1. Introduction
In recent years, the interest in microalgal cultivation has noticeably increased among researchers due to its nutritional values; however, their cultivation only started actively over the last three decades [
1,
2]. Japan was the first country to embark on microalgae cultivation in the 1960s, with Chlorella as the first microalgae cultivated for commercial application [
2,
3]. Microalgae have been reputed as one of the best sources in developing various products (biofuel, fodder additives, polysaccharides, dyes, food additives, unsaturated fatty acids, and antioxidants) used in various industries [
4]. Furthermore, microalgae contribution significantly impacts industries like food and pharmaceuticals, cosmetics, biofertilisers, nutraceutical, bioplastic, and aquaculture feed [
5,
6,
7]. In addition, some microalgae are viable because of their high lipid content for biofuel production [
8,
9] and are utilised as nutritional supplements or as a source of food due to their important nutritional ingredients [
10,
11].
Good and healthy soil is represented by sufficient air, water, minerals, and organic materials to promote and sustain plants. Animal and plant tissues in sediments and soils form humic substances by biochemical transformation. Humus found either in aqueous soil extracts (SE) or dissolved naturally largely consists of biologically active fulvo-humic fractions.
Although microalgae biomass is widely used, applications on a commercial scale are still limited because of the low yield and high cost of microalgal products [
12]. For instance, in the open sea, and thus in aquaculture, microalgae have an essential dietary role to marine animals [
13]. Moreover, all marine invertebrates are microalgae-dependent for their entire lives and utilised as live feed [
14,
15].
Typically, microalgae can be discovered in both freshwater and marine habitats, usually as single-celled photosynthetic autotrophic organisms [
16]. They could produce complex compounds such as carbohydrates, protein, and lipids from simple elements in their surroundings. Microalgae require basic nutrients for growth, besides light, carbon dioxide, and carbon, from the air to produce energy [
6]. The cultivation of marine microalgae requires phosphate fertilisers, nitrate, and some micronutrients such as manganese (Mn), copper (Cu), cobalt (Co), and zinc (Zn). These elements in the growth media [
17] are essential for processes such as energy storage and photosynthesis [
18].
In the growth of microalgal biomass, the culture medium plays a significant role and crucial in microalgae cultivation. The culture medium components should satisfy the basic requirements for optimum energy supply [
19]. However, culture medium alone is not adequate for microalgal growth; most marine algae require certain organic soil extract growing factors. In addition, the source of vitamins, hormones, and other micronutrients found in soil extract further enhanced growth. Similarly, artificial medium could also augment the growth of open ocean and coastal microalgal species.
Few studies also suggest that natural “growth-promoting nutrients” (GPN), such as soil extract, could also enhance the growth of microalgae due to its composition of nitrogen, sulphur, phosphorus, and magnesium [
20,
21]. These components are essential in producing high-quality microalgae biomass [
22] as they promote faster cell division, besides increasing cell density and the growth rate of microalgae [
23]. A previous study showed that soil extract contains the microelements beneficial for microalgae cultivation, and it has been proven that soil extract can establish rapid reproduction and growth in certain microalgae [
2].
The significant growth of industry and urban development leads to the increased use of heavy metals, causing serious environmental problems to the soil and the environment. Because algae play a role as primary producers in most aquatic ecosystems, estimating the heavy metal effects on the growth and photosynthesis of algae is vital in assessing the ecological impact of heavy metals [
24]. Heavy metals are highly abundant in nature, with Cu and Zn functioning as essential elements to microalgae and plants, while chromium (Cr), cadmium (Cd), and lead (Pb) are scarcely beneficial biologically [
25]. Cu is a key component in the electron transport chains and a co-enzyme factor that plays a significant role in plant photosynthesis and respiration. Meanwhile, Zn is a basic component of various enzymes involved in photosynthesis and metabolism, such as carbon anhydride, acidic phosphatase, and alkaline phosphatase. Both Cu and Zn are vital in the promotion of plant growth factors. If lacking either element, the plant’s rate of growth and photosynthesis will be greatly reduced.
Despite their benefits at minute concentrations, both metals may become toxic at higher levels. Unlike Cu and Zn, the heavy metals Cr, Cd, and Pb are unnecessary for plant growth or respiration. However, the rapid industrial growth produces a great quantity of Cr, Cd, and Pb deposited into the environment, resulting in adverse effects on plants. Being the most toxic heavy metal to plants, the toxicity of Cr is form-dependent. Meanwhile, Cd, a potentially toxic metal that is very stable in the aquatic environment, can directly affect the physiological processes of algae. Similarly, Pb is easily accumulated by algae, becoming a toxic influence.
Microalgae are diverse organisms with rapid growth rates and productivity and could live under extreme environments because of their unicellular or simple multicell structure. Microalgae are commonly used as biosensors in environmental analyses and monitoring due to their high sensitivity and specificity in detecting heavy metal pollutants because they are the primary producers at the bottom of the aquatic food chain, first organisms affected by heavy metal pollution. Studies have shown that microalgae possess extracellular and intracellular mechanisms to resist metal toxicity, including the ability to absorb a certain amount of heavy metals. Heavy metals may enter algal cells by active transportation or endocytosis, with the help of chelating proteins such as phytochelatin and class II metallothioneins; thus, reducing heavy metal toxicity. These chelating proteins form complexes with heavy metals and transfer them to intracellular vacuoles. However, toxicity occurs when the amount of accumulated heavy metals exceeds a critical concentration. The binding of heavy metal ions to sulfhydryl groups in the protein cysteine and the disruption of protein structure or displacement of essential elements will eventually affect the physiological and biochemical processes of the microalgae.
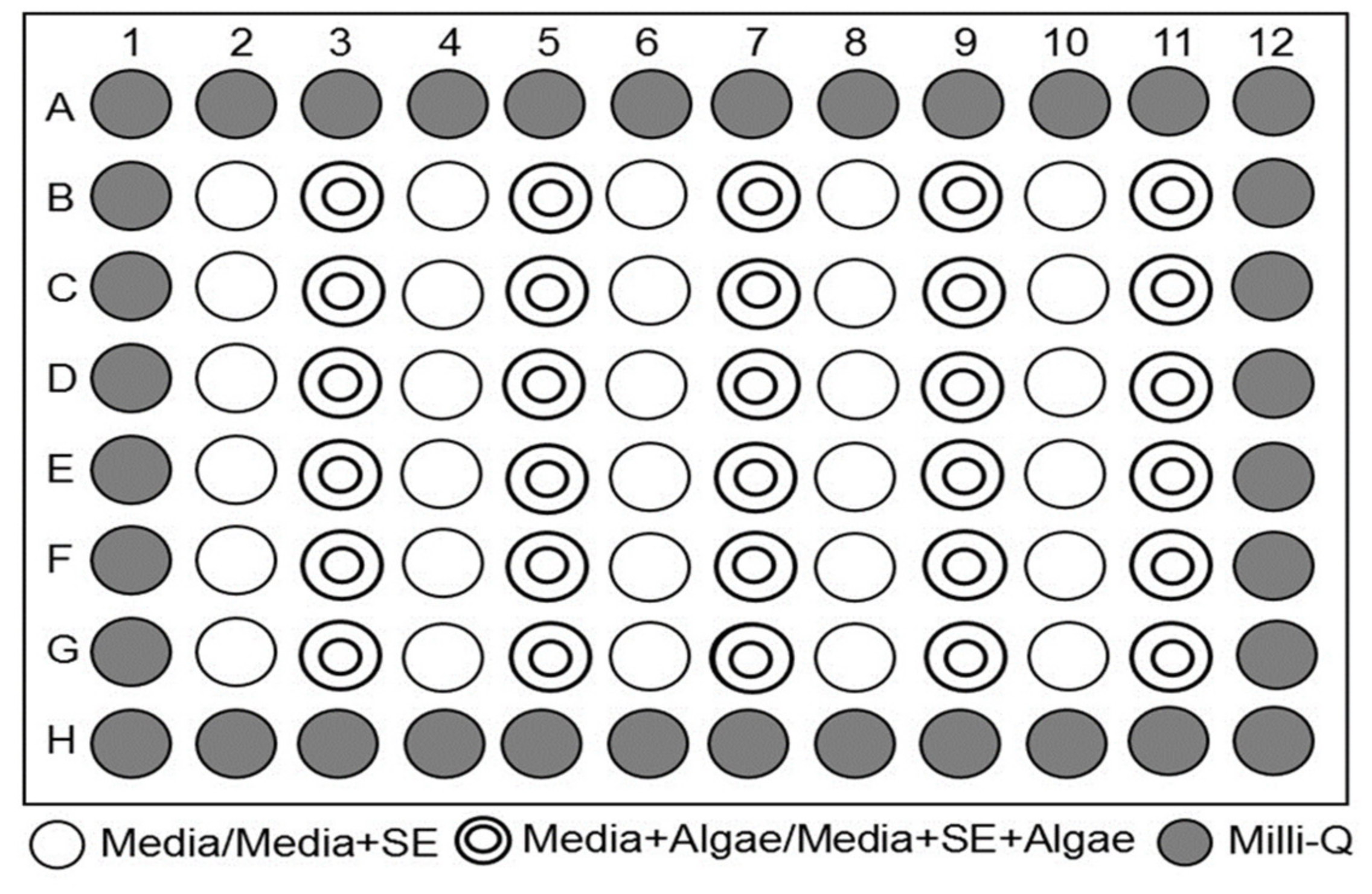
Though the positive effects of Growth-Promoting Agent (GPN) extracted from the soil are well known, very few studies have focussed on applying soil extracts from tropical rainforests and the effect of heavy metals on commercially exploitable, high-value microalgae. To the best of our knowledge, this research is the first attempt in investigating the effect of GNP from the virgin rainforest in a tropical climate. In this regard, this study aims to find the natural growth promoters and study the effectiveness of various soil extracts as well as heavy metals in soil on the targeted microalgae growth. In addition, a 96-well plate culturing method was adopted in this research to decrease the time and cost for a high throughput study of microalgae growth. This has subsequently contributed to the cost-effective production of microalgae with a cheaper medium.
3. Discussion
Microalgae cultivation is becoming more popular around the world as a result of the possible economic and commercial benefits [
26]. Using a unique microplate incubation approach, the current study attempted to assess the enhancing effect of soil extracts and heavy metal content from both RMFR and AHFR on the growth of specific microalgae species.
Soil texture, chemical characteristics, water holding capacity, and bulk density must be determined to understand the roles of soil science. The soil pH of RMFR in this investigation is 3.36, indicating acidic soil. The weathering and leaching processes within the soil could explain this phenomenon. According to the USDA guidelines, the pH levels are in the strongly acidic, moderately acidic, and minor acidic ranges, which differ from the pH levels (5.3 to 6.14) of soil in a previous study [
27]. Similarly, the soil pH level in this study is more acidic than those reported by Ichikogu [
28]. The P ion bound by the sand particles causes the strongly acidic soil in this investigation. The positive connection of sand and P analysis supports this hypothesis. It was discovered that increasing the amount of sand in the soil boosted the P content considerably.
This study determined the total C and N at 5.03 and 0.28 mg/kg for RMFR and at 5.23 mg/kg and 0.35 mg/kg for AHRF. The data are similar to the study by Amlin et al. [
29], where C and N contents were 2.76 and 0.28 mg/kg, respectively. However, the result is low compared to other studies. For example, in the plantation forest of
Khaya ivorensis, Heryati et al. [
30] reported the C and N contents of 12.24 and 1.29 g/kg, respectively. Likewise, the total P in this study (1.07 mg/kg) is lower than reported by Abdu et al. [
31], who studied the secondary forest and plantation forest in Regam (2.13 mg/kg), Durian (2.09 mg/kg), and Padang Besar (1.85 mg/kg). The findings revealed that the fully-managed secondary and plantation forests recover soil nutrients faster than natural forest regeneration.
From the results, Fe was found to be the most predominant trace metals at the soil sampling sites than other heavy metals such as Pb, Zn, and Cu. Fe concentrations are greatest among other heavy metals examined from both Raja Musa and Ayer Hitam locations, with 832.48 mg/kg in Raja Musa soil and 1905.48 mg/kg in Ayer Hitam soil, as shown in
Table 2. These results are lower than the established standard [
32]. The reason for this extreme value of Fe in soil might be due to the sampling procedure as the soils were collected from the depth. This was supported by another study [
23], explained that the highest concentration of Fe is found at 2–15 cm level despites the Fe content varies with the type and depth of the soil, ranging from 20,000 to 550,000 mg/kg [
33] (Xu et al., 2021). Fe is the fourth most prevalent element in the Earth’s crust; therefore, its high level could be attributed to its abundance [
34]. Fe is essential mineral elements for algae growth which promoting photosynthesis, respiration, nitrogen fixation, protein, and nucleic acid synthesis.
Heavy metals possess relatively high densities, and the proportional weight of the atom with an atomic number above 20 is significant. Certain heavy metals, such as Co, Cu, Fe, Mn, Mo, Ni, V, and Zn, are essential for organisms in minimum amounts. Excessive quantities of these metals, however, are harmful to organisms. Other heavy metals such as Pb, Cd, Hg, and As are not advantageous for organisms [
35]. The Pb values in the soil samples analysed are 9.15 and 506.16 mg/kg for Raja Musa and Ayer Hitam soil sample site, respectively (
Table 4). It can be summarised that Pb concentration level in soil between both sampling site was significantly different. According to Singh et al. [
36], the typical mean Pb concentration for surface soils worldwide averages 32 mg/kg and ranges from 10 to 67 mg/kg. However, the value of Pb obtained from Raja Musa soil in this study is lower than in previous studies [
37,
38], with 100 mg/kg suggested as the upper limit [
39]. Meanwhile, a study by Lenart and Wolny-Koładka [
40] indicated a higher Pb concentration than the Ayer Hitam soil sampling site. The availability of Pb in soil can also be affected by the soil type since Pb prefers to absorb on clay and oxides as well as form complex with organic matter. This finding might be the factor that cause Pb concentration was high in Ayer Hitam compared to Raja Musa that has sand-texture soil. Metal availability is influenced by soil conditions in various ways. Harter [
41] reported that soil pH is the most crucial component of metal availability in soil. As reported by Wang et al. [
42], the availability of Cd and Zn decreased with increasing soil pH. Heavy metals have also been significantly associated with soil physical attributes, such as moisture content and water holding capacity [
43].
In Raja Musa and Ayer Hitam soils, Zn concentrations ranged from 4.91 mg/kg to 121.83 mg/kg. The mean concentration of Zn for surface soils worldwide ranges from 5 to 10 mg/kg. The soil collected from RMFR showed the lowest Zn concentration in contrast to soil collected from AHFR. The zinc content found in this study is significantly lower than Ogundele et al. [
44]. The low levels of Zn in these forest sites are likely attributable to the soil’s limited exposure to farming activities in these areas, as well as the widespread use of pesticides and fertilisers.
Cu concentrations ranged from 0.70 mg/kg to 84.41 mg/kg. The soil sample collected from Raja Musa regions had the lowest concentration of Cu, whereas Ayer Hitam soil had the greatest concentration. It was found that the mean concentration of Ayer Hitam soil was above permissible limit set by the Department of Environment in soil (the DOE’s permissible limit of Cu in soils is 13.80 mg/kg) [
45]. The Cu concentration in this Raja Musa soil sample was generally lower than the allowed levels in soil defined in [
46]. All algae have a micronutrient copper requirement for growth and reproduction; however, excess copper could be highly toxic to most algae. For example, Cu plays a very important role in photosynthesis, in a protein involved in photosynthetic electron transport, and serves as a co-factor for several enzymes in the green alga. Cu is required at sufficient concentrations to merit its addition to artificial media.
The finding of this study shows that some of the heavy metal present in the soil such as arsenic, cadmium, chromium, nickel, mercury, and silver are less in concentration which is not determined. This might be attributed to the fact that, undisturbed nature of the forest soil which has been prevented from experiencing illicit felling and lopping of the soil [
47].
According to the work in [
48], the addition of 0.5–2.5 mg/L of mercury inhibited the photosynthetic activity of C. vulgaris by 23.3–71.1 percent. C. vulgaris photosynthetic activity was inhibited by 0.5–2.5 mg/L silver for 12 h, resulting in a 23.1–57.6 percent inhibition. Mercury interferes with chloride binding and harms the oxygen evolution complex [
49]. Mercury inhibits the metabolic fixation of CO2 and the uptake of the carbon source in the dark reaction. According to Singh et al. [
50], even at very low concentrations, mercury inhibits algae growth by affecting nutrient uptake and nitrogen metabolism. It inhibits a variety of enzymatic activities while also inducing oxidative stress, altering the antioxidative defence mechanism.
Over a 72 h exposure time, Ksiazyk et al. [
51] and Ribeiro et al. [
52] reported 50% inhibition of microalgae values of 1.6 and 0.03 mg/L for silver for Pseudokirchneriella subcapitata species, respectively. Silver penetrated the cell wall, causing damage to the cell membrane, membrane integrity loss, and cell lysis.
Lukavsky et al. [
53] found exposure to 2.5 mg/L copper for 12 h showed 44% inhibition of photosynthetic activity of
C. vulgaris. However, the toxicity of arsenic was mainly depended on the exposure time instead of its concentration in the solutions. Therefore, when arsenic concentration in the exposed medium was below 1000 μg/L, no matter how much arsenic was absorbed in the algae, it was able to promote the absorption of Arsenic during the first 10 days of exposure [
54] and after day 10, the toxicity of arsenic caused the algal cells to die gradually. Although the NPK decreased in the soil extract used (except for potassium in RMRF). The decreasing of heavy metal content in extract helps in promoting algae growth as compared in real soil.
NH
+4 and NO
3 might be used as N sources by most phytoplankton species [
55]. Based on our review, some researchers have discovered that for a given N source, macro- and microalgae have a higher photosynthetic or growth rate, as shown by different affinities for NH
+4 and NO
3 uptake, or changes in a wide range of physiological parameters related to growth response to different N sources. According to the work in [
56], NH
+4 is the preferred N source for
Chlamydomonas species and inhibit NO
3 absorption. As the content of both nutrients varies, these preferences could be a factor in competition [
57]. Unlike NO
3, NH
+4 is directly absorbed into amino acids by glutamate condensation, which is catalysed by glutamine synthetase [
56].
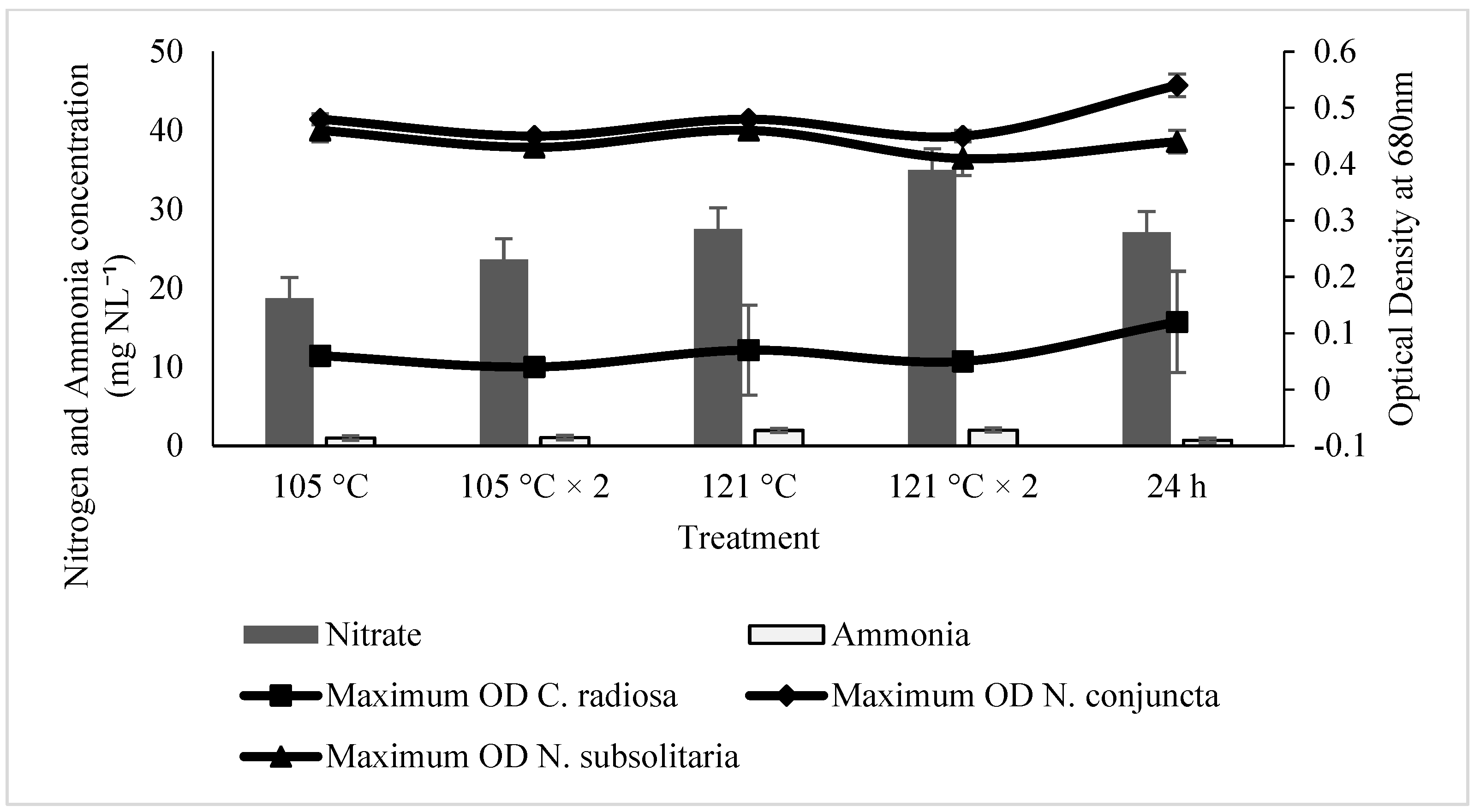
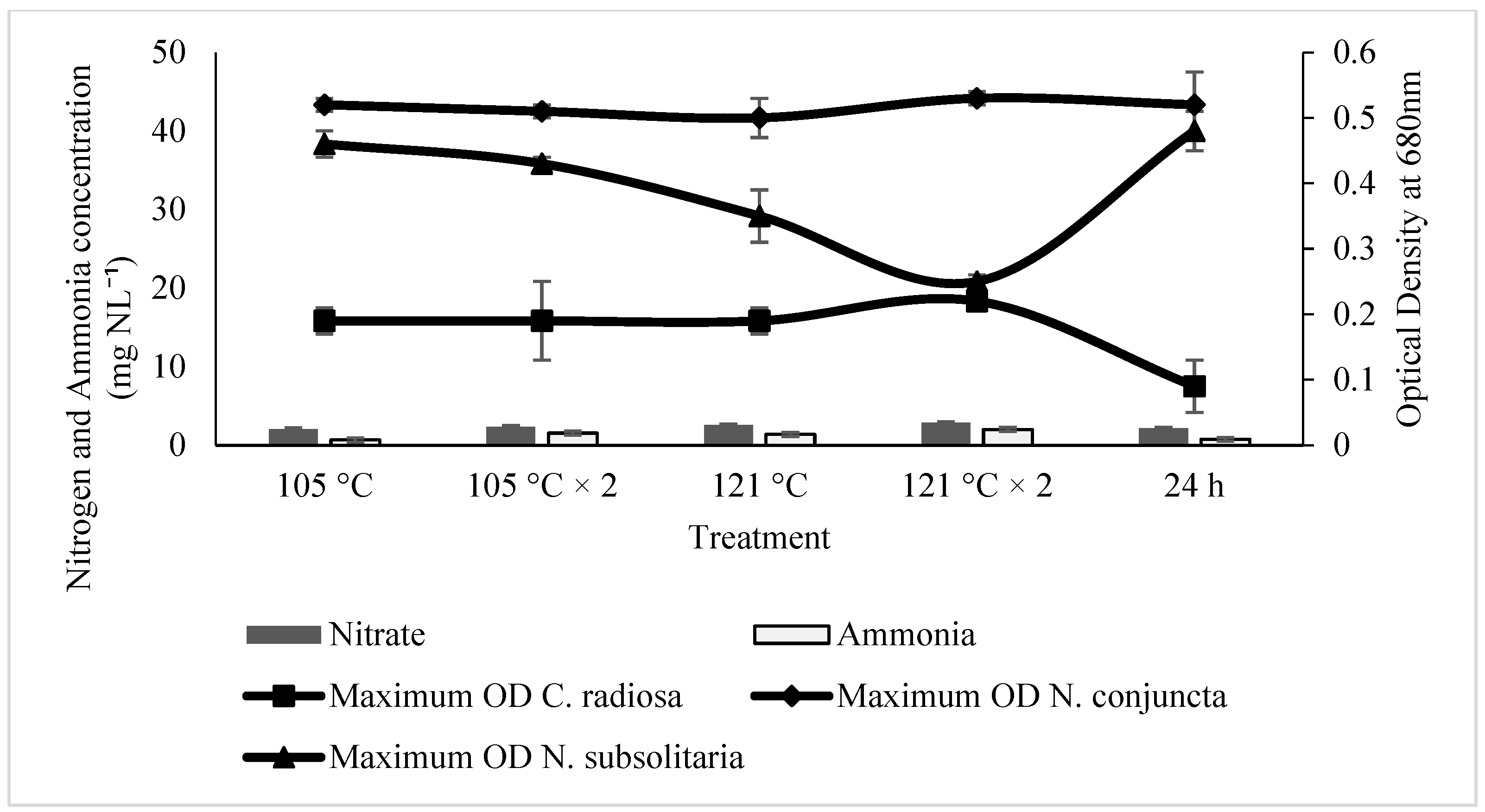
Microalgae prefer ammonium to convert it to nitrate or nitrite. Although ammonium is a reduced form of N, it can be digested directly into amino acids within the cells. On the other hand, nitrate or nitrite must first be reduced to ammonium before being used [
58]. However, in the present study, nitrate is more favourable for the growth of
C. radiosa,
N. conjuncta, and
N. subsolitaria cells, showing approximately the highest maximum OD and growth rate. Therefore, cell growth inhibition upon ammonium supplementation might indicate that it is toxic to the cell. However, the inhibitory level of ammonium on cell growth is different, depending on the microalgal species. Furthermore, in rare situations, ammonium oversaturation in the medium can drastically lower the pH by releasing H+ ions, limiting cell development and even triggering cell lysis [
59,
60]. Several studies reported cell growth inhibition at certain concentrations of ammonium. For example, cell growth suppression was seen in a marine microalga,
Dunaliella tertiolecta, at ammonium concentrations greater than 10 mM [
61]. Conversely, ammonium concentrations of 15 mM were found to limit the growth of a blue-green alga,
Arthrospira platensis [
62], whereas low levels (1.2 mM) were found to be lethal to several marine algae [
63]. This study demonstrated that nitrate is preferred for growth rather than ammonium, similar to previous studies. In comparison, many other microalgal species preferred nitrate over ammonium for growth, such as
D. tertiolecta,
Isochrysis galbana,
Neochloris oleoabundans,
Chlorella sorokiniana, and
Botryococcus braunii [
59,
61,
64,
65,
66].
This soil extract contains unidentified growth factors absent in other materials [
67]. Biomass growth and its chemical composition of microalgae mainly depend on the quality (source) and quantity (intensity) and C and N percentage in the growth media. The algal cell requires inorganic nutrients provided by the growth medium, including essential elements such as N, P, Fe, and silica (SiO
2) [
68]. The highest cell growth rate was merely found by the AHFR due to the ammonium, nitrogen, and various organic matters present from the SE. The species seemingly utilises both N and organic compound-modified SE through metabolism and consequently stimulating cell growth.
Soil extract plays a critical role in the enhanced biomass of
C. radiosa,
N. conjuncta, and
N. subsolitaria due to N, S, P, Ca, and Mg composition. These elements play a vital function in protein formation for microalgal growth. P and Ca are also essential in cell division by promoting faster cell division, increasing cell density, specific growth rate resulting in faster algal growth [
69].
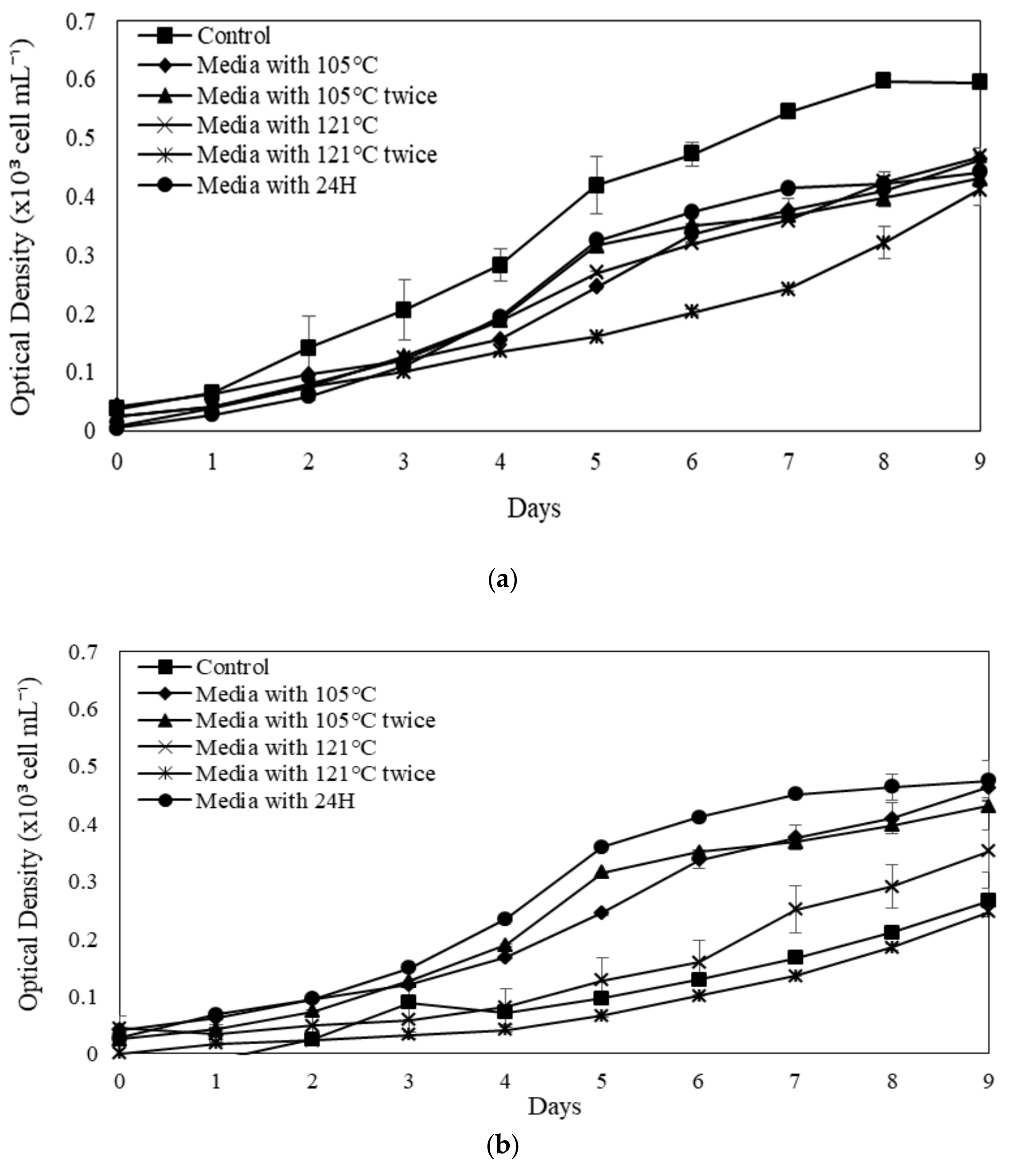
Due to the difference in organic matter content, the Ayer Hitam soil extract promoted microalgal development proportionately to the Raja Musa soil extract in the current investigation. The freshwater nature of the species renders it favourable growth in the AH SE. In some cases, decreasing the pH will consequently inhibit cell growth and even cause cell lysis. However, the development of several microalgae is similar under different soil extraction parameters and the control. Due to high quantities of nutrients combined with specialised extraction procedures, microalgae thrived well in varied extraction settings compared to the controls in some circumstances. Specific extraction conditions are also likely to decrease while precipitated essential minerals and vitamins exist in treated SE [
70].
The maximum OD observed in Ayer Hitam SE is higher than that of Raja Musa SE at different treatment methods for all microalgal species. Maximum OD at specific points has been observed specifically in
N. conjuncta and
N. subsolitaria when treated with Ayer Hitam SE. Several studies have reported using soil extracts for algae cultivation and growth enhancement [
71,
72]. Moreover, the production of microalgae biomass was reported to increase significantly due to its higher levels of N, P, and higher concentrations of micronutrients (Co, Mn, Zn, and Cu), which adversely affect the growth of microalgae, as studied by Lian et al. [
73]. In the present study, media with 121 °C, × 2 and 24 h of treatment method exhibited the maximum OD in Ayer Hitam SE in modified soil extracts. Marjakangas et al. [
74] stated that autoclaving at high temperatures for an extended time could kill bacteria and protozoa; thus, inhibiting the growth of microalgae. Moreover, a drastic increase in dissolved organic matters after autoclaving has been reported [
70,
75,
76].
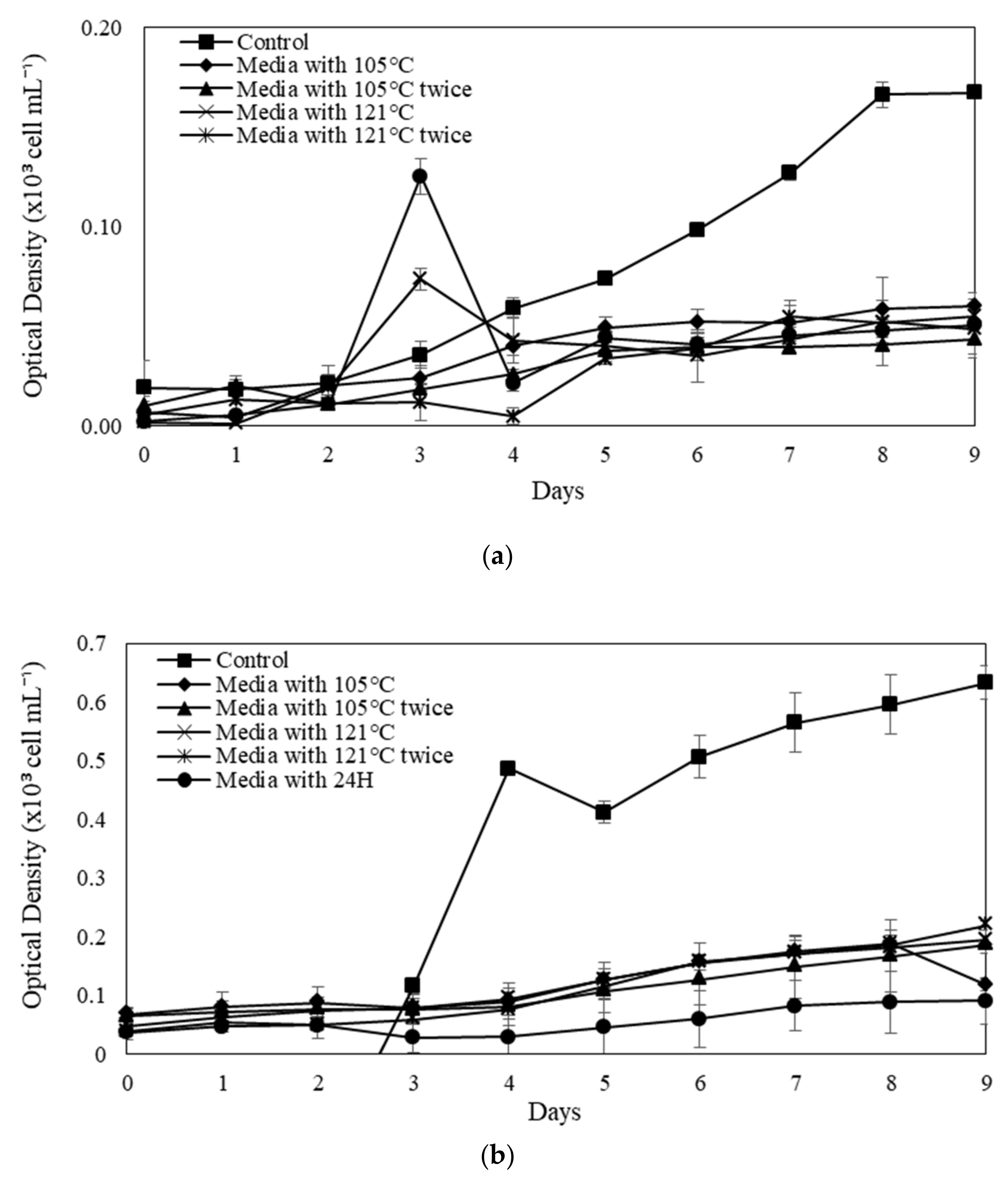
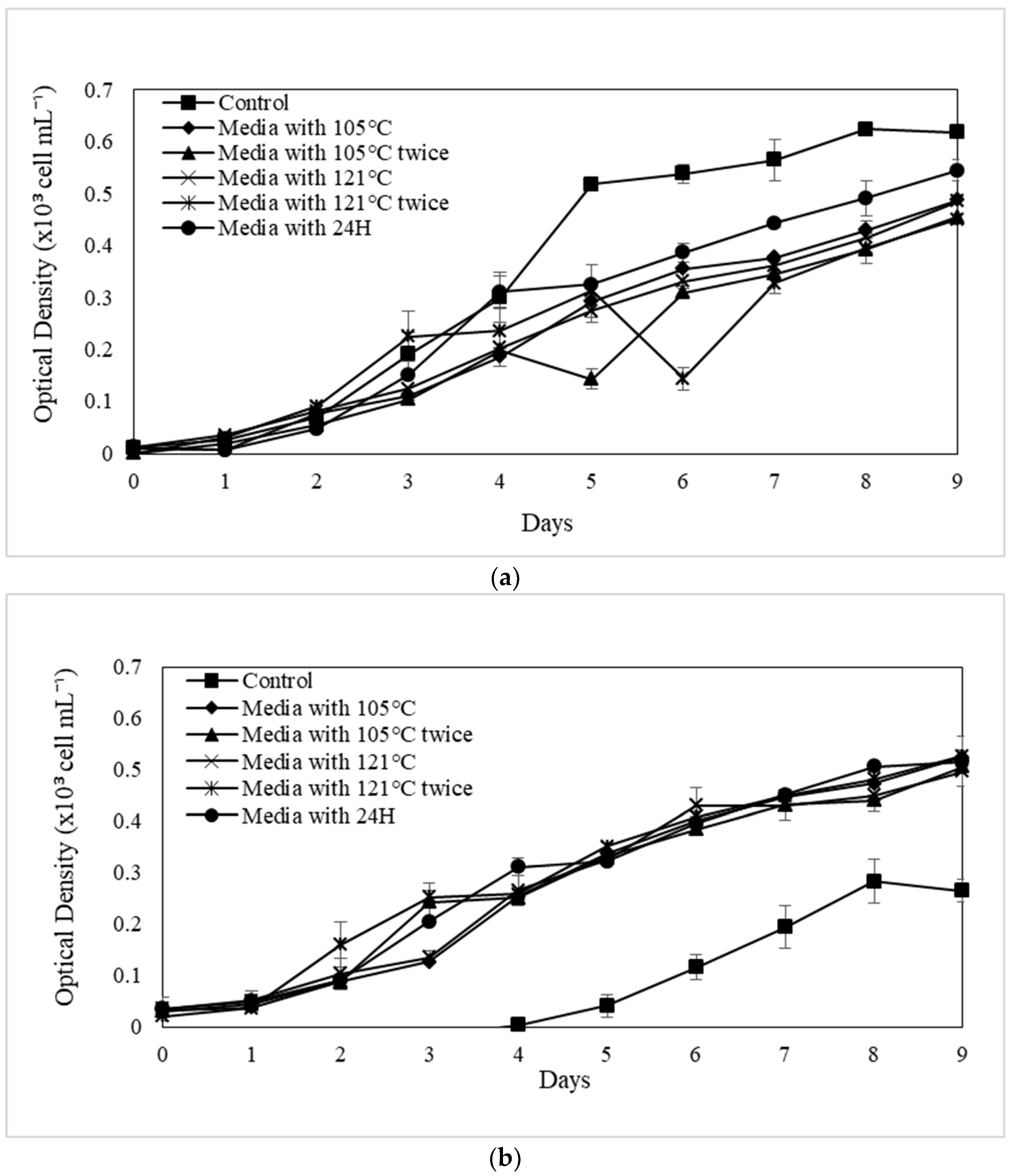
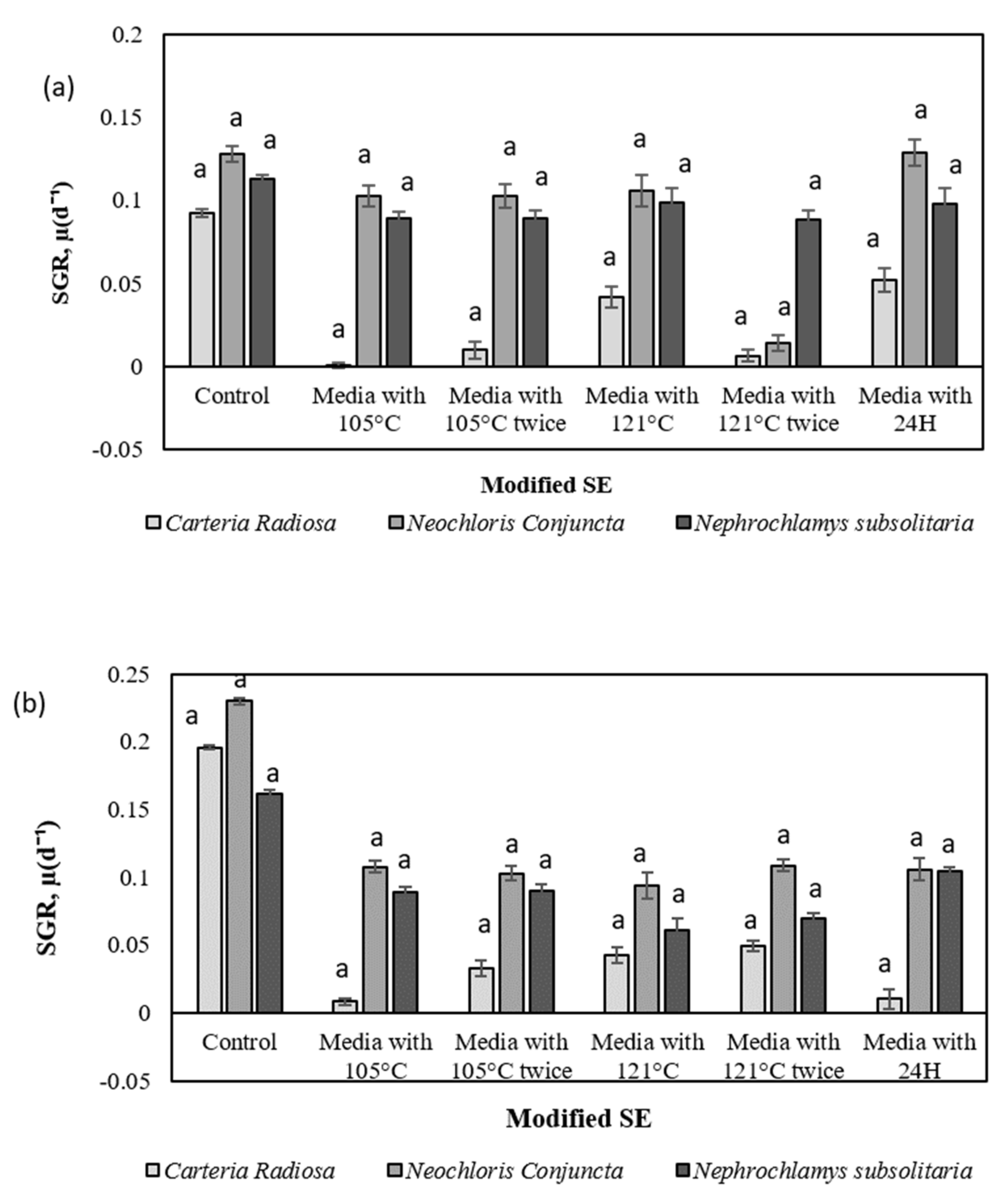
The SGR of microalgal species varies depending on the category of soil extracts, the procedure of augmentation or extraction, and the culture media utilised in the control experiment.
N. conjuncta SGR is in comparable to the control in modified Raja Musa and Ayer Hitam soil extracts (
Figure 6A,B). The significantly greater SGR implies that the extraction parameters and nutrients of further SE that have been modified are essential to the exponential increase. A higher SGR value in the control experiment by some species indicates species dependence and variability. This might be due to the basal medium containing N mainly in the form of NO
3-, which is reduced to ammonia during the cultivation period. These high levels of environmental ammonium inhibit cell growth, resulting in the imbalance of ammonia diffusion across the plasma membrane. At high concentrations, the tricarboxylic acid (TCA) cycle may be disturbed, disrupting cellular respiration. In this scenario, more support for microalgae production will be provided, but overall biomass concentrations will not rise over time. Furthermore, Chew et al. [
77] claimed that high microalgae concentrations could restrict growth rates due to population concentration. The microplate approach utilised in this work may induce certain growth constraints due to the low volume (microwells) and high microalgal biomass concentration on day 9. Namely, the small incubation capacity of the microplate wells may have slowed any growth.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}