

Genome-Wide Identification and Expression Analysis of Calmodulin-Like Gene Family in Paspalums vaginatium Revealed Their Role in Response to Salt and Cold Stress
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of CML Genes in the Paspalum vaginatum Genome
2.2. Sequence Alignment and Phylogenetic Analysis
2.3. Characterization and Assessments of Gene Structure and Conserved Domains of PvCML Genes
2.4. Collinearity and Chromosomal Locations of PvCML Genes
2.5. Analysis of Putative Cis-Acting Elements
2.6. Transcription Expression Analysis of PvCML Genes under the Stress and Cold Treatments
2.7. Plant Materials and Treatments
2.8. Gene Expression Analysis Using Real-Time PCR
3. Results
3.1. Identification and Characterization of CML Proteins in the P. vaginatum Genome
3.2. Phylogenetic Alignments of CMLs among P. vaginatum, Rice and Aribidopsis
3.3. Conserved Motifs and Gene Structure of the PvCMLs
3.4. Chromosomal Location and Collinearity of PvCMLs
3.5. Putative Cis-Element Analysis in Promoter Regions of PvCML Genes
3.6. Transcriptome of PvCMLs under Salt and Cold Treatment Conditions
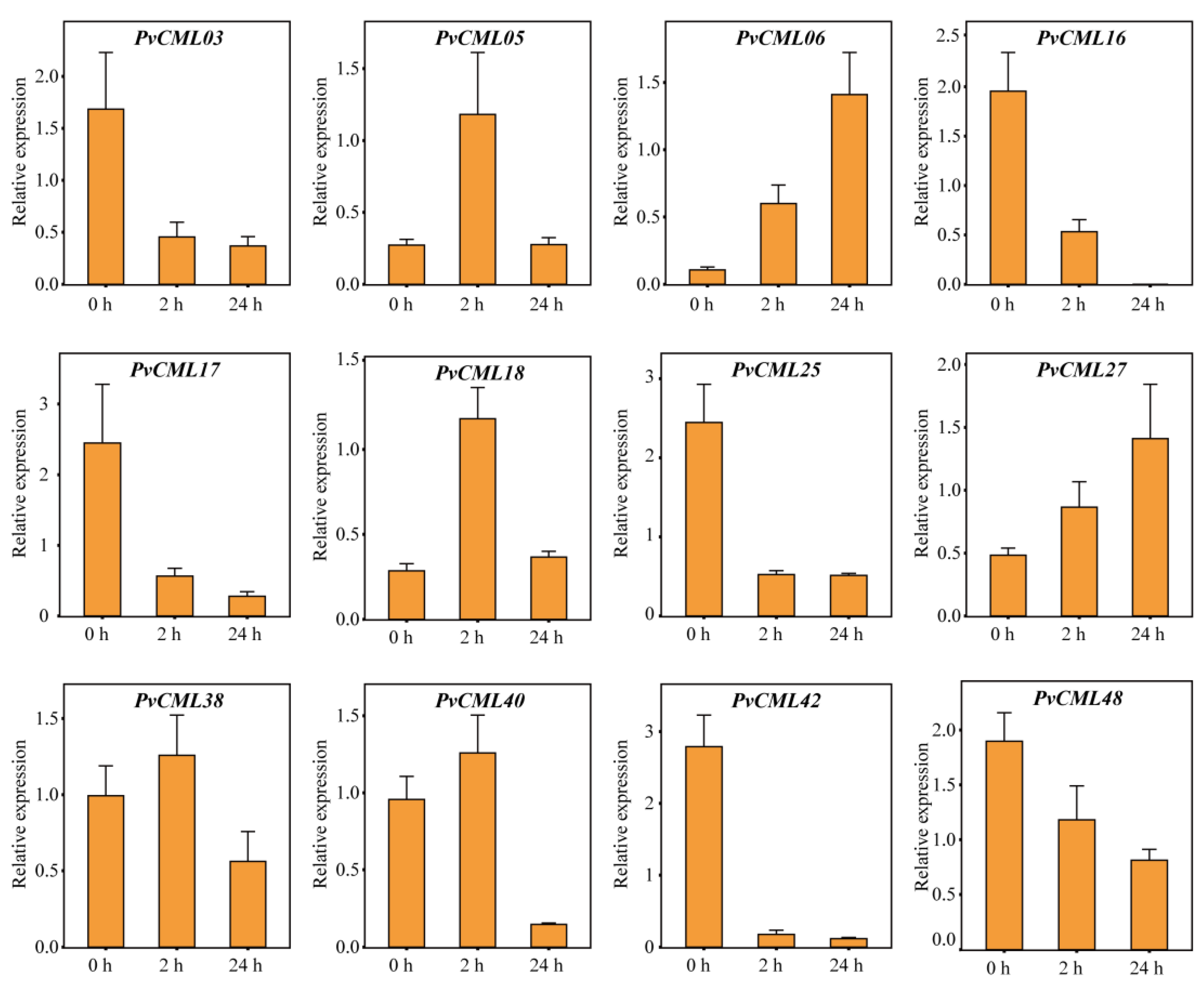
3.7. qRT–PCR of PvCMLs under Salt and Cold Treatment Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, J.-K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, W.; Wang, C.; Gao, Q.; Li, L.; Luan, S. Calcium spikes, waves and oscillations in plant development and biotic interactions. Nat. Plants 2020, 6, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Kudla, J.; Batistič, O.; Hashimoto, K. Calcium Signals: The Lead Currency of Plant Information Processing. Plant Cell 2010, 22, 541–563. [Google Scholar] [CrossRef] [PubMed]
- La Verde, V.; Dominici, P.; Astegno, A. Towards Understanding Plant Calcium Signaling through Calmodulin-Like Proteins: A Biochemical and Structural Perspective. Int. J. Mol. Sci. 2018, 19, 1331. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, K.; Kudla, J. Calcium decoding mechanisms in plants. Biochimie 2011, 93, 2054–2059. [Google Scholar] [CrossRef]
- Perochon, A.; Aldon, D.; Galaud, J.-P.; Ranty, B. Calmodulin and calmodulin-like proteins in plant calcium signaling. Biochimie 2011, 93, 2048–2053. [Google Scholar] [CrossRef]
- Webb, A.A.; McAinsh, M.R.; Taylor, J.E.; Hetherington, A.M. Calcium Ions as Intracellular Second Messengers in Higher Plants. Adv. Bot. Res. 1996, 22, 45–96. [Google Scholar] [CrossRef]
- Hetherington, A.M.; Brownlee, C. The generation of Ca2+ signals in plants. Annu. Rev. Plant Biol. 2004, 55, 401–427. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Poovaiah, B. Calcium/calmodulin-mediated signal network in plants. Trends Plant Sci. 2003, 8, 505–512. [Google Scholar] [CrossRef]
- Edel, K.H.; Marchadier, E.; Brownlee, C.; Kudla, J.; Hetherington, A.M. The Evolution of Calcium-Based Signalling in Plants. Curr. Biol. 2017, 27, R667–R679. [Google Scholar] [CrossRef]
- Luan, S.; Kudla, J.; Rodriguez-Concepcion, M.; Yalovsky, S.; Gruissem, W. Calmodulins and Calcineurin B–like Proteins: Cal-cium sensors for specific signal response coupling in plants. Plant Cell 2002, 14, S389–S400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanta, T.K.; Yadav, D.; Khan, A.L.; Hashem, A.; Abd_Allah, E.; Al-Harrasi, A. Molecular Players of EF-hand Containing Calcium Signaling Event in Plants. Int. J. Mol. Sci. 2019, 20, 1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormack, E.; Braam, J. Calmodulins and related potential calcium sensors of Arabidopsis. New Phytol. 2003, 159, 585–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeFalco, T.A.; Bender, K.W.; Snedden, W.A. Breaking the code: Ca2+ sensors in plant signalling. Biochem. J. 2010, 425, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Dunand, C.; Snedden, W.; Galaud, J.-P. CaM and CML emergence in the green lineage. Trends Plant Sci. 2015, 20, 483–489. [Google Scholar] [CrossRef]
- Zhu, X.; Perez, M.; Aldon, D.; Galaud, J.-P. Respective contribution of CML8 and CML9, two arabidopsis calmodulin-like proteins, to plant stress responses. Plant Signal. Behav. 2017, 12, e1322246. [Google Scholar] [CrossRef] [Green Version]
- Boonburapong, B.; Buaboocha, T. Genome-wide identification and analyses of the rice calmodulin and related potential calcium sensor proteins. BMC Plant Biol. 2007, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Munir, S.; Khan, M.R.G.; Song, J.; Munir, S.; Zhang, Y.; Ye, Z.; Wang, T. Genome-wide identification, characterization and expression analysis of calmodulin-like (CML) proteins in tomato (Solanum lycopersicum). Plant Physiol. Biochem. 2016, 102, 167–179. [Google Scholar] [CrossRef]
- Yadav, M.; Pandey, J.; Chakraborty, A.; Hassan, I.; Kundu, J.K.; Roy, A.; Singh, I.K.; Singh, A. A Comprehensive Analysis of Calmodulin-Like Proteins of Glycine max Indicates Their Role in Calcium Signaling and Plant Defense Against Insect Attack. Front. Plant Sci. 2022, 13, 817950. [Google Scholar] [CrossRef]
- Zeng, H.; Zhang, Y.; Zhang, X.; Pi, E.; Zhu, Y. Analysis of EF-Hand Proteins in Soybean Genome Suggests Their Potential Roles in Environmental and Nutritional Stress Signaling. Front. Plant Sci. 2017, 8, 877. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Tian, J.; Li, S.; Liu, Y.; Feng, T.; Wang, Y.; Li, Y.; Huang, X.; Li, D. Characterization of the Calmodulin/Calmodulin-like Protein (CAM/CML) Family in Ginkgo biloba, and the Influence of an Ectopically Expressed GbCML Gene (Gb_30819) on Seedling and Fruit Development of Transgenic Arabidopsis. Plants 2022, 11, 1506. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, L.; Li, J.; Yang, W.; Ci, J.; Ren, X.; Wang, W.; Wang, Y.; Jiang, L.; Yang, W. Identification and expression analysis revealed drought stress-responsive Calmodulin and Calmodulin-like genes in maize. J. Plant Interact. 2022, 17, 450–461. [Google Scholar] [CrossRef]
- Cai, K.; Kuang, L.; Yue, W.; Xie, S.; Xia, X.; Zhang, G.; Wang, J. Calmodulin and calmodulin-like gene family in barley: Identification, characterization and expression analyses. Front. Plant Sci. 2022, 13, 964888. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Yu, S.; Guo, Z. Calmodulin-Like (CML) Gene Family in Medicago truncatula: Genome-Wide Identification, Characterization and Expression Analysis. Int. J. Mol. Sci. 2020, 21, 7142. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Wang, G.; Zong, M.; Han, S.; Liu, F. Genome-wide identification, and phylogenetic and expression profiling analyses of CaM and CML genes in Brassica rapa and Brassica oleracea. Gene 2018, 677, 232–244. [Google Scholar] [CrossRef] [PubMed]
- Bender, K.W.; Snedden, W.A. Calmodulin-Related Proteins Step Out from the Shadow of Their Namesake. Plant Physiol. 2013, 163, 486–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormack, E.; Tsai, Y.-C.; Braam, J. Handling calcium signaling: Arabidopsis CaMs and CMLs. Trends Plant Sci. 2005, 10, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Chinpongpanich, A.; Limruengroj, K.; Phean-O-Pas, S.; Limpaseni, T.; Buaboocha, T. Expression analysis of calmodulin and calmodulin-like genes from rice, Oryza sativa L. BMC Res. Notes 2012, 5, 625. [Google Scholar] [CrossRef] [Green Version]
- Scholz, S.S.; Vadassery, J.; Heyer, M.; Reichelt, M.; Bender, K.W.; Snedden, W.A.; Boland, W.; Mithöfer, A. Mutation of the Arabidopsis Calmodulin-Like Protein CML37 Deregulates the Jasmonate Pathway and Enhances Susceptibility to Herbivory. Mol. Plant 2014, 7, 1712–1726. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.-Q.; Han, L.-B.; Yang, C.-L.; Wu, X.-M.; Zhong, N.-Q.; Wu, J.-H.; Wang, F.-X.; Wang, H.-Y.; Xia, G.-X. The cotton MYB108 forms a positive feedback regulation loop with CML11 and participates in the defense response against Verticillium dahlia infection. J. Exp. Bot. 2016, 67, 1935–1950. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Cheval, C.; Laohavisit, A.; Hocking, B.; Chiasson, D.; Olsson, T.S.G.; Shirasu, K.; Faulkner, C.; Gilliham, M. A calmodulin-like protein regulates plasmodesmal closure during bacterial immune responses. New Phytol. 2017, 215, 77–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wang, T.; Liu, M.; Sun, W.; Zhang, W.-H. Calmodulin-like gene MtCML40 is involved in salt tolerance by regulating MtHKTs transporters in Medicago truncatula. Environ. Exp. Bot. 2019, 157, 79–90. [Google Scholar] [CrossRef]
- Xu, G.-Y.; Rocha, P.S.C.F.; Wang, M.-L.; Xu, M.-L.; Cui, Y.-C.; Li, L.-Y.; Zhu, Y.-X.; Xia, X. A novel rice calmodulin-like gene, OsMSR2, enhances drought and salt tolerance and increases ABA sensitivity in Arabidopsis. Planta 2011, 234, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Magnan, F.; Ranty, B.; Charpenteau, M.; Sotta, B.; Galaud, J.-P.; Aldon, D.; Ranty, B. Mutations in AtCML9, a calmodulin-like protein from Arabidopsis thaliana, alter plant responses to abiotic stress and abscisic acid. Plant J. 2008, 56, 575–589. [Google Scholar] [CrossRef]
- Sun, Q.; Huang, R.; Zhu, H.; Sun, Y.; Guo, Z. A novel Medicago truncatula calmodulin-like protein (MtCML42) regulates cold tolerance and flowering time. Plant J. 2021, 108, 1069–1082. [Google Scholar] [CrossRef]
- Yu, S.; Wu, J.; Sun, Y.; Zhu, H.; Sun, Q.; Zhao, P.; Huang, R.; Guo, Z. A calmodulin-like protein (CML10) interacts with cytosolic enzymes GSTU8 and FBA6 to regulate cold tolerance. Plant Physiol. 2022, 190, 1321–1333. [Google Scholar] [CrossRef]
- Lee, G.-J.; Carrow, R.N.; Duncan, R.R. Photosynthetic responses to salinity stress of halophytic seashore paspalum ecotypes. Plant Sci. 2004, 166, 1417–1425. [Google Scholar] [CrossRef]
- Wu, P.; Cogill, S.; Qiu, Y.; Li, Z.; Zhou, M.; Hu, Q.; Chang, Z.; Noorai, R.E.; Xia, X.; Saski, C.; et al. Comparative transcriptome profiling provides insights into plant salt tolerance in seashore paspalum (Paspalum vaginatum). BMC Genom. 2020, 21, 131. [Google Scholar] [CrossRef]
- Qi, P.; Eudy, D.; Schnable, J.C.; Schmutz, J.; Raymer, P.L.; Devos, K.M. High Density Genetic Maps of Seashore Paspalum Using Genotyping-By-Sequencing and Their Relationship to The Sorghum Bicolor Genome. Sci. Rep. 2019, 9, 12183. [Google Scholar] [CrossRef] [Green Version]
- Liao, L.; Hu, X.; Hao, J.; Tang, M.; Ren, L.; Pan, L.; Xie, S.; Raymer, P.; Qi, P.; Chen, Z.; et al. The chromosome-based genome of Paspalum vaginatum provides new insights into salt-stress adaptation. bioRxiv 2022. [Google Scholar] [CrossRef]
- Lee, G.-J.; Duncan, R.R.; Carrow, R.N. Salinity Tolerance of Seashore Paspalum Ecotypes: Shoot Growth Responses and Criteria. Hortscience 2004, 39, 1138–1142. [Google Scholar] [CrossRef] [Green Version]
- Goad, D.M.; Baxter, I.; Kellogg, E.A.; Olsen, K.M. Hybridization, polyploidy and clonality influence geographic patterns of diversity and salt tolerance in the model halophyte seashore paspalum (Paspalum vaginatum). Mol. Ecol. 2021, 30, 148–161. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.K.; Juraim, A.S.; Ismail, M.R.; Alam, M.A. The effect of salinity on growth and ion accumulation in six turfgrass species. Plant Omics. 2012, 5, 244–252. [Google Scholar]
- Sun, G.; Wase, N.; Shu, S.; Jenkins, J.; Zhou, B.; Torres-Rodríguez, J.V.; Chen, C.; Sandor, L.; Plott, C.; Yoshinga, Y.; et al. Genome of Paspalum vaginatum and the role of trehalose mediated autophagy in increasing maize biomass. Nat. Commun. 2022, 13, 7731. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.-H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, H.; Bowers, J.E.; Wang, X.; Ming, R.; Alam, M.; Paterson, A.H. Synteny and Collinearity in Plant Genomes. Science 2008, 320, 486–488. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z. PAML 4: Phylogenetic Analysis by Maximum Likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Ngu, D.W.; Carvalho, D.; Liang, Z.; Qiu, Y.; Roston, R.L.; Schnable, J.C. Differentially Regulated Orthologs in Sorghum and the Subgenomes of Maize. Plant Cell 2017, 29, 1938–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- He, X.; Liu, W.; Li, W.; Liu, Y.; Wang, W.; Xie, P.; Kang, Y.; Liao, L.; Qian, L.; Liu, Z.; et al. Genome-wide identification and expression analysis of CaM/CML genes in Brassica napus under abiotic stress. J. Plant Physiol. 2020, 255, 153251. [Google Scholar] [CrossRef]
- Dale, S.; Colin, B.; Jeffrey, F.H. Communicating with Calcium. Plant Cell 1999, 11, 691–706. [Google Scholar]
- Vandelle, E.; Vannozzi, A.; Wong, D.; Danzi, D.; Digby, A.-M.; Santo, S.D.; Astegno, A. Identification, characterization, and expression analysis of calmodulin and calmodulin-like genes in grapevine (Vitis vinifera) reveal likely roles in stress responses. Plant Physiol. Biochem. 2018, 129, 221–237. [Google Scholar] [CrossRef]
- Mohanta, T.K.; Kumar, P.; Bae, H. Genomics and evolutionary aspect of calcium signaling event in calmodulin and calmodulin-like proteins in plants. BMC Plant Biol. 2017, 17, 38. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Damaris, R.N.; Yu, F.; Yang, P. Genome-wide Identification and Expression Analysis of CaM/CML Gene Family in Sacred Lotus (Nelumbo nucifera). Plant Mol. Biol. Rep. 2022, 40, 418–432. [Google Scholar] [CrossRef]
- Bellieny-Rabelo, D.; Oliveira, A.E.A.; Venancio, T.M. Impact of Whole-Genome and Tandem Duplications in the Expansion and Functional Diversification of the F-Box Family in Legumes (Fabaceae). PLoS ONE 2013, 8, e55127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofberger, J.A.; Lyons, E.; Edger, P.P.; Pires, J.C.; Schranz, M.E. Whole Genome and Tandem Duplicate Retention Facilitated Glucosinolate Pathway Diversification in the Mustard Family. Genome Biol. Evol. 2013, 5, 2155–2173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Hao, J.; Pan, L.; Xu, T.; Ren, L.; Chen, Y.; Tang, M.; Liao, L.; Wang, Z. Genome-wide analysis of tandem duplicated genes and their expression under salt stress in seashore paspalum. Front. Plant Sci. 2022, 13, 971999. [Google Scholar] [CrossRef]
- Knight, J.C. Tandem and segmental gene duplication and recombination in the evolution of plant disease resistance genes. Trends Genet. 2004, 20, 113–116. [Google Scholar] [CrossRef]
- Rizzon, C.; Ponger, L.; Gaut, B.S. Striking Similarities in the Genomic Distribution of Tandemly Arrayed Genes in Arabidopsis and Rice. PLoS Comput. Biol. 2006, 2, e115. [Google Scholar] [CrossRef] [Green Version]
- Vanderbeld, B.; Snedden, W.A. Developmental and stimulus-induced expression patterns of Arabidopsis calmodulin-like genes CML37, CML38 and CML39. Plant Mol. Biol. 2007, 64, 683–697. [Google Scholar] [CrossRef]
- Delk, N.A.; Johnson, K.A.; Chowdhury, N.I.; Braam, J. CML24, Regulated in Expression by Diverse Stimuli, Encodes a Potential Ca2+ Sensor That Functions in Responses to Abscisic Acid, Daylength, and Ion Stress. Plant Physiol. 2005, 139, 240–253. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Sun, X.; Duanmu, H.; Zhu, D.; Yu, Y.; Cao, L.; Liu, A.; Jia, B.; Xiao, J.; Zhu, Y. GsCML27, a Gene Encoding a Calcium-Binding Ef-Hand Protein from Glycine soja, Plays Differential Roles in Plant Responses to Bicarbonate, Salt and Osmotic Stresses. PLoS ONE 2015, 10, e0141888. [Google Scholar] [CrossRef] [Green Version]
- Munir, S.; Liu, H.; Xing, Y.; Hussain, S.; Ouyang, B.; Zhang, Y.; Li, H.; Ye, Z. Overexpression of calmodulin-like (ShCML44) stress-responsive gene from Solanum habrochaites enhances tolerance to multiple abiotic stresses. Sci. Rep. 2016, 6, 31772. [Google Scholar] [CrossRef]
- Yang, J.; Ji, L.; Zhu, B.; Yuan, X.; Jin, D.; Xie, G. OsCML16 interacts with a novel CC-NBS-LRR protein OsPi304 in the Ca2+/Mg2+ dependent and independent manner in rice. Biochem. Biophys. Res. Commun. 2018, 504, 346–351. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Locus ID | Length (aa) | MW (kDa) | pI | GRAVY | EF-Hand Domain Number |
|---|---|---|---|---|---|---|
| PvCML01 | Pavag01G036200 | 249 | 26,549.83 | 4.93 | −0.235 | 2 |
| PvCML02 | Pavag01G067400 | 179 | 19,525.77 | 5.66 | −0.444 | 2 |
| PvCML03 | Pavag01G085000 | 155 | 17,270.08 | 3.98 | −0.588 | 4 |
| PvCML04 | Pavag01G238700 | 178 | 20,176.57 | 4.62 | −0.749 | 4 |
| PvCML05 | Pavag01G274800 | 216 | 22,865.16 | 4.54 | −0.289 | 4 |
| PvCML06 | Pavag01G365300 | 189 | 20,387.82 | 4.46 | −0.297 | 3 |
| PvCML07 | Pavag01G374900 | 167 | 17,550.55 | 4.16 | −0.149 | 3 |
| PvCML08 | Pavag02G205000 | 184 | 21,026.19 | 8.76 | −0.649 | 3 |
| PvCML09 | Pavag02G205100 | 98 | 11,265.6 | 5.45 | −0.738 | 1 |
| PvCML10 | Pavag02G351900 | 173 | 19,858.13 | 4.76 | −0.923 | 4 |
| PvCML11 | Pavag02G352100 | 95 | 10,825.18 | 8.18 | −0.674 | 2 |
| PvCML12 | Pavag02G352200 | 120 | 13,500.42 | 9.73 | −0.479 | 2 |
| PvCML13 | Pavag02G392400 | 200 | 21,075.5 | 4.47 | −0.207 | 3 |
| PvCML14 | Pavag03G074400 | 183 | 19,175.09 | 4.48 | −0.538 | 4 |
| PvCML15 | Pavag03G187000 | 239 | 24,717.86 | 4.95 | −0.145 | 4 |
| PvCML16 | Pavag03G287500 | 219 | 23,379.89 | 4.96 | −0.548 | 2 |
| PvCML17 | Pavag03G401100 | 188 | 19,570.96 | 4.88 | −0.233 | 2 |
| PvCML18 | Pavag03G401300 | 202 | 20,647.08 | 4.79 | −0.125 | 4 |
| PvCML19 | Pavag03G405900 | 167 | 17,384.44 | 4.42 | 0.061 | 4 |
| PvCML20 | Pavag03G406300 | 152 | 16,591.57 | 4.2 | −0.296 | 4 |
| PvCML21 | Pavag04G074900 | 230 | 26,143.37 | 4.59 | −0.559 | 3 |
| PvCML22 | Pavag04G268400 | 183 | 19,286.47 | 4.26 | −0.245 | 4 |
| PvCML23 | Pavag05G031700 | 196 | 22,180.57 | 4.48 | −0.645 | 4 |
| PvCML24 | Pavag05G038100 | 313 | 32,468.23 | 6.42 | −0.372 | 2 |
| PvCML25 | Pavag05G038700 | 189 | 19,421.47 | 4.8 | −0.243 | 2 |
| PvCML26 | Pavag05G195800 | 198 | 22,248.02 | 4.57 | −0.366 | 2 |
| PvCML27 | Pavag06G144700 | 263 | 27,960.3 | 4.78 | −0.429 | 4 |
| PvCML28 | Pavag07G020500 | 149 | 16,585.77 | 4.78 | −0.34 | 3 |
| PvCML29 | Pavag07G181000 | 141 | 15,274.29 | 4.53 | −0.025 | 2 |
| PvCML30 | Pavag07G181100 | 141 | 15,343.27 | 4.48 | −0.073 | 2 |
| PvCML31 | Pavag07G181200 | 141 | 15,250.15 | 4.54 | −0.063 | 2 |
| PvCML32 | Pavag07G181300 | 141 | 15,341.34 | 4.53 | −0.021 | 2 |
| PvCML33 | Pavag07G226900 | 243 | 26,137.43 | 6.07 | −0.193 | 2 |
| PvCML34 | Pavag07G229200 | 83 | 8931.05 | 4.29 | −0.15 | 2 |
| PvCML35 | Pavag08G065500 | 206 | 22,885.43 | 4.62 | −0.7 | 4 |
| PvCML36 | Pavag08G137400 | 172 | 18,391.3 | 4.28 | −0.309 | 4 |
| PvCML37 | Pavag09G042600 | 231 | 26,890.24 | 4.95 | −0.685 | 3 |
| PvCML38 | Pavag09G088400 | 227 | 23,777.51 | 5.61 | −0.493 | 4 |
| PvCML39 | Pavag09G124700 | 193 | 20,236.49 | 4.3 | −0.314 | 4 |
| PvCML40 | Pavag09G126600 | 192 | 19,711.92 | 4.7 | −0.184 | 4 |
| PvCML41 | Pavag09G185900 | 154 | 16,812.8 | 4.51 | −0.355 | 2 |
| PvCML42 | Pavag09G246700 | 186 | 19,477.01 | 4.73 | −0.03 | 4 |
| PvCML43 | Pavag10G059200 | 251 | 27,068.03 | 4.51 | −0.524 | 4 |
| PvCML44 | Pavag10G085700 | 99 | 11,520.98 | 9.38 | −0.513 | 2 |
| PvCML45 | Pavag10G188500 | 225 | 25,934.32 | 4.75 | −0.48 | 4 |
| PvCML46 | Pavag10G241600 | 160 | 16,085.05 | 3.87 | 0.316 | 3 |
| PvCML47 | PavagK054400 | 97 | 11,196.58 | 6.41 | −0.635 | 2 |
| PvCML48 | PavagK123100 | 198 | 22,203.98 | 4.57 | −0.368 | 2 |
| PvCML49 | PavagK317500 | 89 | 10,140.33 | 6.41 | −0.741 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.; Chen, J.; Liu, T.; Xiang, L.; Zhou, B.-F. Genome-Wide Identification and Expression Analysis of Calmodulin-Like Gene Family in Paspalums vaginatium Revealed Their Role in Response to Salt and Cold Stress. Curr. Issues Mol. Biol. 2023, 45, 1693-1711. https://doi.org/10.3390/cimb45020109
Yang M, Chen J, Liu T, Xiang L, Zhou B-F. Genome-Wide Identification and Expression Analysis of Calmodulin-Like Gene Family in Paspalums vaginatium Revealed Their Role in Response to Salt and Cold Stress. Current Issues in Molecular Biology. 2023; 45(2):1693-1711. https://doi.org/10.3390/cimb45020109
Chicago/Turabian StyleYang, Meizhen, Jingjin Chen, Tingting Liu, Leilei Xiang, and Biao-Feng Zhou. 2023. "Genome-Wide Identification and Expression Analysis of Calmodulin-Like Gene Family in Paspalums vaginatium Revealed Their Role in Response to Salt and Cold Stress" Current Issues in Molecular Biology 45, no. 2: 1693-1711. https://doi.org/10.3390/cimb45020109