

Hair Growth Effect of DN106212 in C57BL/6 Mouse and Its Network Pharmacological Mechanism of Action

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
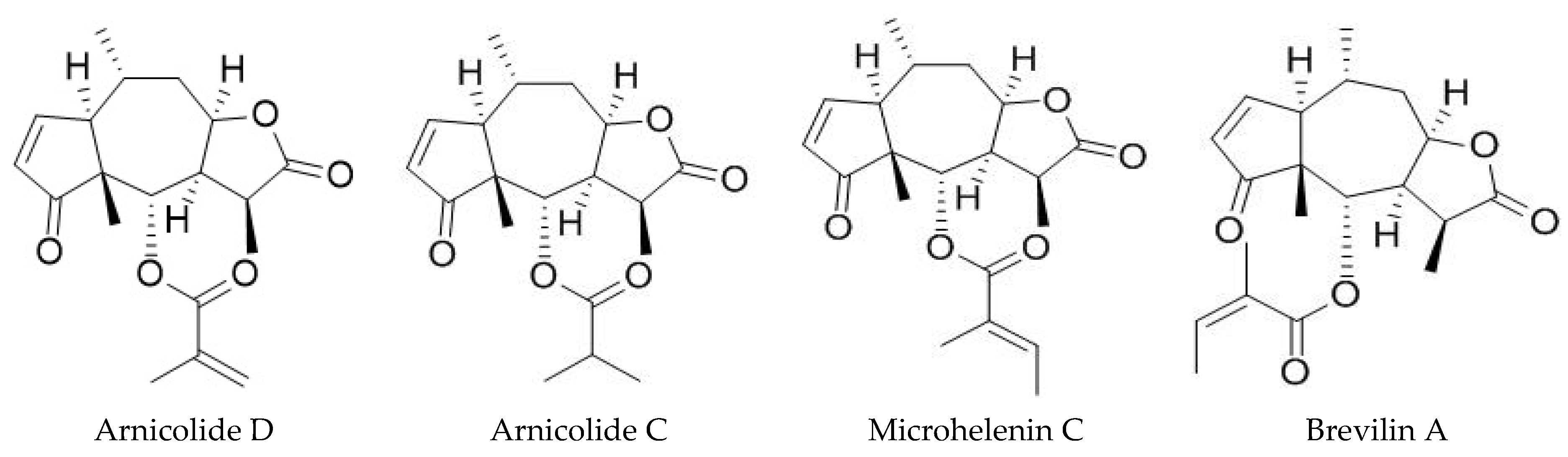
2.1. DN106212 Purified from Centipeda Minima Extraction
2.2. HPLC-UV Conditions
2.3. Experimental Mice and Their Management
2.4. Hair Growth Evaluation
2.5. Histological Study
2.6. Experimental Studies with DN106212
2.7. Quantitative Real-Time Polymerase Chain Reaction (qPCR) Analysis
2.8. Network Pharmacological Analyses
2.9. Statistical Analysis
3. Results
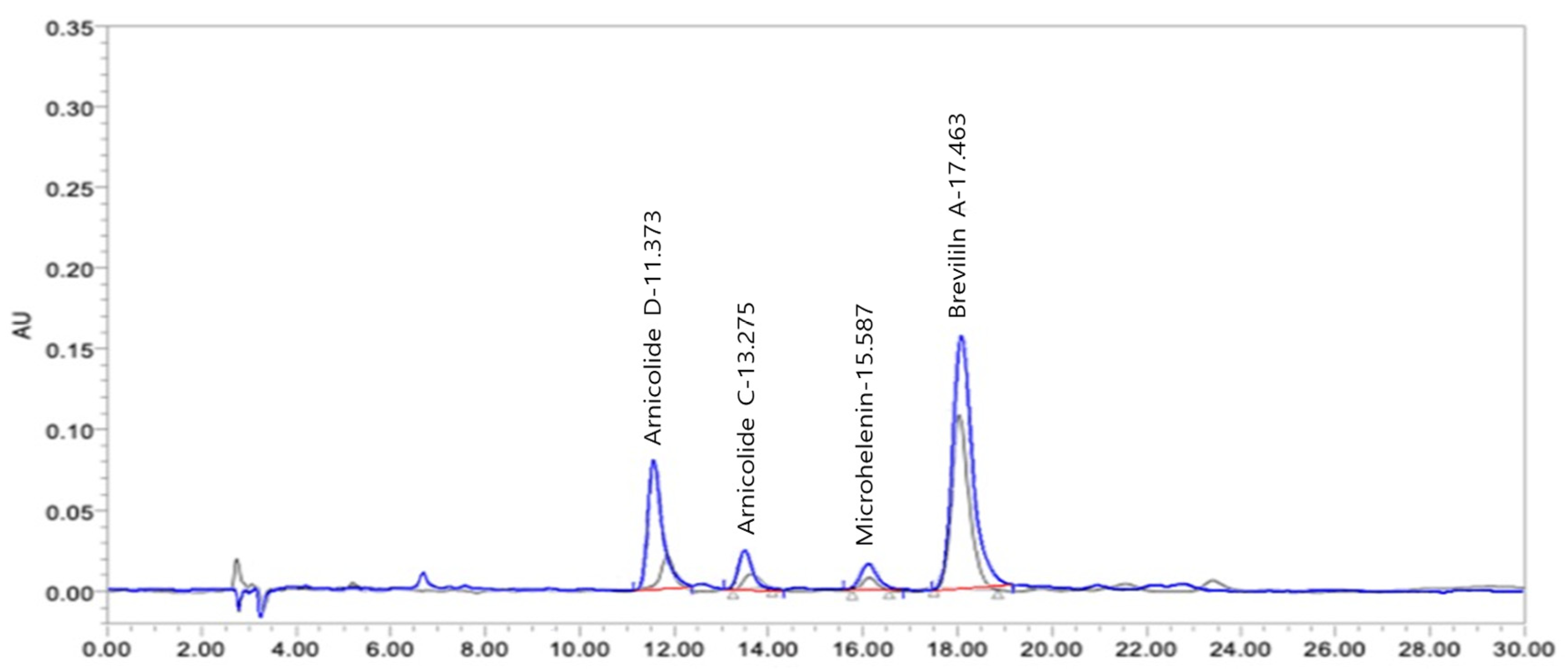
3.1. HPLC Chromatograms of Standards and DN106212
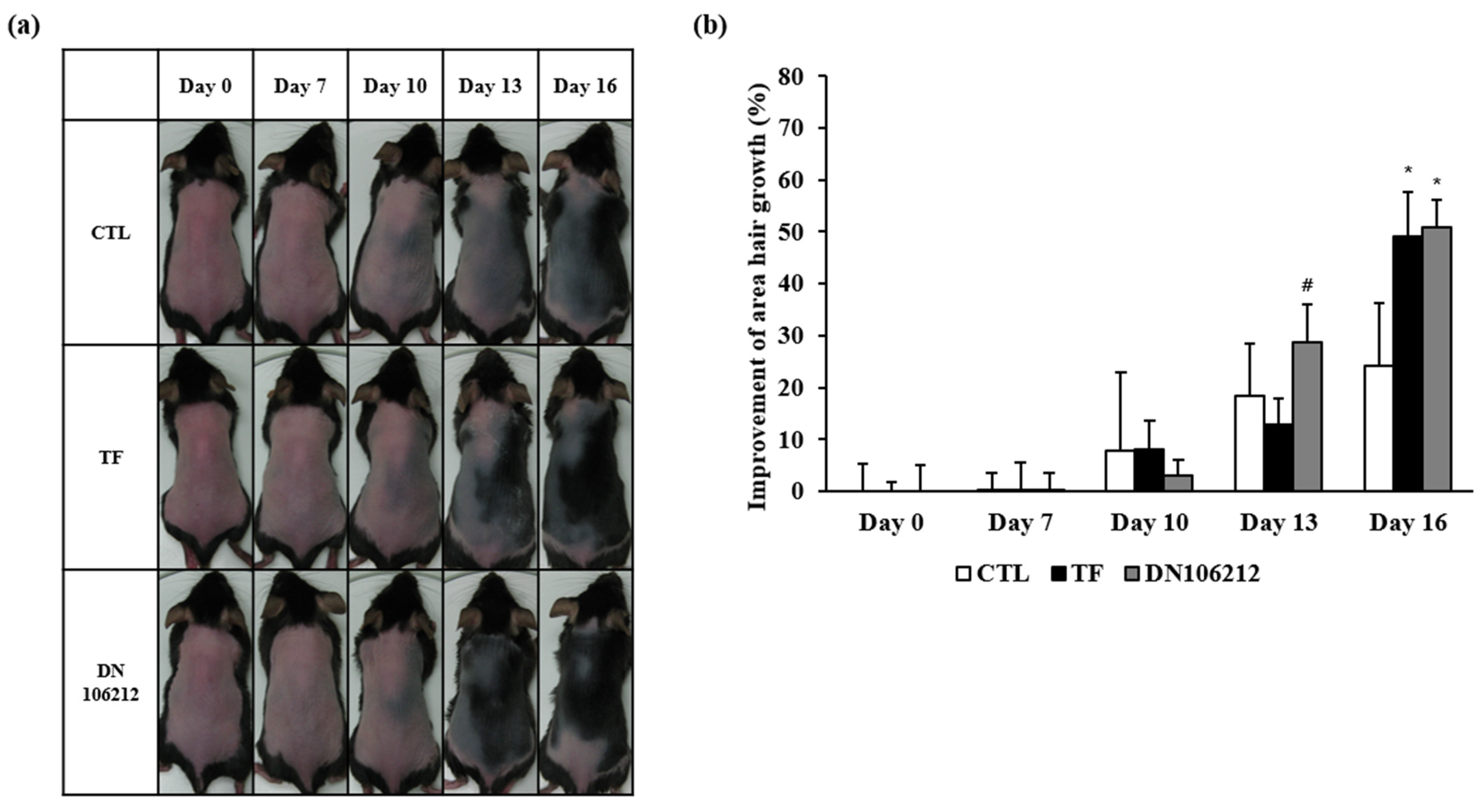
3.2. Effects of DN106212 on Hair Growth in Mice
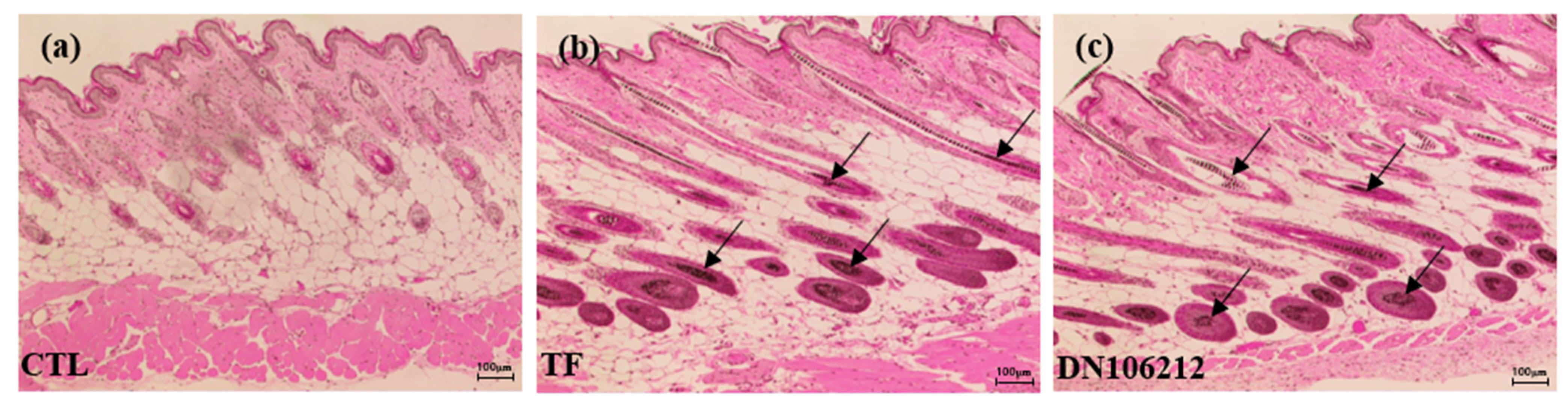
3.3. Effect of DN106212 on Hair Follicles in Mice
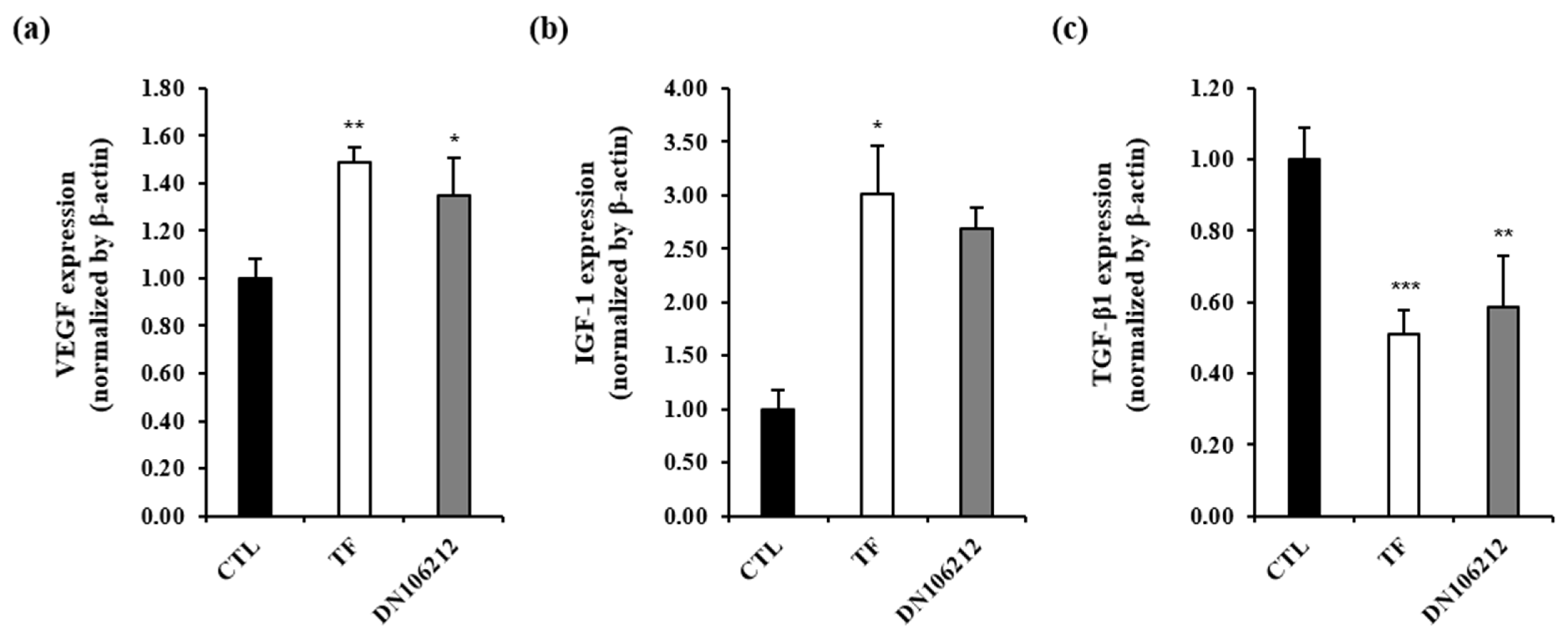
3.4. Effects of DN106212 on Vegfa, Igf1, and Tgfb1 in Dorsal Skin of Mice
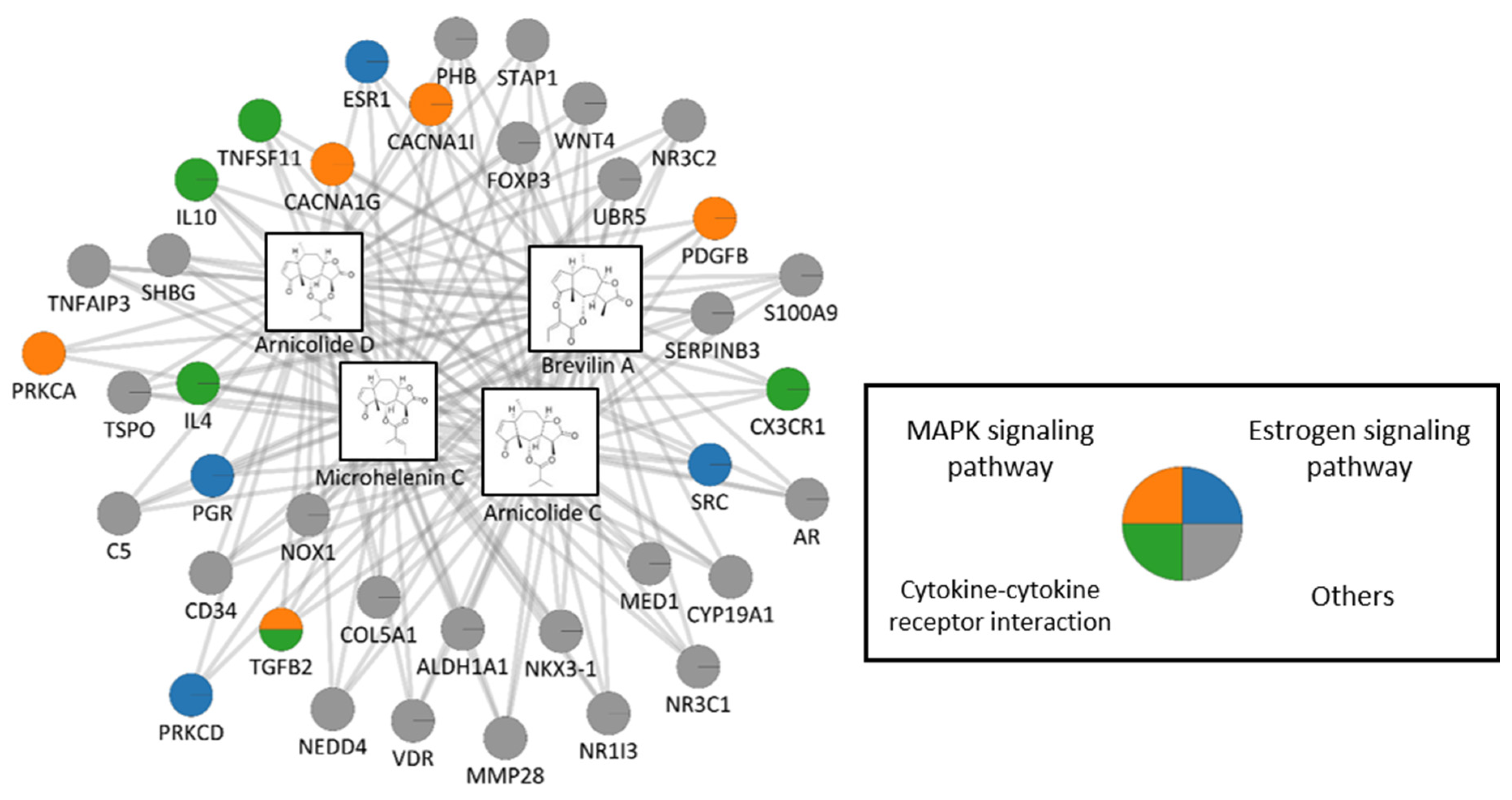
3.5. Network Pharmacology Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Patel, S.; Sharma, V.; Chauhan, N.S.; Thakur, M.; Dixit, V.K. Hair Growth: Focus on Herbal Therapeutic Agent. Curr. Drug. Discov. Technol. 2015, 12, 21–42. [Google Scholar] [CrossRef]
- Olsen, E.A. Female pattern hair loss. Hair Growth Disord. 2008, 45, 171–186. [Google Scholar]
- Hamilton, J.B. Male hormone stimulation is prerequisite and an incitant in common baldness. Am. J. Anat. 1942, 71, 451–480. [Google Scholar] [CrossRef]
- Birch, M.; Lalla, S.; Messenger, A. Female pattern hair loss. Clin. Exp. Dermatol. 2002, 27, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Alkhalifah, A.; Alsantali, A.; Wang, E.; McElwee, K.J.; Shapiro, J. Alopecia areata update: Part I. Clinical picture, histopathology, and pathogenesis. J. Am. Acad. Dermatol. 2010, 62, 177–188. [Google Scholar] [CrossRef]
- Shellow, W.V.; Edwards, J.E.; Koo, J.Y. Profile of alopecia areata: A questionnaire analysis of patient and family. Int. J. Dermatol. 1992, 31, 186–189. [Google Scholar] [CrossRef]
- Harrison, S.; Sinclair, R. Telogen effluvium. Clin. Exp. Dermatol. 2002, 27, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Ha, S. A Study on Influence Factors of Scalp and Hair Loss. Ph.D. Thesis, Deagu Hanny University, Kyoungbuk, Republic of Korea, 2005. [Google Scholar]
- Hattori, M.; Ogawa, H. Biochemical analysis of hair growth from the aspects of aging and enzyme activities. J. Dermatol. 1983, 10, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Kwak, I.-S.; Park, S.-H. The Effects of Combined Exercise and Scalp Care on the Scalp and Hair Condition of Male with Alopecia. Ann. Rom. Soc. Cell Biol. 2021, 25, 993–998. [Google Scholar]
- Dargie, H.; Dollery, C.; Daniel, J. Minoxidil in resistant hypertension. Lancet 1977, 310, 515–518. [Google Scholar] [CrossRef]
- Stenn, K.S.; Paus, R.; Chuong, C.-M.; Randall, V.A.; Widelitz, R.B.; Wu, P.; Jiang, T.-X.; Paul, M.J.; George, N.T.; Zucker, I.; et al. Controls of Hair Follicle Cycling. Physiol. Rev. 2001, 81, 449–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, B.Y. Hair-Growth Potential of Ginseng and Its Major Metabolites: A Review on Its Molecular Mechanisms. Int. J. Mol. Sci. 2018, 19, 2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.M.-L.; Chan, B.D.; Wong, W.-Y.; Qu, Z.; Chan, M.-S.; Leung, T.-W.; Lin, Y.; Mok, D.K.-W.; Chen, S.; Tai, W.C.-S. Anti-cancer Activity of Centipeda minima Extract in Triple Negative Breast Cancer via Inhibition of AKT, NF-κB, and STAT3 Signaling Pathways. Front. Oncol. 2020, 10, 491. [Google Scholar] [CrossRef]
- Huang, S.-S.; Chiu, C.-S.; Lin, T.-H.; Lee, M.-M.; Lee, C.-Y.; Chang, S.-J.; Hou, W.-C.; Huang, G.-J.; Deng, J.-S. Antioxidant and anti-inflammatory activities of aqueous extract of Centipeda minima. J. Ethnopharmacol. 2013, 147, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.-O.; Jin, D.-P.; Dong, N.-P.; Chen, S.-B.; Mok, D.K.W. Qualitative and quantitative analysis of chemical constituents of Centipeda minima by HPLC-QTOF-MS & HPLC-DAD. J. Pharm. Biomed. Anal. 2016, 125, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.G.; Yu, H.M.; Wen, S.L.; Liu, Y.L. Histopathological study on allergic rhinitis treated with Centipeda minima. Zhongguo Zhong yao za zhi= Zhongguo zhongyao zazhi = China J. Chin. Mater. Med. 2005, 30, 292–294. [Google Scholar]
- Qian, Y.; Zhao, C.; Yan, Y. The liver-protective effect of the Centipeda minima on hepatic injury in mice. China Pharm. 2004, 13, 25–26. [Google Scholar]
- Su, M.; Li, Y.; Chung, H.Y.; Ye, W. 2β-(Isobutyryloxy)florilenalin, a Sesquiterpene Lactone Isolated from the Medicinal Plant Centipeda minima, Induces Apoptosis in Human Nasopharyngeal Carcinoma CNE Cells. Molecules 2009, 14, 2135–2146. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R.S.; Towers, G. Antibacterial constituents of the nepalese medicinal herb, Centipeda minima. Phytochemistry 1998, 47, 631–634. [Google Scholar] [CrossRef]
- Wu, J.; Chun, Y.; Ebizuka, Y.; Sankawa, U. Biologically active constituents of centipeda minima: Isolation of a new plewolin ester and the antiallergy activity of sesquiterpene lactones. Chem. Pharm. Bull. 1985, 33, 4091–4094. [Google Scholar] [CrossRef] [Green Version]
- Xue, P.-H.; Zhang, N.; Liu, D.; Zhang, Q.-R.; Duan, J.-S.; Yu, Y.-Q.; Li, J.-Y.; Cao, S.-J.; Zhao, F.; Kang, N.; et al. Cytotoxic and Anti-Inflammatory Sesquiterpenes from the Whole Plants of Centipeda minima. J. Nat. Prod. 2021, 84, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Fang, B.; Wen, S.; Li, Y.; Bai, F.; Wei, Y.; Xiong, Y.; Huang, Q.; Lin, X. Prediction and verification of target of helenalin against hepatic stellate cell activation based on miR-200a-mediated PI3K/Akt and NF-κB pathways. Int. Immunopharmacol. 2021, 92, 107208. [Google Scholar] [CrossRef]
- Zhou, Y.-L.; Yan, Y.-M.; Li, S.-Y.; He, D.-H.; Xiong, S.; Wei, S.-F.; Liu, W.; Hu, L.; Wang, Q.; Pan, H.-F.; et al. 6-O-angeloylplenolin exerts neuroprotection against lipopolysaccharide-induced neuroinflammation in vitro and in vivo. Acta Pharmacol. Sin. 2020, 41, 10–21. [Google Scholar] [CrossRef]
- Chen, X.; Du, Y.; Nan, J.; Zhang, X.; Qin, X.; Wang, Y.; Hou, J.; Wang, Q.; Yang, J. Brevilin A, a Novel Natural Product, Inhibits Janus Kinase Activity and Blocks STAT3 Signaling in Cancer Cells. PLoS ONE 2013, 8, e63697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.J.; Jeon, M.S.; Lee, S.; Kim, J.K.; Jang, T.S.; Chung, K.H.; Kim, K.H. Anti-fibrotic effects of brevilin A in hepatic fibrosis via inhibiting the STAT3 signaling pathway. Bioorg. Med. Chem. Lett. 2021, 41, 127989. [Google Scholar] [CrossRef] [PubMed]
- You, P.; Wu, H.; Deng, M.; Peng, J.; Li, F.; Yang, Y. Brevilin A induces apoptosis and autophagy of colon adenocarcinoma cell CT26 via mitochondrial pathway and PI3K/AKT/mTOR inactivation. Biomed. Pharmacother. 2018, 98, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Su, T.; Wang, Y.P.; Wang, X.N.; Li, C.Y.; Zhu, P.L.; Huang, Y.M.; Yang, Z.Y.; Chen, S.B.; Yu, Z.L. The JAK2/STAT3 pathway is involved in the anti-melanoma effects of brevilin A. Life Sci. 2020, 241, 117169. [Google Scholar] [CrossRef]
- Qu, Z.; Lin, Y.; Mok, D.K.-W.; Bian, Q.; Tai, W.C.-S.; Chen, S. Brevilin A, a Natural Sesquiterpene Lactone Inhibited the Growth of Triple-Negative Breast Cancer Cells via Akt/mTOR and STAT3 Signaling Pathways. OncoTargets Ther. 2020, 13, 5363–5373. [Google Scholar] [CrossRef]
- Zhang, X.; He, J.; Huang, W.; Huang, H.; Zhang, Z.; Wang, J.; Yang, L.; Wang, G.; Wang, Y.; Li, Y. Antiviral Activity of the Sesquiterpene Lactones from Centipeda minima against Influenza a Virus in vitro. Nat. Prod. Commun. 2018, 13, 1934578X1801300201. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.-O.; Xie, X.-J.; Wan, S.-W.; Zhou, G.-L.; Yuen, A.C.-Y.; Mok, D.K.-W.; Chen, S.-B. Qualitative and quantitative analysis of sesquiterpene lactones in Centipeda minima by UPLC–Orbitrap–MS & UPLC-QQQ-MS. J. Pharm. Biomed. Anal. 2019, 174, 360–366. [Google Scholar] [CrossRef]
- Liu, R.; Chan, B.D.; Mok, D.K.-W.; Lee, C.-S.; Tai, W.C.-S.; Chen, S. Arnicolide D, from the herb Centipeda minima, Is a Therapeutic Candidate against Nasopharyngeal Carcinoma. Molecules 2019, 24, 1908. [Google Scholar] [CrossRef] [Green Version]
- Qu, Z.; Lin, Y.; Mok, D.K.-W.; Bian, Q.; Tai, W.C.-S.; Chen, S. Arnicolide D Inhibits Triple Negative Breast Cancer Cell Proliferation by Suppression of Akt/mTOR and STAT3 Signaling Pathways. Int. J. Med. Sci. 2020, 17, 1482–1490. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.H.; Lee, M.J.; Lee, W.-Y.; Pyo, J.; Shin, M.-S.; Hwang, G.S.; Shin, D.; Kim, C.E.; Park, E.-S.; Kang, K.S. Hair Growth Stimulation Effect of Centipeda minima Extract: Identification of Active Compounds and Anagen-Activating Signaling Pathways. Biomolecules 2021, 11, 976. [Google Scholar] [CrossRef] [PubMed]
- Plasari, G.; Edelmann, S.; Högger, F.; Dusserre, Y.; Mermod, N.; Calabrese, A. Nuclear Factor I-C Regulates TGF-β-dependent Hair Follicle Cycling. J. Biol. Chem. 2010, 285, 34115–34125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ru, J.; Li, P.; Wang, J.; Zhou, W.; Li, B.; Huang, C.; Li, P.; Guo, Z.; Tao, W.; Yang, Y.; et al. TCMSP: A database of systems pharmacology for drug discovery from herbal medicines. J. Cheminform. 2014, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Yuan, X.; Tang, Y.; Zhao, Z.; Liu, F.; Shi, W.; Zhang, Y.; Li, J. Identification and verification of EOMEs regulated network in Alopecia areata. Int. Immunopharmacol. 2020, 84, 106544. [Google Scholar] [CrossRef]
- Zhang, R.-Z.; Yu, S.-J.; Bai, H.; Ning, K. TCM-Mesh: The database and analytical system for network pharmacology analysis for TCM preparations. Sci. Rep. 2017, 7, 2821. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Guo, F.; Wang, Y.; Li, C.; Zhang, X.; Li, H.; Diao, L.; Gu, J.; Wang, W.; Li, D.; et al. BATMAN-TCM: A Bioinformatics Analysis Tool for Molecular mechANism of Traditional Chinese Medicine. Sci. Rep. 2016, 6, 21146. [Google Scholar] [CrossRef] [Green Version]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, L.; Reygagne, P.; Benech, P.; Jean-Louis, F.; Scalvino, S.; So, S.L.K.; Hamidou, Z.; Bianovici, S.; Pouch, J.; Ducos, B.; et al. Study of gene expression alteration in male androgenetic alopecia: Evidence of predominant molecular signalling pathways. Br. J. Dermatol. 2017, 177, 1322–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dincer, D.; Tanacan, E.; Ozkan, C.K. Efficacy of systemic minoxidil and tofacitinib combination in treatment-resistant alopecia universalis. J. Cosmet. Dermatol. 2021, 20, 1807–1809. [Google Scholar] [CrossRef] [PubMed]
- Lauer, A.M.; Schrode, K.M. Sex bias in basic and preclinical noise-induced hearing loss research. Noise Health 2017, 19, 207–212. [Google Scholar] [CrossRef]
- Kim, Y.Y.; Noh, S.U.; Kim, M.H.; Kim, H.S.; Kang, H.; Kim, H.O.; Park, Y.M. Effects of topical application of EGCG on testosterone-induced hair loss in a mouse model. Exp. Dermatol. 2011, 20, 1015–1017. [Google Scholar] [CrossRef]
- Tang, Y.; Wang, C.; Desamero, M.J.M.; Kok, M.K.; Chambers, J.K.; Uchida, K.; Kominami, Y.; Ushio, H.; Cervancia, C.; Estacio, M.A.; et al. The Philippines stingless bee propolis promotes hair growth through activation of Wnt/β-catenin signaling pathway. Exp. Anim. 2023, 72, 132–139. [Google Scholar] [CrossRef]
- Miyata, S.; Oda, Y.; Matsuo, C.; Kumura, H.; Kobayashi, K. Stimulatory Effect of Brazilian Propolis on Hair Growth through Proliferation of Keratinocytes in Mice. J. Agric. Food Chem. 2014, 62, 11854–11861. [Google Scholar] [CrossRef]
- Wang, H.; Liu, G.; Yang, X.; Song, Z.; Zhong, H.; Feng, L.; Chen, Y.; Shi, W.; Hao, F. Topical application of 8DSS induces anagen hair growth in telogenic mouse skin. Int. J. Clin. Exp. Med. 2016, 9, 12584–12593. [Google Scholar]
- Kim, B.H.; Lee, W.-Y.; Trinh, T.A.; Pyo, J.S.; Lee, S.; Kim, C.-E.; Lee, D.H.; Park, E.-S.; Kang, K.S. Hair Growth Effect of Emulsion Extracted Brevilin A, a JAK3 Inhibitor, from Centipeda minima. Processes 2020, 8, 767. [Google Scholar] [CrossRef]
- Meephansan, J.; Thummakriengkrai, J.; Ponnikorn, S.; Yingmema, W.; Deenonpoe, R.; Suchonwanit, P. Efficacy of topical tofacitinib in promoting hair growth in non-scarring alopecia: Possible mechanism via VEGF induction. Arch. Dermatol. Res. 2017, 309, 729–738. [Google Scholar] [CrossRef]
- Hu, Z.-Q.; Hou, C.; Miao, Y.; Wang, J.; Wang, X.; Chen, C.-Y. Collagenase IV plays an important role in regulating hair cycle by inducing VEGF, IGF-1, and TGF-β expression. Drug Des. Dev. Ther. 2015, 9, 5373–5383. [Google Scholar] [CrossRef] [Green Version]
- Semon, H.C. An Atlas of the Commoner Skin Diseases: With 120 Plates Reproduced by Direct Colour Photography from the Living Subject; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- MacDonald, O. Alopecia. Br. J. Gen. Pract. 2014, 64, 529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.; Cho, N.; Kim, K.K. Development and Evaluation of the Herbal Medicine for Hair Growth-promoting Activity. KSBB J. 2016, 31, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Moghetti, P.; Castello, R.; Magnani, C.M.; Tosi, F.; Negri, C.; Armanini, D.; Bellotti, G.; Muggeo, M. Clinical and hormonal effects of the 5 alpha-reductase inhibitor finasteride in idiopathic hirsutism. J. Clin. Endocrinol. Metab. 1994, 79, 1115–1121. [Google Scholar]
- Randall, V.A. Androgens and hair: A biological paradox with clinical consequences. Testosterone 2012, 154–176. [Google Scholar] [CrossRef]
- Zhao, J.; Harada, N.; Okajima, K. Dihydrotestosterone inhibits hair growth in mice by inhibiting insulin-like growth factor-I production in dermal papillae. Growth Horm. IGF Res. 2011, 21, 260–267. [Google Scholar] [CrossRef]
- Herman, A.; Herman, A. Mechanism of action of herbs and their active constituents used in hair loss treatment. Fitoterapia 2016, 114, 18–25. [Google Scholar] [CrossRef]
- Avram, M.R.; Cole, J.P.; Chase, C.; Gandelman, M.; Haber, R.; Knudsen, R.; Leavitt, M.L.; Leonard, R.T.; Puig, C.J.; Rose, P.T.; et al. The potential role of minoxidil in the hair transplantation setting. Dermatol. Surg. 2002, 28, 894–900. [Google Scholar]
- Duque-Estrada, B.; Vincenzi, C.; Misciali, C.; Tosti, A. Alopecia secondary to mesotherapy. J. Am. Acad. Dermatol. 2009, 61, 707–709. [Google Scholar] [CrossRef]
- Hagemann, T.; Schlutter-Bohmer, B.; Allam, J.-P.; Bieber, T.; Novak, N. Positive lymphocyte transformation test in a patient with allergic contact dermatitis of the scalp after short-term use of topical minoxidil solution. Contact Dermat. 2005, 53, 53–55. [Google Scholar] [CrossRef]
- McClellan, K.J.; Markham, A. Finasteride. Drugs 1999, 57, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Bannwarth, B.; Kostine, M.; Poursac, N. A pharmacokinetic and clinical assessment of tofacitinib for the treatment of rheumatoid arthritis. Expert Opin. Drug Metab. Toxicol. 2013, 9, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Fleischmann, R. Kinase inhibitors: A new approach to rheumatoid arthritis treatment. Curr. Opin. Rheumatol. 2010, 22, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Harel, S.; Higgins, C.A.; Cerise, J.E.; Dai, Z.; Chen, J.C.; Clynes, R.; Christiano, A.M. Pharmacologic inhibition of JAK-STAT signaling promotes hair growth. Sci. Adv. 2015, 1, e1500973. [Google Scholar] [CrossRef] [Green Version]
- Jabbari, A.; Sansaricq, F.; Cerise, J.; Chen, J.; Bitterman, A.; Ulerio, G.; Borbon, J.; Clynes, R.; Christiano, A.; Mackay-Wiggan, J. An Open-Label Pilot Study to Evaluate the Efficacy of Tofacitinib in Moderate to Severe Patch-Type Alopecia Areata, Totalis, and Universalis. J. Investig. Dermatol. 2018, 138, 1539–1545. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sense (5′→3′) | Antisense (5′→3′) |
|---|---|---|
| Vegfa | CTGGATATGTTTGACTGCTGTGGA | CTGGATATGTTTGACTGCTGTGGA |
| Igf1 | CACTGACATGCCCAAGACTCAGA | TCCGAGTTGCCTCCGTTACC |
| Tgfb1 | CTC CCG TGG CTT CTA GTG C | GCC TTA GTT TGG ACA GGA TCT G |
| Actb | TTCCAGCCTTCCTTCTTG | GGAGCCAGAGCAGTAATC |
| Arnicolide D | Arnicolide C | Microhelenin C | Brevilin A | |
|---|---|---|---|---|
| Content (µg/mL) | 235.9 | 91.8 | 33.2 | 549.3 |
| Term | Overlap | Adjusted p-Value | Combined Score | Genes |
|---|---|---|---|---|
| Estrogen signaling pathway | 4/137 | 0.002401 | 156.9739 | SRC;PRKCD;PGR;ESR1 |
| MAPK signaling pathway | 5/295 | 0.003058 | 86.22343 | CACNA1I;TGFB2;PDGFB;PRKCA;CACNA1G |
| Cytokine–cytokine receptor interaction | 5/294 | 0.003058 | 86.68829 | IL10;IL4;CX3CR1;TGFB2;TNFSF11 |
| Osteoclast differentiation | 3/127 | 0.010617 | 86.71594 | TGFB2;TNFSF11;NOX1 |
| Chemokine signaling pathway | 3/190 | 0.020279 | 47.03707 | CX3CR1;SRC;PRKCD |
| Focal adhesion | 3/199 | 0.022327 | 43.75158 | SRC;PDGFB;PRKCA |
| Rap1 signaling pathway | 3/206 | 0.02344 | 41.43427 | SRC;PDGFB;PRKCA |
| Fc epsilon RI signaling pathway | 2/68 | 0.023807 | 82.16884 | IL4;PRKCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baek, J.Y.; Kim, B.H.; Kim, D.-W.; Lee, W.-Y.; Kim, C.E.; Kim, H.-Y.; Pyo, J.; Park, E.-S.; Kang, K.S. Hair Growth Effect of DN106212 in C57BL/6 Mouse and Its Network Pharmacological Mechanism of Action. Curr. Issues Mol. Biol. 2023, 45, 5071-5083. https://doi.org/10.3390/cimb45060322
Baek JY, Kim BH, Kim D-W, Lee W-Y, Kim CE, Kim H-Y, Pyo J, Park E-S, Kang KS. Hair Growth Effect of DN106212 in C57BL/6 Mouse and Its Network Pharmacological Mechanism of Action. Current Issues in Molecular Biology. 2023; 45(6):5071-5083. https://doi.org/10.3390/cimb45060322
Chicago/Turabian StyleBaek, Ji Yun, Byoung Ha Kim, Dong-Wook Kim, Won-Yung Lee, Chang Eop Kim, Hyun-Young Kim, Jaesung Pyo, Eun-Seok Park, and Ki Sung Kang. 2023. "Hair Growth Effect of DN106212 in C57BL/6 Mouse and Its Network Pharmacological Mechanism of Action" Current Issues in Molecular Biology 45, no. 6: 5071-5083. https://doi.org/10.3390/cimb45060322