
Effects of Exotic and Native Earthworms on Soil Micro-Decomposers in a Subtropical Forest: A Field Mesocosm Experiment

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
- (1)
- Exotic earthworm: 30 exotic earthworms (P. corethrurus) were added;
- (2)
- Native earthworm: 20 native earthworms (Amynthas corticis) were added;
- (3)
- Control: no earthworms.
2.3. Soil Sampling and Laboratory Analysis
2.4. Data Analysis
2.4.1. Nematode Abundance, Community Indices, and Diversity Indices
2.4.2. Analysis of Treatment Effects
3. Results
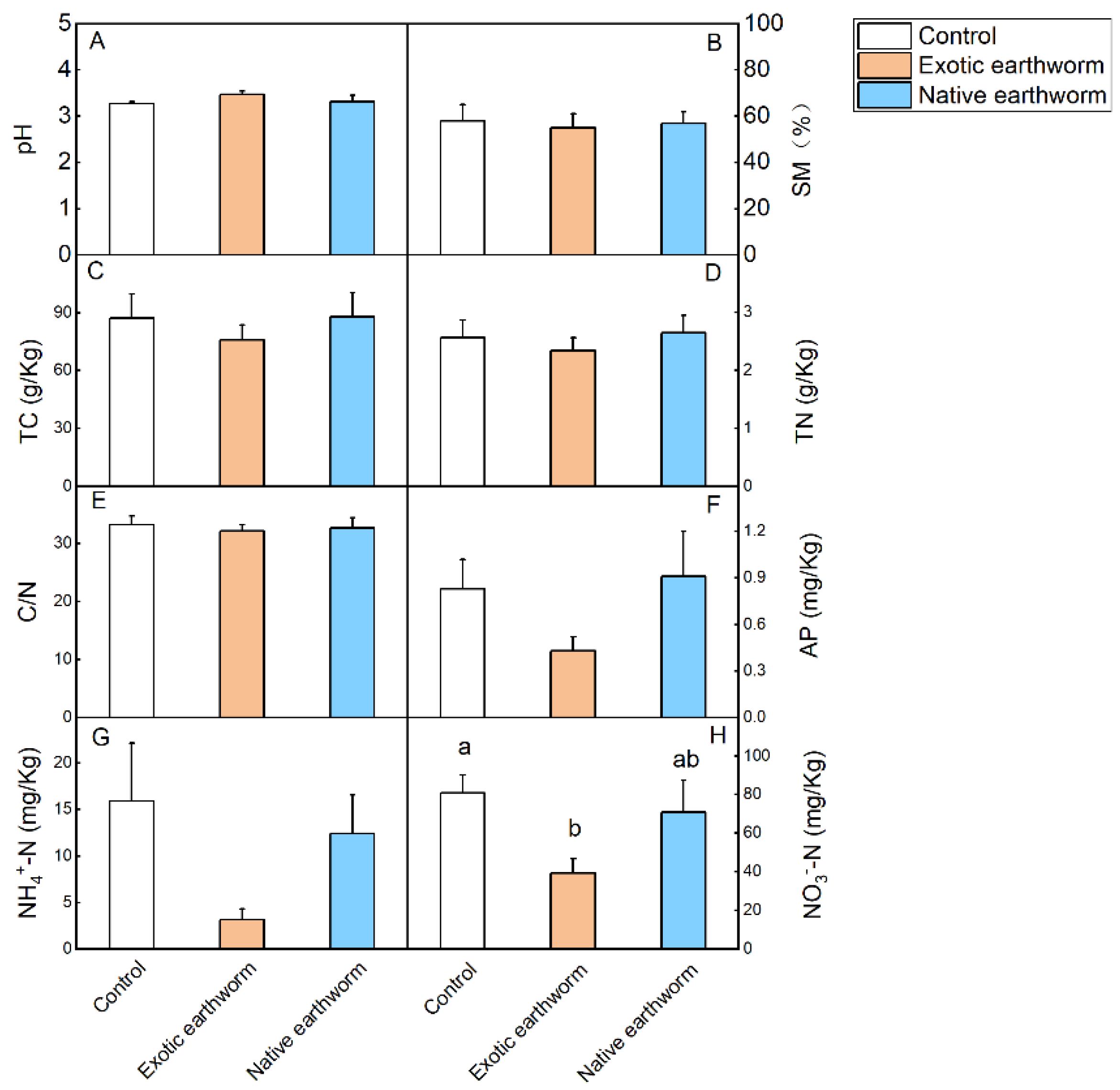
3.1. Soil Chemical Properties
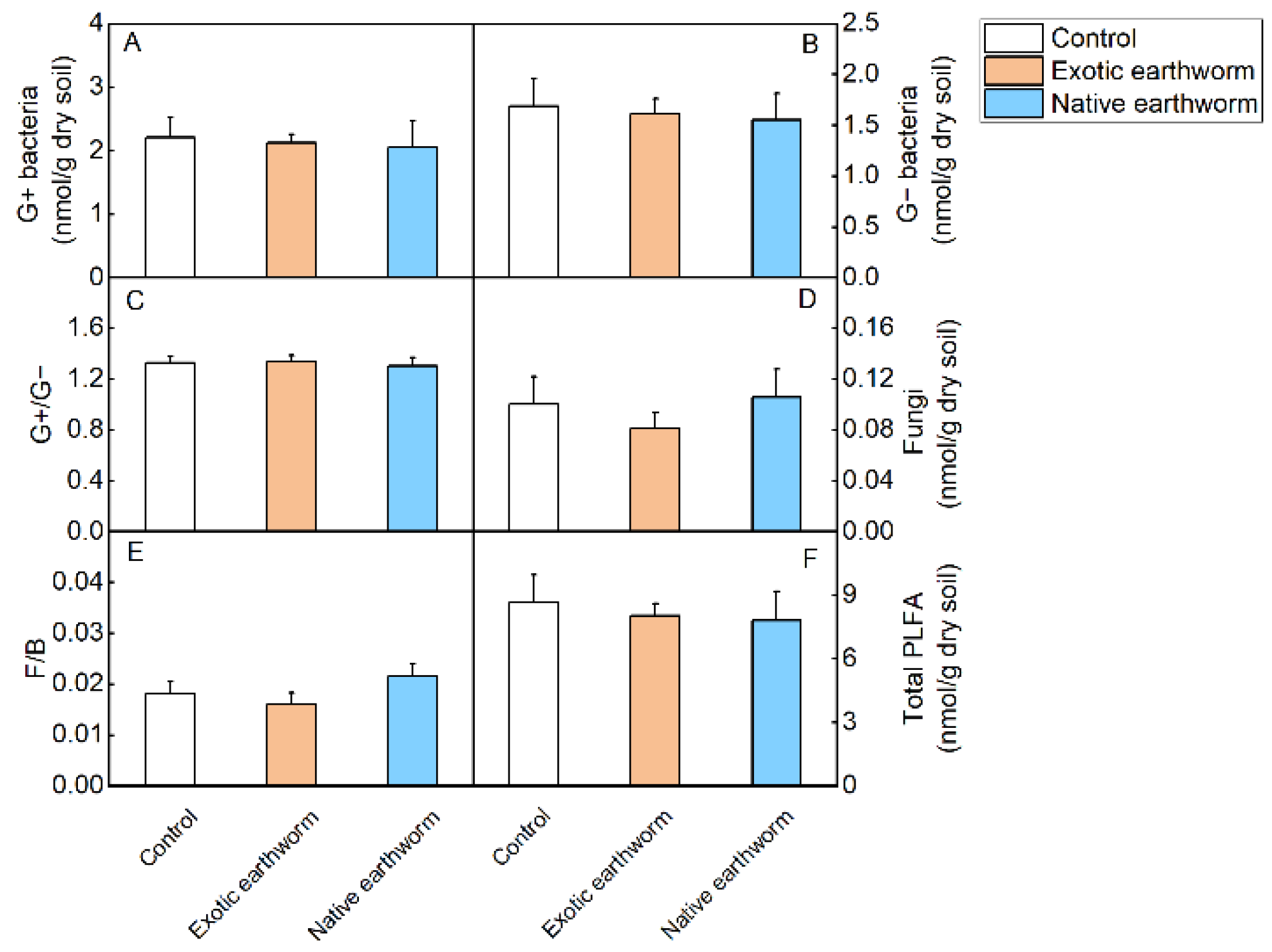
3.2. Soil Microbial Community Characteristics
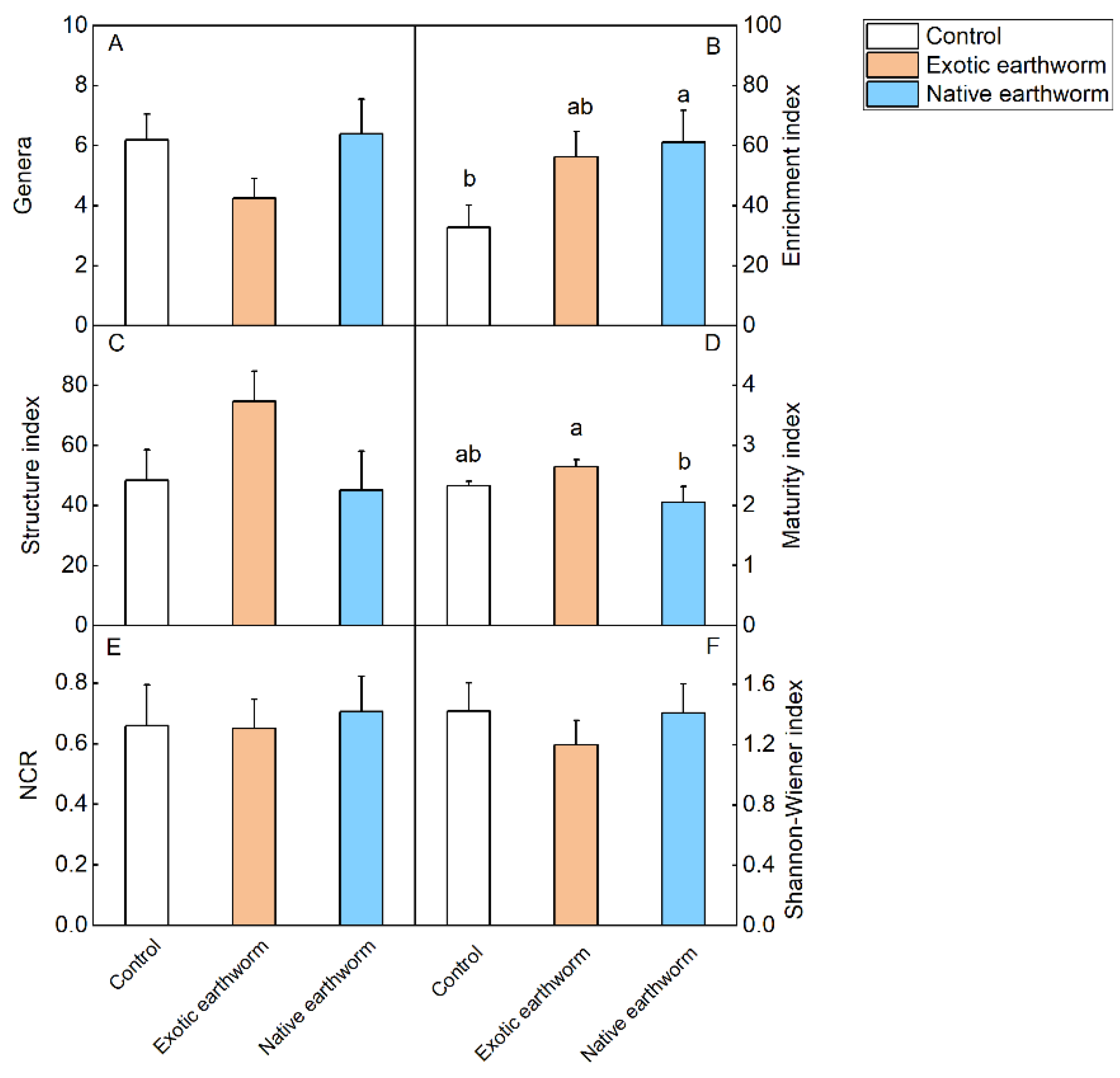
3.3. Soil Nematode Community Characteristics
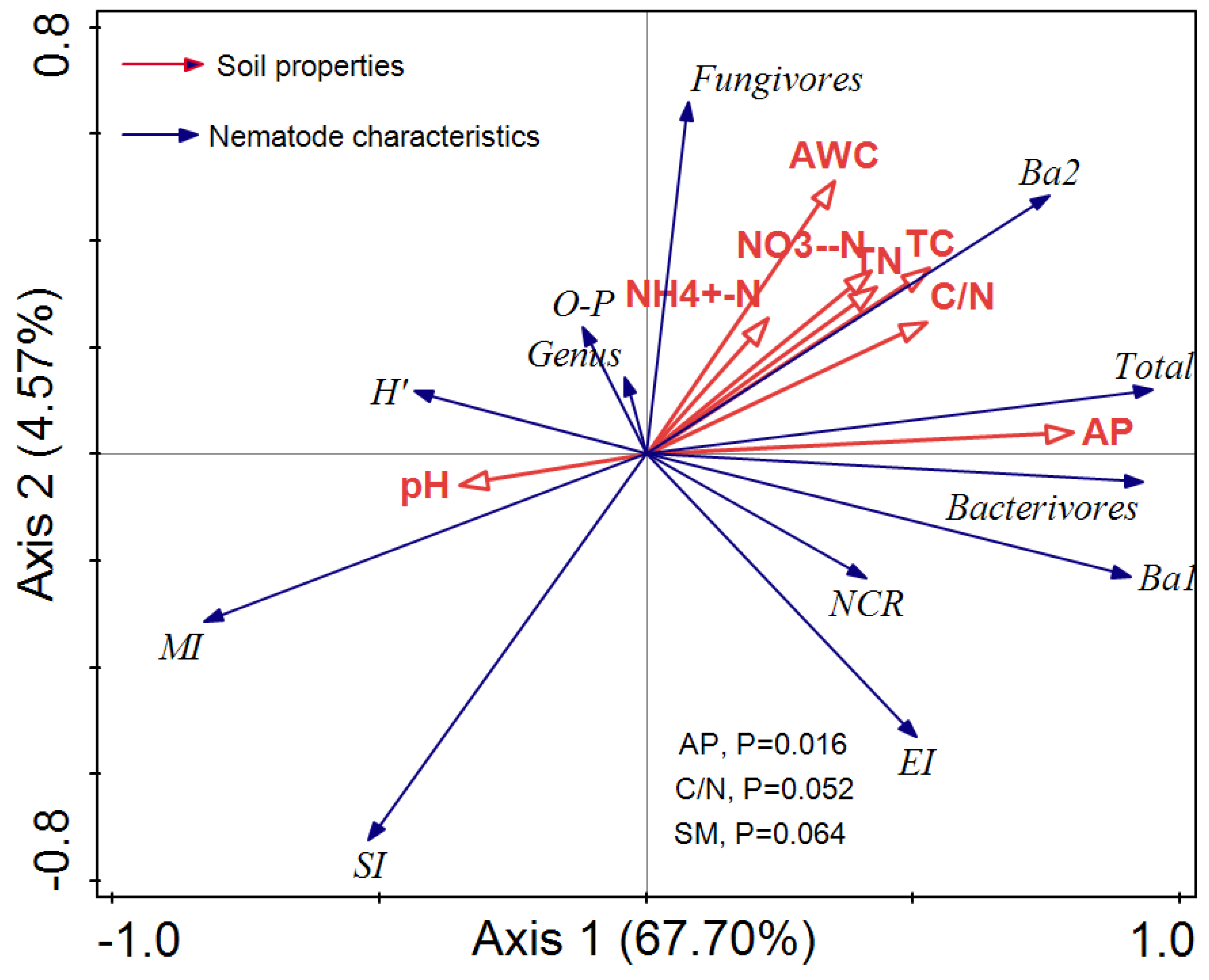
3.4. Associations between Soil Properties and Nematode Abundance and Diversity
4. Discussion
4.1. Effects of Exotic and Native Earthworms on Soil Microbial Community
4.2. Effects of Exotic and Native Earthworms on Soil Nematode Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wardle, D.A.; Bardgett, R.D.; Callaway, R.M.; Van der Putten, W.H. Terrestrial Ecosystem Responses to Species Gains and Losses. Science 2011, 332, 1273–1277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. World Soil Day—Keep soil alive, Protect soil biodiversity. In World Soil Day 2020 Campaign Report; Food and Agriculture Organization of the United Nations: Rome, Italy, 2021. [Google Scholar]
- Ehrenfeld, J.G.; Scott, N. Invasive species and the soil: Effects on organisms and ecosystem processes. Ecol. Appl. 2001, 11, 1259–1260. [Google Scholar] [CrossRef]
- Jamieson, B.G.M. On the phylogeny and higher classification of the oligochaeta. Cladistics 1988, 4, 367–410. [Google Scholar] [CrossRef]
- Reynolds, J.W. Earthworms of the world. Glob. Biodivers. 1994, 4, 11–16. [Google Scholar]
- Eisenhauer, N.; Ferlian, O.; Craven, D.; Hines, J.; Jochum, M. Ecosystem responses to exotic earthworm invasion in northern North American forests. Res. Ideas Outcomes 2019, 5, e34564. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Cameron, E.K.; Martins, I.S.; Lavelle, P.; Mathieu, J.; Tedersoo, L.; Bahram, M.; Gottschall, F.; Guerra, C.A.; Hines, J.; Patoine, G.; et al. Global mismatches in aboveground and belowground biodiversity. Conserv. Biol. 2019, 33, 1187–1192. [Google Scholar] [CrossRef] [Green Version]
- Hendrix, P.F.; Bohlen, P.J. Exotic earthworm invasions in North America: Ecological and policy implications. Bioscience 2002, 52, 801–811. [Google Scholar] [CrossRef] [Green Version]
- Righi, G. Pontoscolex (oligochaeta, glossoscolecidae), a new evaluation. Stud. Neotrop. Fauna Environ. 1984, 19, 159–177. [Google Scholar] [CrossRef]
- Garcia, J.A.; Fragoso, C. Influence of different food substrates on growth and reproduction of two tropical earthworm species (Pontoscolex corethrurus and Amynthas corticis). Pedobiologia 2003, 47, 754–763. [Google Scholar] [CrossRef]
- Lavelle, P.; Pashanasi, B. Soil macrofauna and land management in peruvian amazonia (yurimaguas, loreto). Pedobiologia 1989, 33, 283–291. [Google Scholar]
- Fragoso, C.; Lavelle, P.; Blanchart, E.; Senapati, B.K.; Jimenez, J.J.; Martinez, M.D.; Decaens, T.; Tondoh, J. Earthworm communities of tropical agroecosystems: Origin, structure and influence of management practices. Earthworm Manag. Trop. Agroecosystems 1999, 27–55. [Google Scholar]
- Taheri, S.; Pelosi, C.; Dupont, L. Harmful or useful? A case study of the exotic peregrine earthworm morphospecies Pontoscolex corethrurus. Soil. Biol. Biochem. 2018, 116, 277–289. [Google Scholar] [CrossRef]
- He, X.; Chen, Y.; Liu, S.; Gunina, A.; Wang, X.; Chen, W.; Shao, Y.; Shi, L.; Yao, Q.; Li, J.; et al. Cooperation of earthworm and arbuscular mycorrhizae enhanced plant N uptake by balancing absorption and supply of ammonia. Soil. Biol. Biochem. 2018, 116, 351–359. [Google Scholar] [CrossRef]
- Lavelle, P.; Bignell, D.; Lepage, M.; Wolters, V.; Roger, P.; Ineson, P.; Heal, O.W.; Dhillion, S. Soil function in a changing world: The role of invertebrate ecosystem engineers. Eur. J. Soil. Biol. 1997, 33, 159–193. [Google Scholar]
- Liu, T.; Chen, X.; Gong, X.; Lubbers, I.M.; Jiang, Y.; Feng, W.; Li, X.; Whalen, J.K.; Bonkowski, M.; Griffiths, B.S.; et al. Earthworms Coordinate Soil Biota to Improve Multiple Ecosystem Functions. Curr. Biol. 2019, 29, 3420–3429.e5. [Google Scholar] [CrossRef]
- Puga-Freitas, R.; Barot, S.; Taconnat, L.; Renou, J.-P.; Blouin, M. Signal Molecules Mediate the Impact of the Earthworm Aporrectodea caliginosa on Growth, Development and Defence of the Plant Arabidopsis thaliana. PLoS ONE 2012, 7, e49504. [Google Scholar] [CrossRef] [Green Version]
- Blouin, M.; Hodson, M.E.; Delgado, E.A.; Baker, G.; Brussaard, L.; Butt, K.R.; Dai, J.; Dendooven, L.; Peres, G.; Tondoh, J.E.; et al. A review of earthworm impact on soil function and ecosystem services. Eur. J. Soil. Sci. 2013, 64, 161–182. [Google Scholar] [CrossRef]
- Lubbers, I.M.; van Groenigen, K.J.; Fonte, S.J.; Six, J.; Brussaard, L.; van Groenigen, J.W. Greenhouse-gas emissions from soils increased by earthworms. Nat. Clim. Chang. 2013, 3, 187–194. [Google Scholar] [CrossRef]
- Zaborski, E.R.; Soeken Gittenger, L.A.; Roberts, S.J. A possible Phasmarhabditis sp. (Nematoda: Rhabditidae) isolated from Lumbricus terrestris (Oligochaeta: Lumbricidae). J. Invertebr. Pathol. 2001, 77, 284–287. [Google Scholar] [CrossRef]
- Scheu, S. Effects of earthworms on plant growth: Patterns and perspectives. Pedobiologia 2003, 47, 846–856. [Google Scholar] [CrossRef]
- Ferlian, O.; Thakur, M.P.; Gonzalez, A.C.; San Emeterio, L.M.; Marr, S.; Rocha, B.D.; Eisenhauer, N. Soil chemistry turned upside down: A meta-analysis of invasive earthworm effects on soil chemical properties. Ecology 2020, 101, e02936. [Google Scholar] [CrossRef] [PubMed]
- Ferlian, O.; Eisenhauer, N.; Aguirrebengoa, M.; Camara, M.; Ramirez-Rojas, I.; Santos, F.; Tanalgo, K.; Thakur, M.P. Invasive earthworms erode soil biodiversity: A meta-analysis. J. Anim. Ecol. 2018, 87, 162–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craven, D.; Thakur, M.P.; Cameron, E.K.; Frelich, L.E.; Beausejour, R.; Blair, R.B.; Blossey, B.; Burtis, J.; Choi, A.; Davalos, A.; et al. The unseen invaders: Introduced earthworms as drivers of change in plant communities in North American forests (a meta-analysis). Glob. Chang. Biol. 2017, 23, 1065–1074. [Google Scholar] [CrossRef]
- Bohlen, P.J.; Scheu, S.; Hale, C.M.; McLean, M.A.; Migge, S.; Groffman, P.M.; Parkinson, D. Non-native invasive earthworms as agents of change in northern temperate forests. Front. Ecol. Environ. 2004, 2, 427–435. [Google Scholar] [CrossRef]
- Dalby, P.R.; Baker, G.H.; Smith, S.E. Potential impact of an introduced lumbricid on a native woodland in South Australia. Appl. Soil Ecol. 1998, 9, 351–354. [Google Scholar] [CrossRef]
- Alban, D.H.; Berry, E.C. Effects of earthworm invasion on morphology, carbon, and nitrogen of a forest soil. Appl. Soil Ecol. 1994, 1, 243–249. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Partsch, S.; Parkinson, D.; Scheu, S. Invasion of a deciduous forest by earthworms: Changes in soil chemistry, microflora, microarthropods and vegetation. Soil. Biol. Biochem. 2007, 39, 1099–1110. [Google Scholar] [CrossRef]
- Van den Hoogen, J.; Geisen, S.; Routh, D.; Ferris, H.; Traunspurger, W.; Wardle, D.A.; de Goede, R.G.M.; Adams, B.J.; Ahmad, W.; Andriuzzi, W.S.; et al. Soil nematode abundance and functional group composition at a global scale. Nature 2019, 572, 194–198. [Google Scholar] [CrossRef] [Green Version]
- Yeates, G.W.; Bongers, T.; Degoede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding-habits in soil nematode families and genera—An outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar]
- Yeates, G.W. Nematodes as soil indicators: Functional and biodiversity aspects. Biol. Fert. Soils 2003, 37, 199–210. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, X.; Shao, Y.; Xu, G.; Fu, S. Effects of vegetation removal on soil properties and decomposer organisms. Soil. Biol. Biochem. 2011, 43, 954–960. [Google Scholar] [CrossRef]
- Shao, Y.H.; Zhang, W.X.; Liu, S.J.; Wang, X.L.; Fu, S.L. Diversity and function of soil fauna. Acta Ecol. Sin. 2015, 35, 6614–6625. [Google Scholar]
- Ilieva-Makulec, K.; Makulec, G. Effect of the earthworm Lumbricus rubellus on the nematode community in a peat meadow soil. Eur. J. Soil. Biol. 2002, 38, 59–62. [Google Scholar] [CrossRef]
- Tao, J.; Chen, X.; Liu, M.; Hu, F.; Griffiths, B.; Li, H. Earthworms change the abundance and community structure of nematodes and protozoa in a maize residue amended rice-wheat rotation agro-ecosystem. Soil. Biol. Biochem. 2009, 41, 898–904. [Google Scholar] [CrossRef]
- Shapiro, D.I.; Berry, E.C.; Lewis, L.C. Interactions between nematodes and earthworms-enhanced dispersal of steinernema-carpocapsae. J. Nematol. 1993, 25, 189–192. [Google Scholar]
- Yu, F.; Li, C.; Liu, T.; Li, T.; Hu, F.; Li, H.; Jiao, J. Earthworm mucus interfere in the behavior and physiology of bacterial-feeding nematodes. Appl. Soil Ecol. 2019, 143, 107–115. [Google Scholar] [CrossRef]
- Dash, M.C.; Senapati, B.K.; Mishra, C.C. Nematode feeding by tropical earthworms. Oikos 1980, 34, 322–325. [Google Scholar] [CrossRef]
- Räty, M.; Huhta, V. Earthworms and pH affect communities of nematodes and enchytraeids in forest soil. Biol. Fert. Soils 2003, 38, 52–58. [Google Scholar] [CrossRef]
- Senapati, B.K. Biotic interactions between soil nematodes and earthworms. Soil. Biol. Biochem. 1992, 24, 1441–1444. [Google Scholar] [CrossRef]
- Dempsey, M.A.; Fisk, M.C.; Fahey, T.J. Earthworms increase the ratio of bacteria to fungi in northern hardwood forest soils, primarily by eliminating the organic horizon. Soil. Biol. Biochem. 2011, 43, 2135–2141. [Google Scholar] [CrossRef]
- Lejoly, J.; Quideau, S.; Laganiere, J. Invasive earthworms affect soil morphological features and carbon stocks in boreal forests. Geoderma 2021, 404, 115262. [Google Scholar] [CrossRef]
- Chang, C.H.; Bartz, M.L.C.; Brown, G.; Callaham, M.A.; Cameron, E.K.; Davalos, A.; Dobson, A.; Gorres, J.H.; Herrick, B.M.; Ikeda, H.; et al. The second wave of earthworm invasions in North America: Biology, environmental impacts, management and control of invasive jumping worms. Biol. Invasions 2021, 23, 3291–3322. [Google Scholar] [CrossRef]
- Jochum, M.; Ferlian, O.; Thakur, M.P.; Ciobanu, M.; Klarner, B.; Salamon, J.A.; Frelich, L.E.; Johnson, E.A.; Eisenhauer, N. Earthworm invasion causes declines across soil fauna size classes and biodiversity facets in northern North American forests. Oikos 2021, 130, 766–780. [Google Scholar] [CrossRef]
- Addison, J.A. Distribution and impacts of invasive earthworms in Canadian forest ecosystems. Biol. Invasions 2009, 11, 59–79. [Google Scholar] [CrossRef]
- Hendrix, P.F. Biological invasions belowground—Earthworms as invasive species. Biol. Invasions 2006, 8, 1201–1204. [Google Scholar] [CrossRef]
- Lee, K.E.; Foster, R.C. Soil fauna and soil structure. Aust. J. Soil Res. 1991, 29, 745–775. [Google Scholar] [CrossRef]
- Wang, X.; Fu, S.; Wang, X.; Li, Z.; Li, J.; Zhang, W. One-Year Monitoring of Daily Earthworm Cast Production: Surface Cast Contribution to Soil Fertility in a Subtropical Forest. Forests 2021, 12, 865. [Google Scholar] [CrossRef]
- Zhang, W.X.; Li, J.X.; Guo, M.F.; Liao, C.H. Seasonal variation of earthworm community structure as correlated with environmental factors in three plantation of Heshan, Guangdong, China. Acta Ecol. Sinica 2005, 25, 1362–1370. [Google Scholar]
- Liu, T.; Wang, X.; Zhang, W.; Eisenhauer, N.; Shao, Y.; Zhao, J.; Li, J.; He, X.; Tao, L.; Fu, S.; et al. Earthworms regulate the nematode community by directly enhancing the bacterial-based energy channel rather than through the effect of casts. Pedobiologia 2022, 95, 150843. [Google Scholar] [CrossRef]
- Liu, G. Analysis of Soil Physical and Chemical Properties and Description of Soil Profiles; China Standard: Beijing, China, 1996. [Google Scholar]
- Fang, Y.T.; Gundersen, P.; Mo, J.M.; Zhu, W.X. Input and output of dissolved organic and inorganic nitrogen in subtropical forests of South China under high air pollution. Biogeosciences 2008, 5, 339–352. [Google Scholar] [CrossRef] [Green Version]
- Bossio, D.A.; Scow, K.M. Impacts of carbon and flooding on soil microbial communities: Phospholipid fatty acid profiles and substrate utilization patterns. Microb. Ecol. 1998, 35, 265–278. [Google Scholar] [CrossRef]
- Shi, L.; Zhang, H.; Liu, T.; Mao, P.; Zhang, W.; Shao, Y.; Fu, S. An increase in precipitation exacerbates negative effects of nitrogen deposition on soil cations and soil microbial communities in a temperate forest. Environ. Pollut. 2018, 235, 293–301. [Google Scholar] [CrossRef]
- Barker, K.R. Nematode extraction and bioassays. In An Advanced Treatise on Meloidogyne; Barker, K.R., Carter, C.C., Sasser, J.N., Eds.; North Carolina State University: Raleigh, NC, USA, 1985; Volume 2, pp. 19–35. [Google Scholar]
- Ferris, H.; Bongers, T.; de Goede, R.G.M. A framework for soil food web diagnostics: Extension of the nematode faunal analysis concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Bongers, T. The maturity index—An ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef]
- Neher, D.A.; Darby, B.J. General community indices that can be used for analysis of nematode assemblages. In Nematodes as Environmental Indicators; Wilson, M.J., Kakouli-Duarte, T., Eds.; CABI: Wallingford, UK, 2009; pp. 107–123. [Google Scholar]
- Huang, J.; Zhang, W.; Liu, M.; Briones, M.J.I.; Eisenhauer, N.; Shao, Y.; Cai, X.a.; Fu, S.; Xia, H. Different impacts of native and exotic earthworms on rhizodeposit carbon sequestration in a subtropical soil. Soil. Biol. Biochem. 2015, 90, 152–160. [Google Scholar] [CrossRef]
- Zhang, B.; Li, G.; Shen, T.; Wang, J.; Sun, z. Changes in microbial biomass C, N, and P and enzyme activities in soil incubated with the earthworms Metaphire guillelmi or Eisenia fetida. Soil. Biol. Biochem. 2000, 32, 2055–2062. [Google Scholar] [CrossRef]
- McLean, M.A.; Migge-Kleian, S.; Parkinson, D. Earthworm invasions of ecosystems devoid of earthworms: Effects on soil microbes. Biol. Invasions 2006, 8, 1257–1273. [Google Scholar] [CrossRef]
- Elliott, E.T.; Anderson, R.V.; Coleman, D.C.; Cole, C.V. Habitable pore space and microbial trophic interactions. Oikos 1980, 35, 327–335. [Google Scholar] [CrossRef]
- Niu, X.; Zhai, P.; Zhang, W.; Gu, Y. Effects of Earthworms and Agricultural Plant Species on the Soil Nematode Community in a Microcosm Experiment. Sci. Rep. 2019, 9, 11660. [Google Scholar] [CrossRef] [Green Version]
- Berkelmans, R.; Ferris, H.; Tenuta, M.; van Bruggen, A.H.C. Effects of long-term crop management on nematode trophic levels other than plant feeders disappear after 1 year of disruptive soil management. Appl. Soil Ecol. 2003, 23, 223–235. [Google Scholar] [CrossRef]
- Demetrio, W.C.; Dionisio, J.A.; Maceda, A. Negative effects of earthworms on soil nematodes are dependent on earthworm density, ecological category and experimental conditions. Pedobiologia 2019, 76, 150568. [Google Scholar] [CrossRef]
- Curry, J.P.; Schmidt, O. The feeding ecology of earthworms—A review. Pedobiologia 2007, 50, 463–477. [Google Scholar] [CrossRef]
- Xiao, H.; Wang, W.; Xia, S.; Li, Z.; Gan, J.; Yang, X. Distributional patterns of soil nematodes in relation to environmental variables in forest ecosystems. Soil Ecol. Lett. 2021, 3, 115–124. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Z.; Wang, X.; Zhang, W.; Lu, X.; Liu, T. Effects of Exotic and Native Earthworms on Soil Micro-Decomposers in a Subtropical Forest: A Field Mesocosm Experiment. Forests 2022, 13, 1924. https://doi.org/10.3390/f13111924
Zhao Z, Wang X, Zhang W, Lu X, Liu T. Effects of Exotic and Native Earthworms on Soil Micro-Decomposers in a Subtropical Forest: A Field Mesocosm Experiment. Forests. 2022; 13(11):1924. https://doi.org/10.3390/f13111924
Chicago/Turabian StyleZhao, Zhen, Xiaoling Wang, Weixin Zhang, Xiankai Lu, and Tao Liu. 2022. "Effects of Exotic and Native Earthworms on Soil Micro-Decomposers in a Subtropical Forest: A Field Mesocosm Experiment" Forests 13, no. 11: 1924. https://doi.org/10.3390/f13111924