

Genetic Diversity Analysis of Banana Cultivars (Musa sp.) in Saudi Arabia Based on AFLP Marker
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Site
2.2. Sample Collection
2.3. DNA Extraction
2.4. AFLP Protocol
2.5. Data Analysis
3. Results
3.1. Fragment Analysis and Band Scoring
3.2. Genetic Polymorphism and Diversity
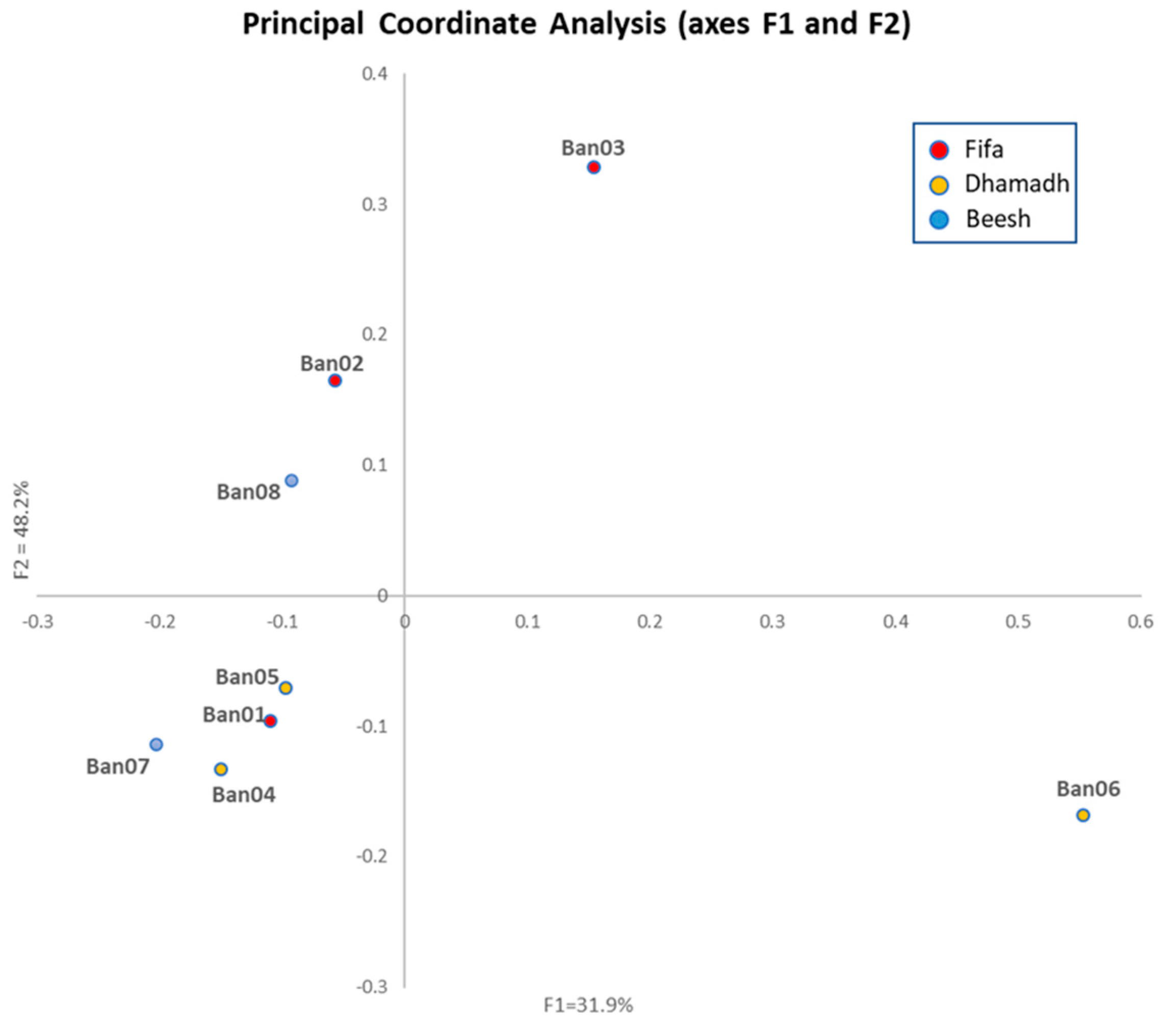
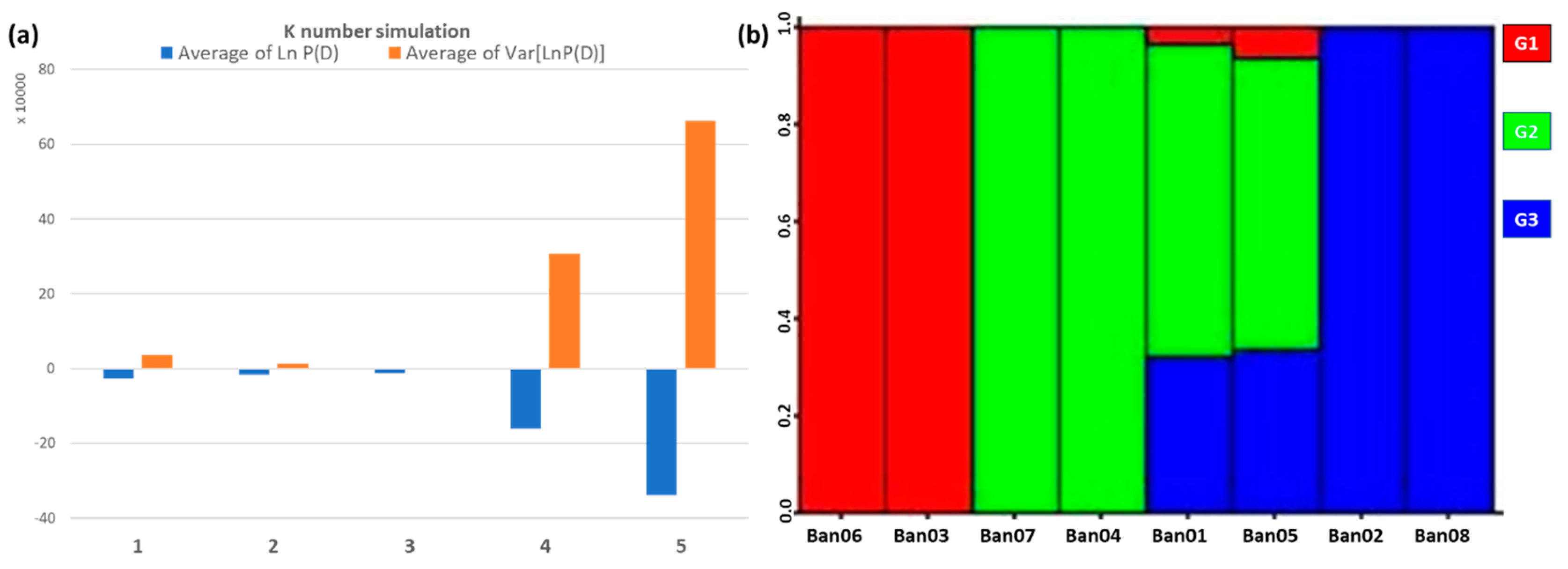
3.3. Population Structure
3.4. Genetic Differentiation and Geographical Influence
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cronquist, A. The Evolution and Classification of Flowering Plants, 2nd ed.; New York Botanical Garden: Bronx, NY, USA, 1988; ISBN 978-0-89327-332-3. [Google Scholar]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#search/banana (accessed on 30 October 2022).
- Christelová, P.; De Langhe, E.; Hřibová, E.; Čížková, J.; Sardos, J.; Hušáková, M.; Van den houwe, I.; Sutanto, A.; Kepler, A.K.; Swennen, R.; et al. Molecular and Cytological Characterization of the Global Musa Germplasm Collection Provides Insights into the Treasure of Banana Diversity. Biodivers. Conserv. 2017, 26, 801–824. [Google Scholar] [CrossRef] [Green Version]
- Abd El-Moneim, D.; Dawood, M.F.A.; Moursi, Y.S. Positive and negative effects of nanoparticles on agricultural crops. Nanotechnol. Environ. Eng. 2021, 6, 21. [Google Scholar] [CrossRef]
- Kodym, A.; Zapata-Arias, F.J. Natural Light as an Alternative Light Source for the in Vitro Culture of Banana (Musa Acuminata Cv. ‘Grande Naine’). Plant Cell Tissue Organ Cult. 1998, 55, 141–145. [Google Scholar] [CrossRef]
- Mahadev, S.R.; Kathithachalam, A.; Marimuthu, M. An Efficient Protocol for Large-Scale Plantlet Production from Male Floral Meristems of Musa Spp. Cultivars Virupakshi and Sirumalai. Vitr. Cell. Dev. Biol. Plant 2011, 47, 611–617. [Google Scholar] [CrossRef]
- Heslop-Harrison, J.S.; Schwarzacher, T. Domestication, Genomics and the Future for Banana. Ann. Bot. 2007, 100, 1073–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Langhe, E.; Vrydaghs, L.; De Maret, P.; Perrier, X.; Denham, T. Why Bananas Matter: An Introduction to the History of Banana Domestication. Ethnobot. Res. Appl. 2009, 7, 165. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, R. Morphological Variation in Musa Germplasm. Genet. Resour. Crop. Evol. 1997, 44, 393–404. [Google Scholar] [CrossRef]
- Pillay, M.; Ogundiwin, E.; Nwakanma, D.C.; Ude, G.; Tenkouano, A. Analysis of Genetic Diversity and Relationships in East African Banana Germplasm. Theor. Appl. Genet. 2001, 102, 965–970. [Google Scholar] [CrossRef]
- Pollefeys, P.; Sharrock, S.; Arnaud, E. Preliminary Analysis of the Literature on the Distribution of Wild Musa Species Using MGIS and DIVA-GIS; INIBAP-International Plant Genetic Resources Institute(IPGRI): Montpellier, France, 2004. [Google Scholar]
- Sardos, J.; Perrier, X.; Doležel, J.; Hřibová, E.; Christelová, P.; Van den houwe, I.; Kilian, A.; Roux, N. DArT Whole Genome Profiling Provides Insights on the Evolution and Taxonomy of Edible Banana (Musa Spp.). Ann. Bot 2016, 118, 1269–1278. [Google Scholar] [CrossRef] [Green Version]
- Lorenzen, J.; Hearne, S.; Mbanjo, G.; Nyine, M.; Close, T. Use of Molecular Markers in Banana and Plantain Improvement. Acta Hortic. 2011, 231–236. [Google Scholar] [CrossRef]
- Hinge, V.R.; Shaikh, I.M.; Chavhan, R.L.; Deshmukh, A.S.; Shelake, R.M.; Ghuge, S.A.; Dethe, A.M.; Suprasanna, P.; Kadam, U.S. Assessment of Genetic Diversity and Volatile Content of Commercially Grown Banana (Musa Spp.) Cultivars. Sci. Rep. 2022, 12, 7979. [Google Scholar] [CrossRef] [PubMed]
- Ruangsuttapha, S.; Eimert, K.; Schröder, M.-B.; Silayoi, B.; Denduangboripant, J.; Kanchanapoom, K. Molecular Phylogeny of Banana Cultivars from Thailand Based on HAT-RAPD Markers. Genet. Resour. Crop. Evol. 2007, 54, 1565–1572. [Google Scholar] [CrossRef]
- D’Hont, A.; Denoeud, F.; Aury, J.-M.; Baurens, F.-C.; Carreel, F.; Garsmeur, O.; Noel, B.; Bocs, S.; Droc, G.; Rouard, M.; et al. The Banana (Musa Acuminata) Genome and the Evolution of Monocotyledonous Plants. Nature 2012, 488, 213–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esmail, S.M.; Aboulila, A.A.; Abd El-Moneim, D. Variation in several pathogenesis-related (PR) protein genes in wheat (Triticum aestivum) involved in defense against Puccinia striiformis f. sp. tritici. Physiol. Mol. Plant Pathol. 2020, 112, 101545. [Google Scholar] [CrossRef]
- Drapal, M.; de Carvalho, E.B.; Rouard, M.; Amah, D.; Sardos, J.; Van den Houwe, I.; Brown, A.; Roux, N.; Swennen, R.; Fraser, P.D. Metabolite Profiling Characterises Chemotypes of Musa Diploids and Triploids at Juvenile and Pre-Flowering Growth Stages. Sci. Rep. 2019, 9, 4657. [Google Scholar] [CrossRef] [Green Version]
- Ude, G.; Pillay, M.; Nwakanma, D.; Tenkouano, A. Analysis of Genetic Diversity and Sectional Relationships in Musa Using AFLP Markers. Theor. Appl. Genet. 2002, 104, 1239–1245. [Google Scholar] [CrossRef]
- Ude, G.; Pillay, M.; Nwakanma, D.; Tenkouano, A. Genetic Diversity in Musa Acuminata Colla and Musa Balbisiana Colla and Some of Their Natural Hybrids Using AFLP Markers. Theor. Appl. Genet. 2002, 104, 1246–1252. [Google Scholar] [CrossRef]
- Loh, J.P.; Kiew, R.; Set, O.; Gan, L.H.; Gan, Y.-Y. Amplified Fragment Length Polymorphism Fingerprinting of 16 Banana Cultivars (Musa Cvs.). Mol. Phylogenet. Evol. 2000, 17, 360–366. [Google Scholar] [CrossRef]
- Khemira, H.; Mars, M. Fig Production in Subtropical South-Western Saudi Arabia. Acta Hortic. 2017, 169–172. [Google Scholar] [CrossRef]
- Safhi, F.A.; ALshamrani, S.M.; Jalal, A.S.; Abd El-Moneim, D.; Alyamani, A.A.; Ibrahim, A.A. Genetic Characterization of Some Saudi Arabia’s Accessions from Commiphora gileadensis Using Physio-Biochemical Parameters, Molecular Markers, DNA Barcoding Analysis and Relative Gene Expression. Genes 2022, 13, 2099. [Google Scholar] [CrossRef]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Friters, A.; Pot, J.; Paleman, J.; Kuiper, M.; et al. AFLP: A New Technique for DNA Fingerprinting. Nucl. Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [Green Version]
- Bonin, A.; Ehrich, D.; Manel, S. Statistical Analysis of Amplified Fragment Length Polymorphism Data: A Toolbox for Molecular Ecologists and Evolutionists. Mol. Ecol. 2007, 16, 3737–3758. [Google Scholar] [CrossRef]
- Abd El-Moneim, D.A.; Mohamed, I.N.; Belal, A.H.; Atta, M.E. Screening bread wheat genotypes for drought tolerance: 1-Germination, radical growth and mean performance of yield and its components. Ann. Agric. Sci. 2008, 53, 171–181. [Google Scholar]
- Schluter, P.M.; Harris, S.A. Analysis of Multilocus Fingerprinting Data Sets Containing Missing Data. Mol. Ecol Notes 2006, 6, 569–572. [Google Scholar] [CrossRef]
- Mesfer, A.S.; Safhi, F.A.; Alshaya, D.S.; Ibrahim, A.A.; Mansour, H.; Abd El Moneim, D. Genetic diversity using biochemical, physiological, karyological and molecular markers of Sesamum indicum L. Front. Genet. 2022, 13, 1035977. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E.L. Arlequin Suite Ver 3.5: A New Series of Programs to Perform Population Genetics Analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Moneim, D.; ELsarag, E.I.S.; Aloufi, S.; El-Azraq, A.M.; ALshamrani, S.M.; Safhi, F.A.A.; Ibrahim, A.A. Quinoa (Chenopodium quinoa Willd.): Genetic Diversity According to ISSR and SCoT Markers, Relative Gene Expression, and Morpho-Physiological Variation under Salinity Stress. Plants 2021, 10, 2802. [Google Scholar] [CrossRef] [PubMed]
- Kaemmer, D.; Afza, R.; Weising, K.; Kahl, G.; Novak, F.J. Oligonucleotide and Amplification Fingerprinting of Wild Species and Cultivars of Banana (Musa Spp.). Nat. Biotechnol. 1992, 10, 1030–1035. [Google Scholar] [CrossRef] [PubMed]
- Wong, C. Genetic Diversity of the Wild Banana Musa Acuminata Colla in Malaysia as Evidenced by AFLP. Ann. Bot. 2001, 88, 1017–1025. [Google Scholar] [CrossRef] [Green Version]
- Jarret, R.L.; Vuylsteke, D.R.; Gawel, N.J.; Pimentel, R.B.; Dunbar, L.J. Detecting Genetic Diversity in Diploid Bananas Using PCR and Primers from a Highly Repetitive DNA Sequence. Euphytica 1993, 68, 69–76. [Google Scholar] [CrossRef]
- Creste, S.; Tulmann Neto, A.; Vencovsky, R.; de Oliveira Silva, S.; Figueira, A. Genetic Diversity of Musa Diploid and Triploid Accessions from the Brazilian Banana Breeding Program Estimated by Microsatellite Markers. Genet. Resour. Crop. Evol. 2004, 51, 723–733. [Google Scholar] [CrossRef]
- Opara, U.L.; Jacobson, D.; Al-Saady, N.A. Analysis of Genetic Diversity in Banana Cultivars (Musa Cvs.) from the South of Oman Using AFLP Markers and Classification by Phylogenetic, Hierarchical Clustering and Principal Component Analyses. J. Zhejiang Univ. Sci. B 2010, 11, 332–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, F.; Megia, R.; Poerba, Y.S. Genetic Diversity of Musa Balbisiana Colla in Indonesia Based on AFLP Marker. HAYATI J. Biosci. 2014, 21, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-L.; Chiang, T.-Y.; Roux, N.; Hao, G.; Ge, X.-J. Genetic Diversity of Wild Banana (Musa Balbisiana Colla) in China as Revealed by AFLP Markers. Genet. Resour. Crop. Evol. 2007, 54, 1125–1132. [Google Scholar] [CrossRef]
- Ge, X.J.; Liu, M.H.; Wang, W.K.; Schaal, B.A.; Chiang, T.Y. Population Structure of Wild Bananas, Musa Balbisiana, in China Determined by SSR Fingerprinting and CpDNA PCR-RFLP: Population structure of wild bananas. Mol. Ecol. 2005, 14, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Bonin, A. Explorative Genome Scan to Detect Candidate Loci for Adaptation Along a Gradient of Altitude in the Common Frog (Rana Temporaria). Mol. Biol. Evol. 2006, 23, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Magdy, M.; Werner, O.; McDaniel, S.; Goffinet, B.; Ros, R. Genomic Scanning Using AFLP to Detect Loci under Selection in the Moss Funaria Hygrometrica along a Climate Gradient in the Sierra Nevada Mountains, Spain. Plant. Biol. 2016, 18, 280–288. [Google Scholar] [CrossRef]
- Magdy, M.; Eshak, M.G.; Rashed, M.A.-S. Genetic Structure of Mugil Cephalus L. Populations from the Northern Coast of Egypt. Vet. World 2016, 9, 53. [Google Scholar] [CrossRef] [Green Version]
- Storz, J.F. INVITED REVIEW: Using Genome Scans of DNA Polymorphism to Infer Adaptive Population Divergence: Genome scans and adaptive population divergence. Mol. Ecol. 2005, 14, 671–688. [Google Scholar] [CrossRef]
- Schlötterer, C. Hitchhiking Mapping—Functional Genomics from the Population Genetics Perspective. Trends Genet. 2003, 19, 32–38. [Google Scholar] [CrossRef]
- Hohenlohe, P.A.; Catchen, J.; Cresko, W.A. Population Genomic Analysis of Model and Nonmodel Organisms Using Sequenced RAD Tags. In Data Production and Analysis in Population Genomics; Pompanon, F., Bonin, A., Eds.; Methods in Molecular BiologyTM; Humana Press: Totowa, NJ, USA, 2012; Volume 888, pp. 235–260. ISBN 978-1-61779-869-6. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Sample ID | Cultivar Name | Species Name | Location |
|---|---|---|---|
| Ban01 | Red Banana | Musa acuminata | Fifa |
| Ban02 | Indian Banana | Musa acuminata | Fifa |
| Ban03 | Baladi Banana | Musa acuminata | Fifa |
| Ban04 | American Banana | Musa paradisiaca | Dhamadh |
| Ban05 | Red Banana | Musa acuminata | Dhamadh |
| Ban06 | French Banana | Musa acuminata | Dhamadh |
| Ban07 | American Banana | Musa paradisiaca | Beesh |
| Ban08 | Indian Banana | Musa acuminata | Beesh |
| Type | EcoRI | 5′-Sequence-3′ | MseI | 5′-Sequence-′3 |
|---|---|---|---|---|
| Adaptors | A1 | CTCGTAGACTGCGTACC | A1 | GACGATGAGTCCTGAG |
| A2 | AATTGGTACGCAGTC | A2 | TACTCAGGACTCAT | |
| 1st PCR | +A | GACTGCGTACCAATTCA | +C | GATGAGTCCTGAGTAC |
| Selective PCR | +ACA | FAM-GACTGCGTACCAATTCAA | +CTC | GATGAGTCCTGAGCTC |
| +AGG | HEX-GACTGCGTACCAATTCAG | +CTA | GATGAGTCCTGAGCTA | |
| +ATA | CY3-GACTGCGTACCAATTCAA | +CTT | GATGAGTCCTGAGCTT |
| Parameter/Location |
Fifa (Ban01–03) | Dhamadh (Ban04–06) |
Beesh (Ban07–08) | Overall |
|---|---|---|---|---|
| Number of polymorphic bands | 862 | 963 | 571 | 1008 |
| Mean effective number of alleles (ne) | 1.470 | 1.525 | 1.390 | 1.461 |
| Standard deviation (ne) | 0.010 | 0.010 | 0.013 | 0.006 |
| Mean heterozygosity (He) | 0.261 | 0.292 | 0.195 | 0.249 |
| Standard deviation (He) | 0.006 | 0.006 | 0.006 | 0.003 |
| Jaccard * | Ban01 | Ban02 | Ban03 | Ban04 | Ban05 | Ban06 | Ban07 | Ban08 |
|---|---|---|---|---|---|---|---|---|
| Ban01 | 0 | 0.571 | 0.636 | 0.528 | 0.492 | 0.761 | 0.522 | 0.541 |
| Ban02 | 0.571 | 0 | 0.62 | 0.565 | 0.56 | 0.755 | 0.577 | 0.483 |
| Ban03 | 0.636 | 0.62 | 0 | 0.642 | 0.618 | 0.71 | 0.647 | 0.636 |
| Ban04 | 0.528 | 0.565 | 0.642 | 0 | 0.517 | 0.773 | 0.447 | 0.551 |
| Ban05 | 0.492 | 0.56 | 0.618 | 0.517 | 0 | 0.75 | 0.513 | 0.532 |
| Ban06 | 0.761 | 0.755 | 0.71 | 0.773 | 0.75 | 0 | 0.812 | 0.762 |
| Ban07 | 0.522 | 0.577 | 0.647 | 0.447 | 0.513 | 0.812 | 0 | 0.556 |
| Ban08 | 0.541 | 0.483 | 0.636 | 0.551 | 0.532 | 0.762 | 0.556 | 0 |
| Dataset | Comparison Scheme | Groups | Among Groups (Va) | Within Groups (Vb) | ΦST (p < 0.00) |
|---|---|---|---|---|---|
| All AFLP | Among locations | 3 | 7% | 93% | 0.07 |
| Among cultivars | 4 | 28.95% | 71.05% | 0.28 | |
| Outliers | Among cultivars | 4 | 100% | 0% | 1.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Safhi, F.A.; Alshamrani, S.M.; Alshaya, D.S.; Hussein, M.A.A.; Abd El-Moneim, D. Genetic Diversity Analysis of Banana Cultivars (Musa sp.) in Saudi Arabia Based on AFLP Marker. Curr. Issues Mol. Biol. 2023, 45, 1810-1819. https://doi.org/10.3390/cimb45030116
Safhi FA, Alshamrani SM, Alshaya DS, Hussein MAA, Abd El-Moneim D. Genetic Diversity Analysis of Banana Cultivars (Musa sp.) in Saudi Arabia Based on AFLP Marker. Current Issues in Molecular Biology. 2023; 45(3):1810-1819. https://doi.org/10.3390/cimb45030116
Chicago/Turabian StyleSafhi, Fatmah Ahmed, Salha Mesfer Alshamrani, Dalal Sulaiman Alshaya, Mohammed A. A. Hussein, and Diaa Abd El-Moneim. 2023. "Genetic Diversity Analysis of Banana Cultivars (Musa sp.) in Saudi Arabia Based on AFLP Marker" Current Issues in Molecular Biology 45, no. 3: 1810-1819. https://doi.org/10.3390/cimb45030116