
Cymbopogon citratus Essential Oil: Extraction, GC–MS, Phytochemical Analysis, Antioxidant Activity, and In Silico Molecular Docking for Protein Targets Related to CNS

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Vegetal Material
2.2. Extraction
2.3. Phytochemical Test
2.4. GC–MS Analyses
2.5. Antioxidant Activity
2.6. In Silico Studies
3. Results and Discussion
3.1. Extraction and Phytochemical Tests
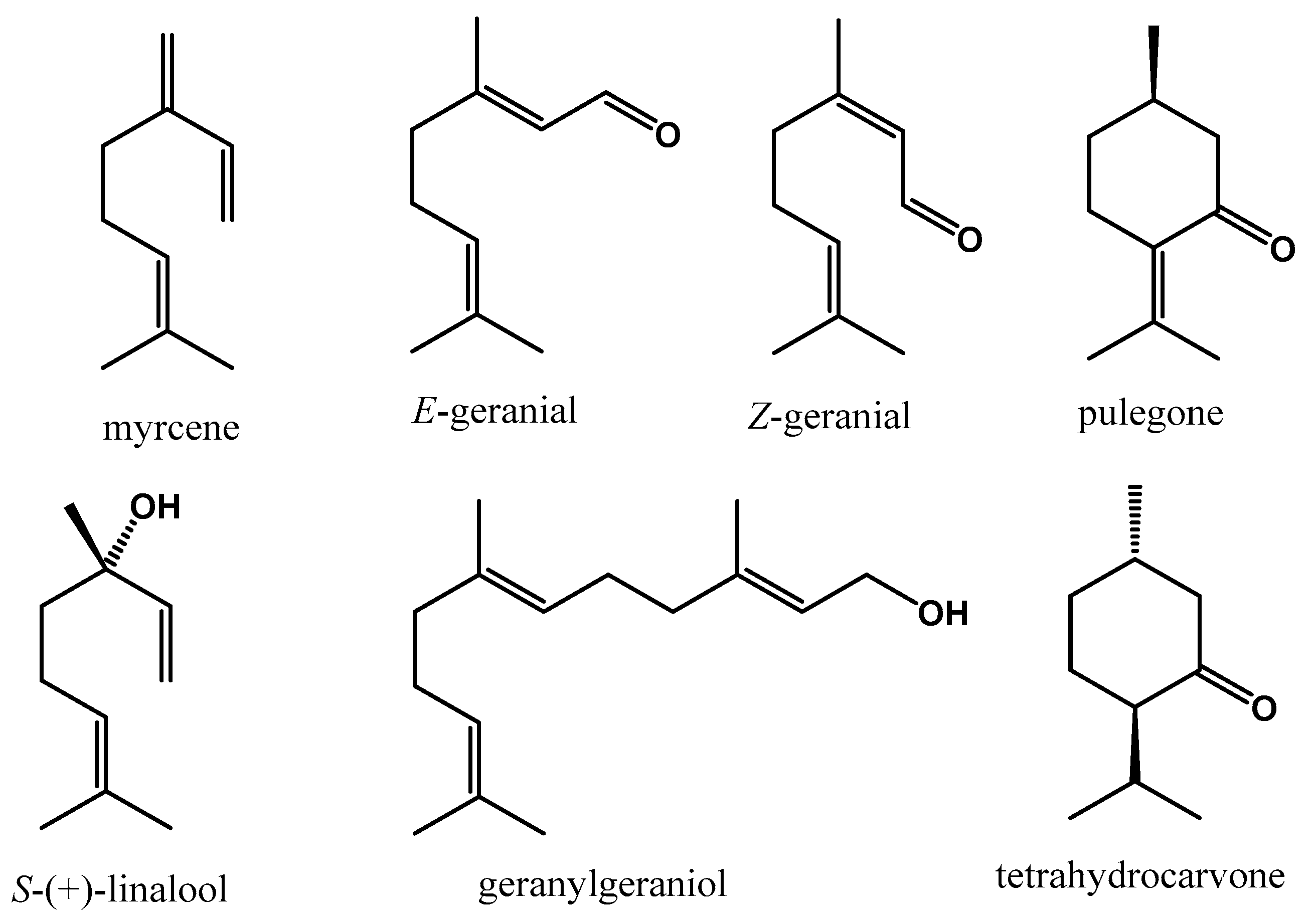
3.2. GC–MS
3.3. Antioxidant Activity
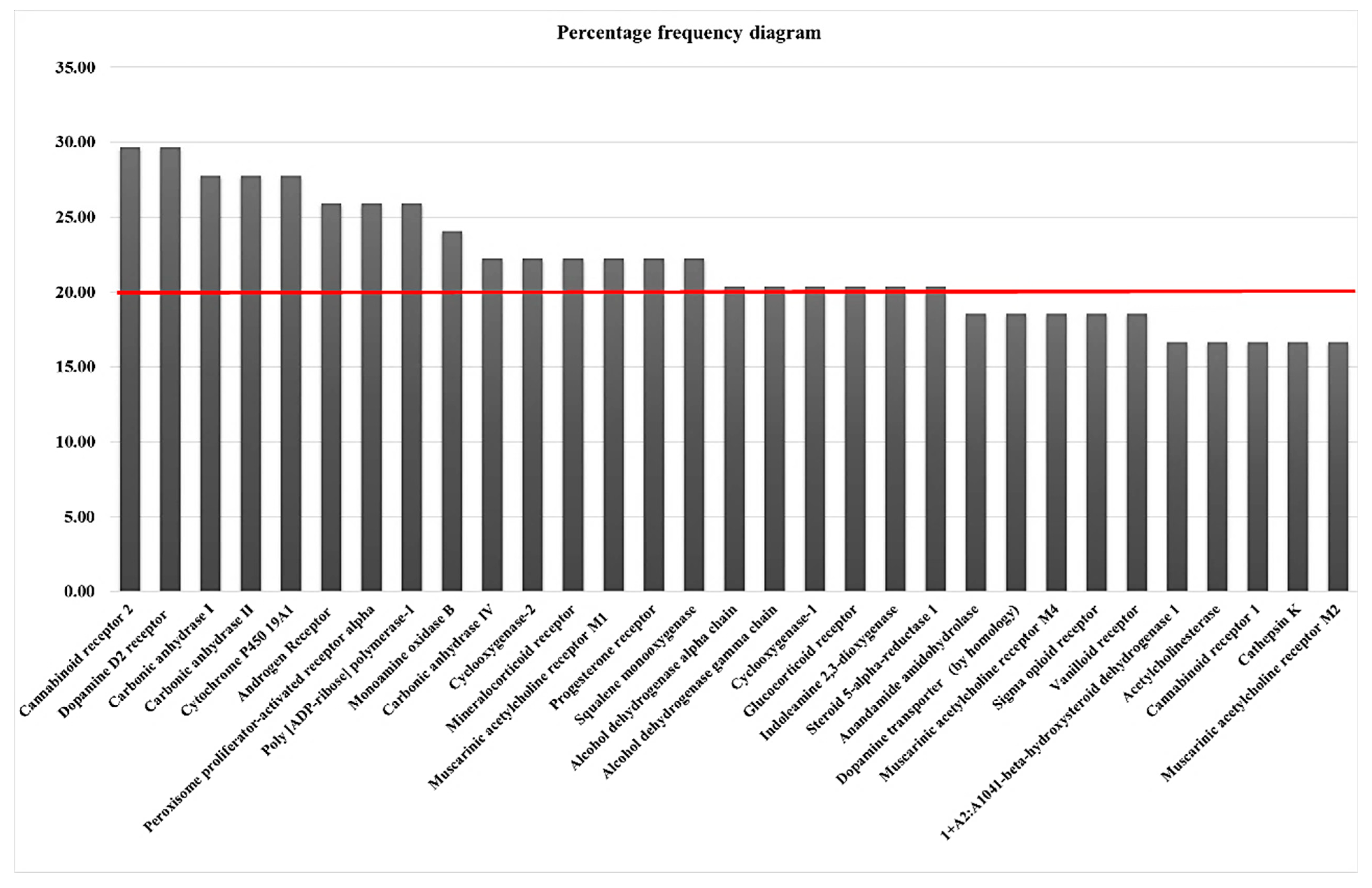
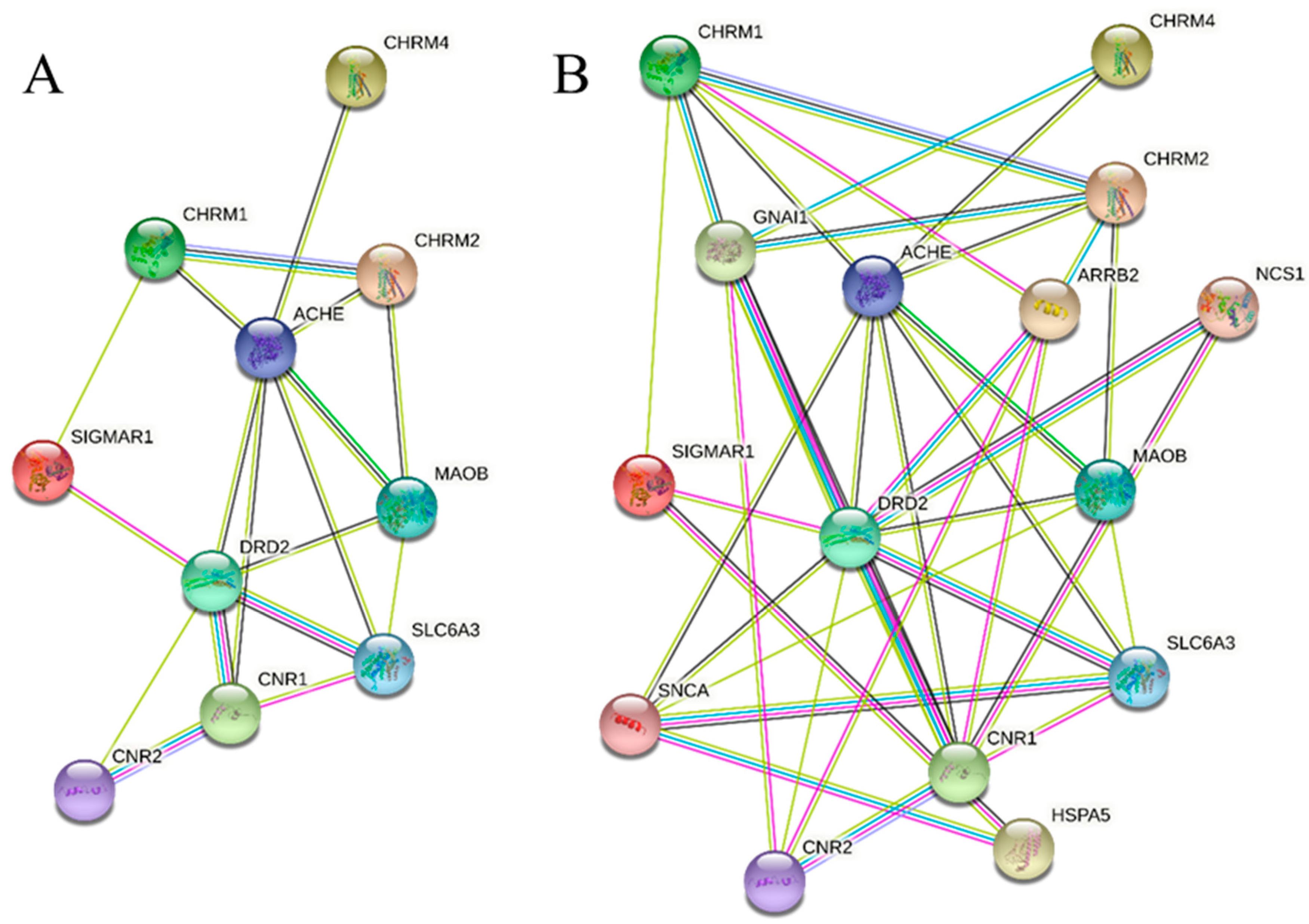
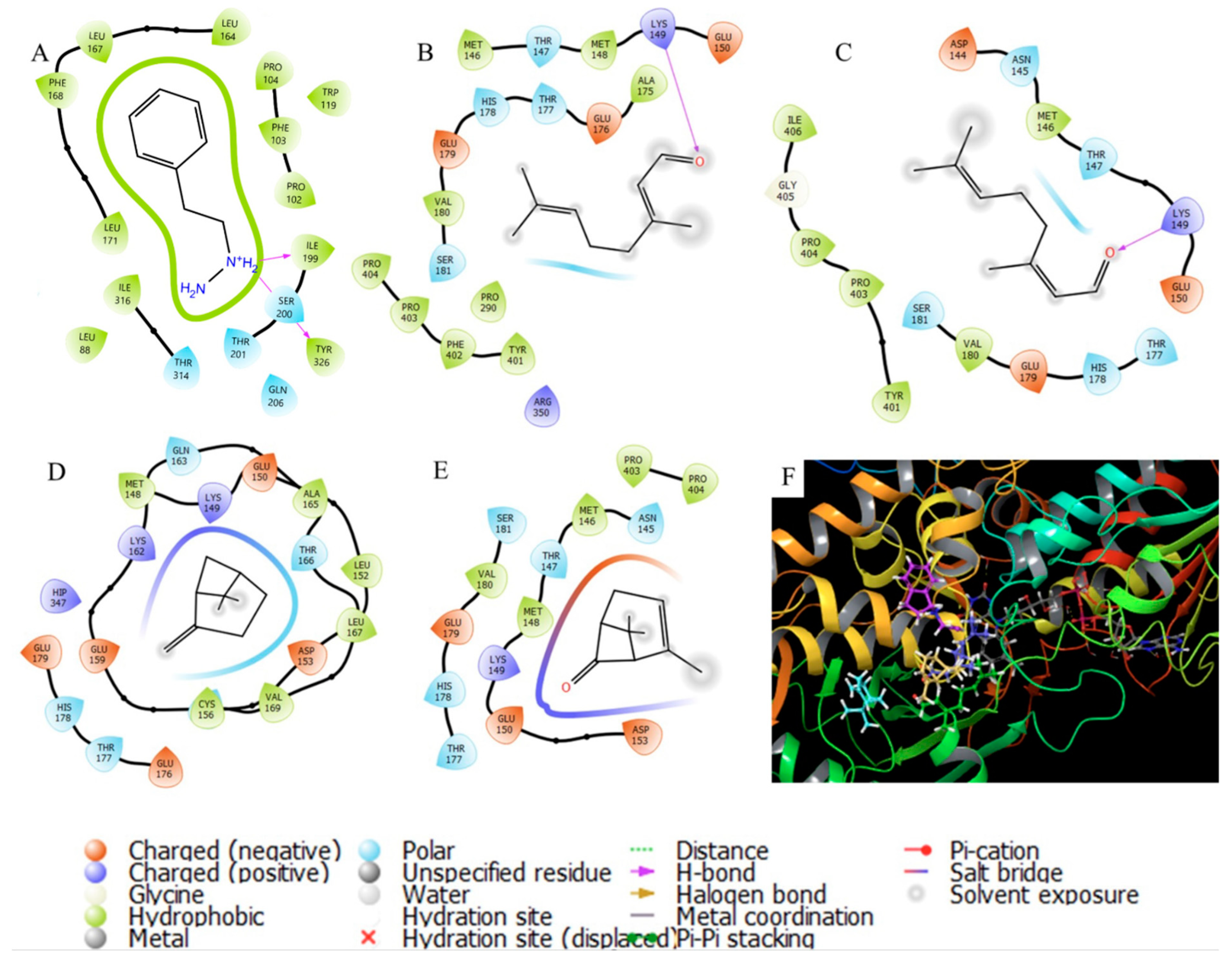
3.4. In Silico Studies

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
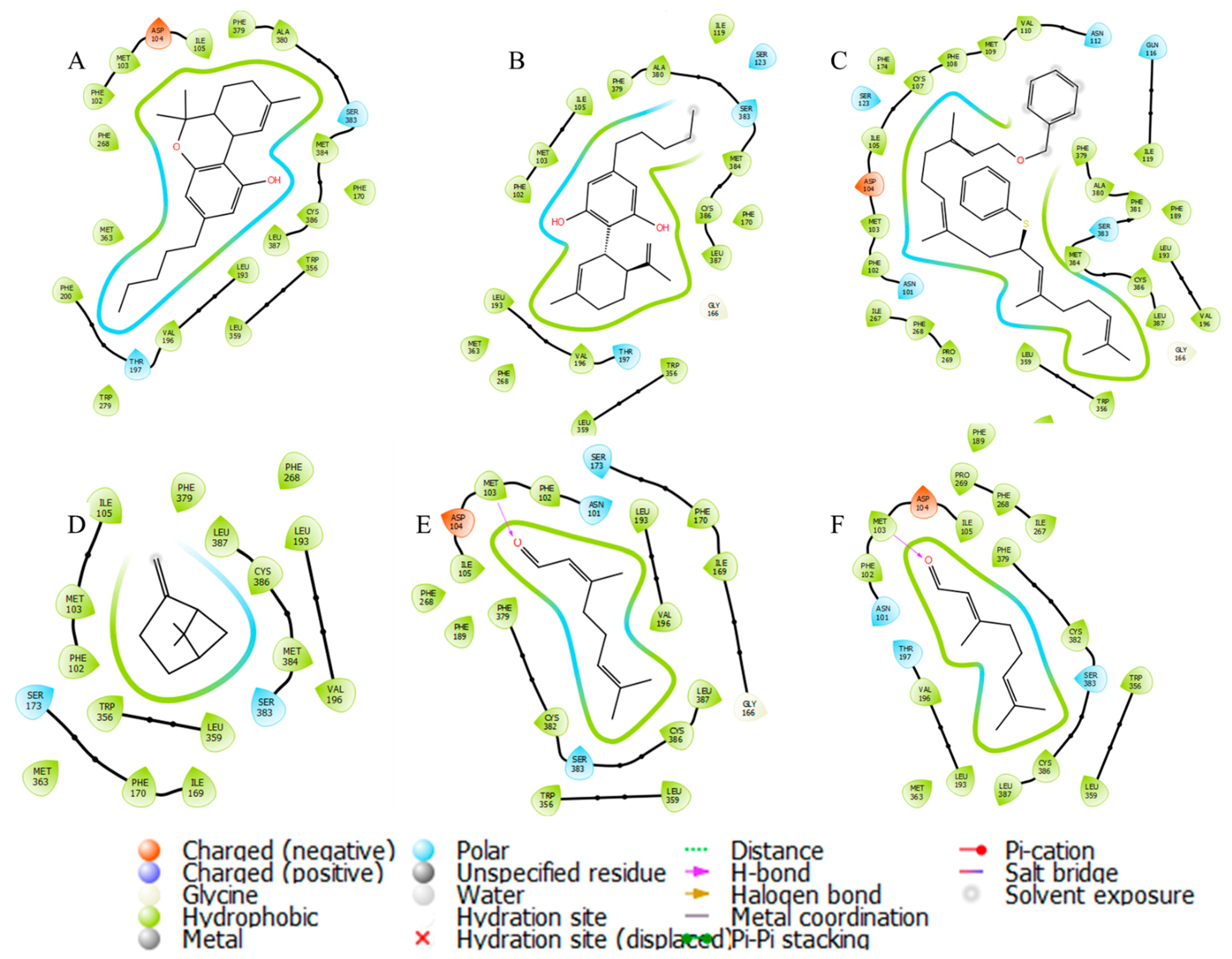{kind=link}
| Target | Frequency (%) | Substrates/Inhibitor | Disease-Related | Biological Functions | Ref. |
|---|---|---|---|---|---|
| Cannabinoid receptor 2 (CNR2) | 29.63 | JWH133, arachidonoyl ethanolamine (AEA) and 2-arachidonoylglycerol (2-AG) | Multiple sclerosis, Alzheimer’s disease (AD) | Neuroinflammation, pain, and anxiety | [57,58] |
| Dopamine D2 receptor (DRD2) | 29.63 | Haloperidol, fluphenazine, chlorpromazine, risperidone, olanzapine, paliperidone, dopamine | Schizophrenia (SCZ) | Cognition, mood, and motor movements | [54,55,56] |
| Monoamine oxidase B (MAOB) | 24.07 | Selegiline, rasagiline | Parkinson’s disease | Dopamine metabolism | [59] |
| Muscarinic acetylcholine receptor M1 (CHRM1) | 22.22 | Xanomeline | Alzheimer’s disease | Learning and memory | [60] |
| Dopamine transporter (SLC6A3) | 18.52 | Dopamine, pramipexole, ropinirole, rotigotine, apomorphine | Parkinson’s disease, bipolar disorder, attention deficit hyperactivity disorder, and dopamine transporter deficiency syndrome. | Translocate dopamine (DA) | [51] |
| Muscarinic acetylcholine receptor M4 (CHRM4) | 18.52 | Pirenzepine, acetylcholine | Alzheimer’s disease, schizophrenia, and drug addiction | Regulation of the transmission | [40,61] |
| Sigma opioid receptor (SIGMAR1) | 18.52 | 1,3 di-O-tolyl guanidine (DTG), haloperidol | Addiction, depression, pain, neurodegenerative conditions, cancer, and amyotrophic lateral sclerosis | Endoplasmic reticulum stress, autophagy, lipid transport, ion channel regulation, cognition, and memory | [52,53] |
| Acetylcholinesterase (ACHE) | 16.67 | Donepezil, galantamine, | Alzheimer’s disease AD, Huntington’s disease, multiple sclerosis, Parkinson’s disease | Development of neuromuscular junctions, thalamocortical connections, axon growth, and apoptosis | [49,50] |
| Cannabinoid receptor 1 (CNR1) | 16.67 | CP55940, JWH-015, WIN55212-2, Arachidonoyl ethanolamine (AEA), 2-Arachidonoylglycerol (2-AG) | Huntington’s disease (HD), multiple sclerosis (MS), Alzheimer’s disease (AD) | Learning, memory, pain, analgesia, anxiety, epilepsy, appetite | [57,62] |
| Muscarinic acetylcholine receptor M2 (CHRM2) | 16.67 | BIBN-99, 3-quinuclidinyl-benzilate, N-methyl scopolamine | Alzheimer’s disease, schizophrenia, Parkinson’s disease, and chronic obstructive pulmonary disease | Cardiovascular function through G-protein-coupled activation | [63,64] |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shah, G.; Shri, R.; Panchal, V.; Sharma, N.; Singh, B.; Mann, A. Scientific basis for the therapeutic use of Cymbopogon citratus, stapf (Lemon grass). J. Adv. Pharm. Technol. Res. 2011, 2, 3. [Google Scholar] [CrossRef]
- Boukhatem, M.N.; Ferhat, M.A.; Kameli, A.; Saidi, F.; Kebir, H.T. Lemon grass (Cymbopogon citratus) essential oil as a potent anti-inflammatory and antifungal drugs. Libyan J. Med. 2014, 9, 25431. [Google Scholar] [CrossRef]
- Pratama, Y.M.; Permana, B.P. Lemongrass (Cymbopogon citratus) Essential Oil Inhibits Candida Albicans Growth In Vitro. Biomedika 2021, 13, 142–147. [Google Scholar] [CrossRef]
- Sharma, A.D.; Kaur, I. Essential oil from Cymbopogon citratus exhibits “anti-aspergillosis” potential: In-silico molecular docking and in vitro studies. Bull. Natl. Res. Cent. 2022, 46, 23. [Google Scholar] [CrossRef]
- Sharma, A.D.; Kaur, I.; Chauhan, A. Anti-aspergillosis and anti-mucormycosis potential of essential oils from two Cymbopogon spp. targeting riboflavin synthesis pathway. Phytomed. Plus. 2003, 3, 100440. [Google Scholar] [CrossRef]
- He, L.-L.; Zhao, Y.; Fan, L.-M.; Zhan, J.-J.; Tao, L.-H.; Yang, Y.-H.; Su, F.-W.; Chen, Q.-B.; Ye, M. In vitro and in vivo antifungal activity of Cymbopogon citrates essential oils from different climate conditions against Botrytis cinerea. Sci. Hortic. 2023, 308, 111544. [Google Scholar] [CrossRef]
- Sawadogo, I.; Paré, A.; Kaboré, D.; Montet, D.; Durand, N.; Bouajila, J.; Zida, E.P.; Sawadogo-Lingani, H.; Nikiéma, P.A.; Nebié, R.H.C.; et al. Antifungal and Antiaflatoxinogenic Effects of Cymbopogon citratus, Cymbopogon nardus, and Cymbopogon schoenanthus Essential Oils Alone and in Combination. J. Fungi 2022, 8, 117. [Google Scholar] [CrossRef]
- Sharma, N.; Sheikh, Z.N.; Alamri, S.; Singh, B.; Kesawat, M.S.; Guleria, S. Chemical Composition, Antibacterial and Combinatorial Effects of the Essential Oils from Cymbopogon spp. and Mentha arvensis with Conventional Antibiotics. Agronomy 2023, 13, 1091. [Google Scholar] [CrossRef]
- Falode, J.A.; Olofinlade, T.B.; Fayeun, G.S.; Adeoye, A.O.; Bamisaye, F.A.; Ajuwon, O.R.; Obafemi, T.O. Free and bound phenols from Cymbopogon citratus mitigated hepatocellular injury in streptozotocin-induced type 1 diabetic male rats via decrease in oxidative stress, inflammation, and other risk markers. Pharmacol. Res. Mod. Chin. Med. 2023, 7, 100234. [Google Scholar] [CrossRef]
- Da Ressurreição, S.; Pedreiro, S.; Batista, M.T.; Figueirinha, A. Effect of Phenolic Compounds from Cymbopogon citratus (DC) Stapf. Leaves on Micellar Solubility of Cholesterol. Molecules 2022, 27, 7338. [Google Scholar] [CrossRef] [PubMed]
- Mukhtar, M.H.; El-Readi, M.Z.; Elzubier, M.E.; Fatani, S.H.; Refaat, B.; Shaheen, U.; Khidir, E.B.A.; Taha, H.H.; Eid, S.Y. Cymbopogon citratus and Citral Overcome Doxorubicin Resistance in Cancer Cells via Modulating the Drug’s Metabolism, Toxicity, and Multidrug Transporters. Molecules 2023, 28, 3415. [Google Scholar] [CrossRef] [PubMed]
- Mouta, L.F.G.L.; Marques, R.S.; Koga-Ito, C.Y.; Salvador, M.J.; Giro, E.M.A.; Brighenti, F.L. Cymbopogon citratus Essential Oil Increases the Effect of Digluconate Chlorhexidine on Microcosm Biofilms. Pathogens 2022, 11, 1067. [Google Scholar] [CrossRef]
- Hacke, A.C.M.; Miyoshi, E.; Marques, J.A.; Pereira, R.P. Cymbopogon citratus (DC.) Stapf, citral and geraniol exhibit anticonvulsant and neuroprotective effects in pentylenetetrazole-induced seizures in zebrafish. J. Ethnopharmacol. 2021, 275, 114142. [Google Scholar] [CrossRef] [PubMed]
- Madi, Y.F.; Choucry, M.A.; El-Marasy, S.A.; Meselhy, M.R.; El-Kashoury, E.-S.A. UPLC-Orbitrap HRMS metabolic profiling of Cymbopogon citratus cultivated in Egypt; neuroprotective effect against AlCl3-induced neurotoxicity in rats. J. Ethnopharmacol. 2020, 259, 112930. [Google Scholar] [CrossRef] [PubMed]
- de Andrade, S.F.; Rocha, C.; Pinheiro, E.J.; Pereira-Leite, C.; Costa, M.D.C.; Rodrigues, L.M. Revealing the Protective Effect of Topically Applied Cymbopogon citratus Essential Oil in Human Skin through A Contact Model. Cosmetics 2023, 10, 29. [Google Scholar] [CrossRef]
- Gaspar, A.L.; Gaspar, A.B.; Contini, L.R.F.; Silva, M.F.; Chagas, E.G.L.; Bahú, J.O.; Concha, V.O.C.; Carvalho, R.A.; Severino, P.; Souto, E.B.; et al. Lemongrass (Cymbopogon citratus)-incorporated chitosan bioactive films for potential skincare applications. Int. J. Pharm. 2022, 628, 122301. [Google Scholar] [CrossRef]
- Li, C.; Luo, Y.; Zhang, W.; Cai, Q.; Wu, X.; Tan, Z.; Chen, R.; Chen, Z.; Wang, S.; Zhang, L. A comparative study on chemical compositions and biological activities of four essential oils: Cymbopogon citratus (DC.) Stapf, Cinnamomum cassia (L.) Presl, Salvia japonica Thunb. and Rosa rugosa Thunb. J. Ethnopharmacol. 2021, 280, 114472. [Google Scholar] [CrossRef]
- de Oliveira Alencar, D.D.; de Souza, E.L.; da Cruz Almeida, E.T.; da Silva, A.L.; Oliveira, H.M.L.; Cavalcanti, M.T. Microencapsulation of Cymbopogon citratus D.C. Stapf Essential Oil with Spray Drying: Development, Characterization, and Antioxidant and Antibacterial Activities. Foods 2022, 11, 1111. [Google Scholar] [CrossRef]
- Pino, J.A.; Fon-Fay, F.M.; Pérez, J.C. Chemical Composition and Biological Activities of Essential Oil from Lemongrass (Cympopogon citratus [D.C.] Stapf.) Leaves Grown in Amazonian Ecuador; Centro Nacional de Investigaciones Científicas, Cuba: La Habana, Cuba, 2018; Volume 49. [Google Scholar]
- Tognolini, M.; Barocelli, E.; Ballabeni, V.; Bruni, R.; Bianchi, A.; Chiavarini, M. Impicciatore, Comparative screening of plant essential oils: Phenylpropanoid moiety as basic core for antiplatelet activity. Life Sci. 2006, 78, 1419–1432. [Google Scholar] [CrossRef]
- Carlini, E.A.; Contar, J.D.D.; Silva-Filho, A.R.; Da Silveira-Filho, N.G.; Frochtengarten, M.L.; Bueno, O.F. Pharmacology of lemongrass (Cymbopogon citratus Stapf). I. Effects of teas prepared from the leaves on laboratory animals. J. Ethnopharmacol. 1986, 17, 37–64. [Google Scholar] [CrossRef]
- Tayeboon, G.S.; Tavakoli, F.; Hassani, S.; Khanavi, M.; Sabzevari, O.; Ostad, S.N. Effects of Cymbopogon citratus and Ferula assa-foetida extracts on glutamate-induced neurotoxicity. In Vitro Cell. Dev. Biol. Anim. 2013, 49, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Bhandary, S.K.; Bhat, V.S.; Sharmila, K.P.; Bekal, M.P. Preliminary phytochemical screening of various extracts of Punica granatum peel, whole fruit and seeds. J. Health Allied Sci. NU 2012, 2, 34–38. [Google Scholar] [CrossRef]
- Bulugahapitiya, V.P. Plants Based Natural Products; University of Ruhuna: Fribourg, Switzerland, 2013. [Google Scholar]
- Marami, L.M.; Dilba, G.M.; Babele, D.A.; Sarba, E.J.; Gizaw, A.; Bune, W.M.; Bayu, M.D.; Admasu, P.; Mekbeb, A.; Tadese, M.; et al. Phytochemical Screening and in-vitro Evaluation of Antibacterial Activities of Echinops amplexicaulis, Ruta chalepensis and Salix subserrata Against Selected Pathogenic Bacterial Strains in West Shewa Zone, Ethiopia. J. Exp. Pharmacol. 2021, 13, 511–520. [Google Scholar] [CrossRef]
- Müller, L.; Fröhlich, K.; Böhm, V. Comparative antioxidant activities of carotenoids measured by ferric reducing antioxidant power (FRAP), ABTS bleaching assay (αTEAC), DPPH assay and peroxyl radical scavenging assay. Food Chem. 2011, 129, 139–148. [Google Scholar] [CrossRef]
- Shimaa, S.M.; Sohair, F.; Amel, S. A Coelomic Fluid of Allolobophora caliginosa as Novel Prospects for Medicinal Antioxidants, Anti-inflammatory, Antiproliferative, Analgesics, and Antipyretics. Biointerface Res. Appl. Chem. 2022, 13, 209. [Google Scholar] [CrossRef]
- Lucas, B.N.; Nora, D.; Boeira, C.P.; Verruck, S.; da Rosa, C.S. Determination of total phenolic compounds in plant extracts via Folin-Ciocalteu’s method adapted to the usage of digital images. Food Sci. Technol. 2022, 42, e35122. [Google Scholar] [CrossRef]
- Xie, Y.; Gong, T.; Liu, H.; Fan, Z.; Chen, Z.; Liu, X. In Vitro and In Vivo Digestive Fate and Antioxidant Activities of Polyphenols from Hulless Barley: Impact of Various Thermal Processing Methods and β-Glucan. J. Agric. Food Chem. 2022, 70, 7683–7694. [Google Scholar] [CrossRef] [PubMed]
- Berker, K.I.; Olgun, F.A.O.; Ozyurt, D.; Demirata, B. Apak, Modified Folin–Ciocalteu Antioxidant Capacity Assay for Measuring Lipophilic Antioxidants. J. Agric. Food Chem. 2013, 61, 4783–4791. [Google Scholar] [CrossRef]
- Zhang, A. Sulphite-corrected, non-phenolic and phenolic antioxidant capacities of fruit wines profiled by differential Folin-Ciocalteu assay. Int. J. Food Sci. Technol. 2022, 57, 1259–1272. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef] [Green Version]
- Tan, T.C.; Spadiut, O.; Gandini, R.; Haltrich, D.; Divne, C. Structural Basis for Binding of Fluorinated Glucose and Galactose to Trametes multicolor Pyranose 2-Oxidase Variants with Improved Galactose Conversion. PLoS ONE 2014, 9, e86736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, T.; Vemuri, K.; Pu, M.; Qu, L.; Han, G.W.; Wu, Y.; Zhao, S.; Shui, W.; Li, S.; Korde, A. Crystal Structure of the Human Cannabinoid Receptor CB1. Cell 2016, 167, 750–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Hua, T.; Vemuri, K.; Ho, J.-H.; Wu, Y.; Wu, L.; Popov, P.; Benchama, O.; Zvonok, N.; Locke, K. Crystal Structure of the Human Cannabinoid Receptor CB2. Cell 2019, 176, 459–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, L.; Tan, L.; Chen, Z.; Qi, J.; Nie, F.; Luo, Z.; Cheng, J.; Wang, S. Haloperidol bound D2 dopamine receptor structure inspired the discovery of subtype selective ligands. Nat. Commun. 2020, 11, 1074. [Google Scholar] [CrossRef] [Green Version]
- Iacovino, L.G.; Reis, J.; Mai, A.; Binda, C.; Mattevi, A. Diphenylene Iodonium Is a Noncovalent MAO Inhibitor: A Biochemical and Structural Analysis. ChemMedChem 2020, 15, 1394–1397. [Google Scholar] [CrossRef]
- Brown, A.J.H.; Bradley, S.J.; Marshall, F.H.; Brown, G.A.; Bennett, K.A.; Brown, J.; Cansfield, J.E.; Cross, D.M.; de Graaf, C.; Hudson, B.D. From structure to clinic: Design of a muscarinic M1 receptor agonist with the potential to treat Alzheimer’s disease. Cell 2021, 184, 5886–5901. [Google Scholar] [CrossRef]
- Suno, R.; Lee, S.; Maeda, S.; Yasuda, S.; Yamashita, K.; Hirata, K.; Horita, S.; Tawaramoto, M.S.; Tsujimoto, H.; Murata, T. Structural insights into the subtype-selective antagonist binding to the M2 muscarinic receptor. Nat. Chem. Biol. 2018, 14, 1150–1158. [Google Scholar] [CrossRef]
- Thal, D.M.; Sun, B.; Feng, D.; Nawaratne, V.; Leach, K.; Felder, C.C.; Bures, M.G.; Evans, D.A.; Weis, W.I.; Bachhawat, P. Crystal structures of the M1 and M4 muscarinic acetylcholine receptors. Nature 2016, 531, 335–340. [Google Scholar] [CrossRef] [Green Version]
- Sastry, G.M.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput. Aided Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef]
- In silico Study of Coumarins Derivatives with Potential Use in Systemic Diseases. Biointerface Res. Appl. Chem. 2022, 13, 240. [CrossRef]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra Precision Glide: Docking and Scoring Incorporating a Model of Hydrophobic Enclosure for Protein−Ligand Complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekpenyong, C.E.; Akpan, E.E. Use of Cymbopogon citratus essential oil in food preservation: Recent advances and future perspectives. Crit. Rev. Food Sci. Nutr. 2017, 57, 2541–2559. [Google Scholar] [CrossRef] [PubMed]
- Gbenou, J.D.; Ahounou, J.F.; Akakpo, H.B.; Laleye, A.; Yayi, E.; Gbaguidi, F.; Baba-Moussa, L.; Darboux, R.; Dansou, P.; Moudachirou, M. Phytochemical composition of Cymbopogon citratus and Eucalyptus citriodora essential oils and their anti-inflammatory and analgesic properties on Wistar rats. Mol. Biol. Rep. 2013, 40, 1127–1134. [Google Scholar] [CrossRef]
- Brügger, B.P.; Martínez, L.C.; Plata-Rueda, A.; Castro, B.M.D.C.E.; Soares, M.A.; Wilcken, C.F.; Carvalho, A.G.; Serrão, J.E.; Zanuncio, J.C. Bioactivity of the Cymbopogon citratus (Poaceae) essential oil and its terpenoid constituents on the predatory bug, Podisus nigrispinus (Heteroptera: Pentatomidae). Sci. Rep. 2019, 9, 8358. [Google Scholar] [CrossRef] [Green Version]
- Casabianca, H.; Graff, J.B.; Faugier, V.; Fleig, F.; Grenier, C. Enantiomeric Distribution Studies of Linalool and Linalyl Acetate. A Powerful Tool for Authenticity Control of Essential Oils. J. High Resolut. Chromatogr. 1998, 21, 107–112. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Y.; Liu, X.; Li, J.; Zhang, J.; Liu, D. Chemical constituents and pharmacological activities of medicinal plants from Rosa genus. Chin. Herb. Med. 2022, 14, 187–209. [Google Scholar] [CrossRef]
- Marucci, G.; Buccioni, M.; Ben, D.D.; Lambertucci, C.; Volpini, R.; Amenta, F. Efficacy of acetylcholinesterase inhibitors in Alzheimer’s disease. Neuropharmacology 2021, 190, 108352. [Google Scholar] [CrossRef]
- Walczak-Nowicka, Ł.J.; Herbet, M. Acetylcholinesterase Inhibitors in the Treatment of Neurodegenerative Diseases and the Role of Acetylcholinesterase in their Pathogenesis. Int. J. Mol. Sci. 2021, 22, 9290. [Google Scholar] [CrossRef]
- Challasivakanaka, S.; Zhen, J.; Smith, M.E.; Reith, M.E.A.; Foster, J.D.; Vaughan, R.A. Dopamine transporter phosphorylation site threonine 53 is stimulated by amphetamines and regulates dopamine transport, efflux, and cocaine analog binding. J. Biol. Chem. 2017, 292, 19066–19075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aydar, E.; Cobos, E.J.; Maurice, T.; Murell-Lagnado, R.D.; Safrany, S.T. Editorial: Sigma Receptors. Front. Pharmacol. 2002, 11, 590519. [Google Scholar] [CrossRef]
- Aishwarya, R.; Abdullah, C.S.; Morshed, M.; Remex, N.S.; Bhuiyan, M.S. Sigmar1′s Molecular, Cellular, and Biological Functions in Regulating Cellular Pathophysiology. Front. Physiol. 2021, 12, 705575. [Google Scholar] [CrossRef] [PubMed]
- Puig, M.V.; Miller, E.K. Neural Substrates of Dopamine D2 Receptor Modulated Executive Functions in the Monkey Prefrontal Cortex. Cereb. Cortex 2015, 25, 2980–2987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitton, A.E.; Green, A.I.; Pizzagalli, D.A.; Roth, R.M.; Williams, J.M.; Brunette, M.F. Potent Dopamine D2 Antagonists Block the Reward-Enhancing Effects of Nicotine in Smokers with Schizophrenia. Schizophr. Bull. 2019, 45, 1300–1308. [Google Scholar] [CrossRef]
- Hussain, S.; Siddiqui, S.A.; Mondal, S.; Millat, M.S.; Marzan, S.; Uddin, M.G.; Aziz, M.A.; Alam, M.F.; Islam, M.S. Association of DRD2 gene polymorphisms with schizophrenia in the young Bangladeshi population: A pilot study. Heliyon 2020, 6, e05125. [Google Scholar] [CrossRef]
- Kendall, D.A.; Yudowski, G.A. Cannabinoid Receptors in the Central Nervous System: Their Signaling and Roles in Disease. Front. Cell. Neurosci. 2017, 10, 294. [Google Scholar] [CrossRef] [Green Version]
- Bie, B.; Wu, J.; Foss, J.F.; Naguib, M. An overview of the cannabinoid type 2 receptor system and its therapeutic potential. Curr. Opin. Anaesthesiol. 2018, 31, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Rathitharan, G.; Meyer, J.H.; Furukawa, Y.; Ang, L.-C.; Boileau, I.; Guttman, M.; Hornykiewicz, O.; Kish, S.J. Brain monoamine oxidase B and A in human parkinsonian dopamine deficiency disorders. Brain 2017, 140, 2460–2474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarpa, M.; Molloy, C.; Jenkins, L.; Strellis, B.; Budgett, R.F.; Hesse, S.; Dwomoh, L.; Marsango, S.; Tejeda, G.S.; Rossi, M. Biased M1 muscarinic receptor mutant mice show accelerated progression of prion neurodegenerative disease. Proc. Natl. Acad. Sci. USA 2021, 118, e2107389118. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.L.; Yeh, H.H. Neurotransmitters and neurotransmission in the developing and adult nervous system. In Conn’s Translational Neuroscience; Academic Press: San Diego, CA, USA, 2017; pp. 49–84. [Google Scholar]
- Njoo, C.; Agarwal, N.; Lutz, B.; Kuner, R. The Cannabinoid Receptor CB1 Interacts with the WAVE1 Complex and Plays a Role in Actin Dynamics and Structural Plasticity in Neurons. PLoS Biol. 2015, 13, e1002286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haga, K.; Kruse, A.C.; Asada, H.; Yurugi-Kobayashi, T.; Shiroishi, M.; Zhang, C.; Weis, W.I.; Okada, T.; Kobilka, B.K.; Haga, T. Structure of the human M2 muscarinic acetylcholine receptor bound to an antagonist. Nature 2012, 482, 547–551. [Google Scholar] [CrossRef] [Green Version]
- Svoboda, J.; Popelikova, A.; Stuchlik, A. Drugs Interfering with Muscarinic Acetylcholine Receptors and Their Effects on Place Navigation. Front. Psychiatry 2017, 8, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabriel, S.M.; Davidson, M.; Haroutunian, V.; Powchik, P.; Bierer, L.M.; Purohit, D.P.; Perl, D.P.; Davis, K.L. Neuropeptide Deficits in Schizophrenia VS. Alzheimer’s Disease Cerebral Cortex. Biol. Psychiatry 1996, 39, 82–91. [Google Scholar] [CrossRef] [PubMed]
- AAT Bioquest, Inc. Quest Database™ Muscarinic Acetylcholine Receptor M4 Inhibitors (IC50, Ki). Available online: https://www.aatbio.com/data-sets/muscarinic-acetylcholine-receptor-m4-inhibitors-ic50-ki (accessed on 22 May 2023).







| Secondary Metabolite Group | Fresh Vegetal Material | Essential Oil |
|---|---|---|
| Phenolic hydroxyls | - | - |
| Unsaturation | ++ | +++ |
| Alkaloids | + | - |
| Steroids and terpenes | +++ | + |
| Carbonyl groups | ++ | +++ |
| Sugars | +++ | + |
| Anthocyanins | ++ | - |
| Concentration (µL EO/mL) | DPPH (%) |
|---|---|
| 150 | 76.30 ± 1.23 |
| 100 | 72.15 ± 2.62 |
| 75 | 59.58 ± 2.25 |
| 50 | 52.11 ± 1.98 |
| 35 | 18.16 ± 3.15 |
| 25 | 8.66 ± 0.87 |
| 10 | 0.0 ± 0.0 |
| RT (min) | Name | Abundance | AChE | CBR2 | DR2 | CBR1 | M1 | M2 | M4 | MAOB |
|---|---|---|---|---|---|---|---|---|---|---|
| 7.75 | β-Myrcene | 8.76 | −4.059 | −6.017 | −5.717 | −5.840 | −6.297 | −7.025 | −6.741 | −2.987 |
| 7.87 | 4-Carene | 0.40 | −4.456 | −6.637 | −5.897 | −6.341 | −6.787 | −7.040 | −7.276 | −3.390 |
| 8.56 | β-Pinene | 1.94 | −4.709 | −6.193 | −5.459 | −5.827 | −6.163 | −6.762 | −6.931 | −3.173 |
| 9.07 | α-Ocimene | 0.40 | −1.969 | −3.778 | −3.422 | −3.315 | −3.196 | −4.042 | −3.971 | −0.510 |
| 9.18 | β-Ocimene | 0.40 | −2.376 | −4.469 | −4.005 | −4.170 | −3.947 | −4.691 | −3.841 | −0.932 |
| 9.41 | Seudenone | 0.07 | −4.584 | −5.897 | −5.906 | −4.940 | −6.193 | −6.324 | −6.312 | −3.512 |
| 10.42 | 1,2,3,3a,4,6a-hexahydropentalene | 0.20 | −4.013 | −5.900 | −5.607 | −5.511 | −6.106 | −6.403 | −6.428 | −2.949 |
| 16.89 | d-Linalool | 1.24 | −2.649 | −4.713 | −3.538 | −4.894 | −4.478 | −4.332 | −4.822 | −1.569 |
| 11.52 | Alloocimene | 0.38 | −3.153 | −5.191 | −4.324 | −5.478 | −5.089 | −5.323 | −5.404 | −1.780 |
| 12.05 | 3,3,5-Trimethyl-1,4-hexadiene | 0.31 | −2.900 | −4.449 | −4.072 | −4.109 | −4.443 | −4.905 | −4.628 | −1.855 |
| 12.27 | 3,5-Dimethyl-1,6-heptadiene | 0.80 | −1.027 | −2.954 | −2.231 | −1.742 | −2.830 | −2.724 | −3.042 | 0.212 |
| 12.69 | Ethenylcyclohexane | 0.81 | −4.425 | −5.844 | −5.079 | −5.891 | −6.138 | −6.510 | −6.508 | −3.306 |
| 12.96 | 5-Dodecyne | 0.98 | 1.056 | −0.991 | −0.398 | 0.796 | 0.173 | −1.252 | −1.262 | 3.277 |
| 13.29 | Pulegone | 1.00 | −4.594 | −6.959 | −6.407 | −6.483 | −6.581 | −7.366 | −7.774 | −3.612 |
| 13.56 | Tetrahydrocarvone | 1.73 | −4.813 | −6.692 | −5.646 | −5.456 | −6.533 | −6.429 | −7.075 | −3.061 |
| 15.76 | Z-Geranial | 27.58 | −2.263 | −4.050 | −4.437 | −4.384 | −4.935 | −4.555 | −4.756 | −1.062 |
| 17.7 | E-Geranial | 38.62 | −2.787 | −4.535 | −4.359 | −5.533 | −4.187 | −4.679 | −4.793 | −1.283 |
| 18.18 | 3-Ethoxybenzenamine | 1.23 | −4.391 | −5.220 | −5.261 | −5.832 | −5.407 | −5.257 | −5.438 | −3.109 |
| 18.37 | Grandlure IV | 0.22 | −4.029 | −6.262 | −5.123 | −5.166 | −5.782 | −7.163 | −6.447 | −2.975 |
| 18.73 | Carvotanacetone | 0.48 | −5.141 | −7.020 | −6.368 | −6.551 | −7.282 | −7.194 | −7.761 | −3.574 |
| 18.99 | Ethyl nerate | 0.29 | −3.835 | −5.454 | −4.512 | −6.495 | −4.719 | −5.766 | −5.264 | −1.607 |
| 19.13 | Verbenyl ethyl ether | 0.74 | −4.573 | −6.224 | −4.924 | −5.807 | −6.031 | −7.097 | −7.071 | −2.783 |
| 19.75 | 2,3-Dimethyl-3-buten-2-ol | 0.76 | −3.739 | −4.383 | −3.571 | −4.536 | −4.426 | −4.825 | −4.758 | −2.254 |
| 20.26 | (Z)-2-Butenoic acid, methyl ester, | 1.33 | −3.615 | −3.541 | −3.838 | −3.949 | −4.181 | −3.854 | −4.308 | −2.088 |
| 20.62 | Dehydrolinalool | 0.22 | −2.662 | −4.383 | −3.847 | −4.739 | −4.288 | −4.654 | −4.763 | −2.590 |
| 20.85 | 2-Butenoic acid, hexyl ester | 0.77 | 1.204 | −0.796 | −0.220 | −1.109 | 0.259 | −0.893 | −0.726 | 2.892 |
| 21.03 | Chrysanthenone | 0.30 | −5.398 | −6.865 | −4.983 | −6.817 | −6.342 | −7.531 | −7.678 | −3.517 |
| 21.12 | Safranal | 0.30 | −5.559 | −6.299 | −5.963 | −5.861 | −6.250 | −7.021 | −7.470 | −3.710 |
| 21.47 | Butyl crotonate | 0.61 | −2.258 | −3.482 | −2.791 | −3.802 | −3.056 | −3.889 | −4.022 | −0.556 |
| 21.83 | (Z)-3,7-Dimethyl-2,6-octadienal | 0.08 | −2.263 | −4.050 | −4.437 | −4.384 | −4.935 | −4.555 | −4.756 | −1.062 |
| 23.12 | 2-Tridecanone | 0.81 | 0.890 | −1.035 | −0.113 | −1.947 | −0.851 | −1.690 | −1.094 | 2.938 |
| 25.68 | Caryophyllene oxide | 0.07 | −4.565 | −6.944 | −4.517 | −6.219 | −3.925 | −8.261 | −7.851 | −2.936 |
| 26.67 | α-Guajene | 0.09 | −5.065 | −7.370 | −6.424 | −6.932 | −6.372 | −8.089 | −8.295 | −3.151 |
| 28.64 | 2-Pentadecanone | 0.05 | 0.437 | −1.584 | 0.262 | −2.187 | −0.710 | −1.950 | −1.508 | 2.572 |
| 33.72 | 2,2,3,5,6-Pentamethyl-3-heptene | 0.03 | −4.172 | −6.157 | −5.614 | −5.976 | −5.908 | −6.434 | −6.610 | −1.934 |
| 37.31 | 1-Benzyloxy-9-(phenylthio)-3,7,11,15-tetramethyl-2,6,10,14-hexadecatetraene | 0.22 | −3.920 | −10.474 | −4.460 | −9.091 | −7.890 | −9.524 | −8.581 | −1.828 |
| 37.94 | Geranylgeraniol | 1.56 | −2.125 | −4.829 | −4.066 | −4.879 | −4.051 | −4.577 | −4.736 | −0.007 |
| 38.36 | 2-cis-Geranylgeraniol | 0.27 | −2.250 | −4.800 | −3.307 | −5.289 | −4.246 | −4.117 | −4.591 | 1.017 |
| 38.6 | 2,6-Dimethyl-6-(4-methyl-3- pentenyl)-2-cyclohexene- 1-carboxaldehyde | 0.48 | −4.219 | −6.127 | −5.912 | −6.670 | −6.351 | −7.792 | −7.059 | −2.108 |
| 38.76 | Geranyllinalool | 0.52 | −1.561 | −4.981 | −3.221 | −5.242 | −4.102 | −4.056 | −5.000 | 0.133 |
| 39.18 | 2,6,10,15,19,23-Hexamethyl- 2,6,10,14,18,22-tetracosahexaene (all-E) | 1.39 | −4.129 | −8.479 | −5.427 | −7.232 | −6.022 | −7.048 | −1.266 | |
| 39.47 | 2,6,10,14,18-Pentamethyl-2,6,10,14,18-eicosapentaene | 0.85 | −4.031 | −8.025 | −3.890 | −7.795 | −5.945 | −4.998 | −7.618 | −1.129 |
| 40.05 | Peruviol | 0.26 | −1.133 | −4.067 | −1.594 | −3.935 | −2.811 | −3.895 | −4.044 | 1.063 |
| 40.24 | Lavandulol | 0.13 | −2.795 | −4.559 | −4.117 | −4.453 | −4.722 | −5.130 | −5.083 | −2.489 |
| 41.14 | 2-Methyl-2-(4-methyl-3-pentenyl)cyclopropanemethanol | 0.11 | −3.923 | −5.160 | −4.973 | −5.131 | −6.096 | −5.762 | −5.946 | −2.383 |
| 41.97 | 4,8-Dimethyl-3,7-nonadien-2-ol | 0.06 | −4.114 | −5.556 | −4.824 | −5.864 | −5.379 | −5.578 | −5.993 | −2.674 |
| - | ligand/substratum | * | −3.934 | −7.224 | −8.16 | −5.107 | −4.19 | −4.484 | −6.04 | - |
| - | blocker/inhibitor | + | −5.459 | −8.085 | −9.27 | −7.686 | −4.613 | −9.538 | −4.665 | −5.766 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortes-Torres, A.G.; López-Castillo, G.N.; Marín-Torres, J.L.; Portillo-Reyes, R.; Luna, F.; Baca, B.E.; Sandoval-Ramírez, J.; Carrasco-Carballo, A. Cymbopogon citratus Essential Oil: Extraction, GC–MS, Phytochemical Analysis, Antioxidant Activity, and In Silico Molecular Docking for Protein Targets Related to CNS. Curr. Issues Mol. Biol. 2023, 45, 5164-5179. https://doi.org/10.3390/cimb45060328
Cortes-Torres AG, López-Castillo GN, Marín-Torres JL, Portillo-Reyes R, Luna F, Baca BE, Sandoval-Ramírez J, Carrasco-Carballo A. Cymbopogon citratus Essential Oil: Extraction, GC–MS, Phytochemical Analysis, Antioxidant Activity, and In Silico Molecular Docking for Protein Targets Related to CNS. Current Issues in Molecular Biology. 2023; 45(6):5164-5179. https://doi.org/10.3390/cimb45060328
Chicago/Turabian StyleCortes-Torres, Ana G., Guiee N. López-Castillo, Josefina L. Marín-Torres, Roberto Portillo-Reyes, Felix Luna, Beatriz E. Baca, Jesús Sandoval-Ramírez, and Alan Carrasco-Carballo. 2023. "Cymbopogon citratus Essential Oil: Extraction, GC–MS, Phytochemical Analysis, Antioxidant Activity, and In Silico Molecular Docking for Protein Targets Related to CNS" Current Issues in Molecular Biology 45, no. 6: 5164-5179. https://doi.org/10.3390/cimb45060328