


Effects of Tree Species on Moso Bamboo (Phyllostachys edulis (Carriere) J. Houzeau) Fine Root Morphology, Biomass, and Soil Properties in Bamboo–Broadleaf Mixed Forests

Abstract
:1. Introduction
2. Materials and Methods
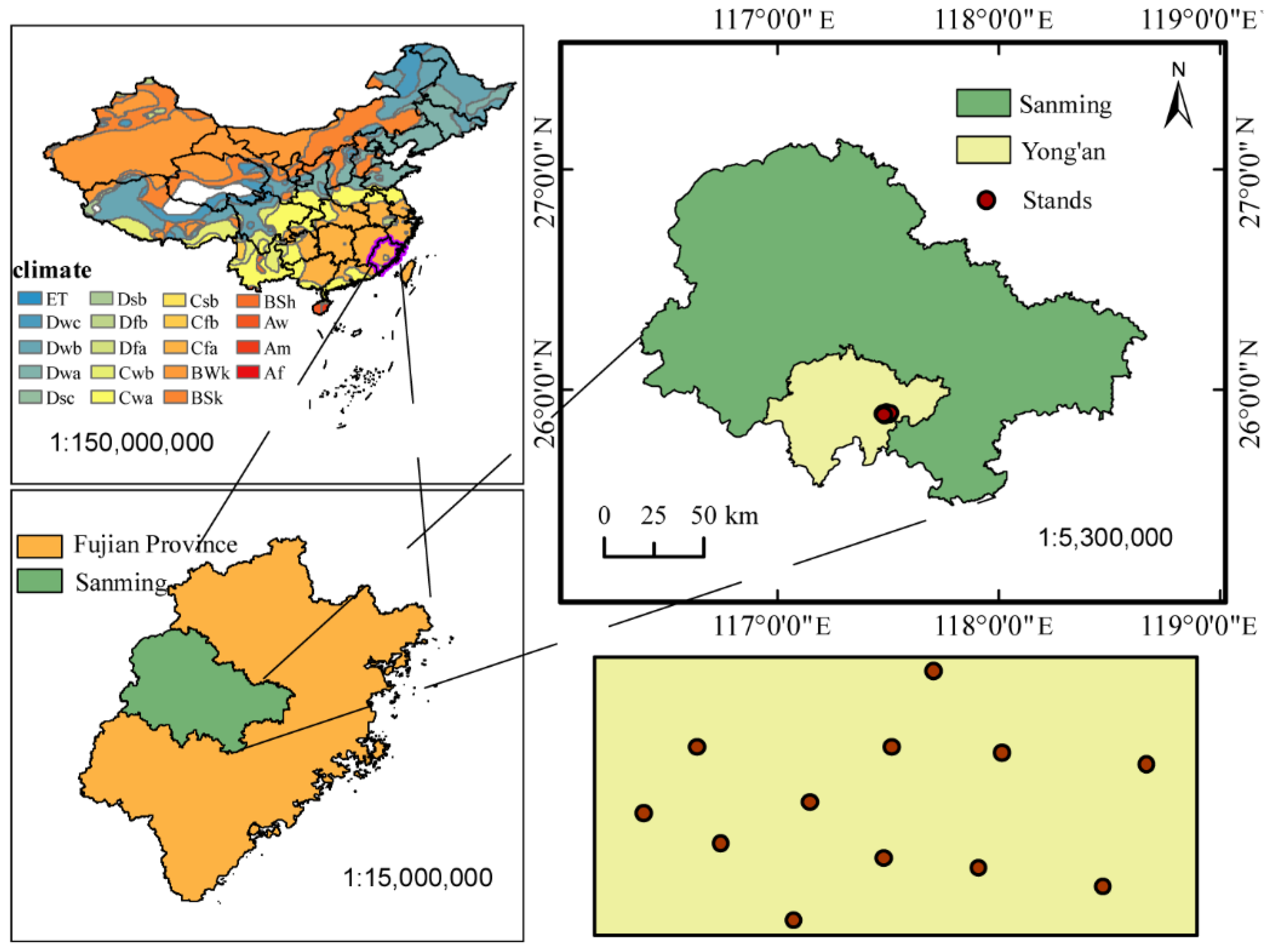
2.1. Site Description
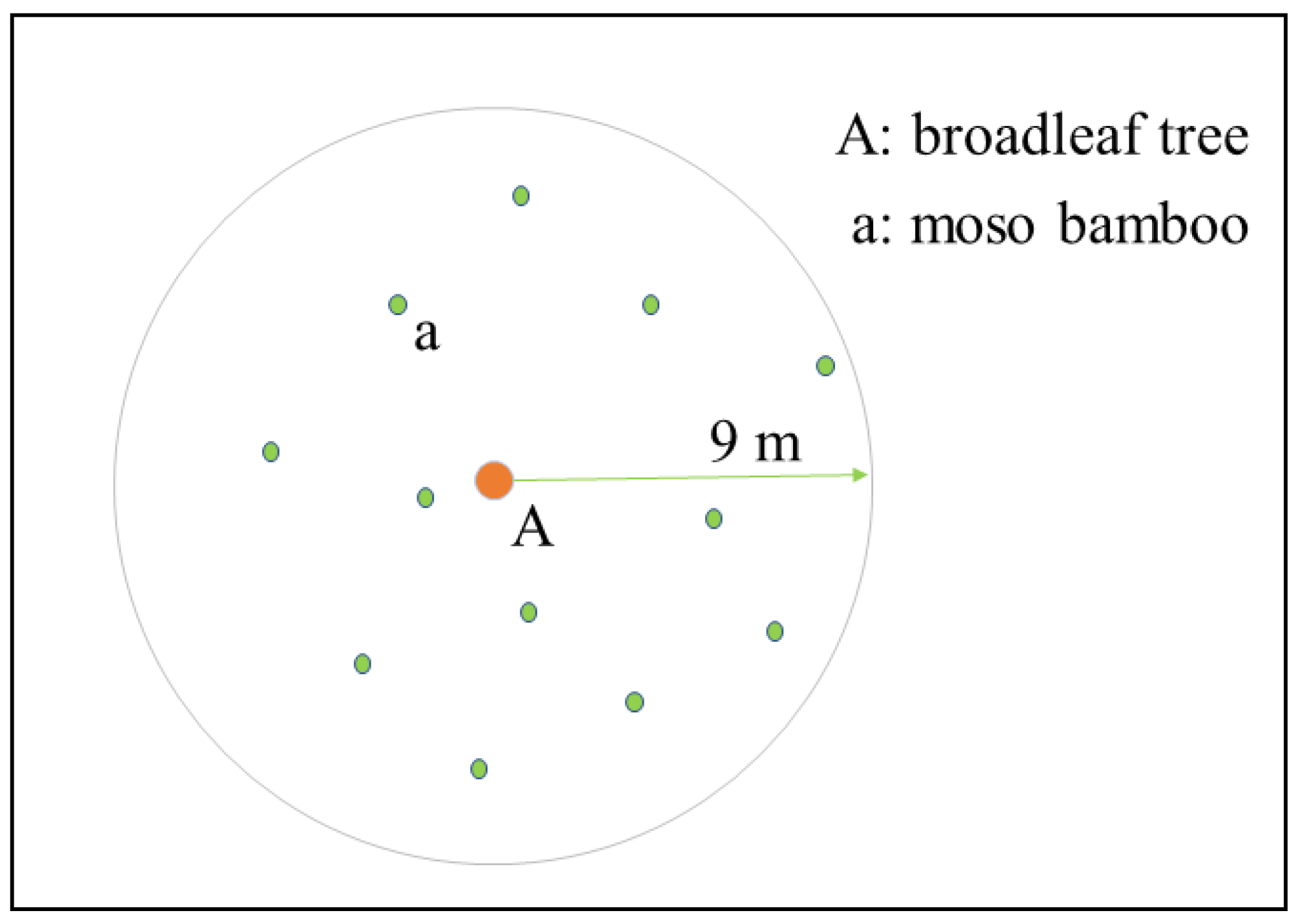
2.2. Experiment Design
2.3. Fine Root Sampling and Analysis
2.4. Soil Sampling and Analysis
2.5. Data Analysis
3. Results
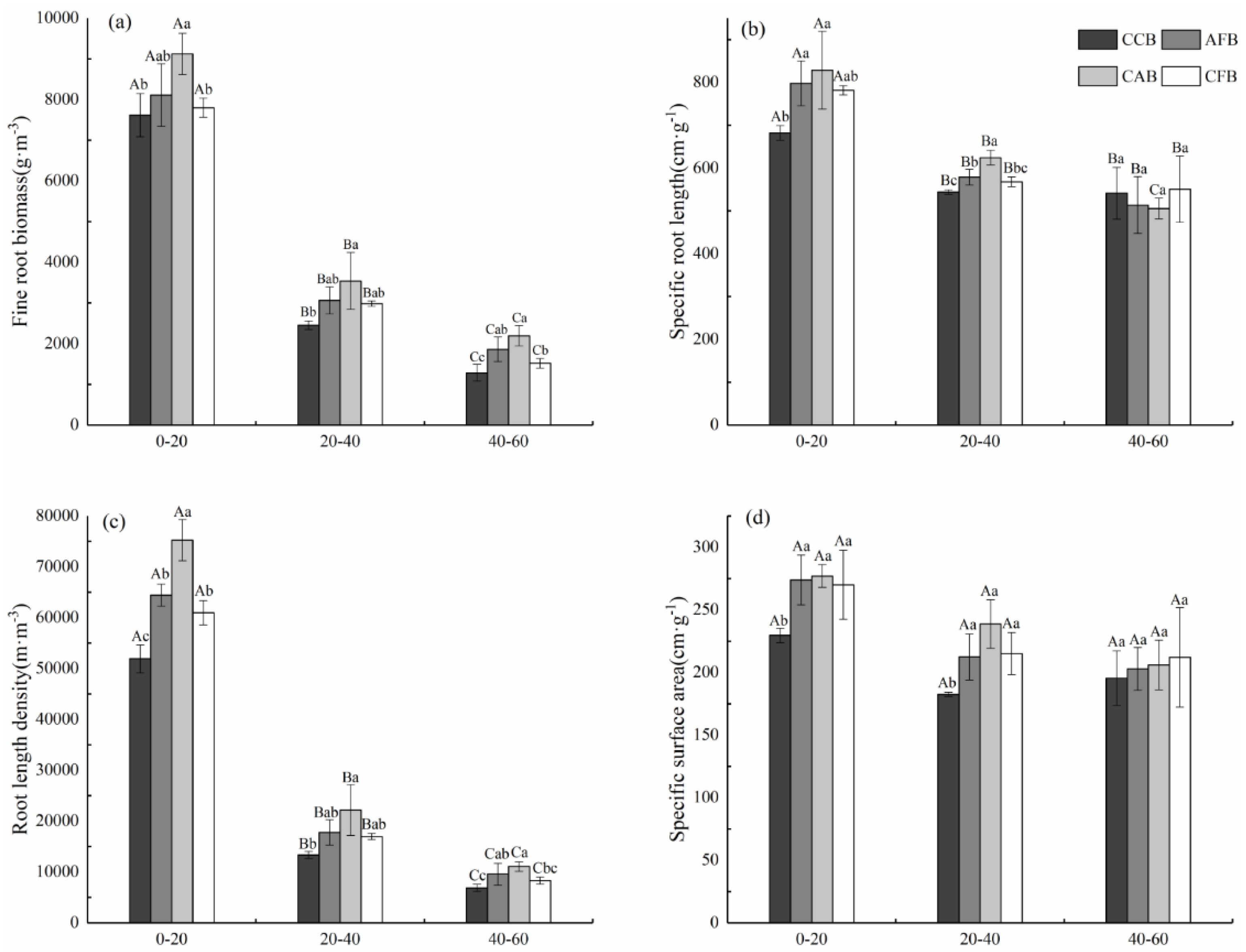
3.1. Fine Root Traits
3.2. Soil Properties
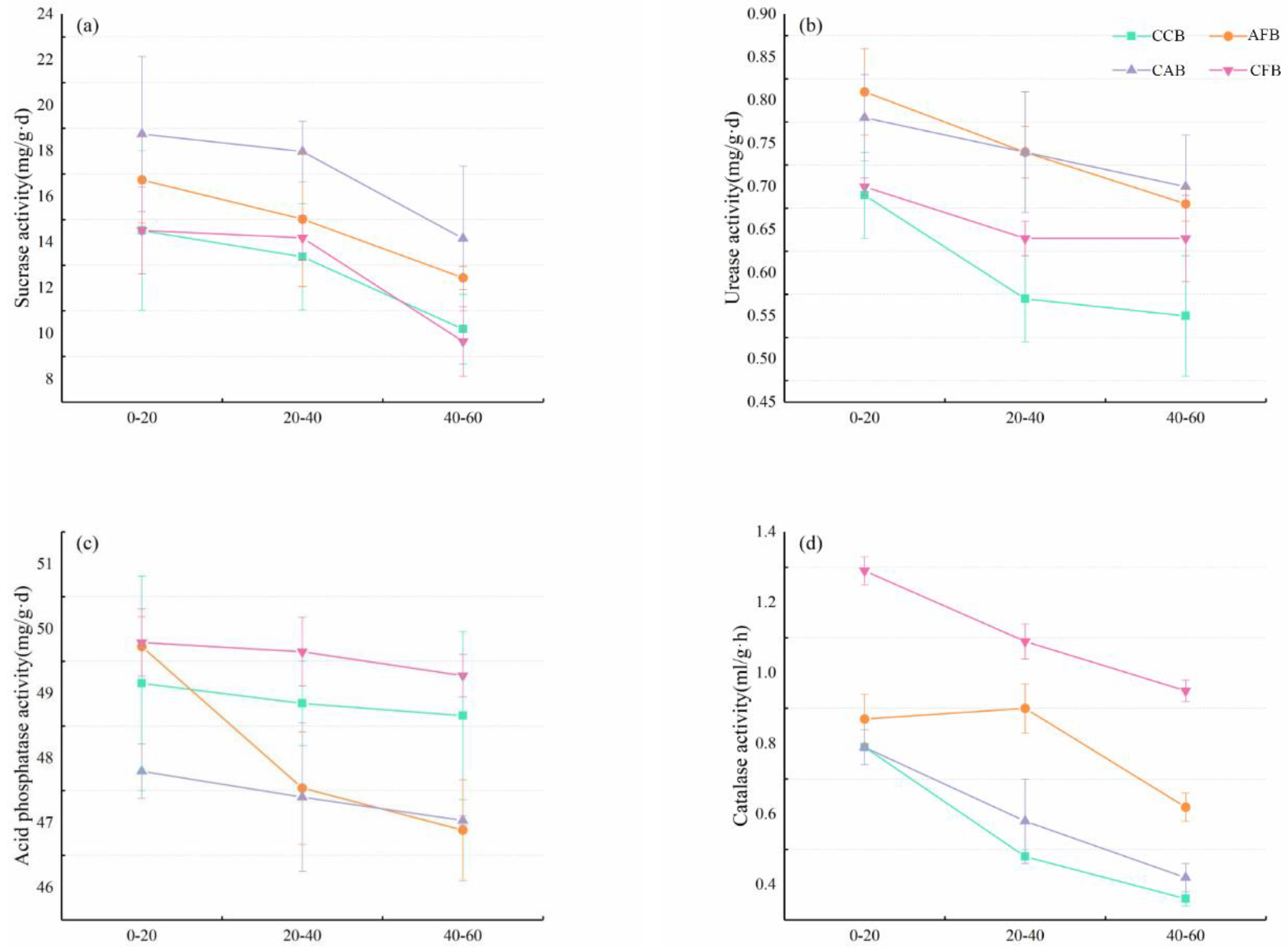
3.3. Soil Enzyme Activity
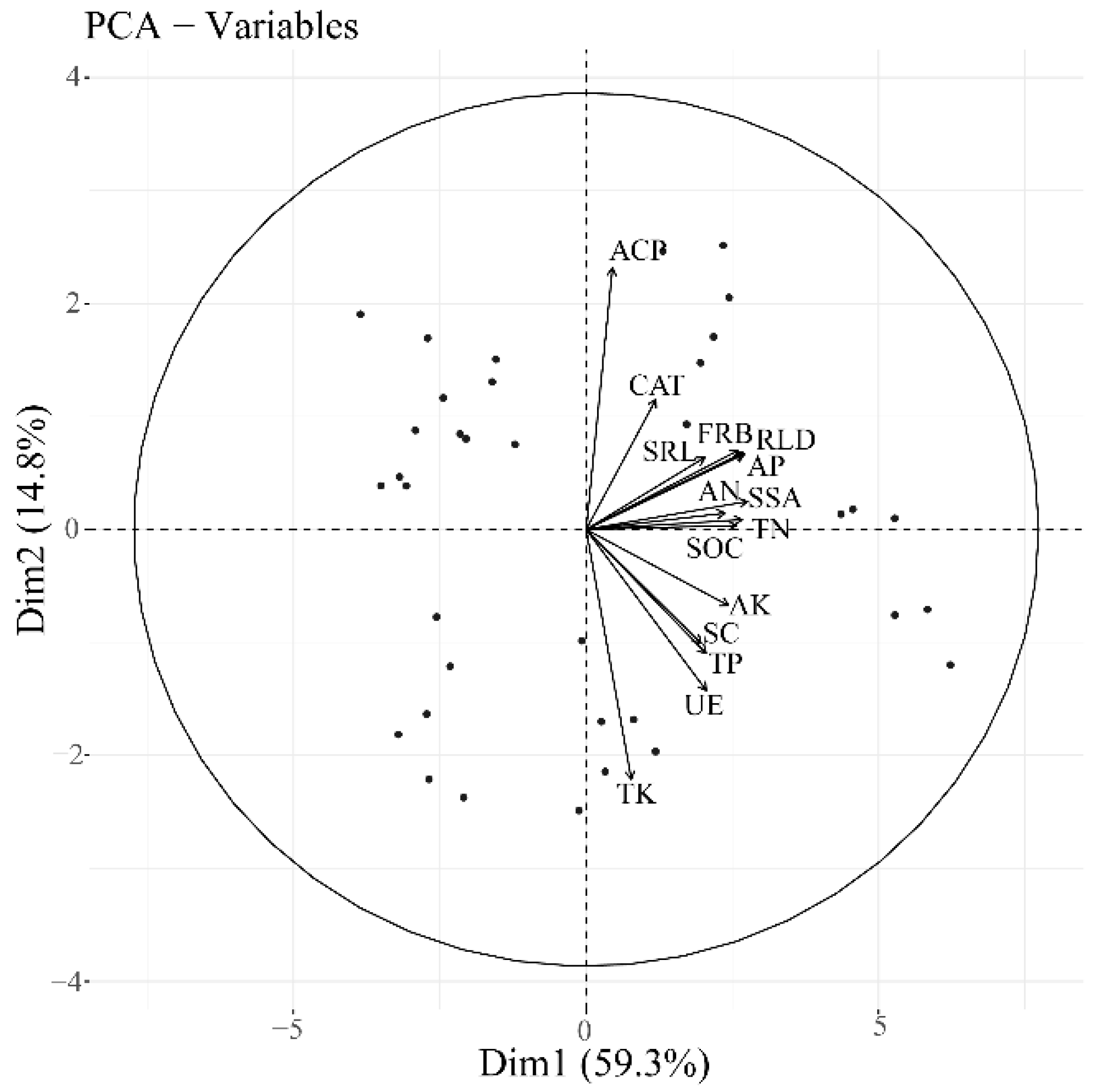
3.4. Associations between Stand Fine Root Traits and Soil Properties
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Río, M.; Pretzsch, H.; Alberdi, I.; Bielak, K.; Bravo, F.; Brunner, A.; Condés, S.; Ducey, M.J.; Fonseca, T.; von Lüpke, N.; et al. Characterization of mixed forests. In Dynamics, Silviculture and Management of Mixed Forests; Springer: Cham, Switzerland, 2018; pp. 27–71. [Google Scholar]
- Williams, L.J.; Paquette, A.; Cavender-Bares, J.; Messier, C.; Reich, P.B. Spatial complementarity in tree crowns explains overyielding in species mixtures. Nat. Ecol. Evol. 2017, 1, 1–7. [Google Scholar] [CrossRef]
- Jactel, H.; Bauhus, J.; Boberg, J.; Bonal, D.; Castagneyrol, B.; Gardiner, B.; Gonzalez-Olabarria, J.R.; Koricheva, J.; Meurisse, N.; Brockerhoff, E.G. Tree diversity drives forest stand resistance to natural disturbances. Curr. For. Rep. 2017, 3, 223–243. [Google Scholar] [CrossRef]
- Forrester, D.I. The spatial and temporal dynamics of species interactions in mixed-species forests: From pattern to process. For. Ecol. Manag. 2014, 312, 282–292. [Google Scholar] [CrossRef]
- Pretzsch, H.; Steckel, M.; Heym, M.; Biber, P.; Ammer, C.; Ehbrecht, M.; Bielak, K.; Bravo, F.; Ordóñez, C.; Collet, C.; et al. Stand growth and structure of mixed-species and monospecific stands of Scots pine (Pinus sylvestris L.) and oak (Q. robur L., Quercus petraea (Matt.) Liebl.) analysed along a productivity gradient through Europe. Eur. J. For. Res. 2020, 139, 349–367. [Google Scholar] [CrossRef] [Green Version]
- Primicia, I.; Camarero, J.J.; Imbert, J.B.; Castillo, F.J. Effects of thinning and canopy type on growth dynamics of Pinus sylvestris: Inter-annual variations and intra-annual interactions with microclimate. Eur. J. For. Res. 2013, 132, 121–135. [Google Scholar] [CrossRef]
- Sterck, F.J.; Duursma, R.A.; Pearcy, R.W.; Valladares, F.; Cieslak, M.; Weemstra, M. Plasticity influencing the light compensation point offsets the specialization for light niches across shrub species in a tropical forest understorey. J. Ecol. 2013, 101, 971–980. [Google Scholar] [CrossRef] [Green Version]
- Valverde-Barrantes, O.J.; Raich, J.W.; Russell, A.E. Fine-root mass, growth and nitrogen content for six tropical tree species. Plant Soil 2007, 290, 357–370. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Mommer, L.; de Vries, F.T. Going underground: Root traits as drivers of ecosystem processes. Trends Ecol. Evol. 2014, 29, 692–699. [Google Scholar] [CrossRef]
- Hallett, P.D.; Marin, M.; Bending, G.D.; George, T.S.; Collins, C.D.; Otten, W. Building soil sustainability from root–soil interface traits. Trends Plant Sci. 2022, 27, 688–698. [Google Scholar] [CrossRef]
- Casper, B.B.; Jackson, R.B. Plant competition underground. Annu. Rev. Ecol. Evol. Syst. 1997, 545–570. [Google Scholar] [CrossRef]
- de Kroon, H. How do roots interact? Science 2007, 318, 1562–1563. [Google Scholar] [CrossRef] [PubMed]
- Messier, C.; Coll, L.; Poitras-Larivière, A.; Bélanger, N.; Brisson, J. Resource and non-resource root competition effects of grasses on early-versus late-successional trees. J. Ecol. 2009, 97, 548–554. [Google Scholar] [CrossRef] [Green Version]
- Harper, J.L.; Jones, M.; Hamilton, N.R. The Evolution of Roots and The Problems of Analyzing Their Behavior, Plant Root Growth, An Ecological Perspective. Spec. Publ. Br. Ecol. Soc. 1991, 10, 3–22. [Google Scholar]
- Børja, I.; De Wit, H.A.; Steffenrem, A.; Majdi, H. Stand age and fine root biomass, distribution and morphology in a Norway spruce chronosequence in southeast Norway. Tree Physiol. 2008, 28, 773–784. [Google Scholar] [CrossRef]
- Persson, H.; Ahlström, K. Fine-root response to nitrogen supply in nitrogen manipulated Norway spruce catchment areas. For. Ecol. Manag. 2002, 168, 29–41. [Google Scholar] [CrossRef]
- Freschet, G.T.; Roumet, C.; Comas, L.H.; Weemstra, M.; Bengough, A.G.; Rewald, B.; Bardgett, R.D.; de Deyn, G.B.; Johnson, D.; Klimešová, J.; et al. Root traits as drivers of plant and ecosystem functioning: Current understanding, pitfalls and future research needs. New Phytol. 2021, 232, 1123–1158. [Google Scholar] [CrossRef]
- Bergmann, J.; Weigelt, A.; van der Plas, F.; Laughlin, D.C.; Kuyper, T.W.; Guerrero-Ramirez, N.; Valverde-Barrantes, O.J.; Bruelheide, H.; Freschet, G.T.; Iversen, C.M.; et al. The fungal collaboration gradient dominates the root economics space in plants. Sci. Adv. 2020, 6, eaba3756. [Google Scholar] [CrossRef]
- Zhang, X.; Dippold, M.A.; Kuzyakov, Y.; Razavi, B.S. Spatial pattern of enzyme activities depends on root exudate composition. Soil Biol. Biochem. 2019, 133, 83–93. [Google Scholar] [CrossRef]
- Ni, H.; Su, W.; Fan, S.; Chu, H. Effects of intensive management practices on rhizosphere soil properties, root growth, and nutrient uptake in Moso bamboo plantations in subtropical China. For. Ecol. Manag. 2021, 493, 119083. [Google Scholar] [CrossRef]
- Roumet, C.; Birouste, M.; Picon-Cochard, C.; Ghestem, M.; Osman, N.; Vrignon-Brenas, S.; Cao, K.-F.; Stokes, A. Root structure–function relationships in 74 species: Evidence of a root economics spectrum related to carbon economy. New Phytol. 2016, 210, 815–826. [Google Scholar] [CrossRef]
- Bahamondez, C.; Álvarez, O.; Itzelcoaut, M. Global Forest Resources Assessment 2010, Main Report; FAO: Rome, Italy, 2010. [Google Scholar]
- Jiang, Z.H. Bamboo and Rattan in the World; China Forest Publishing House: Beijng, China, 2007. [Google Scholar]
- Yang, C.; Ni, H.; Zhong, Z.; Zhang, X.; Bian, F. Changes in soil carbon pools and components induced by replacing secondary evergreen broadleaf forest with Moso bamboo plantations in subtropical China. Catena 2019, 180, 309–319. [Google Scholar] [CrossRef]
- Li, W.; Bai, Z.; Jin, C.; Zhang, X.; Guan, D.; Wang, A.; Yuan, F.; Wu, J. The influence of tree species on small scale spatial heterogeneity of soil respiration in a temperate mixed forest. Sci. Total Environ. 2017, 590, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Tan, R.; Yang, Y.; Wang, J. Plant diversity as a good indicator of vegetation stability in a typical plateau wetland. J. Mt. Sci. Engl. 2014, 11, 464–474. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, J.; Chang, S.-X.; Jiang, P.; Zhou, G.; Fu, S.; Yan, E.; Wu, J.; Lin, L. Long-term intensive management effects on soil organic carbon pools and chemical composition in Moso bamboo (Phyllostachys pubescens) forests in subtropical China. For. Ecol. Manag. 2013, 303, 121–130. [Google Scholar] [CrossRef]
- Zhang, M.-M.; Fan, S.-H.; Yan, X.-R.; Zhou, Y.-Q.; Guan, F.-Y. Relationships between stand spatial structure characteristics and influencing factors of bamboo and broadleaf mixed forest. J. For. Res. Jpn. 2020, 25, 1–9. [Google Scholar]
- IUSS Working Group WRB 2014. World Reference Base for Soil Resources 2014. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; FAO: Rome, Italy, 2014. [Google Scholar]
- Schad, P. The international soil classification system WRB 2014. In Novel Methods for Monitoring and Managing Land and Water Resources in Siberia; Springer: Cham, Switzerland, 2016; pp. 563–571. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Elsevier: New York, NY, USA, 2012. [Google Scholar]
- Shaohui, F.; Fuming, X.; Silong, W.; Wenhui, S.; Xiaojun, Y.; Zhengqi, S. Fine Root Biomass and Turnover in Moso Bamboo Plantation in Huitong Forest Station, Hunan Province. Sci. Silvae Sin. 2009, 45, 1–6. [Google Scholar]
- Liu, G.; Fan, S.; Cai, C.; Liu, X.; Li, Y.; Luo, T. Fine Root Biomass Distribution of Moso Bamboo at Different Ages. J. Trop. Subtrop. Bot. 2017, 25, 472–479. [Google Scholar]
- Zhao, L.; Wang, J. Research on Relations Between Growth Effect and Soil Enzyme Activities and Soil Nutrient Factors in Mixed Poplar and Black Locust Plantations. J. Beijing For. Univ. 1995, 17, 1–8. [Google Scholar]
- Yuan, W.; Li, X.; Zhang, J.; Rong, L. Preliminary studies on carbon reserves of litterfall and fine root in an age series of Eucalyptus grandis plantation. Sci. Silvae Sin. 2009, 22, 385–389. [Google Scholar]
- Miao, Y.; Chen, Y.-L.; Li, X.W.; Fan, C.; Liu, Y.-K.; Yang, Z.-J.; Zhang, J.; Cai, X.-L. Effects of fertilization on Alnus formosana fine root morphological characteristics, biomass and issue content of C N under A. formosana-Hemarthria compressa compound mode. Chin. J. Plant Ecol. 2013, 37, 674–683. [Google Scholar]
- Wijesinghe, D.K.; John, E.A.; Beurskens, S.; Hutchings, M.J. Root system size and precision in nutrient foraging: Responses to spatial pattern of nutrient supply in six herbaceous species. J. Ecol. 2001, 89, 972–983. [Google Scholar] [CrossRef]
- Ma, Z.; Chen, H.Y.H. Effects of species diversity on fine root productivity increase with stand development and associated mechanisms in a boreal forest. J. Ecol. 2017, 105, 237–245. [Google Scholar] [CrossRef]
- Zhou, Y.; Guan, F.; Fan, S. Niche characteristics of Phyllostachys edulis and its associated tree species in Tianbaoyan bamboo and broadleaved mixed forest. J. Beijing For. Univ. 2017, 39, 46–53. [Google Scholar]
- Wang, L.; Zhong, C.; Cai, J. Spatial distribution and morphological variations of the fine root in walnut-wheat intercropping agroforestry ecosystem. J. Northwest A F Univ. Nat. Sci. Ed. 2011, 39, 64–70. [Google Scholar]
- Zamora, D.S.; Jose, S.; Nair, P.K.R. Morphological plasticity of cotton root in response to interspecific competition with pecan in an alleycropping system in the southern United States. Agrofor. Syst. 2007, 69, 107–116. [Google Scholar] [CrossRef]
- Davis, J.P.; Haines, B.; Coleman, D.; Hendrick, R. Fine root dynamics along an elevational gradient in the southern Appalachian Mountains, USA. For. Ecol. Manag. 2004, 187, 19–33. [Google Scholar] [CrossRef]
- Curt, T.; Prévosto, B. Rooting strategy of naturally regenerated beech in Silver birch and Scots pine woodlands. In Roots: The Dynamic Interface between Plants and the Earth; Springer: Dordrecht, The Netherlands, 2003; pp. 265–279. [Google Scholar]
- Zheng, Y.; Guan, F.; Fan, S.; Zhou, Y.; Jing, X. Functional trait responses to strip clearcutting in a Moso bamboo forest. Forests 2021, 12, 793. [Google Scholar] [CrossRef]
- Kooch, Y.; Tarighat, F.S.; Hosseini, S.M. Tree species effects on soil chemical, biochemical and biological features in mixed Caspian lowland forests. Trees 2017, 31, 863–872. [Google Scholar] [CrossRef]
- Miller, H.G. Dynamics of nutrient cycling in plantation ecosystems. In Nutrition of Plantation Forests; Bowen, G.D., Nambiar, E.K.S., Eds.; FAO: Rome, Italy, 1984. [Google Scholar]
- Wang, B.; Xue, S.; Liu, G.B.; Zhang, G.H.; Li, G.; Ren, Z.P. Changes in soil nutrient and enzyme activities under different vegetations in the Loess Plateau area, Northwest China. Catena 2012, 92, 186–195. [Google Scholar] [CrossRef]
- Hao, C.; Dong, S.; Lei, L.; Mo, C.; Zhang, T.; Zhu, X.; Mo, J. Effects of Experimental Nitrogen and Phosphorus Addition on Litter Decomposition in an Old-Growth Tropical Forest. PLoS ONE 2013, 8, e84101. [Google Scholar]
- Zhang, M.; Fan, S.; Guan, F.; Yan, X.; Yin, Z. Soil bacterial community structure of mixed bamboo and broad-leaved forest based on tree crown width ratio. Sci. Rep. 2020, 10, 6522. [Google Scholar]
- Moghimian, N.; Hosseini, S.M.; Kooch, Y.; Darki, B.Z. Impacts of changes in land use/cover on soil microbial and enzyme activities. Catena 2017, 157, 407–414. [Google Scholar] [CrossRef]
- Rong, L.; Shou-Jian, L.I.; Xian-Wei, L.I. Soil enzyme dynamics during fine root (including grass root) decomposition in different farmland-to-forest/grassland conversions in the rainy zone of western China. Chin. J. Plant Ecol. 2010, 34, 642–650. [Google Scholar]
- Dick, R.P. Soil enzyme activities as indicators of soil quality. Soil Sci. Soc. Am. J. 1994, 58, 107–124. [Google Scholar]
- Avazpoor, Z.; Moradi, M.; Basiri, R.; Mirzaei, J.; Taghizadeh-Mehrjardi, R.; Kerry, R. Soil enzyme activity variations in riparian forests in relation to plant species and soil depth. Arab. J. Geosci. 2019, 12, 708. [Google Scholar] [CrossRef]
- Ushio, M.; Kitayama, K.; Balser, T.C. Tree species effects on soil enzyme activities through effects on soil physicochemical and microbial properties in a tropical montane forest on Mt. Kinabalu, Borneo. Pedobiologia 2010, 53, 227–233. [Google Scholar] [CrossRef]
- Behbahani, S.M.; Moradi, M.; Basiri, R.; Mirzaei, J. Sand mining disturbances and their effects on the diversity of arbuscular mycorrhizal fungi in a riparian forest of Iran. J. Arid Land 2017, 9, 837–849. [Google Scholar] [CrossRef] [Green Version]
- Sardans, J.; Peñuelas, J. Plant-soil interactions in Mediterranean forest and shrublands: Impacts of climatic change. Plant Soil 2013, 365, 1–33. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Type | Age of Broadleaf Tree (a) | DBH of Broadleaf Tree (cm) | Average DBH of Bamboo (cm) | Culm Density (culm ha−1) | Altitude (m) | Slope (°) | Aspect |
|---|---|---|---|---|---|---|---|
| CCB | 53 | 40.82 | 8.89 | 2829 | 796 | 20 | adret |
| AFB | 55 | 49.11 | 9.15 | 2712 | 770 | 25 | adret |
| CAB | 44 | 41.36 | 9.48 | 2868 | 778 | 22 | adret |
| CFB | 56 | 53.55 | 9.37 | 2987 | 789 | 23 | adret |
| Stand Type | Soil Layer | SOC | TN | TP | TK | AN | AP | AK |
|---|---|---|---|---|---|---|---|---|
| cm | (g/kg) | (g/kg) | (g/kg) | (g/kg) | (mg/kg) | (mg/kg) | (mg/kg) | |
| 0–20 | 30.38 ± 1.03 Ab | 2.12 ± 0.10 Ac | 0.33 ± 0.01 Ab | 12.99 ± 2.39 Ac | 144.91 ± 2.34 Aa | 1.69 ± 0.17 Ad | 32.77 ± 2.68 Ab | |
| CCB | 20–40 | 23.43 ± 2.78 Bab | 1.87 ± 0.05 Bb | 0.30 ± 0.01A Bc | 13.25 ± 0.82 Ad | 114.89 ± 5.45 Ba | 0.88 ± 0.05 Bc | 21.97 ± 4.32 Bb |
| 40–60 | 14.93 ± 0.83 Ca | 1.84 ± 0.12 Ba | 0.29 ± 0.03 Ba | 12.67 ± 1.53 Ad | 101.73 ± 3.19 Ca | 0.13 ± 0.02 Cab | 18.44 ± 3.70 Bb | |
| 0–20 | 34.66 ± 2.52 Ab | 2.49 ± 0.24 Ab | 0.39 ± 0.01 Aa | 36.27 ± 4.56 Aa | 127.99 ± 1.16 Ab | 3.71 ± 0.10 Aa | 45.97 ± 5.88 Aab | |
| AFB | 20–40 | 24.30 ± 2.51 Bab | 1.93 ± 0.15 Bab | 0.39 ± 0.03 Aa | 37.08 ± 3.53 Aa | 105.05 ± 2.76 Bab | 1.24 ± 0.09 Ba | 36.18 ± 2.22 Ba |
| 40–60 | 17.10 ± 1.30 Ca | 1.84 ± 0.07 Ba | 0.30 ± 0.02 Ba | 37.48 ± 4.41 Aa | 85.61 ± 1.20 Cc | 0.15 ± 0.01 Cab | 17.69 ± 1.23 Cb | |
| 0–20 | 46.71 ± 7.53 Aa | 2.81 ± 0.11 Aa | 0.39 ± 0.01 Aa | 29.13 ± 3.27 Ab | 127.93 ± 1.45 Ab | 3.44 ± 0.09 Ab | 54.39 ± 12.25 Aa | |
| CAB | 20–40 | 27.19 ± 3.25 Ba | 2.17 ± 0.21 Ba | 0.36 ± 0.01 Bb | 28.54 ± 2.79 Ab | 105.09 ± 6.98 Bab | 1.11 ± 0.14 Bab | 43.27 ± 4.29 ABa |
| 40–60 | 17.29 ± 1.86 Ca | 1.83 ± 0.17 Ca | 0.27 ± 0.01 Cab | 29.29 ± 3.46 Ab | 90.92 ± 2.59 Cb | 0.17 ± 0.03 Ca | 28.9 ± 3.35 Ba | |
| 0–20 | 30.23 ± 1.77 Ab | 2.26 ± 0.18 Abc | 0.28 ± 0.01 Ac | 19.88 ± 4.31 Ac | 104.25 ± 7.09 Ac | 2.07 ± 0.05 Ac | 35.66 ± 0.62 Ab | |
| CFB | 20–40 | 20.05 ± 1.30 Bb | 1.95 ± 0.08 Bab | 0.27 ± 0.01 Ac | 21.12 ± 4.21 Ac | 94.82 ± 6.81 Bb | 1.03 ± 0.04 Bbc | 24.17 ± 6.24 Bb |
| 40–60 | 16.06 ± 0.61 Ca | 1.80 ± 0.08 Ba | 0.24 ± 0.02 Bb | 20.96 ± 4.00 Ac | 82.07 ± 1.89 Cc | 0.12 ± 0.02 Cb | 29.93 ± 1.62 ABa |
| FRB | SRL | RLD | SSA | SOC | TN | TP | TK | AN | AP | AK | SC | UE | ACP | CAT | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dim. 1 | 9.8908 | 9.2673 | 10.0738 | 7.7663 | 9.7735 | 9.0609 | 5.7811 | 0.8213 | 10.4241 | 5.6534 | 8.1469 | 5.3079 | 5.7967 | 0.2791 | 1.9562 |
| Dim. 2 | 2.5446 | 2.6883 | 2.4592 | 0.1151 | 0.0410 | 0.0067 | 6.6185 | 26.8863 | 0.3373 | 2.2351 | 2.4934 | 5.5981 | 11.1667 | 29.5147 | 7.2946 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Guan, F.; Li, Z.; Zheng, Y.; Zhou, X.; Zhang, X. Effects of Tree Species on Moso Bamboo (Phyllostachys edulis (Carriere) J. Houzeau) Fine Root Morphology, Biomass, and Soil Properties in Bamboo–Broadleaf Mixed Forests. Forests 2022, 13, 1834. https://doi.org/10.3390/f13111834
Zhou Y, Guan F, Li Z, Zheng Y, Zhou X, Zhang X. Effects of Tree Species on Moso Bamboo (Phyllostachys edulis (Carriere) J. Houzeau) Fine Root Morphology, Biomass, and Soil Properties in Bamboo–Broadleaf Mixed Forests. Forests. 2022; 13(11):1834. https://doi.org/10.3390/f13111834
Chicago/Turabian StyleZhou, Yang, Fengying Guan, Zhen Li, Yaxiong Zheng, Xiao Zhou, and Xuan Zhang. 2022. "Effects of Tree Species on Moso Bamboo (Phyllostachys edulis (Carriere) J. Houzeau) Fine Root Morphology, Biomass, and Soil Properties in Bamboo–Broadleaf Mixed Forests" Forests 13, no. 11: 1834. https://doi.org/10.3390/f13111834