

The Factors Influencing Wildlife to Use Existing Bridges and Culverts in Giant Panda National Park

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Camera Trap Monitoring
2.3. Traffic Volume
2.4. Land Cover
2.5. Statistical Analysis
3. Results
3.1. Richness and Abundance
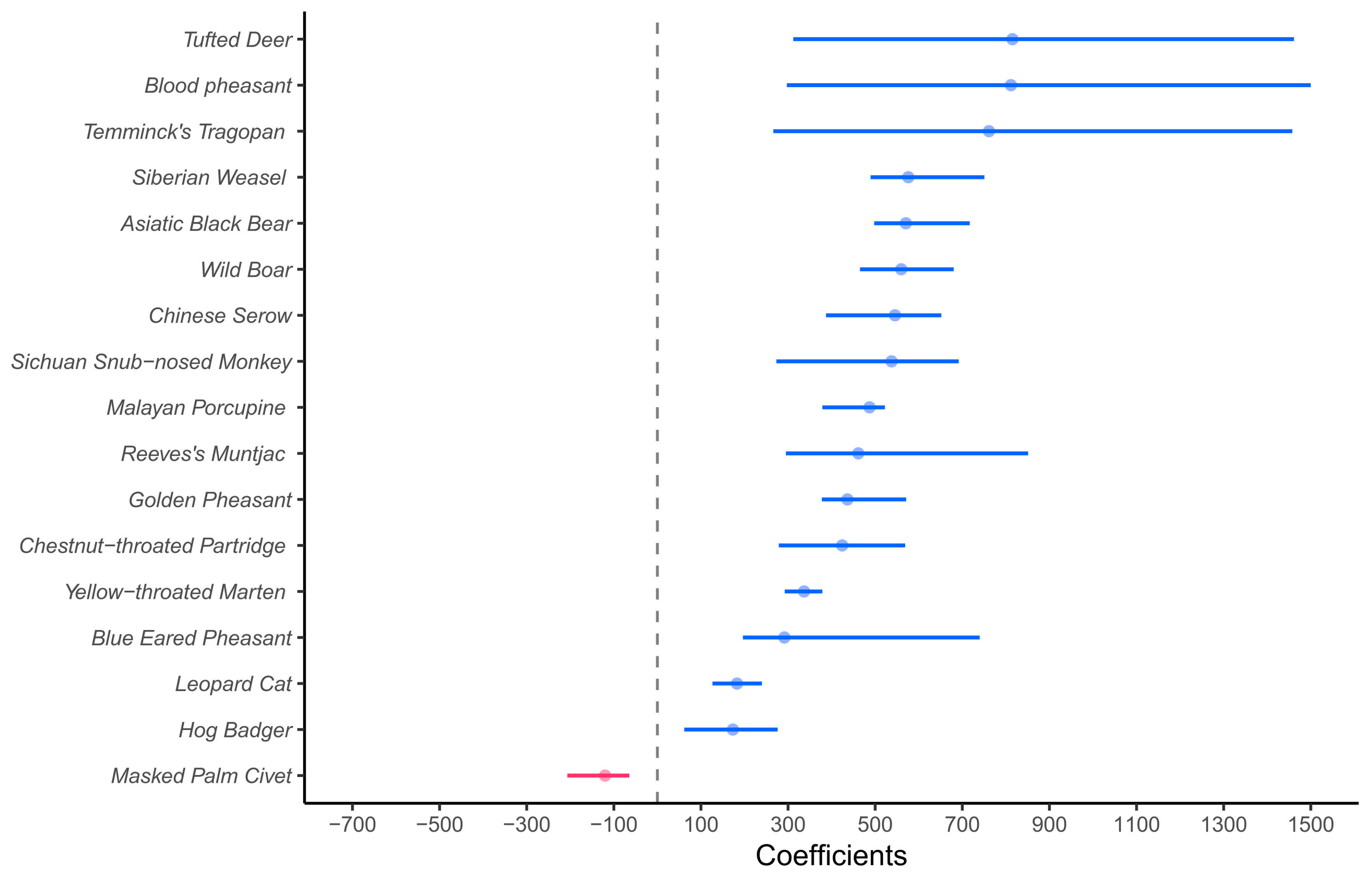
3.2. Factors Affecting Wildlife Visiting
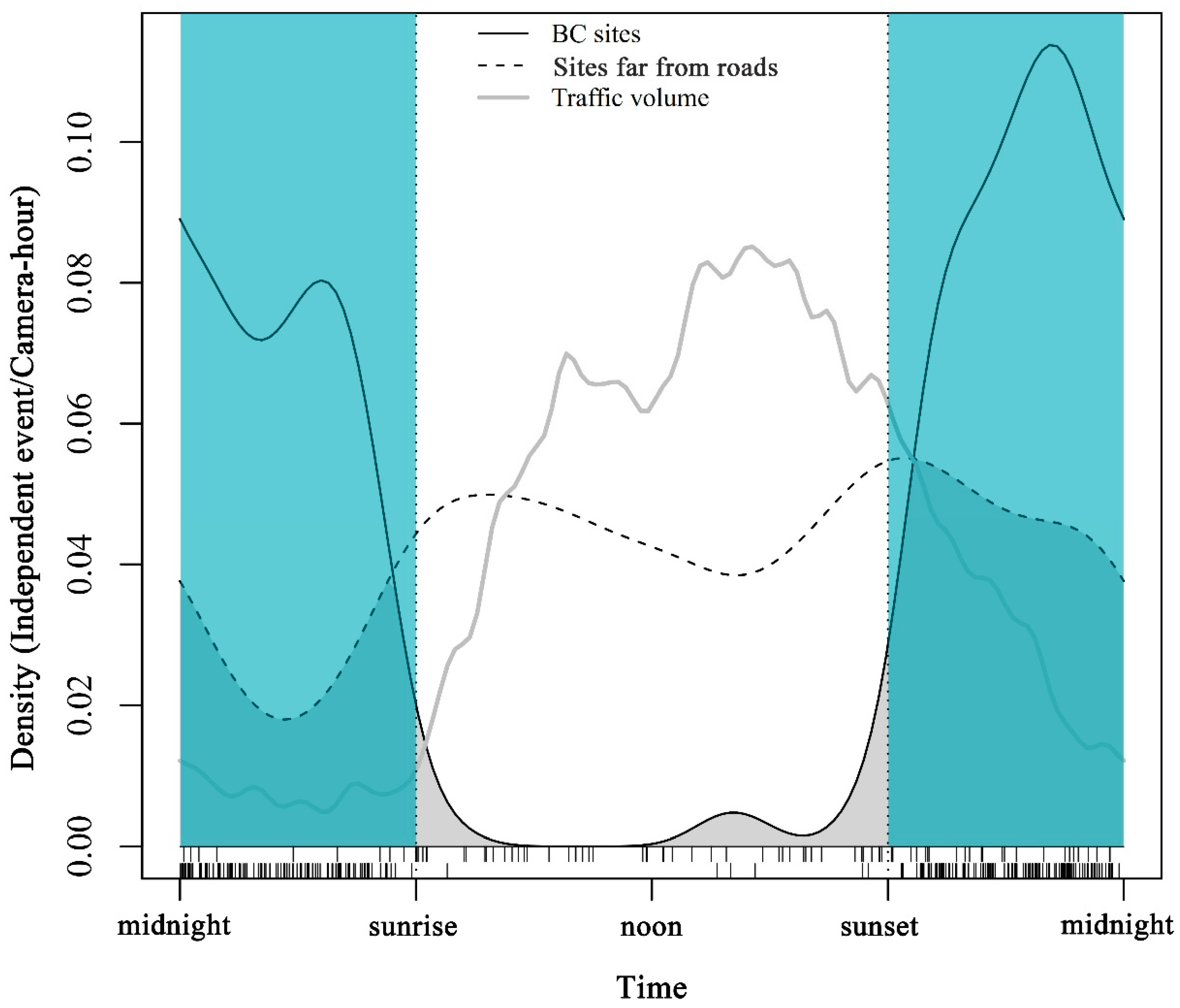
3.3. Daily Pattern of Visiting
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trombulak, S.C.; Frissell, C.A. Review of Ecological Effects of Roads on Terrestrial and Aquatic Communities. Conserv. Biol. 2000, 14, 18–30. [Google Scholar] [CrossRef] [Green Version]
- Forman, R.T.T. Estimate of the Area Affected Ecologically by the Road System in the United States. Conserv. Biol. 2000, 14, 31–35. [Google Scholar] [CrossRef]
- Forman, R.T.; Alexander, L.E.J. Roads and their major ecological effects. Annu. Rev. Ecol. Syst. 1998, 29, 207–231. [Google Scholar] [CrossRef] [Green Version]
- Laurance, W. If you can’t build well, then build nothing at all. Nature 2018, 563, 295. [Google Scholar] [CrossRef] [Green Version]
- Russo, L.F.; Barrientos, R.; Fabrizio, M.; Di Febbraro, M.; Loy, A. Prioritizing road-kill mitigation areas: A spatially explicit national-scale model for an elusive carnivore. Divers. Distrib. 2020, 26, 1093–1103. [Google Scholar] [CrossRef]
- Clevenger, A.P.; Waltho, N. Factors Influencing the Effectiveness of Wildlife Underpasses in Banff National Park, Alberta, Canada. Conserv. Biol. 2000, 14, 47–56. [Google Scholar] [CrossRef]
- Coffin, A.W. From roadkill to road ecology: A review of the ecological effects of roads. J. Transp. Geogr. 2007, 15, 396–406. [Google Scholar] [CrossRef]
- Wilcox, B.A.; Murphy, D.D. Conservation Strategy: The Effects of Fragmentation on Extinction. Am. Nat. 1985, 125, 879–887. [Google Scholar] [CrossRef]
- Madadi, H.; Moradi, H.; Soffianian, A.; Salmanmahiny, A.; Senn, J.; Geneletti, D. Degradation of natural habitats by roads: Comparing land-take and noise effect zone. Environ. Impact Assess. Rev. 2017, 65, 147–155. [Google Scholar] [CrossRef]
- Geneletti, D.; Dawa, D. Environmental impact assessment of mountain tourism in developing regions: A study in Ladakh, Indian Himalaya. Environ. Impact Assess. Rev. 2009, 29, 229–242. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, e15000522015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceia-Hasse, A.; Navarro, L.M.; Borda-de-Água, L.; Pereira, H.M. Population persistence in landscapes fragmented by roads: Disentangling isolation, mortality, and the effect of dispersal. Ecol. Model. 2018, 375, 45–53. [Google Scholar] [CrossRef]
- Braschler, B.; Dolt, C.; Baur, B. The Function of A Set-Aside Railway Bridge in Connecting Urban Habitats for Animals: A Case Study. Sustainability 2020, 12, 1194. [Google Scholar] [CrossRef] [Green Version]
- Ceia-Hasse, A.; Borda-de-Água, L.; Grilo, C.; Pereira, H.M. Global exposure of carnivores to roads. Glob. Ecol. Biogeogr. 2017, 26, 592–600. [Google Scholar] [CrossRef] [Green Version]
- Rytwinski, T.; Fahrig, L. Do species life history traits explain population responses to roads? A meta-analysis. Biol. Conserv. 2012, 147, 87–98. [Google Scholar] [CrossRef]
- Corlatti, L.; Hackländer, K.; Frey-Roos, F. Ability of Wildlife Overpasses to Provide Connectivity and Prevent Genetic Isolation. Conserv. Biol. 2009, 23, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Glista, D.J.; DeVault, T.L.; DeWoody, J.A. A review of mitigation measures for reducing wildlife mortality on roadways. Landsc. Urban Plan. 2009, 91, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sawaya, M.A.; Clevenger, A.P.; Kalinowski, S.T. Demographic Connectivity for Ursid Populations at Wildlife Crossing Structures in Banff National Park. Conserv. Biol. 2013, 27, 721–730. [Google Scholar] [CrossRef]
- van der Grift, E.A.; van der Ree, R.; Fahrig, L.; Findlay, S.; Houlahan, J.; Jaeger, J.A.G.; Klar, N.; Madriñan, L.F.; Olson, L. Evaluating the effectiveness of road mitigation measures. Biodivers. Conserv. 2013, 22, 425–448. [Google Scholar] [CrossRef] [Green Version]
- Seidler, R.G.; Green, D.S.; Beckmann, J.P. Highways, crossing structures and risk: Behaviors of Greater Yellowstone pronghorn elucidate efficacy of road mitigation. Glob. Ecol. Conserv. 2018, 15, e00416. [Google Scholar] [CrossRef]
- Ascensão, F.; Mira, A. Factors affecting culvert use by vertebrates along two stretches of road in southern Portugal. Ecol. Res. 2007, 22, 57–66. [Google Scholar] [CrossRef]
- Clevenger, A.P.; Chruszcz, B.; Gunson, K. Drainage culverts as habitat linkages and factors affecting passage by mammals. J. Appl. Ecol. 2001, 38, 1340–1349. [Google Scholar] [CrossRef]
- Brunen, B.; Daguet, C.; Jaeger, J.A.G. What attributes are relevant for drainage culverts to serve as efficient road crossing structures for mammals? J. Environ. Manag. 2020, 268, 110423. [Google Scholar] [CrossRef]
- Ng, S.J.; Dole, J.W.; Sauvajot, R.M.; Riley, S.P.D.; Valone, T.J. Use of highway undercrossings by wildlife in southern California. Biol. Conserv. 2004, 115, 499–507. [Google Scholar] [CrossRef]
- Grilo, C.; Bissonette, J.A.; Santos-Reis, M. Response of carnivores to existing highway culverts and underpasses: Implications for road planning and mitigation. Biodivers. Conserv. 2008, 17, 1685–1699. [Google Scholar] [CrossRef]
- Villalva, P.; Reto, D.; Santos-Reis, M.; Revilla, E.; Grilo, C. Do dry ledges reduce the barrier effect of roads? Ecol. Eng. 2013, 57, 143–148. [Google Scholar] [CrossRef]
- Wang, Y.; Guan, L.; Chen, J.; Kong, Y. Influences on mammals frequency of use of small bridges and culverts along the Qinghai–Tibet railway, China. Ecol. Res. 2018, 33, 879–887. [Google Scholar] [CrossRef]
- Bhardwaj, M.; Olsson, M.; Seiler, A. Ungulate use of non-wildlife underpasses. J. Environ. Manag. 2020, 273, 111095. [Google Scholar] [CrossRef]
- Clevenger, A.; Barrueto, M. Trans-Canada Highway Wildlife and Monitoring Research, Final Report. Part B: Research; Western Transportation Institute: Bozeman, MT, USA, 2014. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J.J.N. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Xu, W.; Viña, A.; Kong, L.; Pimm, S.L.; Zhang, J.; Yang, W.; Xiao, Y.; Zhang, L.; Chen, X.; Liu, J.; et al. Reassessing the conservation status of the giant panda using remote sensing. Nat. Ecol. Evol. 2017, 1, 1635–1638. [Google Scholar] [CrossRef]
- He, K.; Dai, Q.; Gu, X.; Zhang, Z.; Zhou, J.; Qi, D.; Gu, X.; Yang, X.; Zhang, W.; Yang, B.; et al. Effects of roads on giant panda distribution: A mountain range scale evaluation. Sci. Rep. 2019, 9, 1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.; Swaisgood, R.R.; Dai, Q.; Yang, Z.; Yuan, S.; Owen, M.A.; Pilfold, N.W.; Yang, X.; Gu, X.; Zhou, H.; et al. Giant panda distributional and habitat-use shifts in a changing landscape. Conserv. Lett. 2018, 11, e12575. [Google Scholar] [CrossRef]
- Qiu, L.; Han, H.; Zhou, H.; Hong, M.; Zhang, Z.; Yang, X.; Gu, X.; Zhang, W.; Wei, W.; Dai, Q. Disturbance control can effectively restore the habitat of the giant panda (Ailuropoda melanoleuca). Biol. Conserv. 2019, 238, 108233. [Google Scholar] [CrossRef]
- Administration, S.F. The Fourth National Giant Panda Survey. State Forestry Administration of China Press: Beijing, China, 2015. [Google Scholar]
- Tanwar, K.S.; Sadhu, A.; Jhala, Y.V. Camera trap placement for evaluating species richness, abundance, and activity. Sci. Rep. 2021, 11, 23050. [Google Scholar] [CrossRef]
- Burton, A.C.; Neilson, E.; Moreira, D.; Ladle, A.; Steenweg, R.; Fisher, J.T.; Bayne, E.; Boutin, S. REVIEW: Wildlife camera trapping: A review and recommendations for linking surveys to ecological processes. J. Appl. Ecol. 2015, 52, 675–685. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote. Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Huang, H.; Chen, Y.; Clinton, N.; Wang, J.; Wang, X.; Liu, C.; Gong, P.; Yang, J.; Bai, Y.; Zheng, Y.; et al. Mapping major land cover dynamics in Beijing using all Landsat images in Google Earth Engine. Remote Sens. Environ. 2017, 202, 166–176. [Google Scholar] [CrossRef]
- Lovell, C.; Li, S.; Turner, J.; Carbone, C. The effect of habitat and human disturbance on the spatiotemporal activity of two urban carnivores: The results of an intensive camera trap study. Ecol. Evol. 2022, 12, e8746. [Google Scholar] [CrossRef]
- Greenville, A.C.; Wardle, G.M.; Dickman, C.R. Desert mammal populations are limited by introduced predators rather than future climate change. R. Soc. Open Sci. 2017, 4, 170384. [Google Scholar] [CrossRef] [Green Version]
- Etana, B.; Atickem, A.; Tsegaye, D.; Bekele, A.; De Beenhouwer, M.; Hundera, K.; Lens, L.; Fashing, P.J.; Stenseth, N.C. Traditional shade coffee forest systems act as refuges for medium- and large-sized mammals as natural forest dwindles in Ethiopia. Biol. Conserv. 2021, 260, 109219. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R.; Springer: New York, NY, USA, 2009; Volume 574. [Google Scholar]
- Reed, D.; Ward, A. Efficacity of methods advocated to reduce deer vehicle accidents: Research and rationale in the USA. In Proceedings of the Highway and Wildlife Relationships, Symposium, Strasbourg, France, 5–7 June 1985. [Google Scholar]
- Gehrt, S.D.; Prange, S.J.B.E. Interference competition between coyotes and raccoons: A test of the mesopredator release hypothesis. Behav. Ecol. 2007, 18, 204–214. [Google Scholar] [CrossRef] [Green Version]
- Ripple, W.J.; Estes, J.A.; Beschta, R.L.; Wilmers, C.C.; Ritchie, E.G.; Hebblewhite, M.; Berger, J.; Elmhagen, B.; Letnic, M.; Nelson, M.P. Status and ecological effects of the world’s largest carnivores. Science 2014, 343, 1241484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansfield, E.R.; Helms, B.P. Detecting Multicollinearity. Am. Stat. 1982, 36, 158–160. [Google Scholar] [CrossRef]
- Barrow, L.N.; McNew, S.M.; Mitchell, N.; Galen, S.C.; Lutz, H.L.; Skeen, H.; Valqui, T.; Weckstein, J.D.; Witt, C.C. Deeply conserved susceptibility in a multi-host, multi-parasite system. Ecol. Lett. 2019, 22, 987–998. [Google Scholar] [CrossRef] [PubMed]
- Golivets, M.; Woodall, C.W.; Wallin, K.F. Functional form and interactions of the drivers of understory non-native plant invasions in northern US forests. J. Appl. Ecol. 2019, 56, 2596–2608. [Google Scholar] [CrossRef]
- Bürkner, P.-C. brms: An R Package for Bayesian Multilevel Models Using Stan. J. Stat. Softw. 2017, 80, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Meredith, M.; Ridout, M. Estimating overlap of daily activity patterns from camera trap data. J. Agric. Biol. Environ. Stat. 2018, 14, 322–337. [Google Scholar]
- Ridout, M.S.; Linkie, M. Estimating overlap of daily activity patterns from camera trap data. JABES 2009, 14, 322–337. [Google Scholar] [CrossRef]
- Dodd, C.K.; Barichivich, W.J.; Smith, L.L. Effectiveness of a barrier wall and culverts in reducing wildlife mortality on a heavily traveled highway in Florida. Biol. Conserv. 2004, 118, 619–631. [Google Scholar] [CrossRef]
- Sołowczuk, A.J.S. Effect of Landscape Elements and Structures on the Acoustic Environment on Wildlife Overpasses Located in Rural Areas. Sustainability 2020, 12, 7866. [Google Scholar] [CrossRef]
- Barrueto, M.; Ford, A.T.; Clevenger, A.P. Anthropogenic effects on activity patterns of wildlife at crossing structures. Ecosphere 2014, 5, art27. [Google Scholar] [CrossRef]
- Gagnon, J.W.; Dodd, N.L.; Ogren, K.S.; Schweinsburg, R.E. Factors associated with use of wildlife underpasses and importance of long-term monitoring. J. Wildl. Manag. 2011, 75, 1477–1487. [Google Scholar] [CrossRef]
- Barber, J.R.; Crooks, K.R.; Fristrup, K.M. The costs of chronic noise exposure for terrestrial organisms. Trends Ecol. Evol. 2010, 25, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Hammitt, W.E.; Cole, D.N.; Monz, C.A. Wildland Recreation: Ecology and Management; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Erb, P.L.; McShea, W.J.; Guralnick, R.P. Anthropogenic influences on macro-level mammal occupancy in the Appalachian trail corridor. PLoS ONE 2012, 7, e42574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queiroz, C.; Beilin, R.; Folke, C.; Lindborg, R. Farmland abandonment: Threat or opportunity for biodiversity conservation? A global review. Front. Ecol. Environ. 2014, 12, 288–296. [Google Scholar] [CrossRef]
- Gordon, I.J. Review: Livestock production increasingly influences wildlife across the globe. Animal 2018, 12, s372–s382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schieltz, J.M.; Rubenstein, D.I. Evidence based review: Positive versus negative effects of livestock grazing on wildlife. What do we really know? Environ. Res. Lett. 2016, 11, 113003. [Google Scholar] [CrossRef] [Green Version]
- Clevenger, A.P.; Waltho, N. Dry drainage culvert use and design considerations for small-and medium-sized mammal movement across a major transportation corridor. In Proceedings of the Third International Conference on Wildlife Ecology and Transportation Florida Department of Transportation US Department of Transportation US Forest Service Defenders of Wildlife, Missoula, MT, USA, 13–16 September 1999. [Google Scholar]
- McDonald, W.; Clair, C.C.S. Elements that promote highway crossing structure use by small mammals in Banff National Park. J. Appl. Ecol. 2004, 41, 82–93. [Google Scholar] [CrossRef]
- Roads, roadsides and wildlife conservation: A review: A.F. Bennett, in: Nature conservation 2: The role of corridors, ed D.A. Saunders & R.J. Hobbs, (Surrey Beatty). Biol. Conserv. 1991, 60, 64. [CrossRef]
- Gunson, K.E.; Mountrakis, G.; Quackenbush, L.J. Spatial wildlife-vehicle collision models: A review of current work and its application to transportation mitigation projects. J. Environ. Manag. 2011, 92, 1074–1082. [Google Scholar] [CrossRef]
- Ramp, D.; Wilson, V.K.; Croft, D.B. Assessing the impacts of roads in peri-urban reserves: Road-based fatalities and road usage by wildlife in the Royal National Park, New South Wales, Australia. Biol. Conserv. 2006, 129, 348–359. [Google Scholar] [CrossRef]
- Galinskaitė, L.; Ulevičius, A.; Valskys, V.; Samas, A.; Busher, P.E.; Ignatavičius, G. The Influence of Landscape Structure on Wildlife–Vehicle Collisions: Geostatistical Analysis on Hot Spot and Habitat Proximity Relations. ISPRS Int. J. Geo Inf. 2022, 11, 63. [Google Scholar]
- Gu, H.; Dai, Q.; Wang, Q.; Wang, Y. Factors contributing to amphibian road mortality in a wetland. Curr. Zool. 2011, 57, 768–774. [Google Scholar] [CrossRef]
- Li, T.; Shilling, F.; Thorne, J.; Li, F.; Schott, H.; Boynton, R.; Berry, A.M. Fragmentation of China’s landscape by roads and urban areas. Landsc. Ecol. 2010, 25, 839–853. [Google Scholar] [CrossRef] [Green Version]
- Forman, R.T.; Sperling, D.; Bissonette, J.A.; Clevenger, A.P.; Cutshall, C.D.; Dale, V.H.; Fahrig, L.; France, R.L.; Goldman, C.R.; Heanue, K. Road Ecology: Science and Solutions; Island Press: Washington, DC, USA, 2003. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Model | Variable | Coefficient | SE | Lower95% | Upper95% | R2 |
|---|---|---|---|---|---|---|
| Richness~site | Site: Far | 280.65 | 55.08 | 171.74 | 390.29 | 0.50 |
| Model | LOOIC | WAIC | Δelpd (SE) | R2 | Variables | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Human | Traffic: Low | DA | Openness | Cover50 | Cover1 km | Zi-Cover1 km | |||||
| Global | 251.70 | 248.60 | −0.30 (0.70) | 0.49 | −0.17 (−0.30, −0.04) | 0.45 (0.15, 0.75) | 0.22 (0.05, 0.39) | −0.56 (−4.29, 3.41) | 0.79 (−19.16, 21.22) | −19.60 (−92.98, 39.78) | 19.12 (0.50, 37.69) |
| Reduced | 251.10 | 248.00 | 0 | 0.49 | −0.17 (−0.31, −0.04) | 0.46 (0.17, 0.76) | 0.21 (0.05, 0.37) | - | - | - | 20.93 (3.89, 38.05) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, S.; Yue, Y.; Wang, Y.; Zhang, D.; Yang, B.; Yu, Z.; Lin, H.; Dai, Q. The Factors Influencing Wildlife to Use Existing Bridges and Culverts in Giant Panda National Park. Diversity 2023, 15, 487. https://doi.org/10.3390/d15040487
Lu S, Yue Y, Wang Y, Zhang D, Yang B, Yu Z, Lin H, Dai Q. The Factors Influencing Wildlife to Use Existing Bridges and Culverts in Giant Panda National Park. Diversity. 2023; 15(4):487. https://doi.org/10.3390/d15040487
Chicago/Turabian StyleLu, Song, Ying Yue, Yihong Wang, Dawei Zhang, Biao Yang, Zhen Yu, Honghui Lin, and Qiang Dai. 2023. "The Factors Influencing Wildlife to Use Existing Bridges and Culverts in Giant Panda National Park" Diversity 15, no. 4: 487. https://doi.org/10.3390/d15040487