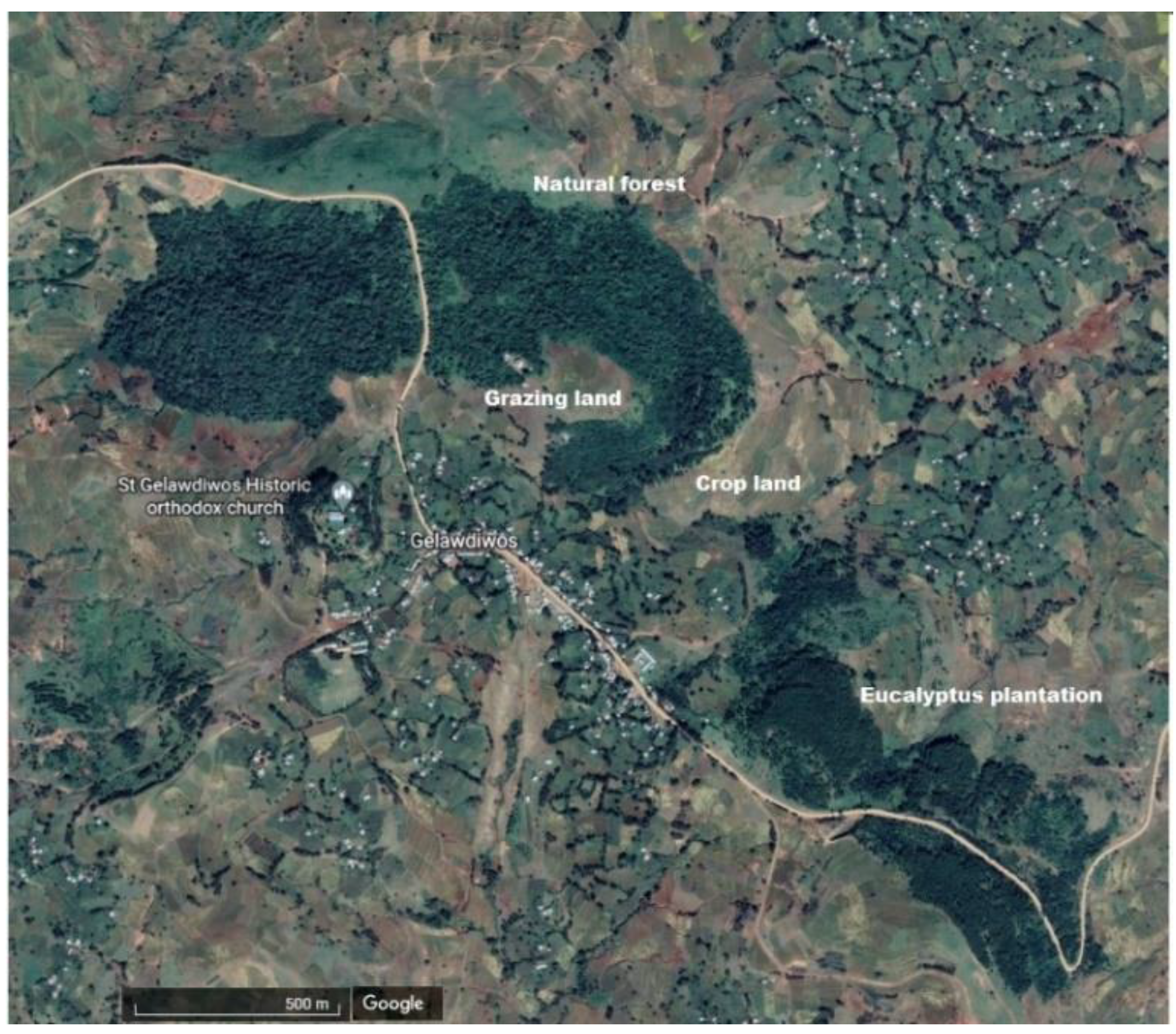
2.1. Study Area
2.2. Soil Sampling, C and N Analysis
2.3. Sequential Chemical Extraction
2.4. Derivatization and GC/MS Analysis
2.5. Calculated Ratios and Indices
2.5.1. Microbial Biomarkers
2.5.2. Suberin and Cutin Monomers
2.5.3. Lignin-Derived Phenols
2.6. Statistical Analysis
3. Results
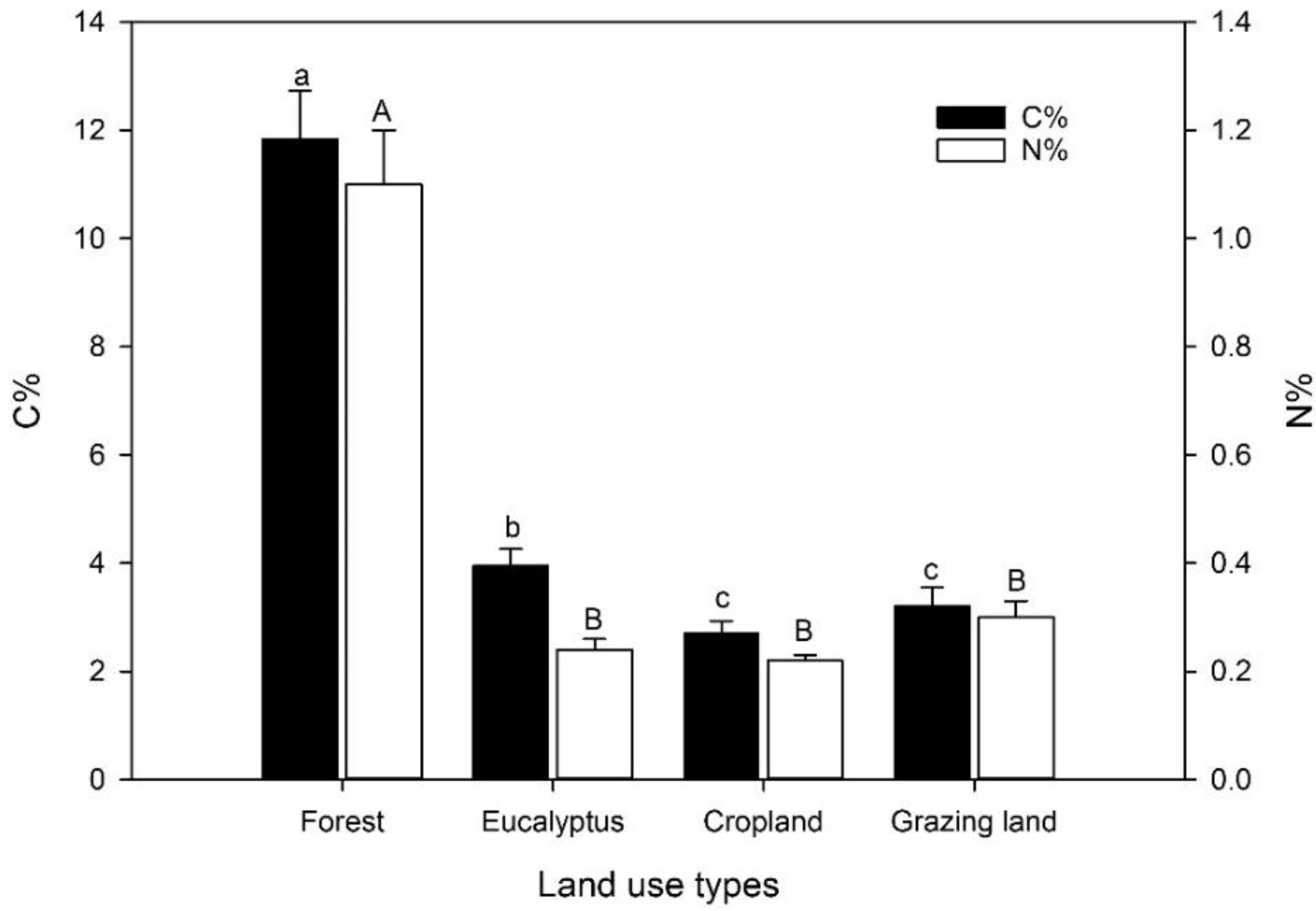
3.1. Soil C and N Content, and Yields of Sequential Extractions
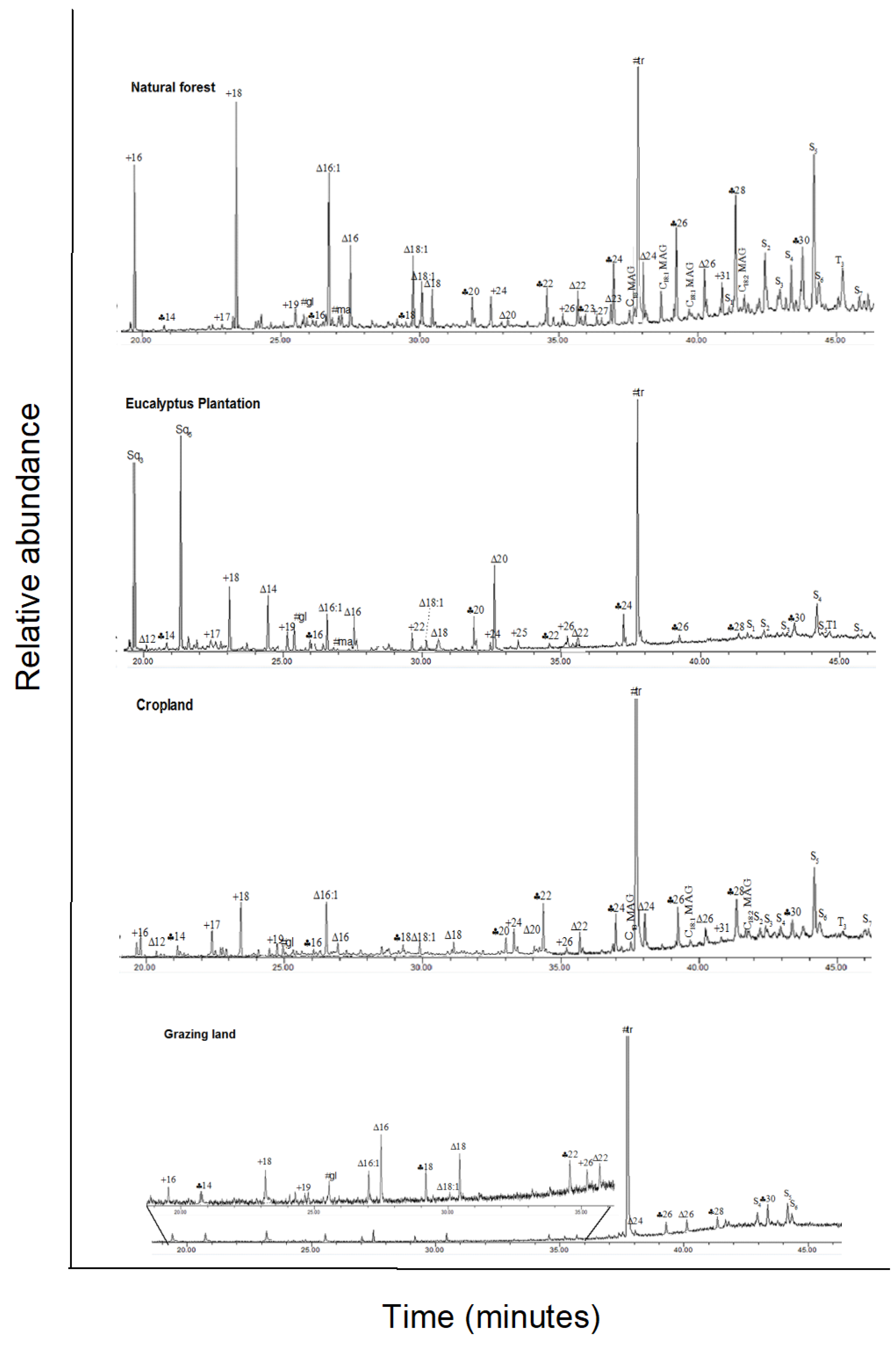
3.2. Solvent-Extractable Compounds
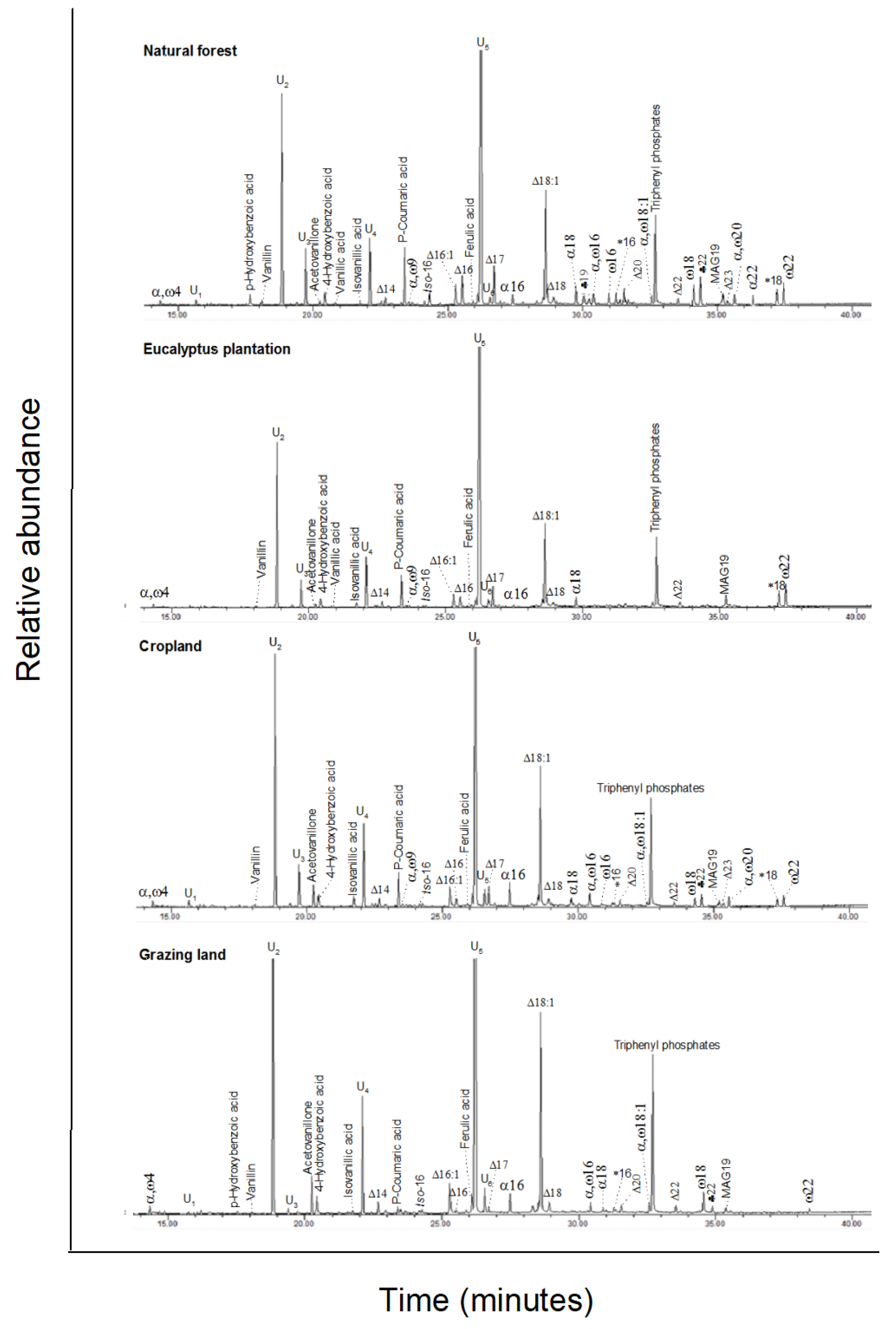
3.3. Base Hydrolysis Extractable Compounds
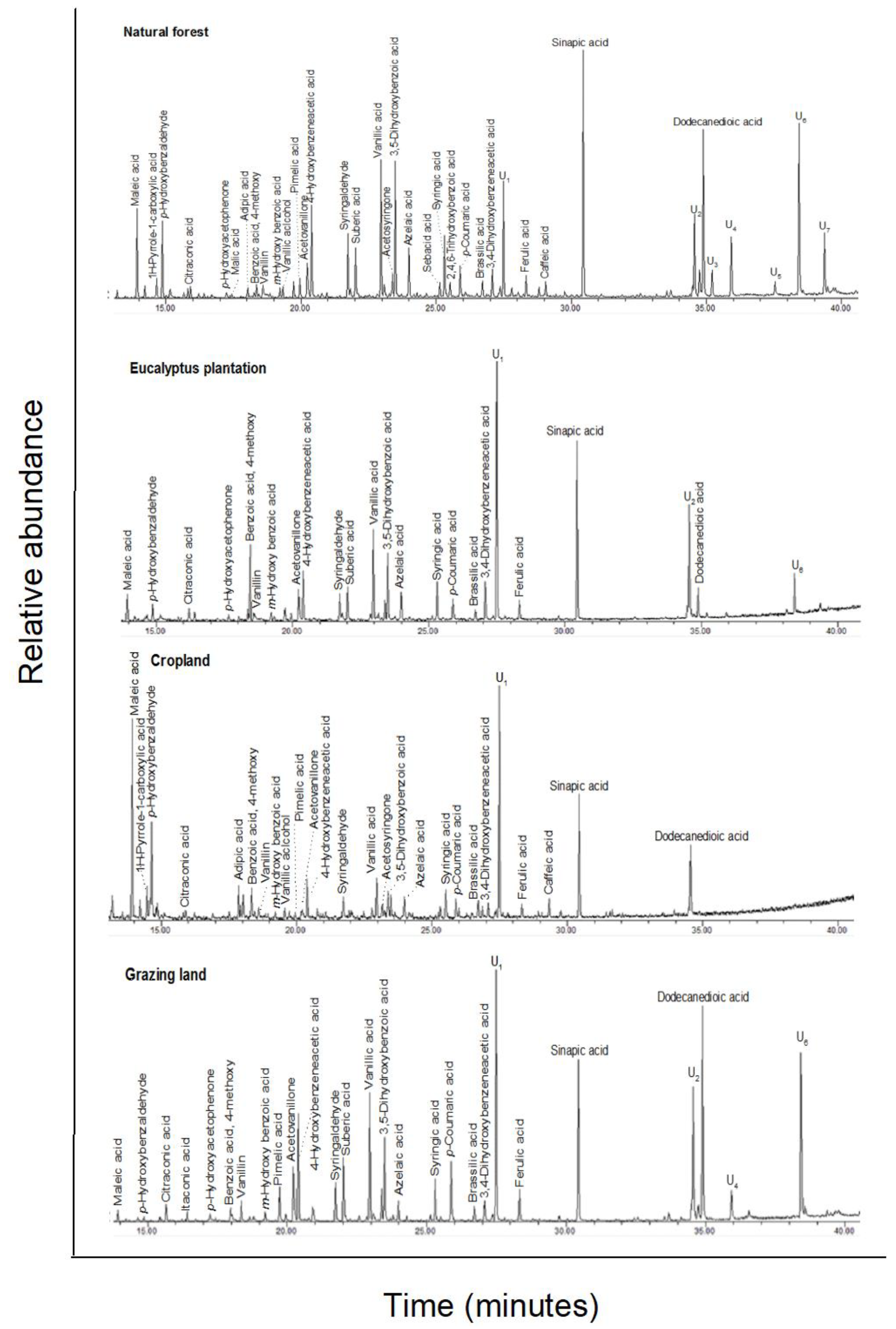
3.4. CuO Oxidation Extractable Compounds
4. Discussion
4.1. Above- and Belowground Inputs
4.2. Microbial Inputs
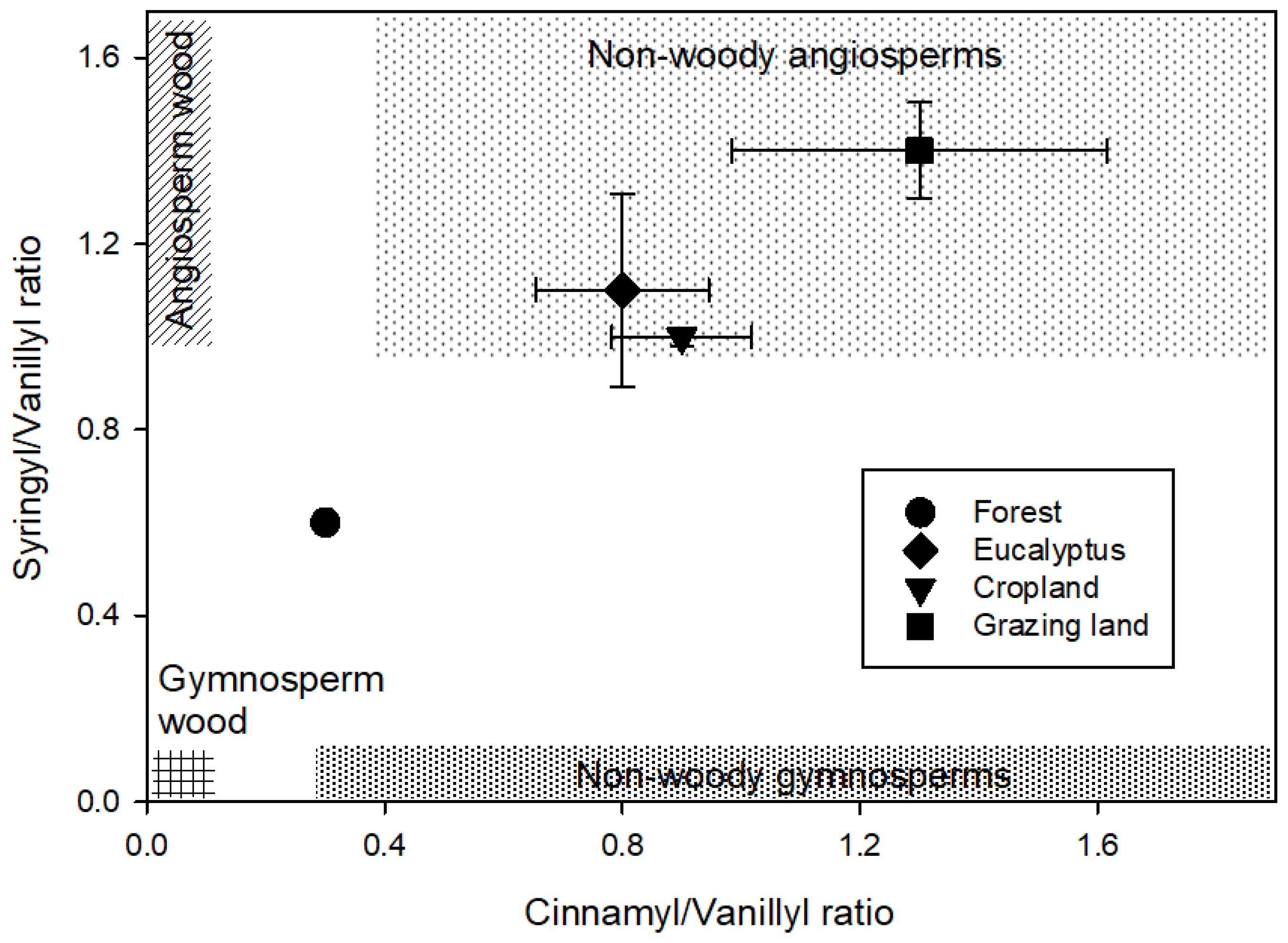
4.3. Angiosperms and Gymnosperms
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

 , n-alkanols; +, n-alkanes; Δ, n-alkanoic acids; #,
carbohydrates (gl, glucose; ma, mannose; su, sucrose); MAG, monoacylglycerides;
S1–S7, steroids; T1–T5,
triterpenoids; U1, unidentified. Numbers refer to total carbon
numbers in aliphatic lipid series. Detailed description of each compound with
its quantity, molecular formula, and molecular weight in Appendix A Table A1.
, n-alkanols; +, n-alkanes; Δ, n-alkanoic acids; #,
carbohydrates (gl, glucose; ma, mannose; su, sucrose); MAG, monoacylglycerides;
S1–S7, steroids; T1–T5,
triterpenoids; U1, unidentified. Numbers refer to total carbon
numbers in aliphatic lipid series. Detailed description of each compound with
its quantity, molecular formula, and molecular weight in Appendix A Table A1.
, n-alkanols; +, n-alkanes; Δ, n-alkanoic acids; #,
carbohydrates (gl, glucose; ma, mannose; su, sucrose); MAG, monoacylglycerides;
S1–S7, steroids; T1–T5,
triterpenoids; U1, unidentified. Numbers refer to total carbon
numbers in aliphatic lipid series. Detailed description of each compound with
its quantity, molecular formula, and molecular weight in Appendix A Table A1.
, n-alkanols; +, n-alkanes; Δ, n-alkanoic acids; #,
carbohydrates (gl, glucose; ma, mannose; su, sucrose); MAG, monoacylglycerides;
S1–S7, steroids; T1–T5,
triterpenoids; U1, unidentified. Numbers refer to total carbon
numbers in aliphatic lipid series. Detailed description of each compound with
its quantity, molecular formula, and molecular weight in Appendix A Table A1.
 , n-alkanols;
+, n-alkanes; Δ, n-alkanoic acids; iso-alkanoic acids; α-alkanoic acids; α,ω-alkanedioic acids; ω-hydroxyalkanoic acids; * Mid chain substituted
hydroxy acids; steroids (β-sitpsterol);
organophosphate; phenols; and U1–U5, unidentified.
Numbers refer to total carbon numbers in aliphatic lipid series. Detailed
description of each compound, with its quantity, molecular formula, and
molecular weight, is in Appendix A Table A2.
, n-alkanols;
+, n-alkanes; Δ, n-alkanoic acids; iso-alkanoic acids; α-alkanoic acids; α,ω-alkanedioic acids; ω-hydroxyalkanoic acids; * Mid chain substituted
hydroxy acids; steroids (β-sitpsterol);
organophosphate; phenols; and U1–U5, unidentified.
Numbers refer to total carbon numbers in aliphatic lipid series. Detailed
description of each compound, with its quantity, molecular formula, and
molecular weight, is in Appendix A Table A2.
, n-alkanols;
+, n-alkanes; Δ, n-alkanoic acids; iso-alkanoic acids; α-alkanoic acids; α,ω-alkanedioic acids; ω-hydroxyalkanoic acids; * Mid chain substituted
hydroxy acids; steroids (β-sitpsterol);
organophosphate; phenols; and U1–U5, unidentified.
Numbers refer to total carbon numbers in aliphatic lipid series. Detailed
description of each compound, with its quantity, molecular formula, and
molecular weight, is in Appendix A Table A2.
, n-alkanols;
+, n-alkanes; Δ, n-alkanoic acids; iso-alkanoic acids; α-alkanoic acids; α,ω-alkanedioic acids; ω-hydroxyalkanoic acids; * Mid chain substituted
hydroxy acids; steroids (β-sitpsterol);
organophosphate; phenols; and U1–U5, unidentified.
Numbers refer to total carbon numbers in aliphatic lipid series. Detailed
description of each compound, with its quantity, molecular formula, and
molecular weight, is in Appendix A Table A2.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound 3 | MF | MW | Forest | Eucalyptus | Cropland | Grazing Land |
|---|---|---|---|---|---|---|
| n-Alkanols | ||||||
| 2-Tetradecanol | C14H30O | 226 | 65.3 ± 16.4 | 5.8 1 | ||
| 1-Hexadecanol | C16H34O | 242 | 43.3 2 | 3.7 1 | ||
| 1-Octadecanol | C18H38O | 270 | 2.1 1 | 6.0 1 | 25.1 1 | |
| n-Eicosanol | C20H42O | 298 | 9.2 2 | 50.7 2 | 5.1 1 | |
| n-Docosanol | C22H46O | 326 | 45.2 ± 25.4 | 96.8 ± 8.0 | 55.1 ± 10.2 | 46.9 ± 19.3 |
| 1-Tricosanol | C23H48O | 340 | 6.7 2 | 6.2 1 | ||
| n-Tetracosanol | C24H50O | 354 | 41.9 ± 29.9 | 80.2 ± 20.1 | 40.6 ± 18.1 | 69.8 ± 15.7 |
| n-Hexacosanol | C26H54O | 382 | 84.0 ± 38.2 | 73.0 2 | 47.4 ± 11.7 | 60.5 2 |
| n-Octacosanol | C28H58O | 410 | 93.7 ± 46.7 | 77.6 2 | 86.0 ± 39.5 | 66.2 ± 14.8 |
| n-Tricontanol | C30H62O | 438 | 41.4 2 | 33.4 1 | 81.4 ± 15.3 | |
| Total Alkanols | 324 ± 172 | 520 ± 212 | 250 ± 86 | 356 ± 103 | ||
| n-Alkanes | ||||||
| n-Heptadecane | C17H36 | 240 | 162 ± 72 | 16.0 2 | ||
| Octadecane | C18H38 | 254 | 1.1 1 | 58.6 2 | ||
| Nonadecane | C19H40 | 268 | 81.1 2 | 8.6 1 | ||
| n-Docosane | C22H46 | 310 | 4.8 1 | 3.5 1 | ||
| Tricosane | C23H48 | 324 | 4.0 1 | 3.1 1 | ||
| n-Tetracosane | C24H50 | 338 | 6.2 2 | |||
| n-Pentacosane | C25H52 | 352 | 16.9 1 | 18.1 ± 0.8 | 11.7 2 | |
| n-Hexacosane | C26H54 | 366 | 19.6 1 | 29.6 ± 8.2 | ||
| n-Heptacosane | C27H56 | 380 | 201 ± 76 | 45.0 1 | 39.9 1 | 1.9 1 |
| Hentriacontane | C31H64 | 436 | 13.6 2 | 27.2 1 | 27.6 1 | 8.0 1 |
| Total Alkanes | 222 ± 71 | 391 ± 78 | 139 ± 66 | 58 ± 15 | ||
| n-Alkanoic Acids | ||||||
| Nonanoic acid | C9H18O2 | 158 | 8.7 2 | 14.5 1 | 38.7 ± 6.2 | 37.6 ± 13.1 |
| Dodecanoic acid | C12H24O2 | 200 | 188.3 ± 33.6 | 19.4 2 | 10.9 2 | |
| n-Tetradecanoic acid | C14H28O2 | 228 | 10.0 2 | 149.3 ± 18.5 | 4.3 1 | 11.2 1 |
| n-Hexadecanoic acid | C16H32O2 | 256 | 48.9 ± 25.4 | 169.8 ± 22.1 | 63.9 ± 22.0 | 34.0 ± 2.4 |
| n-Octadecanoic acid (18:1) | C18H34O2 | 282 | 20.5 2 | 132.4 ± 17.3 | 24.9 2 | |
| 9-Octadecanoic acid (Z) | C18H34O2 | 282 | 10.3 2 | 89.9 ± 20.9 | 12.5 2 | |
| n-Octadecanoic acid | C18H36O2 | 284 | 21.1 ± 7.5 | 114.7 ± 41.7 | 38.4 ± 14.0 | 48.4 ± 23.8 |
| n-Eicosanoic acid | C20H40O2 | 312 | 0.4 1 | 14.8 1 | 3.0 1 | |
| n-Docosanoic acid | C22H44O2 | 340 | 132.0 2 | 131.9 ± 44.9 | 25.8 ± 9.9 | 18.8 ± 1.3 |
| Tricosanoic acid | C23H46O2 | 354 | 11.8 2 | |||
| n-Tetracosanoic acid | C24H48O2 | 368 | 75.5 ± 46.0 | 81.3 2 | 11.6 2 | 27.2 ± 2.9 |
| n-Hexacosanoic acid | C26H52O2 | 396 | 60.3 ± 14.0 | 42.5 1 | 5.2 1 | 43.8 ± 19.4 |
| Total Alkanoic Acids | 449 ± 184 | 1129 ± 128 | 207 ± 50 | 272 ± 31 | ||
| Iso-Alkanoic Acids | ||||||
| Iso-heptadecanoic acid | C17H34O2 | 270 | 24.5 2 | |||
| Monoacylglycerides | ||||||
| (±)-2,3-Dihydroxypropyl hexadecanoate (C16) | C19H38O4 | 330 | 9.2 2 | 19.1 1 | 37.1 2 | |
| Monostearin (C18:1) | C21H42O4 | 358 | 16.1 2 | 134.4 ± 26.1 | 97.0 ± 19.14 | 64.9 ± 8.9 |
| Linolein, 1-mono (C18:1) | C21H38O4 | 354 | 20.6 ± 9.5 | 31.9 1 | 9.3 1 | |
| 2-Monolinolenin (C18:2) | C21H36O4 | 352 | 6.4 1 | 74.3 ± 16.0 | 9.7 1 | |
| Total Monoacylglycerides | 52 ± 26 | 260 ± 19 | 134 ± 46 | 84 ± 2 | ||
| Carbohydrates | ||||||
| D-Glucose | C6H12O6 | 180 | 16.2 2 | 90.1 ± 32.9 | 37.8 ± 16.6 | 46.7 ± 14.7 |
| Mannose | C6H12O6 | 180 | 16.9 ± 7.0 | 212 ± 78.4 | 78.4 ± 25.3 | 46.0 ± 14.0 |
| Sucrose | C12H22O11 | 342 | 7.7 2 | 19.6 1 | 19.5 2 | 33.0 ± 11.3 |
| Trehalose | C12H22O11 | 342 | 3895 ± 712 1 | 2107 ± 217 2 | 1684 ± 191 2 | 1768 ± 157 2 |
| Total Carbohydrates | 3936 ± 703 | 2429 ± 256 | 18120 ± 229 | 1894 ± 192 | ||
| Steroids and Terpenoids | ||||||
| Aromadendrene | C15H24 | 204 | 0.5 1 | 125 2 | 13.8 1 | |
| γ-Elemene | C15H24 | 204 | 183.0 ± 81.1 | 13.5 1 | ||
| Ledol | C15H26O | 222 | 157.2 ± 32.9 | 9.3 1 | ||
| (-)-Globulol | C15H26O | 222 | 72.0 ± 7.5 | 17.1 2 | 11.8 1 | |
| trans-Farnesol | C15H26O | 222 | 753 ± 64.9 | 48.2 1 | ||
| Cholesterol | C27H46O | 386 | 11.4 2 | 98.8 ± 33.0 | 62.2 2 | 23.6 2 |
| Ergostrol | C28H44O | 396 | 36.8 2 | 94.7 1 | ||
| Campesterol | C28H48O | 400 | 32.9 2 | 117.3 ± 28.7 | 87.9 2 | 104.7 ± 0.3 |
| Stigmasterol | C29H48O | 412 | 56.0 2 | 243.1 ± 30.4 | 110.2 ± 17.4 | 138.9 ± 18.3 |
| ß-Sitosterol | C29H50O | 414 | 232.2 ± 93.8 | 243.2 ± 6.0 | 237.9 ± 46.1 | 142.9 ± 25.7 |
| Stigmastanol | C29H52O | 416 | 119.4 ± 34.2 | |||
| ß-Amyrin | C30H50O | 426 | 81.2 2 | 161.8 ± 1.8 | 98.9 ± 16.3 | |
| α-Amyrin | C30H50O | 426 | 14.7 2 | 66.1 ± 17.9 | ||
| Lupeol | C30H50O | 426 | 9.7 1 | 87.3 2 | 13.3 1 | 28.0 1 |
| Sitosterone | C29H48O | 412 | 13.3 1 | 73.6 2 | 17.0 1 | |
| Erthrodiol | C30H50O2 | 442 | 47.4 2 | 343.6 ± 100.3 | ||
| Oleanolic acid | C30H48O3 | 456 | 85.4 ± 52.5 | 642.1 ± 132.5 | 46.9 2 | |
| Total Steroids and Terpenoids | 622 ± 267 | 3461 ± 341 | 729 ± 80 | 616 ± 616 | ||
| Unidentified | 37.6 ± 3.4 | 126.2 ± 10.8 | 238.1 ± 99.6 | 67.7 2 | ||
| Total aliphatic lipids | 5007 ± 746 | 4729 ± 531 | 2550 ± 430 | 2664 ± 300 | ||
| Total solvent extracts | 5629 ± 740 | 8190 ± 857 | 3279 ± 508 | 3280 ± 394 | ||
| Compound | MF | MW | Forest | Eucalyptus | Cropland | Grazing Land |
|---|---|---|---|---|---|---|
| n-Alkanols3 | ||||||
| n-Hexadecanol | C16H34O | 242 | 7.4 1 | 20.2 | 66.4 2 | 75.3 2 |
| n-Octadecanol | C18H38O | 270 | 9.1 1 | 10.6 | 10.8 1 | |
| n-Nonadecanol | C19H40O | 284 | 40.5 ± 10.5 | 85.6 ± 12.9 | 80.7 2 | |
| n-Docosanol | C22H46O | 326 | 47.1 ± 5.5 | 121 ± 8 | 129 ± 36 | 552 ± 324 |
| n-Tetracosanol-1 | C24H50O | 354 | 13.0 1 | |||
| n-Hexacosanol | C26H54O | 382 | 14.6 1 | 14.3 1 | ||
| n-Octacosanol | C28H58O | 410 | 13.0 1 | |||
| Total n-Alkanols | 104 ± 21 | 265 ± 12 | 314 ± 21 | 627 ± 362 | ||
| n-Alkanoic Acids | ||||||
| n-Tetradecanoic acid | C14H28O2 | 228 | 128 ± 16 | 352 ± 20.2 | 540 ± 118 | 362.0 ± 92 |
| n-Hexadecanoic acid (C16:1) | C16H30O2 | 254 | 390 ± 50 | 1030 ± 40 | 969 ± 18 | 1569 ± 127 |
| n-Hexacosanoic acid | C16H32O2 | 256 | 579 ± 301 | 11.5 ± 49.1 | 651.2 ± 17.6 | 128 ± 70 |
| n-Heptadecanoic acid | C17H34O2 | 270 | 472 ± 65.8 | 514 ± 27.4 | 592 ± 82.7 | 62.3 ± 57.0 |
| n-Octadecanoic acid, cis-9 (C18:1) | C18H34O2 | 282 | 54.6 ± 12.2 | 106 ± 11 | 193 ± 38 | 321 ± 78 |
| n-Octadecanoic acid (C18:1) | C18H34O2 | 282 | 2710 ±790 | 6095 ± 118 | 6631 ± 437.1 | 8089 ± 655 |
| n-Octadecanoic acid | C18H36O2 | 284 | 187 ± 40.1 | 408 ± 5 | 621 ± 12.4 | 661 ± 53 |
| n-Nonadecanoic acid | C19H38O2 | 298 | 22.7 2 | 41.9 2 | ||
| n-Eicosanoic acid | C20H40O2 | 312 | 83.2 ± 13.4 | 213 ± 6.7 | 322.6 ± 74.7 | 347 ± 36 |
| n-Docosanoic acid | C22H44O2 | 340 | 93.3 ± 28.1 | 324 ± 61 | 292 ± 108 | 342 ± 76 |
| n-Triacosanoic acid | C23H46O2 | 354 | 47.8 ± 4.1 | 139 ± 9.3 | 204 ± 52.2 | 206 ± 26.8 |
| n-Tetracosanoic acid | C24H48O2 | 368 | 7.6 1 | 26.9 1 | 57.7 2 | |
| n-Hexacosanoic acid | C26H52O2 | 396 | 5.5 1 | 14.6 1 | ||
| n-Octacosanoic acid | C28H56O2 | 424 | 7.3 1 | 21.3 1 | 46.1 1 | |
| n-Triacontanoic acid | C30H60O2 | 302 | 11.0 1 | |||
| Total Alkanoic Acids | 4788 ± 1180 | 9998 ± 179 | 11063 ± 452 | 12157 ± 1173 | ||
| α-Alkanoic Acids | ||||||
| α-Hydroxy-Hexadecanoic acid | C16H32O3 | 272 | 66.7 2 | 105 ± 7.7 | 189 ± 37.4 | 383 ± 225 |
| α-Hydroxyoctadecanoic acid | C18H36O3 | 300 | 280 ± 30 | 398 ± 12 | 388 ± 64.4 | 88.0 1 |
| α-Hydroxydocosanoic acid | C22H44O3 | 356 | 17.4 2 | 12.5 1 | ||
| α-Hydroxytetracosanoic acid | C24H48O3 | 384 | 75.6 2 | |||
| α-hydroxypentacosanoic acid | C25H50O3 | 398 | 24.5 1 | |||
| Total α-Alkanoic Acids | 364 ± 60 | 515 ± 124 | 576 ± 100 | 572 ± 357 | ||
| α,ω-Alkanedioic Acids | ||||||
| α,ω-Butanedioic acid | C4H6O4 | 118 | 62.2 ± 20.8 | 123.7 ± 5.2 | 60.3 ± 14.3 | 178.7 ± 67.9 |
| α,ω-Nonanedioic acid | C9H16O4 | 188 | 39.0 ± 12.0 | 94.5 ± 4.2 | 109.2 ± 41.7 | 93.4 ± 24.4 |
| α,ω-Hexadecanedioic acid | C16H30O4 | 286 | 64.4 ± 9.1 | 110.6 ± 23.3 | 181.3 ± 42.3 | 248.4 ± 9.1 |
| α,ω-Octadec-9-enedioic acid (C18:1) | C18H32O4 | 312 | 120.0 ± 18.5 | 151.8 ± 52.7 | 93.7 2 | |
| α,ω-Eicosanedioc acid | C20H38O4 | 342 | 56.5 ± 11.7 | 161.4 ± 9.4 | 144.4 ± 15.6 | 157.8 ± 17.9 |
| Total α,ω-Alkanedioic Acids | 342 ± 49 | 642 ± 41 | 589 ± 125 | 678 ± 70 | ||
| ω-Hydroxyalkanoic Acids3 | ||||||
| ω-Hydroxyhexadecanoic acid | C16H32O3 | 272 | 97.5 ± 27.7 | 81.4 2 | 137.5 2 | 124.6 2 |
| ω-Hydroxyoctadecanoic acid | C18H34O3 | 298 | 41.9 ± 4.4 | 98.4 ± 13.7 | 103.9 ± 48.2 | 8.4 1 |
| ω-Hydroxydocosanoic acid | C22H44O3 | 358 | 83.1 ± 19.2 | 125 ± 28 | 106 ± 30 | 10.8 1 |
| ω-Hydroxyoctacosanoic acid | C28H56O3 | 440 | 136 ± 72 | 235 ± 35 | 478 ± 43 | 75.2 1 |
| ω-Hydroxytriacontanoic acid | C30H60O3 | 468 | 377 ± 70 | 565.2 ± 13.9 | 518.7 ± 69.9 | 78.7 1 |
| Total ω-Hydroxyalkanoic Acids | 736 ± 31 | 1105 ± 26 | 1344 ± 30 | 298 ± 141 | ||
| Mid-Chain Substituted Hydroxy Acids | ||||||
| 10,16-Dihydroxyhexadecanoic | C16H32O4 | 288 | 58.8 2 | 152.8 2 | 182.1 2 | 223.3 2 |
| 8-Hydroxyhexadecane-1,16-dioic acid | C16H30O5 | 302 | 67.4 ± 13.3 | 115 ± 26.6 | 123 ± 4.1 | |
| Dihydroxymethoxyoctadecanoic acid | C18H36O5 | 332 | 3.1 1 | 11.2 1 | ||
| 9,10,18-Trihydroxyoctadecanoic acid | C18H36O5 | 332 | 32.2 1 | 90.9 2 | 38.1 2 | |
| 9,10-Dihydroxyoctadecane-1,18-dioic acid | C19H38O4 | 330 | 65.5 ± 41.1 | 222 2 | 78.3 2 | 85.9 2 |
| Total Mid-Chain Substituted Hydroxy Acids | 227 ± 111 | 592 ± 122 | 422 ± 160 | 309 ± 80 | ||
| Monoacylglycerides | C19H38O4 | 330 | 238 ± 59 | 487 ± 128 | 432 ± 177 | |
| Benzyles and Phenols | ||||||
| p-Hydroxyacetophenone | C8H8O2 | 136 | 21.7 ± 4.1 | 53.6 ± 5.7 | 93.0 ± 17.0 | 55.3 2 |
| Vanillin | C8H8O3 | 152 | 4.7 1 | 38.0 2 | ||
| Acetovanillone | C8H10O3 | 154 | 51.4 2 | 122 ± 32.7 | 137 2 | 1080 ± 468 |
| 4-Hydroxybenzoic acid | C7H6O3 | 138 | 204 ± 27 | 609 ± 34 | 692 ± 51 | 916 ± 116 |
| Vanillic acid | C8H8O4 | 168 | 25.4 ± 10.7 | 74.3 ± 20.6 | 39.2 2 | 53.0 2 |
| Isovanillic acid | C8H8O4 | 168 | 60.1 ± 5.1 | 217 ± 33 | 254 ± 71 | 40.1 1 |
| p-Coumaric acid | C9H8O3 | 164 | 19.1 2 | 60.6 2 | 66.7 2 | 52.3 1 |
| Ferulic acid | C10H10O4 | 194 | 40.9 ± 5.1 | 96.8 ± 1.4 | 109 ± 29 | 67.5 2 |
| Total Benzyles and Phenols | 428 ± 53 | 1233 ± 57 | 1390 ± 200 | 2303 ± 493 | ||
| Compound | MF | MW | Forest * | Eucalyptus | Cropland | Grazing Land |
|---|---|---|---|---|---|---|
| Organophosphates | ||||||
| Triphenyl phosphate (mg g−1) | C18H15O4P | 326 | 1.5 ± 0.2 | 4.5 ± 0.1 | 5.6 ± 0.3 | 6.8 ± 0.6 |
| Hydroxy Benzene Products | ||||||
| p-Hydroxyacetophenone | C8H8O2 | 136 | 198 | 145 1 | nb | 483 2 |
| Benzoic acid, 4-methoxy-/p-Anisic acid | C8H8O3 | 152 | 524 | 233 1 | 301 1 | 537 2 |
| m-Hydroxy benzoic acid | C7H6O3 | 138 | 271 | 509 ± 61 | 625 ± 2 | 241 1 |
| 3,5-dihydroxybenzoic acid | C7H6O4 | 154 | 2991 | 665 ± 119 | 469 2 | 360 1 |
| 2,4,6-Trihydroxybenzoic acid | C7H6O5 | 170 | 181 | 442 ± 27 | 247 1 | 705 ± 145 |
| 3,4-Dihydroxybenzeneacetic acid | C8H8O4 | 168 | 592 | 801 ± 180 | 432 2 | 751 ± 214 |
| Total Hydroxy Benzene Products | 31218 | 2795 ± 413 | 2074 ± 417 | 3078 ± 910 | ||
| Protein and Polysaccharide Products | ||||||
| Benzoic acid | C7H6O2 | 122 | 626 | |||
| Butan-1,4-dioic acid/succinic acid | C4H6O4 | 118 | 2584 | 420 2 | 642 ± 15 | 242 1 |
| 2-Butenedioic acid/fumaric acid | C4H4O4 | 116 | 126 | 400 2 | 840 ± 55 | 175 1 |
| Maleic acid | C4H4O4 | 116 | 1591 | 183 1 | 519 1 | 237 1 |
| 1H-pyrrole-1-carboxylic acid | C5H5NO2 | 111 | 902 | 532 ± 64 | 658 ± 62 | 196 1 |
| p-Hydroxybenzaldehyde | C7H6O2 | 122 | 997 | 365 2 | 213 1 | 203 1 |
| 4-Hydroxybenzeneacetic acid | C8H8O3 | 152 | 1449 | 837 ± 183 | 742 ± 74 | 554 ± 35 |
| 2-Buten-1,4-dioic acid/citraconic acid | C5H6O4 | 130 | 165 | 151 1 | 200 1 | |
| Penten-1,5-dioic acid/itaconic acid | C5H6O4 | 130 | 121 | 499 ± 44 | 488 2 | 375 2 |
| Hydroxybutan-1,4-dioic acid/Malic acid | C4H6O5 | 134 | 150 | 301 2 | 425 2 | 518 ± 19 |
| Total Protein and Polysaccharide Products | 8713 | 3686 | 4528 | 2700 | ||
| Lignin Monomers | ||||||
| Vanillin/vanillaldehyde | C8H8O3 | 152 | 564 | 555 ± 78 | 219 1 | 576 ± 36 |
| Vanillic alcohol | C8H10O3 | 154 | 188 | 292 2 | nd | nd |
| Acetovanillone | C8H10O3 | 154 | 573 | 744 ± 155 | 654 ± 35 | 587 2 |
| Vanillic acid | C8H8O4 | 168 | 1169 | 178 1 | 725 ± 110 | |
| Total Vanillyls | 2493 | 1768 ± 353 | 1598 ± 364 | 1163 ± 384 | ||
| Syringaldehyde | C9H10O4 | 182 | 272 | 177 1 | 252 1 | 389 1 |
| Acetosyringone | C10H12O4 | 196 | 491 | 576 ± 89 | 707 ± 52 | 1091 ± 221 |
| Syringic acid | C9H10O5 | 198 | 619 | 741 ± 157 | 617 ± 36 | 199 1 |
| Total Syringyls | 1382 | 1494 ± 70 | 1576 ± 339 | 1827 ± 691 | ||
| Cinnamyls | ||||||
| P-Coumaric acid | C9H8O3 | 164 | 299 | 556 ± 72 | 660 ± 62 | 253 2 |
| Ferulic acid | C10H10O4 | 194 | 267 | 611 ± 83 | 656 ± 29 | 1015 ± 271 |
| Caffeic acid | C9H8O4 | 180 | 388 | 139 1 | 336 2 | |
| Sinapic acid | C11H12O5 | 284 | 4132 1 | 941 ± 295 2 | 932 ± 87 2 | 973 ± 145 2 |
| Total Cinnamyls | 5086 | 2247 ± 339 | 2249 ± 91 | 2577 ± 179 | ||
| Total Benzyles and Phenols | 48892 | 11991 ± 1394 | 12025 ± 1693 | 11344 ± 3471 | ||
| Dicarboxylic Acids | ||||||
| Adipic acid | C6H10O4 | 146 | 439 | 603 2 | 863 2 | 167 1 |
| Pimelic acid | C7H12O4 | 160 | 303 | 303 2 | 402 2 | 254 1 |
| Suberic acid | C8H14O4 | 174 | 818 | 708 ± 165 | 706 ± 108 | 760 2 |
| Azelaic acid | C9H16O4 | 118 | 1120 | 893 ± 214 | 672 ± 53 | 451 2 |
| Sebacic acid | C10H18O4 | 202 | 294 | 147 1 | 217 1 | |
| Brassilic acid | C11H20O4 | 216 | 292 | 527 ± 66 | 657 ± 44 | 790 ± 114 |
| Dodecanedioic acid | C12H22O4 | 230 | 1802 | 753 ± 176 | 469 2 | 1357 ± 240 |
| Total Dicarboxylic Acids | 5069 | 3933 ± 995 | 3769 ± 362 | 3995 ± 911 | ||
| Identified Total CuO Products | 53961 | 15924 ± 1700 | 15794 ± 1278 | 15340 ± 4044 | ||
| Unidentified | 8475 | 5425 ± 507 | 5638 ± 258 | 6250 ± 1147 | ||
| Total CuO Products | 62436 | 21349 ± 507 | 21432 ± 258 | 21590 ± 1147 | ||
References
- Guo, L.B.; Gifford, R.M. Soil carbon stocks and land use change: A meta analysis. Glob. Change Biol. 2002, 8, 345–360. [Google Scholar] [CrossRef]
- Shiferaw, B.; Holden, S. Soil erosion and smallholders’ conservation decisions in the highlands of Ethiopia. World Dev. 1999, 27, 739–752. [Google Scholar] [CrossRef]
- Awulachew, S.B.; McCartney, M.; Steenhuis, T.S.; Ahmed, A.A. A Review of Hydrology, Sediment and Water Resource Use in the Blue Nile Basin; International Water Management Institute (IWMI): Colombo, Sri Lanka, 2009. [Google Scholar]
- Lemenih, M.; Kassa, H. Re-Greening Ethiopia: History, Challenges and Lessons. Forests 2014, 5, 1896–1909. [Google Scholar] [CrossRef]
- Assefa, D.; Rewald, B.; Sandén, H.; Rosinger, C.; Abiyu, A.; Yitaferu, B.; Godbold, D.L. Deforestation and land use strongly effect soil organic carbon and nitrogen stock in Northwest Ethiopia. Catena 2017, 153, 89–99. [Google Scholar] [CrossRef]
- Bekele, M. Forest Plantations and Woodlots in Ethiopia; Regional Soil Conservation Unit, Swedish International Development Authority: Nairobi, Kenya, 2011. [Google Scholar]
- Desta, L.; Kassie, M.; Beninc, S.; Penderd, J. Land degradation in the highlands of Amhara region and strategies for sustainable land management. In Policies for Sustainable Land Management in the Highlands of Ethiopia; Jabbar, M.A., Pender, J., Ehiu, S.K., Eds.; International Livestock Research Institute: Addis Ababa, Ethiopia, 2000. [Google Scholar]
- Wassie, A.; Sterck, F.J.; Teketay, D.; Bongers, F. Tree regeneration in church forests of Ethiopia: Effects of microsites and management. Biotropica 2009, 41, 110–119. [Google Scholar] [CrossRef]
- Aerts, R.; Van Overtveld, K.; November, E.; Wassie, A.; Abiyu, A.; Demissew, S.; Daye, D.D.; Giday, K.; Haile, M.; Tewolde Berhan, S.; et al. Conservation of the Ethiopian church forests: Threats, opportunities and implications for their management. Sci. Total Environ. 2016, 551, 404–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohjonen, V.; Pukkala, T. Eucalyptus globulus In Ethiopian forestry. For. Ecol. Manag. 1990, 36, 19–31. [Google Scholar] [CrossRef]
- Solomon, D.; Fritzsche, F.; Lehmann, J.; Tekalign, M.; Zech, W. Soil organic matter dynamics in the subhumid agroecosystems of the Ethiopian highlands: Evidence from natural 13C abundance and particle-size fractionation. Soil Sci. Soc. Am. J. 2002, 66, 969–978. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, I.U.; Assefa, D.; Godbold, D.L. Land-Use Change Depletes Quantity and Quality of Soil Organic Matter Fractions in Ethiopian Highlands. Forests 2022, 13, 69. [Google Scholar] [CrossRef]
- Tolimir, M.; Kresović, B.; Životić, L.; Dragović, S.; Dragović, R.; Sredojević, Z.; Gajić, B. The conversion of forestland into agricultural land without appropriate measures to conserve SOM leads to the degradation of physical and rheological soil properties. Sci. Rep. 2020, 10, 13668. [Google Scholar] [CrossRef]
- Delelegn, Y.T.; Purahong, W.; Blazevic, A.; Yitaferu, B.; Wubet, T.; Göransson, H.; Godbold, D.L. Changes in land use alter soil quality and aggregate stability in the highlands of northern Ethiopia. Sci. Rep. 2017, 7, 13602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, M.W.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kogel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, A.; Simpson, M.J. Sources and composition of hydrolysable aliphatic lipids and phenols in soils from western Canada. Org. Geochem. 2006, 37, 385–407. [Google Scholar] [CrossRef]
- Otto, A.; Simpson, M.J. Evaluation of CuO oxidation parameters for determining the source and stage of lignin degradation in soil. Biogeochem 2006, 80, 121–142. [Google Scholar] [CrossRef]
- Mueller, K.E.; Polissar, P.J.; Oleksyn, J.; Freeman, K.H. Differentiating temperate tree species and their organs using lipid biomarkers in leaves, roots and soil. Org. Geochem. 2012, 52, 130–141. [Google Scholar] [CrossRef]
- Jansen, B.; Wiesenberg, G.B.L. Opportunities and limitations related to the application of plant-derived lipid molecular proxies in soil science. Soil 2017, 3, 211–234. [Google Scholar] [CrossRef] [Green Version]
- Rasse, D.P.; Rumpel, C.; Dignac, M.F. Is soil carbon mostly root carbon? Mechanisms for a specific stabilisation. Plant. Soil 2005, 269, 341–356. [Google Scholar] [CrossRef]
- Kögel-Knabner, I. The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter. Soil Biol. Biochem. 2002, 34, 139–162. [Google Scholar] [CrossRef]
- Feng, X.; Simpson, M.J. The distribution and degradation of biomarkers in Alberta grassland soil profiles. Org. Geochem. 2007, 38, 1558–1570. [Google Scholar] [CrossRef]
- Amelung, W.; Brodowski, S.; Sandhage-Hofmann, A.; Bol, R. Combining biomarker with stable isotope analyses for assessing the transformation and turnover of soil organic matter. In Advances in Agronomy; Sparks, D., Ed.; Academic Press: Cambridge, MA, USA, 2008; pp. 155–250. [Google Scholar]
- Angst, G.; Mueller, K.E.; Nierop, K.G.; Simpson, M.J. Plant-or microbial-derived? A review on the molecular composition of stabilized soil organic matter. Soil Biol. Biochem. 2021, 156, 108189. [Google Scholar] [CrossRef]
- Tareq, S.M.; Kitagawa, H.; Ohta, K. Lignin biomarker and isotopic records of paleovegetation and climate changes from Lake Erhai, southwest China, since 18.5kaBP. Quat. Int. 2011, 229, 47–56. [Google Scholar] [CrossRef]
- Feng, X.; Simpson, M.J. Molecular-level methods for monitoring soil organic matter responses to global climate change. J. Environ. Monit. 2011, 13, 1246–1254. [Google Scholar] [CrossRef]
- Poirier, N.; Sohi, S.P.; Gaunt, J.L.; Mahieu, N.; Randall, E.W.; Powlson, D.S.; Evershed, R.P. The chemical composition of measurable soil organic matter pools. Org. Geochem. 2005, 36, 1174–1189. [Google Scholar] [CrossRef]
- Kögel-Knabner, I. Analytical approaches for characterizing soil organic matter. Org. Geochem. 2000, 31, 609–625. [Google Scholar] [CrossRef]
- Otto, A.; Simpson, M.J. Analysis of soil organic matter biomarkers by sequential chemical degradation and gas chromatography–mass spectrometry. J. Sep. Sci. 2007, 30, 272–282. [Google Scholar] [CrossRef]
- Angst, G.; John, S.; Mueller, C.W.; Kögel-Knabner, I.; Rethemeyer, J. Tracing the sources and spatial distribution of organic carbon in subsoils using a multi-biomarker approach. Sci. Rep. 2016, 6, 29478. [Google Scholar] [CrossRef] [Green Version]
- Goñi, M.A.; Teixeira, M.J.; Perkey, D.W. Sources and distribution of organic matter in a river-dominated estuary (Winyah Bay, SC, USA). Estuar. Coast. Shelf Sci. 2003, 57, 1023–1048. [Google Scholar] [CrossRef]
- Otto, A.; Shunthirasingham, C.; Simpson, M.J. A comparison of plant and microbial biomarkers in grassland soils from the Prairie Ecozone of Canada. Org. Geochem. 2005, 36, 425–448. [Google Scholar] [CrossRef]
- Thevenot, M.; Dignac, M.F.; Rumpel, C. Fate of lignins in soils: A review. Soil Biol. Biochem. 2010, 42, 1200–1211. [Google Scholar] [CrossRef]
- Hedges, J.I.; Mann, D.C. The characterization of plant tissues by their lignin oxidation products. Geochim. Cosmochim. Acta 1979, 43, 1803–1807. [Google Scholar] [CrossRef]
- Tareq, S.M.; Tanaka, N.; Ohta, K. Biomarker signature in tropical wetland: Lignin phenol vegetation index (LPVI) and its implications for reconstructing the paleoenvironment. Sci. Total Environ. 2004, 324, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 4, 439–473. [Google Scholar] [CrossRef] [Green Version]
- WRB. World Reference Base for Soil Resources 2014, International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; Food and Agriculture Organization of the United Nations: Rome, Italy, 2014. [Google Scholar]
- Assefa, D. Carbon Dynamics of Different Land Use Systems in NW Ethiopian. Ph.D. Thesis, University of Natural Resources and Life Sciences, Vienna, Austria, 2017. [Google Scholar]
- Feng, X.; Simpson, M.J. Temperature responses of individual soil organic matter components. J. Geophys. Res. Biogeosci. 2008, 113. [Google Scholar] [CrossRef] [Green Version]
- Pautler, B.G.; Austin, J.; Otto, A.; Stewart, K.; Lamoureux, S.F.; Simpson, M.J. Biomarker assessment of organic matter sources and degradation in Canadian High Arctic littoral sediments. Biogeochem. 2010, 100, 75–87. [Google Scholar] [CrossRef]
- Assefa, D.; Rewald, B.; Sandén, H.; Godbold, D.L. Fine root dynamics in afromontane forest and adjacent land uses in the northwest ethiopian highlands. Forests 2017, 8, 249. [Google Scholar] [CrossRef] [Green Version]
- Hamer, U.; Rumpel, C.; Dignac, M.-F. Cutin and suberin biomarkers as tracers for the turnover of shoot and root derived organic matter along a chronosequence of Ecuadorian pasture soils. Eur. J. Soil Sci. 2012, 63, 808–819. [Google Scholar] [CrossRef]
- Godbold, D.L.; Hoosbeek, M.R.; Lukac, M.; Cotrufo, M.F.; Janssens, I.A.; Ceulemans, R.; Polle, A.; Velthorst, E.J.; Scarascia-Mugnozza, G.; De Angelis, P.; et al. Mycorrhizal hyphal turnover as a dominant process for carbon input into soil organic matter. Plant Soil 2006, 281, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Berhongaray, G.; Cotrufo, F.M.; Janssens, I.A.; Ceulemans, R. Below-ground carbon inputs contribute more than above-ground inputs to soil carbon accrual in a bioenergy poplar plantation. Plant Soil 2019, 434, 363–378. [Google Scholar] [CrossRef] [Green Version]
- Mendez-Millan, M.; Dignac, M.-F.; Rumpel, C.; Rasse, D.P.; Bardoux, G.; Derenne, S. Contribution of maize root derived C to soil organic carbon throughout an agricultural soil profile assessed by compound specific 13 C analysis. Org. Geochem. 2012, 42, 1502–1511. [Google Scholar] [CrossRef]
- Quenea, K.; Mathieu, J.; Derenne, S. Soil lipids from accelerated solvent extraction: Influence of temperature and solvent on extract composition. Org. Geochem. 2012, 44, 45–52. [Google Scholar] [CrossRef]
- Wallander, H.; Ekblad, A.; Godbold, D.L.; Johnson, D.; Bahr, A.; Baldrian, P.; Björk, R.G.; Kieliszewska-Rokicka, B.; Kjøller, R.; Kraigher, H. Evaluation of methods to estimate production, biomass and turnover of ectomycorrhizal mycelium in forests soils–A review. Soil Biol. Biochem. 2013, 57, 1034–1047. [Google Scholar] [CrossRef]
- Feng, X.; Nielsen, L.L.; Simpson, M.J. Responses of soil organic matter and microorganisms to freeze–thaw cycles. Soil Biol. Biochem. 2007, 39, 2027–2037. [Google Scholar] [CrossRef]
- Ahmed, I.U.; Mengistie, H.K.; Godbold, D.L.; Sandén, H. Soil moisture integrates the influence of land-use and season on soil microbial community composition in the Ethiopian highlands. App. Soil Ecol. 2019, 135, 85–90. [Google Scholar] [CrossRef]
- Elaissi, A.; Rouis, Z.; Salem, N.A.B.; Mabrouk, S.; Ben Salem, Y.; Salah, K.B.H.; Aouni, M.; Farhat, F.; Chemli, R.; Harzallah-Skhiri, F.; et al. Chemical composition of 8 eucalyptus species’ essential oils and the evaluation of their antibacterial, antifungal and antiviral activities. BMC Complement. Altern. Med. 2012, 12, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, A.; Wang, Y.; Liu, Y. Study on the chemical constituents of the essential oil of the leaves of Eucalyptus globulus Labill from China. Asian J. Tradit. Med. 2009, 4, 134–140. [Google Scholar]
- Jäger, S.; Trojan, H.; Kopp, T.; Laszczyk, M.N.; Scheffler, A. Pentacyclic Triterpene Distribution in Various Plants—Rich Sources for a New Group of Multi-Potent Plant Extracts. Molecules 2009, 14, 2016–2031. [Google Scholar] [CrossRef] [Green Version]
- Goñi, M.A.; Hedges, J.I. Lignin dimers: Structures, distribution, and potential geochemical applications. Geochim. Cosmochim. Acta 1992, 56, 4025–4043. [Google Scholar] [CrossRef]
- Goñi, M.A.; Montgomery, S. Alkaline CuO oxidation with a microwave digestion system: Lignin analyses of geochemical samples. Anal. Chem. 2000, 72, 3116–3121. [Google Scholar] [CrossRef]
- Li, F.; Pan, B.; Zhang, D.; Yang, X.; Li, H.; Liao, S.; Ghaffar, A.; Peng, H.; Xing, B. Organic matter source and degradation as revealed by molecular biomarkers in agricultural soils of Yuanyang terrace. Sci. Rep. 2015, 5, 11074. [Google Scholar] [CrossRef] [Green Version]
- Goñi, M.A.; Yunker, M.B.; Macdonald, R.W.; Eglinton, T.I. Distribution and sources of organic biomarkers in arctic sediments from the Mackenzie River and Beaufort Shelf. Mar. Chem. 2000, 71, 23–51. [Google Scholar] [CrossRef]
- Tesfay, H.M.; Konrad, H.; Lapin, K. Strategy for the Provision of Reproductive Material for Juniperus procera in Ethiopia; Report of the JUPRET Project (Juniperus procera—Development of a Concept for the Provision of Forest repRoductive Material in Ethiopia); Austrian Research Centre for Forests: Vienna, Austria, 2021; 35p, ISBN 978-3-903258-53-2. [Google Scholar]


| Extraction | Forest | Eucalyptus | Cropland | Grazing Land |
|---|---|---|---|---|
| Solvent | 5.6 ± 0.7 ab | 8.2 ± 0.9 b | 3.3 ± 0.5 a | 3.2 ± 0.4 a |
| Base hydrolysis | 30.0 ± 3.6 a | 82.0 ± 1.0 b | 97.4 ± 4.4 bc | 111.3 ± 8.2 c |
| CuO oxidation | 36.0 (33.2–38.7) | 21.3 ± 2.1 a | 21.4 ± 1.0 a | 21.4 ± 4.9 a |
| Compound 1 | Forest | Eucalyptus | Cropland | Grazing Land |
|---|---|---|---|---|
| n-Alkanols (C14–C30) | 324 ± 171 a | 520 ± 212 a | 250 ± 86 a | 356 ± 103 a |
| n-Alkanes (C17–C31) | 222 ± 71 ab | 391 ± 78 a | 139 ± 66 ab | 58 ± 15 b |
| n-Alkanoic acids (C9–C26) | 448 ± 184 b | 1129 ± 128 a | 207 ± 49 b | 272 ± 31 b |
| Sum | 995 ± 288 ab | 2040 ± 400 b | 596 ± 198 a | 686 ± 120 a |
| Sum < C20 | 172 ± 55 a | 1269 ± 221 b | 205 ± 60 a | 205 ± 59 a |
| Sum ≥ C20 | 823 ± 236 a | 771 ± 180 a | 391 ± 140 a | 482 ± 81 a |
| <C20:≥C20 ratio (s) | 0.21 ± 0.01 a | 1.69 ± 0.09 c | 0.55 ± 0.07 b | 0.43 ± 0.10 ab |
| Monoacylglycerides (C19–C21) | 52 ± 26 a | 260 ± 19 b | 134 ± 46 ab | 84 ± 2 a |
| Sugars | 3936 ± 703 a | 2429 ± 256 ab | 1820 ± 229 b | 1894 ± 192 b |
| Steroids and Terpenoids | 622 ± 267 a | 3461 ± 341 b | 729 ± 80 a | 616 ± 99 a |
| Compound | Forest | Eucalyptus | Cropland | Grazing Land |
|---|---|---|---|---|
| µg g−1 C | ||||
| n-Alkanols (C16–C28) 1 | 104 ± 21 a | 265 ± 12 a | 314 ± 21 a | 627 ± 362 a |
| n-Alkanoic acids (C14–C30) 2 | 4788 ± 1180 a | 9998 ± 179 b | 11063 ± 452 b | 12157 ± 1173 b |
| Branched alkanoic acids (iso-C16) | 11 a | 38 a | 16 a | nd |
| α-Alkanoic acids (C16–C25) 3 | 364 ± 60 a | 515 ± 124 a | 576 ± 100 a | 571 ± 357 a |
| Sum <C20 | 4901 ± 1113 a | 9799 ± 107 b | 10789 ± 357 b | 11664 ± 1317 b |
| Sum ≥C20 | 309 ± 43 a | 873 ± 79 bc | 994 ± 237 bc | 1616 ± 447 c |
| <C20:≥C20 ratio (bh) | 15.9 ± 3.4 a | 11.4 ± 1.0 a | 11.9 ± 2.2 a | 7.9 ± 1.2 a |
| α,ω-Alkanedioic acids (C4–C20) 3 | 342 ± 49 a | 642 ± 41 a | 589 ± 125 a | 678 ± 69 a |
| ω-Hydroxyalkanoic acids (C16–C30) 3 | 736 ± 31 a | 1105 ± 26 b | 1344 ± 30 b | 298 ± 141 c |
| Mid-chain substituted hydroxy acids | 227 ± 111 a | 592 ± 122 a | 422 ± 160 a | 309 ± 80 a |
| Monoacylglycerides (C19) | 238 ± 59 | 487 ± 128 | 432 ± 177 | nd |
| Benzyles and phenols | 428 ± 53 | 1233 ± 57 | 1390 ± 200 | 2303 ± 493 |
| mg g−1 C | ||||
| Organophosphates | 1.5 ± 0.2 a | 4.5 ± 0.1 b | 5.6 ± 0.3 bc | 6.8 ± 0.6 c |
| Suberin and Cutin Monomers | Forest | Eucalyptus | Cropland | Grazing Land |
|---|---|---|---|---|
| Suberin 1 | 653 ± 56 a | 1087 ± 63 b | 1247 ± 15 b | 322 ± 137 c |
| Suberin 2 | 792 ± 42 a | 1267 ± 27 b | 1488 ± 36 b | 456 ± 129 a |
| Cutin 1 | 126 ± 40 a | 268 ± 96 a | 305 ± 90 a | 223 ± 17 a |
| Cutin 2 | 158 ± 71 a | 359 ± 88 a | 344 ± 109 a | 223 ± 17 a |
| Suberin/Cutin ratio 1 | 6.1 ± 1.4 a | 6.3 ± 3.1 a | 5.3 ± 2.2 a | 1.0 ± 0.4 a |
| Suberin/Cutin ratio 2 | 7.1 ± 2.5 a | 3.9 ± 0.9 a | 6.1 ± 2.8 a | 1.5 ± 0.6 a |
| Index | Forest | Eucalyptus | Cropland | Grazing Land |
|---|---|---|---|---|
| S/V | 0.6 (0.5–0.7) | 1.1 (0.7–1.4) | 1.0 (1.0–1.0) | 1.4 (1.2–1.6) |
| C/V | 0.3 (0.2–0.3) | 0.8 (0.5–1.0) | 0.9 (0.6–1.0) | 1.3 (0.9–1.9) |
| V:S:C | 4:2:1 | 1:1:1 | 1:1:1 | 1:1:1 |
| LPVI | 70 (50–90) | 992 (301–1674) | 880 (548–1087) | 2080 (1323–2941) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Assefa, D.; Mentler, A.; Sandén, H.; Rewald, B.; Godbold, D.L. The Biological Origins of Soil Organic Matter in Different Land-Uses in the Highlands of Ethiopia. Forests 2022, 13, 560. https://doi.org/10.3390/f13040560
Assefa D, Mentler A, Sandén H, Rewald B, Godbold DL. The Biological Origins of Soil Organic Matter in Different Land-Uses in the Highlands of Ethiopia. Forests. 2022; 13(4):560. https://doi.org/10.3390/f13040560
Chicago/Turabian StyleAssefa, Dessie, Axel Mentler, Hans Sandén, Boris Rewald, and Douglas L. Godbold. 2022. "The Biological Origins of Soil Organic Matter in Different Land-Uses in the Highlands of Ethiopia" Forests 13, no. 4: 560. https://doi.org/10.3390/f13040560