
The Role of Mesenchymal Stromal Cells and Their Products in the Treatment of Injured Spinal Cords

Abstract
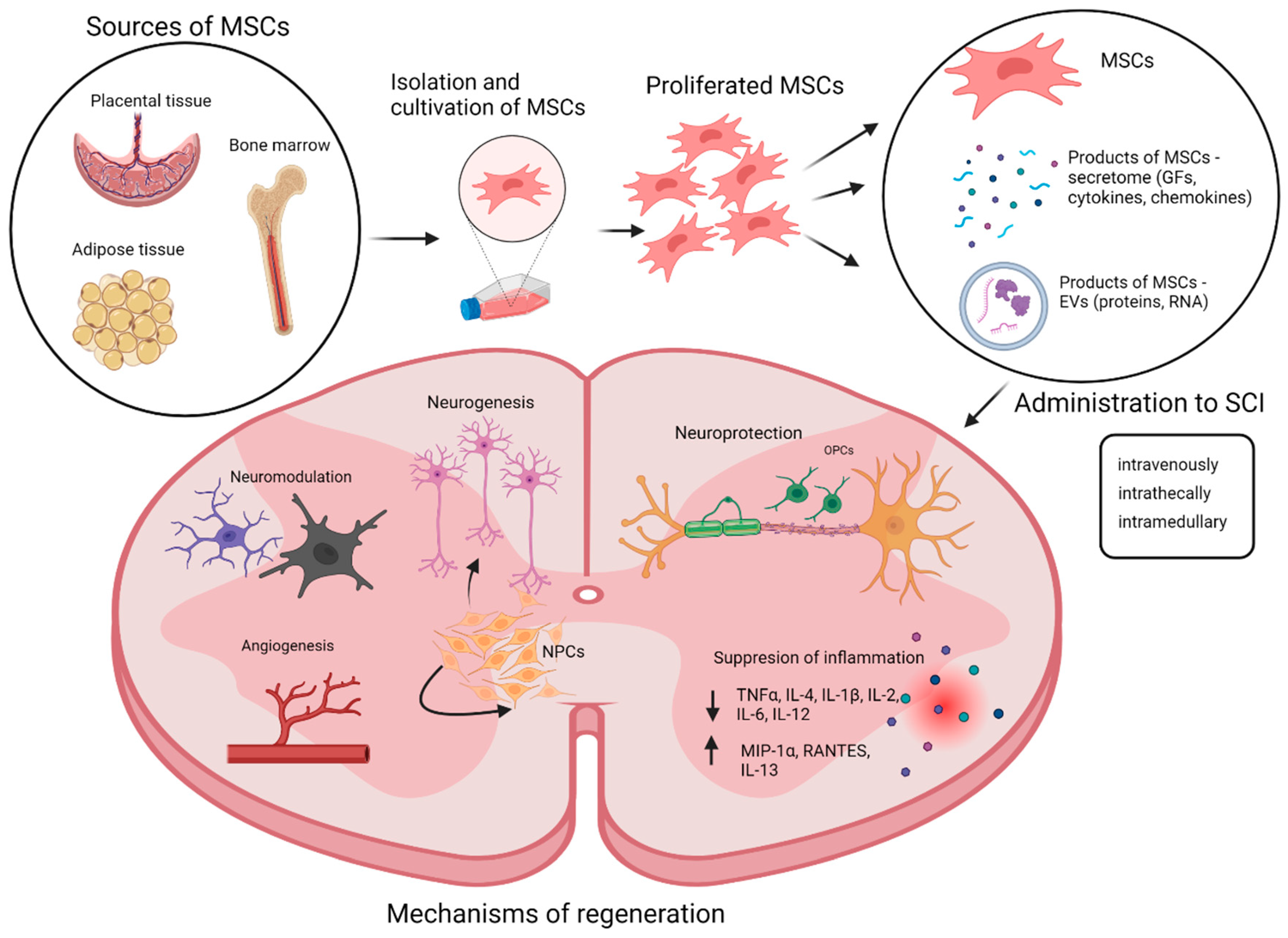
:1. Introduction
2. Mesenchymal Stromal Cells (MSCs)
3. MSC Therapy
3.1. MSC Doses, Method, and Time Frame of the Application
3.2. MSCs with Biomaterials and Scaffold
4. Paracrine Effect of MSCs
4.1. Secretome

{kind=link}
| Factor | Effect | References |
|---|---|---|
| vascular endothelial growth factor (VEGF) | supports angiogenesis, enhances the autophagic flux and reduces inflammation reaction, promotes neuronal survival | [101,102] |
| basic fibroblast growth factor (bFGF) | modulates neural progenitor proliferation and differentiation | [103] |
| nerve growth factor (NGF) | promotes angiogenic activity and reduces apoptosis, promotes neuronal cell survival and neuritogenesis | [99,104,105] |
| brain-derived neurotrophic factor (BDNF) | neuroprotective potential on primary auditory neurons increases neuroregeneration and synapses using BDNF-dependent mechanisms ppAKT and ppERK 1/2 | [99,106,107] |
| hepatocyte growth factor (HGF) | mediates functional recovery and remyelination through HGF/cMet signaling pathway | [108,109] |
| chemokine ligand 1 (CX3CL1) | activation of microglia to a neuroprotective phenotype | [93] |
4.2. Extracellular Vesicles (EVs)
5. Clinical Studies
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Rowland, J.W.; Hawryluk, G.W.J.; Kwon, B.; Fehlings, M.G. Current status of acute spinal cord injury pathophysiology and emerging therapies: Promise on the horizon. Neurosurg. Focus 2008, 25, E2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, J.W.; Sadowsky, C. Spinal-cord injury. Lancet 2002, 359, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Dai, Y.; Chen, G.; Cui, S. Dissecting the Dual Role of the Glial Scar and Scar-Forming Astrocytes in Spinal Cord Injury. Front Cell. Neurosci. 2020, 14, 78. [Google Scholar] [CrossRef] [Green Version]
- Gerber, Y.N.; Saint-Martin, G.P.; Bringuier, C.M.; Bartolami, S.; Goze-Bac, C.; Noristani, H.N.; Perrin, F.E. CSF1R Inhibition Reduces Microglia Proliferation, Promotes Tissue Preservation and Improves Motor Recovery After Spinal Cord Injury. Front. Cell. Neurosci. 2018, 12, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.-S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Guo, W.; Zhu, Z.; Hu, Y.; Wang, Y.; Zhang, X.; Wang, W.; Du, N.; Song, T.; Yang, K.; et al. Macrophage migration inhibitory factor facilitates production of CCL5 in astrocytes following rat spinal cord injury. J. Neuroinflamm. 2018, 15, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, A.P.; Warren, P.M.; Silver, J. New insights into glial scar formation after spinal cord injury. Cell Tissue Res. 2021, 387, 319–336. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, Z.-L.; Xie, H.; Tian, X.-B.; Xu, H.-L.; Li, W.; Yao, S. Microglial depletion impairs glial scar formation and aggravates inflammation partly by inhibiting STAT3 phosphorylation in astrocytes after spinal cord injury. Neural Regen. Res. 2023, 18, 1325–1331. [Google Scholar] [CrossRef]
- Bracken, M.B.; Shepard, M.J.; Holford, T.R.; Leo-Summers, L.; Aldrich, E.F.; Fazl, M.; Fehlings, M.G.; Herr, D.L.; Hitchon, P.W.; Marshall, L.F.; et al. Methylprednisolone or tirilazad mesylate administration after acute spinal cord injury: 1-year follow up. Results of the third National Acute Spinal Cord Injury randomized controlled trial. J. Neurosurg. 1998, 89, 699–706. [Google Scholar] [CrossRef]
- Bartlett, R.D.; Burley, S.; Ip, M.; Phillips, J.B.; Choi, D. Cell Therapies for Spinal Cord Injury: Trends and Challenges of Current Clinical Trials. Neurosurgery 2020, 87, E456–E472. [Google Scholar] [CrossRef]
- Nori, S.; Khazaei, M.; Ahuja, C.S.; Yokota, K.; Ahlfors, J.-E.; Liu, Y.; Wang, J.; Shibata, S.; Chio, J.; Hettiaratchi, M.H.; et al. Human Oligodendrogenic Neural Progenitor Cells Delivered with Chondroitinase ABC Facilitate Functional Repair of Chronic Spinal Cord Injury. Stem Cell Rep. 2018, 11, 1433–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nori, S.; Okada, Y.; Nishimura, S.; Sasaki, T.; Itakura, G.; Kobayashi, Y.; Renault-Mihara, F.; Shimizu, A.; Koya, I.; Yoshida, R.; et al. Long-term safety issues of iPSC-based cell therapy in a spinal cord injury model: Oncogenic transformation with epithelial-mesenchymal transition. Stem Cell Rep. 2015, 4, 360–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alagesan, S.; Griffin, M.D. Autologous and allogeneic mesenchymal stem cells in organ transplantation: What do we know about their safety and efficacy? Curr. Opin. Organ. Transpl. 2014, 19, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, S.; Shi, Y.; Galipeau, J.; Krampera, M.; Leblanc, K.; Martin, I.; Nolta, J.; Phinney, D.G.; Sensebe, L. Mesenchymal stem versus stromal cells: International Society for Cell & Gene Therapy (ISCT(R)) Mesenchymal Stromal Cell committee position statement on nomenclature. Cytotherapy 2019, 21, 1019–1024. [Google Scholar] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Stewart, M.C.; Stewart, A.A. Mesenchymal stem cells: Characteristics, sources, and mechanisms of action. Veter-Clin. N. Am. Equine Pract. 2011, 27, 243–261. [Google Scholar] [CrossRef]
- Mushahary, D.; Spittler, A.; Kasper, C.; Weber, V.; Charwat, V. Isolation, cultivation, and characterization of human mesenchymal stem cells. Cytom. Part A 2017, 93, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Gu, L.-H.; Zhang, T.-T.; Li, Y.; Yan, H.-J.; Qi, H.; Li, F.-R. Immunogenicity of allogeneic mesenchymal stem cells transplanted via different routes in diabetic rats. Cell. Mol. Immunol. 2014, 12, 444–455. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.-L.; Liu, Y.; Yang, S.-G.; Zhao, Q.-J.; Wang, X.; Gong, W.; Han, Z.-B.; Xu, Z.-S.; Lu, Y.-X.; Liu, D.; et al. Isolation and characterization of human umbilical cord mesenchymal stem cells with hematopoiesis-supportive function and other potentials. Haematologica 2006, 91, 1017–1026. [Google Scholar]
- Barkholt, L.; Flory, E.; Jekerle, V.; Lucas-Samuel, S.; Ahnert, P.; Bisset, L.; Büscher, D.; Fibbe, W.; Foussat, A.; Kwa, M.; et al. Risk of tumorigenicity in mesenchymal stromal cell–based therapies—Bridging scientific observations and regulatory viewpoints. Cytotherapy 2013, 15, 753–759. [Google Scholar] [CrossRef]
- Marques, J.C.M.T.; Cornélio, D.A.; Silbiger, V.N.; Luchessi, A.D.; de Souza, S.; de Medeiros, S.R.B. Identification of new genes associated to senescent and tumorigenic phenotypes in mesenchymal stem cells. Sci. Rep. 2017, 7, 17837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berebichez-Fridman, R.; Montero-Olvera, P.R. Sources and Clinical Applications of Mesenchymal Stem Cells: State-of-the-art review. Sultan Qaboos Univ. Med. J. 2018, 18, e264–e277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyano, K.; Ikehata, M.; Ohshima, K.; Yoshida, Y.; Nose, Y.; Yoshihara, S.-I.; Oki, K.; Shiraishi, S.; Uzu, M.; Nonaka, M.; et al. Intravenous administration of human mesenchymal stem cells derived from adipose tissue and umbilical cord improves neuropathic pain via suppression of neuronal damage and anti-inflammatory actions in rats. PLoS ONE 2022, 17, e0262892. [Google Scholar] [CrossRef]
- Albu, S.; Kumru, H.; Coll, R.; Vives, J.; Vallés, M.; Benito-Penalva, J.; Rodríguez, L.; Codinach, M.; Hernández, J.; Navarro, X.; et al. Clinical effects of intrathecal administration of expanded Wharton jelly mesenchymal stromal cells in patients with chronic complete spinal cord injury: A randomized controlled study. Cytotherapy 2021, 23, 146–156. [Google Scholar] [CrossRef]
- DiGirolamo, C.M.; Stokes, D.; Colter, D.; Phinney, D.G.; Class, R.; Prockop, D.J. Propagation and senescence of human marrow stromal cells in culture: A simple colony-forming assay identifies samples with the greatest potential to propagate and differentiate. Br. J. Haematol. 1999, 107, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Syková, E.; Homola, A.; Mazanec, R.; Lachmann, H.; Konrádová, L.; Kobylka, P.; Pádr, R.; Neuwirth, J.; Komrska, V.; Vávra, V.; et al. Autologous bone marrow transplantation in patients with subacute and chronic spinal cord injury. Cell Transplant. 2006, 15, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Kokai, L.E.; Marra, K.; Rubin, J.P. Adipose stem cells: Biology and clinical applications for tissue repair and regeneration. Transl. Res. 2014, 163, 399–408. [Google Scholar] [CrossRef]
- Jin, H.J.; Bae, Y.K.; Kim, M.; Kwon, S.-J.; Jeon, H.B.; Choi, S.J.; Kim, S.W.; Yang, Y.S.; Oh, W.; Chang, J.W. Comparative analysis of human mesenchymal stem cells from bone marrow, adipose tissue, and umbilical cord blood as sources of cell therapy. Int. J. Mol. Sci. 2013, 14, 17986–18001. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.-L.; Luo, H.-S.; Li, J.-T.; Xia, Y.-Z.; Li, L.; Zhang, L.-J.; Meng, H.; Cui, G.-Y.; Chen, Z.; Wu, N.; et al. Functional recovery in acute traumatic spinal cord injury after transplantation of human umbilical cord mesenchymal stem cells. Crit. Care Med. 2010, 38, 2181–2189. [Google Scholar] [CrossRef]
- Reyhani, S.; Abbaspanah, B.; Mousavi, S.H. Umbilical cord-derived mesenchymal stem cells in neurodegenerative disorders: From literature to clinical practice. Regen. Med. 2020, 15, 1561–1578. [Google Scholar] [CrossRef]
- Stenderup, K.; Justesen, J.; Clausen, C.; Kassem, M. Aging is associated with decreased maximal life span and accelerated senescence of bone marrow stromal cells. Bone 2003, 33, 919–926. [Google Scholar] [CrossRef] [PubMed]
- English, K.; Wood, K.J. Mesenchymal stromal cells in transplantation rejection and tolerance. Cold Spring Harb. Perspect. Med. 2013, 3, a015560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ankrum, J.; Ong, J.F.; Karp, J.M. Mesenchymal stem cells: Immune evasive, not immune privileged. Nat. Biotechnol. 2014, 32, 252–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Huang, X.; Wang, H.; Liu, X.; Zhang, T.; Wang, Y.; Hu, D. The challenges and promises of allogeneic mesenchymal stem cells for use as a cell-based therapy. Stem Cell Res. Ther. 2015, 6, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Wang, Z.; Zheng, X.; Ding, L.; Han, D.; Yan, H.; Guo, Z.; Wang, H. Haploidentical hematopoietic stem cell transplant with umbilical cord-derived multipotent mesenchymal cell infusion for the treatment of high-risk acute leukemia in children. Leuk. Lymphoma 2015, 56, 1346–1352. [Google Scholar] [CrossRef]
- Li, C.; Zhao, H.; Cheng, L.; Bin Wang, B. Allogeneic vs. autologous mesenchymal stem/stromal cells in their medication practice. Cell Biosci. 2021, 11, 187. [Google Scholar] [CrossRef]
- Qi, X.; Shao, M.; Peng, H.; Bi, Z.; Su, Z.; Li, H. In vitro differentiation of bone marrow stromal cells into neurons and glial cells and differential protein expression in a two-compartment bone marrow stromal cell/neuron co-culture system. J. Clin. Neurosci. 2010, 17, 908–913. [Google Scholar] [CrossRef]
- Liau, L.L.; Looi, Q.H.; Chia, W.C.; Subramaniam, T.; Ng, M.H.; Law, J.X. Treatment of spinal cord injury with mesenchymal stem cells. Cell Biosci. 2020, 10, 112. [Google Scholar] [CrossRef]
- Sanchez-Ramos, J.; Song, S.; Cardozo-Pelaez, F.; Hazzi, C.; Stedeford, T.; Willing, A.; Freeman, T.; Saporta, S.; Janssen, W.; Patel, N.; et al. Adult bone marrow stromal cells differentiate into neural cells in vitro. Exp. Neurol. 2000, 164, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Krabbe, C.; Zimmer, J.; Meyer, M. Neural transdifferentiation of mesenchymal stem cells—A critical review. Apmis 2005, 113, 831–844. [Google Scholar] [CrossRef]
- Lee, H.-S.; Kim, K.-S.; Eun-Ju, O.; Joe, Y.-A. A Comparison of ROCK Inhibitors on Human Bone Marrow-Derived Mesenchymal Stem Cell Differentiation into Neuron-Like Cells. Biomol. Ther. 2010, 18, 386–395. [Google Scholar] [CrossRef] [Green Version]
- Wislet-Gendebien, S.; Hans, G.; Leprince, P.; Rigo, J.; Moonen, G.; Rogister, B. Plasticity of cultured mesenchymal stem cells: Switch from nestin-positive to excitable neuron-like phenotype. Stem Cells 2005, 23, 392–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Xu, C.; Liu, Z.; Yang, L.; Hong, Y.; Liu, G.; Zhong, H.; Cai, X.; Lin, X.; Chen, X.; et al. Neural differentiation of bone marrow mesenchymal stem cells with human brain-derived neurotrophic factor gene-modified in functionalized self-assembling peptide hydrogel in vitro. J. Cell. Biochem. 2019, 120, 2828–2835. [Google Scholar] [CrossRef] [PubMed]
- Lattanzi, W.; Concetta Geloso, M.; Saulnier, N.; Giannetti, S.; Puglisi, M.A.; Corvino, V.; Gasbarrini, A.; Michetti, F. Neurotrophic features of human adipose tissue-derived stromal cells: In vitro and in vivo studies. J. Biomed. Biotechnol. 2011, 2011, 468705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.-L.; Pan, X.-M.; Chen, H.-H.; Fu, X.-Y.; Jiang, J.; Ding, M.-X. Repairing and Analgesic Effects of Umbilical Cord Mesenchymal Stem Cell Transplantation in Mice with Spinal Cord Injury. BioMed Res. Int. 2020, 2020, 7650354. [Google Scholar] [CrossRef]
- Yousefifard, M.; Nasirinezhad, F.; Manaheji, H.S.; Janzadeh, A.; Hosseini, M.; Keshavarz, M. Human bone marrow-derived and umbilical cord-derived mesenchymal stem cells for alleviating neuropathic pain in a spinal cord injury model. Stem Cell Res. Ther. 2016, 7, 36. [Google Scholar] [CrossRef] [Green Version]
- Upadhyayula, P.S.; Martin, J.R.; Rennert, R.C.; Ciacci, J.D. Review of operative considerations in spinal cord stem cell therapy. World J. Stem Cells 2021, 13, 168–176. [Google Scholar] [CrossRef]
- Ra, J.C.; Shin, I.S.; Kim, S.H.; Kang, S.K.; Kang, B.C.; Lee, H.Y.; Kim, Y.J.; Jo, J.Y.; Yoon, E.J.; Choi, H.J.; et al. Safety of intravenous infusion of human adipose tissue-derived mesenchymal stem cells in animals and humans. Stem Cells Dev. 2011, 20, 1297–1308. [Google Scholar] [CrossRef]
- Choi, E.W.; Lee, H.W.; Shin, I.S.; Park, J.H.; Yun, T.W.; Youn, H.Y.; Kim, S.-J. Comparative Efficacies of Long-Term Serial Transplantation of Syngeneic, Allogeneic, Xenogeneic, or CTLA4Ig-Overproducing Xenogeneic Adipose Tissue-Derived Mesenchymal Stem Cells on Murine Systemic Lupus Erythematosus. Cell Transplant. 2016, 25, 1193–1206. [Google Scholar] [CrossRef] [Green Version]
- Wagner, B.; Henschler, R. Fate of intravenously injected mesenchymal stem cells and significance for clinical application. Adv. Biochem. Eng. Biotechnol. 2013, 130, 19–37. [Google Scholar]
- Leibacher, J.; Dauber, K.; Ehser, S.; Brixner, V.; Kollar, K.; Vogel, A.; Spohn, G.; Schäfer, R.; Seifried, E.; Henschler, R. Human mesenchymal stromal cells undergo apoptosis and fragmentation after intravenous application in immune-competent mice. Cytotherapy 2017, 19, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.; Pahwa, B.; Agrawal, D.; Singh, P.; Gurjar, H.; Mishra, S.; Jagdevan, A.; Misra, M.C. Safety and feasibility of intramedullary injected bone marrow–derived mesenchymal stem cells in acute complete spinal cord injury: Phase 1 trial. J. Neurosurg. Spine 2022, 37, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Martinez, A.M.B.; Ramalho, B.D.S.; de Almeida, F.M.; Sales, C.M.; de Lima, S. Injection of bone marrow mesenchymal stem cells by intravenous or intraperitoneal routes is a viable alternative to spinal cord injury treatment in mice. Neural Regen. Res. 2018, 13, 1046–1053. [Google Scholar] [CrossRef]
- Ninomiya, K.; Iwatsuki, K.; Ohnishi, Y.-I.; Ohkawa, T.; Yoshimine, T. Intranasal delivery of bone marrow stromal cells to spinal cord lesions. J. Neurosurg. Spine 2015, 23, 111–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, L.; Peng, Y.; Xu, W.; He, P.; Li, T.; Lu, X.; Chen, G. Progress in Stem Cell Therapy for Spinal Cord Injury. Stem Cells Int. 2020, 2020, 2853650. [Google Scholar] [CrossRef] [PubMed]
- Antonic, A.; Sena, E.S.; Lees, J.S.; Wills, T.E.; Skeers, P.; Batchelor, P.E.; Macleod, M.R.; Howells, D.W. Stem cell transplantation in traumatic spinal cord injury: A systematic review and meta-analysis of animal studies. PLoS Biol. 2013, 11, e1001738. [Google Scholar] [CrossRef] [Green Version]
- Moon, S.M.; Kim, W.; Chung, J.Y.; Im, W.; Yoo, D.Y.; Jung, H.Y.; Won, M.-H.; Choi, J.H.; Hwang, I.K. Neuroprotective Effects of Adipose-Derived Stem Cells Are Maintained for 3 Weeks against Ischemic Damage in the Rabbit Spinal Cord. BioMed Res. Int. 2014, 2014, 539051. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Rong, L.-M.; Li, M.-M.; Cao, T.-T.; Chen, H.; Pang, M.; Xu, S.-S.; Wen, H.-Q. Dose optimization of intrathecal administration of human umbilical cord mesenchymal stem cells for the treatment of subacute incomplete spinal cord injury. Neural Regen. Res. 2022, 17, 1785–1794. [Google Scholar] [CrossRef]
- Vaquero, J.; Zurita, M.; Rico, M.A.; Aguayo, C.; Bonilla, C.; Marin, E.; Tapiador, N.; Sevilla, M.; Vazquez, D.; Carballido, J.; et al. Intrathecal administration of autologous mesenchymal stromal cells for spinal cord injury: Safety and efficacy of the 100/3 guideline. Cytotherapy 2018, 20, 806–819. [Google Scholar] [CrossRef]
- Jamali, F.; Alqudah, M.; Rahmeh, R.; Bawaneh, H.; Al-Shudifat, A.; Samara, O.; Awidi, A. Safe Reversal of Motor and Sensory Deficits by Repeated High Doses of Mesenchymal Stem Cells in a Patient with Chronic Complete Spinal Cord Injury. Am. J. Case Rep. 2023, 24, e938576. [Google Scholar] [CrossRef]
- Yang, Y.; Pang, M.; Du, C.; Liu, Z.-Y.; Chen, Z.-H.; Wang, N.-X.; Zhang, L.-M.; Chen, Y.-Y.; Mo, J.; Dong, J.-W.; et al. Repeated subarachnoid administrations of allogeneic human umbilical cord mesenchymal stem cells for spinal cord injury: A phase 1/2 pilot study. Cytotherapy 2021, 23, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Hirota, R.; Sasaki, M.; Kataoka-Sasaki, Y.; Oshigiri, T.; Kurihara, K.; Fukushi, R.; Oka, S.; Ukai, R.; Yoshimoto, M.; Kocsis, J.D.; et al. Enhanced Network in Corticospinal Tracts after Infused Mesenchymal Stem Cells in Spinal Cord Injury. J. Neurotrauma 2022, 39, 1665–1677. [Google Scholar] [CrossRef] [PubMed]
- Slovinska, L.; Harvanova, D.; Janockova, J.; Matejova, J.; Cibur, P.; Moravek, M.; Spakova, T.; Rosocha, J. Mesenchymal Stem Cells in the Treatment of Human Spinal Cord Injury: The Effect on Individual Values of pNF-H, GFAP, S100 Proteins and Selected Growth Factors, Cytokines and Chemokines. Curr. Issues Mol. Biol. 2022, 44, 578–596. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Tang, F.; Zhao, Y.; Han, G.; Yin, N.; Li, X.; Chen, B.; Han, S.; Jiang, X.; Yun, C.; et al. Significant Improvement of Acute Complete Spinal Cord Injury Patients Diagnosed by a Combined Criteria Implanted with NeuroRegen Scaffolds and Mesenchymal Stem Cells. Cell Transplant. 2018, 27, 907–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phedy, P.; Djaja, Y.P.; Gatam, L.; Kusnadi, Y.; Wirawan, R.P.; Tobing, I.M.; Subakir, N.; Mappalilu, A.; Prawira, M.A.; Yauwenas, R.; et al. Motoric Recovery After Transplantation of Bone Marrow Derived Mesenchymal Stem Cells in Chronic Spinal Cord Injury: A Case Report. Am. J. Case Rep. 2019, 20, 1299–1304. [Google Scholar] [CrossRef] [PubMed]
- Blando, S.; Anchesi, I.; Mazzon, E.; Gugliandolo, A. Can a Scaffold Enriched with Mesenchymal Stem Cells Be a Good Treatment for Spinal Cord Injury? Int. J. Mol. Sci. 2022, 23, 7545. [Google Scholar] [CrossRef]
- Liu, T.; Zhu, W.; Zhang, X.; He, C.; Liu, X.; Xin, Q.; Chen, K.; Wang, H. Recent Advances in Cell and Functional Biomaterial Treatment for Spinal Cord Injury. BioMed Res. Int. 2022, 2022, 5079153. [Google Scholar] [CrossRef]
- Prang, P.; Mueller, R.; Eljaouhari, A.; Heckmann, K.; Kunz, W.; Weber, T.; Faber, C.; Vroemen, M.; Bogdahn, U.; Weidner, N. The promotion of oriented axonal regrowth in the injured spinal cord by alginate-based anisotropic capillary hydrogels. Biomaterials 2006, 27, 3560–3569. [Google Scholar] [CrossRef]
- Blaško, J.; Szekiova, E.; Slovinska, L.; Kafka, J.; Cizkova, D. Axonal outgrowth stimulation after alginate/mesenchymal stem cell therapy in injured rat spinal cord. Acta Neurobiol. Exp. 2017, 77, 337–350. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Vázquez, M.; Vega-Ruiz, B.; Ramos-Zúñiga, R.; Saldaña-Koppel, D.A.; Quiñones-Olvera, L.F. Chitosan and Its Potential Use as a Scaffold for Tissue Engineering in Regenerative Medicine. BioMed Res. Int. 2015, 2015, 821279. [Google Scholar] [CrossRef] [Green Version]
- Boido, M.; Ghibaudi, M.; Gentile, P.; Favaro, E.; Fusaro, R.; Tonda-Turo, C. Chitosan-based hydrogel to support the paracrine activity of mesenchymal stem cells in spinal cord injury treatment. Sci. Rep. 2019, 9, 6402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grulova, I.; Slovinska, L.; Blaško, J.; Devaux, S.; Wisztorski, M.; Salzet, M.; Fournier, I.; Kryukov, O.; Cohen, S.; Cizkova, D. Delivery of Alginate Scaffold Releasing Two Trophic Factors for Spinal Cord Injury Repair. Sci. Rep. 2015, 5, 13702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Che, M.-T.; Zeng, X.; Qiu, X.-C.; Feng, B.; Lai, B.-Q.; Shen, H.-Y.; Ling, E.-A.; Zeng, Y.-S. Neurotrophin-3 released from implant of tissue-engineered fibroin scaffolds inhibits inflammation, enhances nerve fiber regeneration, and improves motor function in canine spinal cord injury. J. Biomed. Mater. Res. Part A 2018, 106, 2158–2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Mei, X.; Lv, S.; Zhang, Z.; Xu, J.; Sun, D.; Xu, J.; He, X.; Chi, G.; Li, Y. Rat vibrissa dermal papilla cells promote healing of spinal cord injury following transplantation. Exp. Ther. Med. 2018, 15, 3929–3939. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Gao, W.; Yue, B.; Jiang, J.; Gu, Y.; Dai, J.; Chen, L.; Shi, Q. Promotion of neurological recovery in rat spinal cord injury by mesenchymal stem cells loaded on nerve-guided collagen scaffold through increasing alternatively activated macrophage polarization. J. Tissue Eng. Regen. Med. 2018, 12, e1725–e1736. [Google Scholar] [CrossRef]
- Liu, D.; Li, X.; Xiao, Z.; Yin, W.; Zhao, Y.; Tan, J.; Chen, B.; Jiang, X.; Dai, J. Different functional bio-scaffolds share similar neurological mechanism to promote locomotor recovery of canines with complete spinal cord injury. Biomaterials 2019, 214, 119230. [Google Scholar] [CrossRef]
- Yari-Ilkhchi, A.; Ebrahimi-Kalan, A.; Farhoudi, M.; Mahkam, M. Design of graphenic nanocomposites containing chitosan and polyethylene glycol for spinal cord injury improvement. RSC Adv. 2021, 11, 19992–20002. [Google Scholar] [CrossRef]
- Cofano, F.; Boido, M.; Monticelli, M.; Zenga, F.; Ducati, A.; Vercelli, A.; Garbossa, D. Mesenchymal Stem Cells for Spinal Cord Injury: Current Options Limitations, and Future of Cell Therapy. Int. J. Mol. Sci. 2019, 20, 2698. [Google Scholar] [CrossRef] [Green Version]
- Wood, C.R.; Al Delfi, I.R.; Innes, J.F.; Myint, P.; Johnson, W.E. Exposing mesenchymal stem cells to chondroitin sulphated proteoglycans reduces their angiogenic and neuro-adhesive paracrine activity. Biochimie 2018, 155, 26–36. [Google Scholar] [CrossRef]
- Lim, H.-S.; Joe, Y.A. A ROCK Inhibitor Blocks the Inhibitory Effect of Chondroitin Sulfate Proteoglycan on Morphological Changes of Mesenchymal Stromal/Stem Cells into Neuron-Like Cells. Biomol. Ther. 2013, 21, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-S.; Yi, T.G.; Lee, H.-J.; Kim, S.-N.; Park, S.; Jeon, M.-S.; Song, S.U. Mesenchymal stem cells infected with Mycoplasma arginini secrete complement C3 to regulate immunoglobulin production in b lymphocytes. Cell Death Dis. 2014, 5, e1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caires, H.R.; Esteves, T.; Quelhas, P.; Barbosa, M.A.; Navarro, M.; Almeida, C.R. Macrophage interactions with polylactic acid and chitosan scaffolds lead to improved recruitment of human mesenchymal stem/stromal cells: A comprehensive study with different immune cells. J. R. Soc. Interface 2016, 13, 20160570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adutler-Lieber, S.; Ben-Mordechai, T.; Naftali-Shani, N.; Asher, E.; Loberman, D.; Raanani, E.; Leor, J. Human macrophage regulation via interaction with cardiac adipose tissue-derived mesenchymal stromal cells. J. Cardiovasc. Pharmacol. Ther. 2012, 18, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Xu, Y.; Liu, Q.; Zhang, Q. Mesenchymal Stem Cell-Macrophage Crosstalk and Maintenance of Inflammatory Microenvironment Homeostasis. Front. Cell Dev. Biol. 2021, 9, 681171. [Google Scholar] [CrossRef]
- Luque-Campos, N.; Bustamante-Barrientos, F.A.; Pradenas, C.; García, C.; Araya, M.J.; Bohaud, C.; Contreras-López, R.; Elizondo-Vega, R.; Djouad, F.; Luz-Crawford, P.; et al. The Macrophage Response Is Driven by Mesenchymal Stem Cell-Mediated Metabolic Reprogramming. Front. Immunol. 2021, 12, 624746. [Google Scholar] [CrossRef]
- Nakajima, H.; Uchida, K.; Guerrero, A.R.; Watanabe, S.; Sugita, D.; Takeura, N.; Yoshida, A.; Long, G.; Wright, K.T.; Johnson, W.E.; et al. Transplantation of Mesenchymal Stem Cells Promotes an Alternative Pathway of Macrophage Activation and Functional Recovery after Spinal Cord Injury. J. Neurotrauma 2012, 29, 1614–1625. [Google Scholar] [CrossRef] [Green Version]
- Urdzíková, L.M.; Růžička, J.; Labagnara, M.; Kárová, K.; Kubinová, Š.; Jiráková, K.; Murali, R.; Syková, E.; Jhanwar-Uniyal, M.; Jendelová, P. Human Mesenchymal Stem Cells Modulate Inflammatory Cytokines after Spinal Cord Injury in Rat. Int. J. Mol. Sci. 2014, 15, 11275–11293. [Google Scholar] [CrossRef] [Green Version]
- Neuhuber, B.; Himes, B.T.; Shumsky, J.S.; Gallo, G.; Fischer, I. Axon growth and recovery of function supported by human bone marrow stromal cells in the injured spinal cord exhibit donor variations. Brain Res. 2005, 1035, 73–85. [Google Scholar] [CrossRef]
- Fontaine, M.J.; Shih, H.; Schäfer, R.; Pittenger, M.F. Unraveling the Mesenchymal Stromal Cells’ Paracrine Immunomodulatory Effects. Transfus. Med. Rev. 2015, 30, 37–43. [Google Scholar] [CrossRef]
- Han, Y.; Yang, J.; Fang, J.; Zhou, Y.; Candi, E.; Wang, J.; Hua, D.; Shao, C.; Shi, Y. The secretion profile of mesenchymal stem cells and potential applications in treating human diseases. Signal Transduct. Target. Ther. 2022, 7, 92. [Google Scholar] [CrossRef]
- Pajer, K.; Bellák, T.; Nógrádi, A. Stem Cell Secretome for Spinal Cord Repair: Is It More than Just a Random Baseline Set of Factors? Cells 2021, 10, 3214. [Google Scholar] [CrossRef] [PubMed]
- Kiselevskii, M.V.; Vlasenko, R.Y.; Stepanyan, N.G.; Shubina, I.Z.; Sitdikova, S.M.; Kirgizov, K.I.; Varfolomeeva, S.R. Secretome of Mesenchymal Bone Marrow Stem Cells: Is It Immunosuppressive or Proinflammatory? Bull Exp. Biol. Med. 2021, 172, 250–253. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Xu, G.; Guo, J.; Xie, M.; Chen, L.; Xu, W. Mesenchymal Stem Cells Modulate Light-induced Activation of Retinal Microglia Through CX3CL1/CX3CR1 Signaling. Ocul. Immunol. Inflamm. 2016, 24, 684–692. [Google Scholar] [CrossRef]
- Han, D.; Wu, C.; Xiong, Q.; Zhou, L.; Tian, Y. Anti-inflammatory Mechanism of Bone Marrow Mesenchymal Stem Cell Transplantation in Rat Model of Spinal Cord Injury. Cell Biochem. Biophys. 2015, 71, 1341–1347. [Google Scholar] [CrossRef] [PubMed]
- Delfi, I.; Wood, C.R.; Johnson, L.D.V.; Snow, M.D.; Innes, J.F.; Myint, P.; Johnson, W.E.B. An In Vitro Comparison of the Neurotrophic and Angiogenic Activity of Human and Canine Adipose-Derived Mesenchymal Stem Cells (MSCs): Translating MSC-Based Therapies for Spinal Cord Injury. Biomolecules 2020, 10, 1301. [Google Scholar] [CrossRef] [PubMed]
- Taei, A.A.; Dargahi, L.; Khodabakhsh, P.; Kadivar, M.; Farahmandfar, M. Hippocampal neuroprotection mediated by secretome of human mesenchymal stem cells against experimental stroke. CNS Neurosci. Ther. 2022, 28, 1425–1438. [Google Scholar] [CrossRef]
- Székiová, E.; Michalová, Z.; Blaško, J.; Mucha, R.; Slovinská, L.; Kello, M.; Vanický, I. Characterisation of mesenchymal stem cells conditioned media obtained at different conditioning times: Their effect on glial cells in in vitro scratch model. Growth Factors 2023, 1–14. [Google Scholar] [CrossRef]
- Baldassarro, V.A.; Perut, F.; Cescatti, M.; Pinto, V.; Fazio, N.; Alastra, G.; Parziale, V.; Bassotti, A.; Fernandez, M.; Giardino, L.; et al. Intra-individual variability in the neuroprotective and promyelinating properties of conditioned culture medium obtained from human adipose mesenchymal stromal cells. Stem Cell Res. Ther. 2023, 14, 128. [Google Scholar] [CrossRef]
- Crigler, L.; Robey, R.C.; Asawachaicharn, A.; Gaupp, D.; Phinney, D.G. Human mesenchymal stem cell subpopulations express a variety of neuro-regulatory molecules and promote neuronal cell survival and neuritogenesis. Exp. Neurol. 2006, 198, 54–64. [Google Scholar] [CrossRef]
- Wood, C.R.; Juárez, E.H.; Ferrini, F.; Myint, P.; Innes, J.; Lossi, L.; Merighi, A.; Johnson, W.E. Mesenchymal stem cell conditioned medium increases glial reactivity and decreases neuronal survival in spinal cord slice cultures. Biochem. Biophys. Rep. 2021, 26, 100976. [Google Scholar] [CrossRef]
- Wang, H.; Wang, Y.; Li, D.; Liu, Z.; Zhao, Z.; Han, D.; Yuan, Y.; Bi, J.; Mei, X. VEGF inhibits the inflammation in spinal cord injury through activation of autophagy. Biochem. Biophys. Res. Commun. 2015, 464, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, S.; Zhao, Z.; Luo, Y.; Hou, Y.; Li, H.; He, L.; Zhou, L.; Wu, W. Effect of VEGF on Inflammatory Regulation, Neural Survival, and Functional Improvement in Rats following a Complete Spinal Cord Transection. Front. Cell. Neurosci. 2017, 11, 381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, F.G.; Carvalho, M.M.; Neves-Carvalho, A.; Panchalingam, K.M.; Behie, L.A.; Pinto, L.; Sousa, N.; Salgado, A.J. Secretome of mesenchymal progenitors from the umbilical cord acts as modulator of neural/glial proliferation and differentiation. Stem Cell Rev. Rep. 2014, 11, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Villatoro, A.J.; Alcoholado, C.; Martín-Astorga, M.; Fernández, V.; Cifuentes, M.; Becerra, J. Comparative analysis and characterization of soluble factors and exosomes from cultured adipose tissue and bone marrow mesenchymal stem cells in canine species. Veter-Immunol. Immunopathol. 2018, 208, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Zha, K.; Yang, Y.; Tian, G.; Sun, Z.; Yang, Z.; Li, X.; Sui, X.; Liu, S.; Zhao, J.; Guo, Q. Nerve growth factor (NGF) and NGF receptors in mesenchymal stem/stromal cells: Impact on potential therapies. Stem Cells Transl. Med. 2021, 10, 1008–1020. [Google Scholar] [CrossRef] [PubMed]
- Scheper, V.; Schwieger, J.; Hamm, A.; Lenarz, T.; Hoffmann, A. BDNF-overexpressing human mesenchymal stem cells mediate increased neuronal protection in vitro. J. Neurosci. Res. 2019, 97, 1414–1429. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Bayarsaikhan, D.; Lee, J.; Bayarsaikhan, G.; Lee, B. Brain-Derived Neurotrophic Factor Secreting Human Mesenchymal Stem Cells Improve Outcomes in Rett Syndrome Mouse Models. Front. Neurosci. 2021, 15, 725398. [Google Scholar] [CrossRef]
- Bai, L.; Lennon, D.P.; Caplan, A.I.; DeChant, A.; Hecker, J.; Kranso, J.; Zaremba, A.; Miller, R.H. Hepatocyte growth factor mediates mesenchymal stem cell–induced recovery in multiple sclerosis models. Nat. Neurosci. 2012, 15, 862–870. [Google Scholar] [CrossRef] [Green Version]
- Song, P.; Han, T.; Xiang, X.; Wang, Y.; Fang, H.; Niu, Y.; Shen, C. The role of hepatocyte growth factor in mesenchymal stem cell-induced recovery in spinal cord injured rats. Stem Cell Res. Ther. 2020, 11, 178. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Rohde, E.; Pachler, K.; Gimona, M. Manufacturing and characterization of extracellular vesicles from umbilical cord–derived mesenchymal stromal cells for clinical testing. Cytotherapy 2019, 21, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef] [PubMed]
- Hade, M.D.; Suire, C.N.; Suo, Z. Mesenchymal Stem Cell-Derived Exosomes: Applications in Regenerative Medicine. Cells 2021, 10, 1959. [Google Scholar] [CrossRef]
- Liu, W.-Z.; Ma, Z.-J.; Li, J.-R.; Kang, X.-W. Mesenchymal stem cell-derived exosomes: Therapeutic opportunities and challenges for spinal cord injury. Stem Cell Res. Ther. 2021, 12, 102. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Waheed, S.; Wang, C.; Shekh, M.; Li, Z.; Wu, J. Exosomes and Their Bioengineering Strategies in the Cutaneous Wound Healing and Related Complications: Current Knowledge and Future Perspectives. Int. J. Biol. Sci. 2023, 19, 1430–1454. [Google Scholar] [CrossRef]
- Zhao, L.; Hu, C.; Zhang, P.; Jiang, H.; Chen, J. Genetic communication by extracellular vesicles is an important mechanism underlying stem cell-based therapy-mediated protection against acute kidney injury. Stem Cell Res. Ther. 2019, 10, 119. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-J.; Wang, C. A review of the regulatory mechanisms of extracellular vesicles-mediated intercellular communication. Cell Commun. Signal. 2023, 21, 77. [Google Scholar] [CrossRef]
- Chang, Q.; Hao, Y.; Wang, Y.; Zhou, Y.; Zhuo, H.; Zhao, G. Bone marrow mesenchymal stem cell-derived exosomal microRNA-125a promotes M2 macrophage polarization in spinal cord injury by downregulating IRF5. Brain Res. Bull. 2021, 170, 199–210. [Google Scholar] [CrossRef]
- Deng, J.; Li, M.; Meng, F.; Liu, Z.; Wang, S.; Zhang, Y.; Li, M.; Li, Z.; Zhang, L.; Tang, P. 3D spheroids of human placenta-derived mesenchymal stem cells attenuate spinal cord injury in mice. Cell Death Dis. 2021, 12, 1096. [Google Scholar] [CrossRef]
- Staubach, S.; Bauer, F.N.; Tertel, T.; Börger, V.; Stambouli, O.; Salzig, D.; Giebel, B. Scaled preparation of extracellular vesicles from conditioned media. Adv. Drug Deliv. Rev. 2021, 177, 113940. [Google Scholar] [CrossRef]
- Banks, W.A.; Sharma, P.; Bullock, K.M.; Hansen, K.M.; Ludwig, N.; Whiteside, T.L. Transport of Extracellular Vesicles across the Blood-Brain Barrier: Brain Pharmacokinetics and Effects of Inflammation. Int. J. Mol. Sci. 2020, 21, 4407. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Deng, X.; Liu, M.; He, M.; Long, W.; Xu, Z.; Zhang, K.; Liu, T.; So, K.-F.; Fu, Q.-L.; et al. Intranasal delivery of BDNF-loaded small extracellular vesicles for cerebral ischemia therapy. J. Control. Release 2023, 357, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jiang, W.; Lu, Y.; Mao, T.; Gu, Y.; Ju, D.; Dong, C. Exosomes combined with biomaterials in the treatment of spinal cord injury. Front. Bioeng. Biotechnol. 2023, 11, 1077825. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, Y.; Mu, J.; Chen, J.; Zhang, C.; Cao, H.; Gao, J. Transplantation of Human Mesenchymal Stem-Cell-Derived Exosomes Immobilized in an Adhesive Hydrogel for Effective Treatment of Spinal Cord Injury. Nano Lett. 2020, 20, 4298–4305. [Google Scholar] [CrossRef]
- Demyanenko, S.V.; Pitinova, M.A.; Kalyuzhnaya, Y.N.; Khaitin, A.M.; Batalshchikova, S.A.; Dobaeva, N.M.; Shevtsova, Y.A.; Goryunov, K.V.; Plotnikov, E.Y.; Pashkevich, S.G.; et al. Human Multipotent Mesenchymal Stromal Cell–Derived Extracellular Vesicles Enhance Neuroregeneration in a Rat Model of Sciatic Nerve Crush Injury. Int. J. Mol. Sci. 2022, 23, 8583. [Google Scholar] [CrossRef] [PubMed]
- Terstappen, G.C.; Meyer, A.H.; Bell, R.D.; Zhang, W. Strategies for delivering therapeutics across the blood–brain barrier. Nat. Rev. Drug Discov. 2021, 20, 362–383. [Google Scholar] [CrossRef]
- Sung, S.-E.; Seo, M.-S.; Kim, Y.-I.; Kang, K.-K.; Choi, J.-H.; Lee, S.; Sung, M.; Yim, S.-G.; Lim, J.-H.; Seok, H.-G.; et al. Human Epidural AD–MSC Exosomes Improve Function Recovery after Spinal Cord Injury in Rats. Biomedicines 2022, 10, 678. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhang, J. Mesenchymal stem cell-derived exosomes containing miR-145-5p reduce inflammation in spinal cord injury by regulating the TLR4/NF-kappaB signaling pathway. Cell Cycle 2021, 20, 993–1009. [Google Scholar] [CrossRef]
- Deng, G.; Gao, Y.; Cen, Z.; He, J.; Cao, B.; Zeng, G.; Zong, S. miR-136-5p Regulates the Inflammatory Response by Targeting the IKKbeta/NF-kappaB/A20 Pathway After Spinal Cord Injury. Cell Physiol. Biochem. 2018, 50, 512–524. [Google Scholar] [CrossRef]
- Xin, H.; Li, Y.; Buller, B.; Katakowski, M.; Zhang, Y.; Wang, X.; Shang, X.; Zhang, Z.G.; Chopp, M. Exosome-Mediated Transfer of miR-133b from Multipotent Mesenchymal Stromal Cells to Neural Cells Contributes to Neurite Outgrowth. Stem Cells 2012, 30, 1556–1564. [Google Scholar] [CrossRef] [Green Version]
- Han, M.; Zhang, Z.; Liu, Z.; Liu, Y.; Zhao, H.; Wang, B.; Zhang, C.; Shang, H.; Li, Y.; Wang, S.; et al. Three-dimensional-cultured MSC-derived exosome with hydrogel for cerebral ischemia repair. Biomater. Adv. 2023, 149, 213396. [Google Scholar] [CrossRef] [PubMed]
- Ran, N.; Li, W.; Zhang, R.; Lin, C.; Zhang, J.; Wei, Z.; Li, Z.; Yuan, Z.; Wang, M.; Fan, B.; et al. Autologous exosome facilitates load and target delivery of bioactive peptides to repair spinal cord injury. Bioact. Mater. 2023, 25, 766–782. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, L.; Wang, S.; Han, Q.; Zhao, R.C. Exosomes secreted by mesenchymal stem cells promote endothelial cell angiogenesis by transferring miR-125a. J. Cell Sci. 2016, 129, 2182–2189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Kheir, W.A.; Gabr, H.; Awad, M.R.; Ghannam, O.; Barakat, Y.; Farghali, H.A.M.A.; El Maadawi, Z.M.; Ewes, I.; Sabaawy, H.E. Autologous bone marrow-derived cell therapy combined with physical therapy induces functional improvement in chronic spinal cord injury patients. Cell Transplant. 2014, 23, 729–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geffner, L.F.; Santacruz, P.; Izurieta, M.; Flor, L.; Maldonado, B.; Auad, A.H.; Montenegro, X.; Gonzalez, R.; Silva, F. Administration of autologous bone marrow stem cells into spinal cord injury patients via multiple routes is safe and improves their quality of life: Comprehensive case studies. Cell Transplant. 2008, 17, 1277–1293. [Google Scholar] [CrossRef] [Green Version]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slovinska, L.; Harvanova, D. The Role of Mesenchymal Stromal Cells and Their Products in the Treatment of Injured Spinal Cords. Curr. Issues Mol. Biol. 2023, 45, 5180-5197. https://doi.org/10.3390/cimb45060329
Slovinska L, Harvanova D. The Role of Mesenchymal Stromal Cells and Their Products in the Treatment of Injured Spinal Cords. Current Issues in Molecular Biology. 2023; 45(6):5180-5197. https://doi.org/10.3390/cimb45060329
Chicago/Turabian StyleSlovinska, Lucia, and Denisa Harvanova. 2023. "The Role of Mesenchymal Stromal Cells and Their Products in the Treatment of Injured Spinal Cords" Current Issues in Molecular Biology 45, no. 6: 5180-5197. https://doi.org/10.3390/cimb45060329