1. Introduction
Globally, urban land expansion causes the loss of natural habitat, the fragmentation of remaining natural areas, and the alteration of natural areas’ abiotic and biotic characteristics [
1,
2,
3]. This has had and will continue to have a significant influence on biodiversity, both directly and indirectly [
4]. On the one hand, biodiversity is an important part of any healthy ecosystem and is thus essential to the provision of ecosystem services [
5]. On the other hand, urban biodiversity is crucial for human well-being because high biodiversity enables residents to enjoy nature in cities.
Several recent studies have focused on the analysis of the relationship between bird diversity and urban landscape patterns at different scales [
6,
7,
8]. On the landscape scale, patch area, connectivity, and level of urbanization in the surrounding matrix were found to be the most important variables in explaining bird species diversity in urban areas [
9,
10,
11]. Meanwhile, open features, especially open water, could be an important factor affecting bird diversity in forests in urban areas [
12]. Bird distribution patterns in urban areas were strongly associated with habitat evenness, the largest patch index for woodland at the 400 m scale, and the contagion index at the 400 and 1000 m scales [
13].
At the patch scale, attention was paid to management intensity and habitat characteristics, like the vertical heterogeneity of forest patches [
10,
14,
15]. A study of bird communities and habitat vegetation composition in Shanghai’s urban park found that vegetation complexity and structure positively affected bird community composition [
14]. Bird species richness was related not only to the amount of vegetation structure but also to whether the green space was cultivated or natural, and it was discovered that cultivated greenery leads to homogeneous bird communities in the urban landscape [
15].
The appropriate way to quantify and enhance the function of urban green space as a wildlife habitat has been frequently mentioned. Many studies suggest that species diversity estimates based on landscape indicators should be multi-scaled [
16,
17,
18]; however, there are few case studies, particularly on birds [
19,
20,
21]. One of the reasons is that the statistical approaches required to manage these challenges are still being researched [
17].
In recent years, several studies have focused on bird diversity in specific urban green spaces, such as streets, small parks, and backyards. A study analyzing the taxonomic, functional, and phylogenetic diversity of birds influenced by street design suggested minimizing paved road areas to boost bird diversity in streets [
22]. The abundance of bird species in small urban parks may be predicted most accurately by two factors: the area of the park and the proportion of canopy cover [
7]. A study explored the contribution of residential backyards to urban bird diversity, finding that backyards with the largest areas, tallest trees and bushes, and proximity to other green places increase species diversity [
23].
Studies have revealed that the urbanization process impacted the decrease of bird functional diversity through biotic homogenization [
24,
25]. Functional diversity (FD) is a measure of the diversity of species traits found in communities that provides a link between species’ traits and how they influence ecosystem functions [
26]. Taxonomic and functional diversity may respond differently to anthropogenic effects and therefore necessitate different conservation strategies.
Urban rivers are essential for preserving urban biodiversity and promoting the health of urban ecosystems [
27,
28]. Urban rivers have relatively complete habitat structures and are ecological corridors that support a high level of biodiversity and connect fragmented urban green space, reducing isolation, and thus maintaining urban biodiversity [
29,
30]. However, human activities change the form and function of urban river landscapes, such as buried, cut, and hardened rivers, and increase the surface coverage of roads and buildings [
31,
32]. This reduces the naturalness, variety, sustainability, and landscape quality of rivers, ultimately decreasing biodiversity [
33]. Due to these challenges, researchers are becoming increasingly interested in urban river corridors and are paying attention to the relationship between river network connectivity and bird community structure [
27]; the landscape-scale characteristics of suburban rivers impacting bird diversity [
29]; and the impact of urban structure, combination of land use, and infrastructure on avian riparian habitat [
34]. However, few studies have analyzed the impact of the plan, design, and management of multi-scale urban riverside green space on bird communities by assessing several facets of diversity, such as taxonomic and functional.
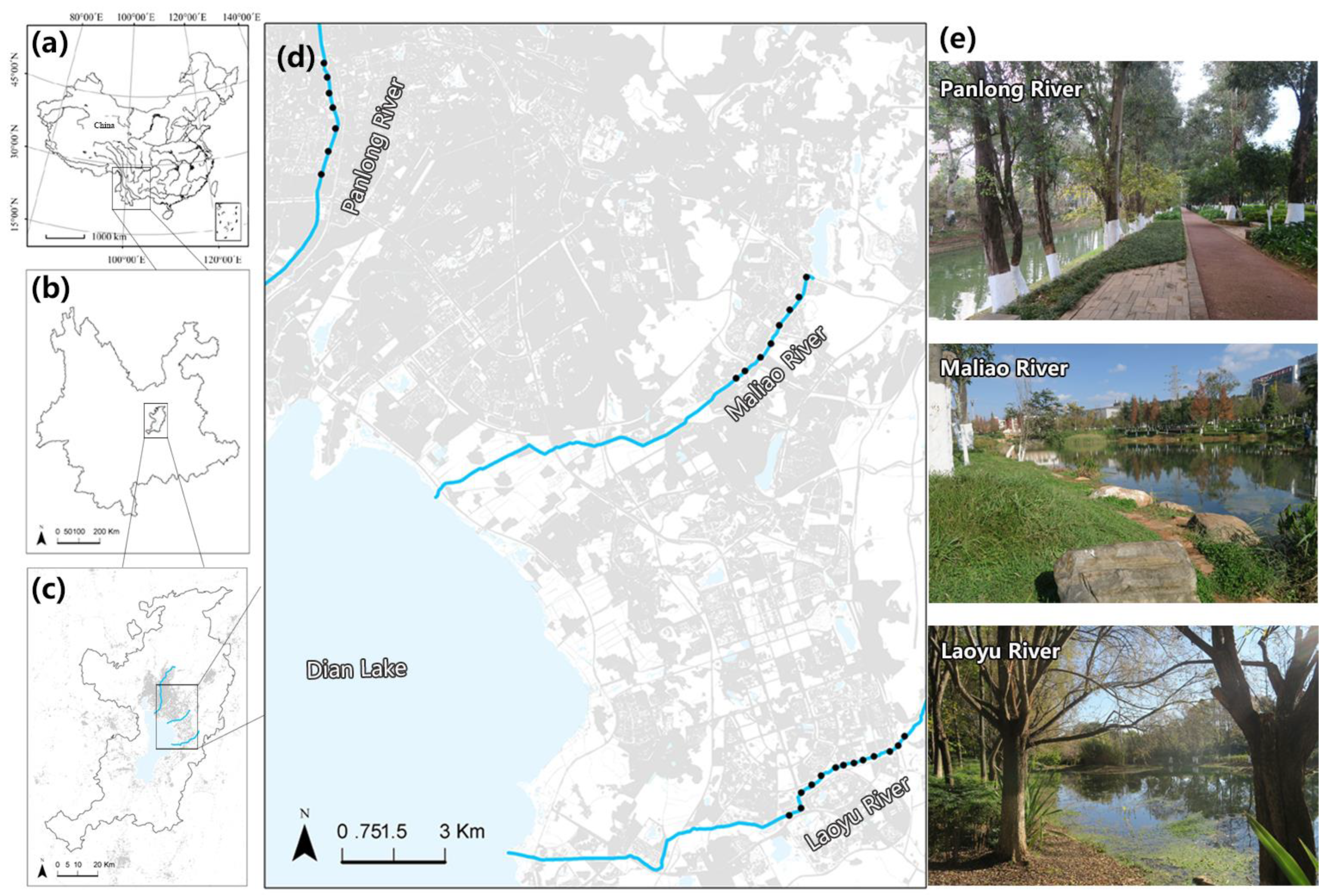
In this study, we explored bird community variation (taxonomic and functional) among the three rivers with different levels of human disturbance along the urban development gradient. This study related bird species richness and FD to environmental variables at three spatial scales: landscape metrics at two different landscape scales (500 and 100 m radius) and vegetation structure at patch scales (20 m radius).
3. Result
3.1. Species Diversity and Functional Diversity and Their Variation among Rivers
We recorded 71 species (3387 counts) from 34 families and 12 orders in 12 replicated surveys covering 28 sampling sites during the breeding and non-breeding seasons (see
Appendix A). The bird species richness ranged from 11 to 32 (mean ± s.e. = 19.11 ± 1.05) for the different sites. The Panlong River (a high-human disturbance river) contained 38 species; the Maliao River (a medium-human disturbance river) contained 46 species; and the Laoyu River (a low-human disturbance river) contained 57 species. We recorded a total of 45 bird species in the breeding season and 63 species in the non-breeding season across all sampling sites in the three rivers (
Figure 2). The range of species observed at each sampling site in the breeding season was between 7 and 21 (mean ± s.e. = 12.32 ± 0.67), while in the non-breeding season, the range was between 5 and 27 (mean ± s.e. = 14.00 ± 0.99). Meanwhile, Moran’s S from the spatial correlation analysis was not significant (
p = 0.118), indicating that species richness was spatially independent.
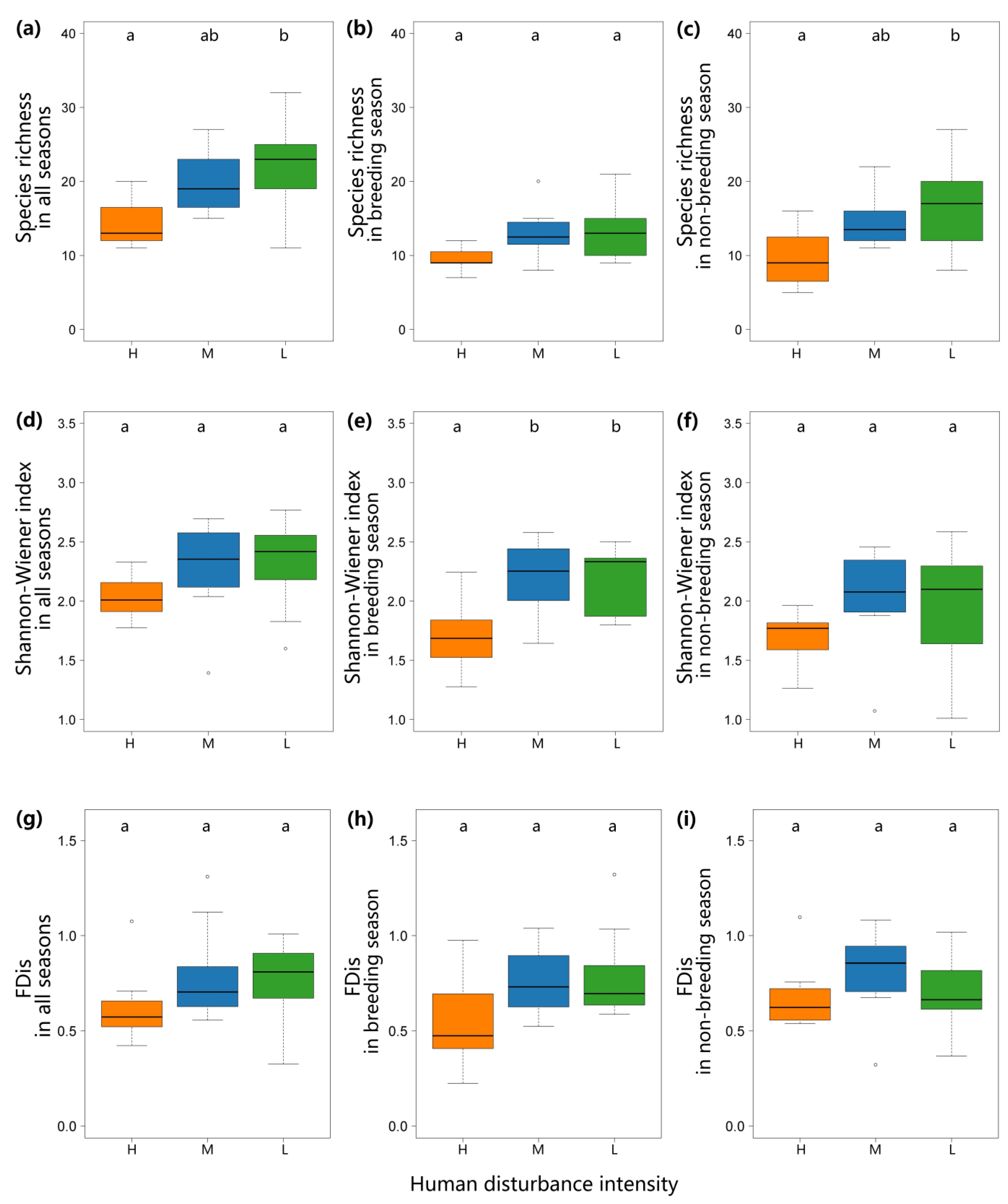
During both seasons, the bird species richness along the low-human disturbance river was highest, followed by the medium-human disturbance river and the high-human disturbance river. The degree of human disturbance had a significant effect on bird richness according to the analysis of the one-way ANOVA (
Figure 2;
F2,25 = 4.239,
p = 0.026). Tukey’s HSD post hoc test revealed that the low-human disturbance river had a significantly higher bird richness than the high-human disturbance river (
p = 0.022;
Figure 2). The richness of the three types of rivers during the breeding season has no significant difference. In the non-breeding season, Tukey’s HSD post hoc test revealed that the low-human disturbance river had a significantly higher bird richness than the high-human disturbance river (
p = 0.030;
Figure 2).
Throughout the year and the non-breeding season of birds, the Shannon–Wiener index showed no significant difference. However, in the breeding season, low- and medium-human disturbance rivers had a significantly higher Shannon–Wiener index than the high-human disturbance river (p = 0.006, p = 0.009). There was no significant difference in FDis among the three types of rivers during the annual cycle, the breeding or non-breeding season.
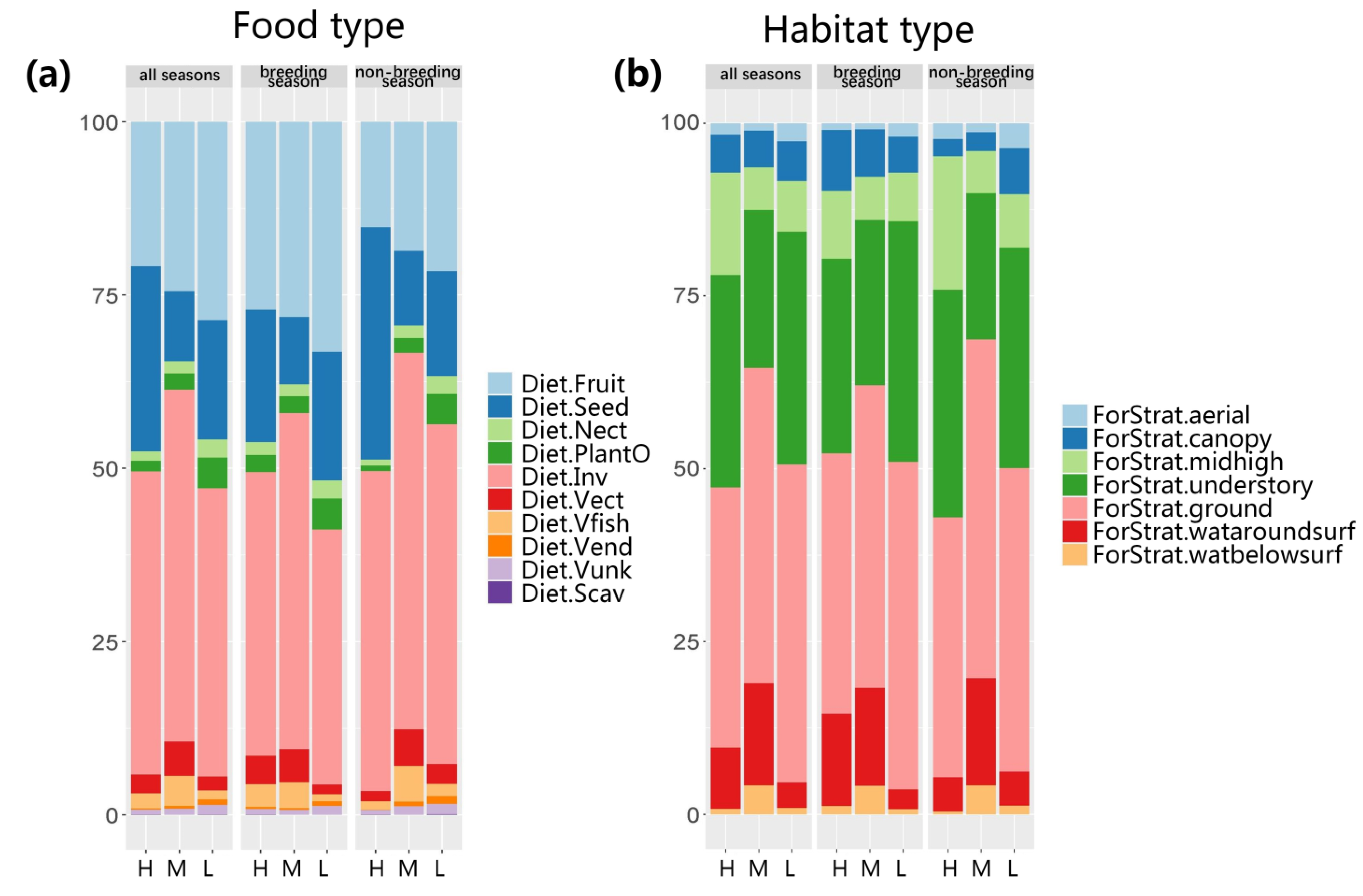
In the three rivers, the food source of bird species was dominated by invertebrates, followed by fruits, seeds, amphibians, fish, other plants, nectar, birds, and corrosion. With the increase in human disturbance, the relative abundance of fruit-eating and nectar-eating birds decreased while that of seed-eating birds increased (
Figure 3). Considering the vertical foraging strata, with the increase in human disturbance, the relative abundance of time spent on the ground stratum by bird species increased (
Figure 3; low-human disturbance rivers = 46.01%, medium-human disturbance rivers = 45.62%, high-human disturbance rivers = 37.62%).
3.2. PLSR Linked Species Diversity and Functional Diversity with Landscape- and Patch-Level Variables
First, we used richness and FDis as the response variables and landscape- and patch-level variables as the explanatory variables to perform PLSR, and the first components explained 38.8% and 16.2% of the variation in the response variable for richness and FDis, respectively. There were 19 and 18 metrics with VIP ≥ 1 (i.e., the important explanatory variable; see
Table A3 and
Table A4).
For bird richness, in 19 important explanatory variables, there were 8 landscape indicators on the 100 m scale, 6 landscape indicators on the 500 m scale, respectively, and 5 patch-level variables on the 20 m scale (that is, 42.1%, 31.6%, and 26.3% of the 19 important explanatory variables). For open green space and aquatic plants, the effect in the 100 m radius was more significant than that in the 500 m radius (VIP ≥ 1). For constructed land, the three landscape indices on both the 500 and 100 m scales were important. The AREA_MN of the forest was important on both the 500 and 100 m scales. In the 20 m radius, litter layer and ground herbaceous plant coverage, as well as a vertical range of 3–10 m vegetation cover, were important explanatory variables.
For bird FDis, in the 18 important explanatory variables, there were 8 landscape indicators on the 100 m scale, 7 landscape indicators on the 500 m scale, and 3 landscape indicators on the 20 m scale (that is, 44.4%, 38.9%, and 16.7% of the 18 important explanatory variables, respectively). Similar to bird species richness, the AREA_MN of the forest was also significant for bird FDis at both the 500- and 1000-m scales, and metrics at the 100-m scale were more significant for open green space. For constructed land, the ED and PLAND in both the 500 m and 100 m scales are important, and the metrics in the 500 m scale are more important than those in the 100 m scale for aquatic plants. On the 20 m scale, the litter layer, foliage cover in 0.5–1 m, and foliage cover in 4–6 m are important.
3.3. Multiple Regression Linked Species Diversity and Functional Diversity with Landscape- and Patch-Level Variables
For richness as a response variable, PLSR and VIP approaches were used to derive 19 explanatory variables of interest (i.e., VIP ≥ 1). To avoid multicollinearity, we selected indicators with variance inflation factors (VIFs) < 10 (n = 15) [
66] to create multiple linear regression models. We obtained 7 candidate models (i.e., top models) with independent variable counts less than or equal to 3 and ΔAICc < 2 (
Table 2). Then we applied the model averaging to the top model set, and the independent variables in the average model were mean foliage cover in 4–6 m in the 20 m scale (
p < 0.05), ED of constructed land in the 100 m scale (
p < 0.05), AREA_MN of open green space in the 100 m scale, AREA_MN of forest in the 100 m scale (
p < 0.05), ED of aquatic plant in the 100 m scale, ED of forest in the 500 m scale, and AREA_MN of constructed land in the 500 m scale (
Table 3).
For FDis as the response variable, PLSR and VIP approaches were used to derive 19 explanatory variables of interest (i.e., VIP ≥ 1). To avoid multicollinearity, we selected 15 indicators whose VIFs < 10 [
66] to create multiple linear regression models. We obtained 4 candidate models (i.e., top models) with independent variable counts less than or equal to 3 and ΔAICc < 2 (
Table 4). Then we applied the model averaging to the top model set, and independent variables in the average model are ED of forest in the 100 m scale, PLAND of forest in the 100 m scale (
p < 0.05), ED of open green spaces in the 100 m scale (
p < 0.05), mean foliage cover in 4–6 m in the 20 m scale (
p < 0.05), and ED of constructed land in the 100 m scale (
Table 5).
4. Discussion
As the scale dependency of ecological processes and multiple environmental factors at different spatial scales affects bird diversity, multi-scale approaches are better than single-scale models due to the models they produce to assess the effects of multi-scale landscape structure on bird distribution and habitat use [
18,
67,
68]. This study linked environmental variables both at small- and medium-sized landscape scales and patch scales with river bird species richness and functional diversity (FDis). We found that the landscape characteristics at 500 and 100 m scales, and patch attributes at 20 m scale had a significant effect on bird species richness, while FD was significantly affected by indexes at 100 and 20 m scales. To improve the urban riverfront landscape management and design strategy, multi-scale habitat attributes should be considered to address how to optimize land uses, which are mainly human driven at multiple scales of space, time, and intricacy of management.
There were significant differences in richness among three different intensities of human disturbance of rivers, either throughout the year or during the non-breeding season, respectively. The species richness of the low-human disturbance river (the Laoyu River) was significantly higher than that of the high-human disturbance river (the Panlong River). During the non-breeding season, some migratory birds, such as shorebirds, prefer to stay in the low-human disturbance river. Low-human disturbance rivers in urban areas are a refuge for birds [
48], especially during the non-breeding season [
69].
There was no significant difference for FDis among the three rivers, whether for the whole year or in the different seasons. As urbanization increases the functional similarity of the bird community [
3,
20], the differences in FDis among the sampling sites were not significant. Unlike bird species richness, the FDis was significantly influenced by explanatory variables on 100 and 20-m scales but not on the 500-m scale (
Table 5). Functional diversity has a complex relationship with habitat variables because it depends on the habitat and landscape structural needs of species in each functional category at the survey site [
70]. For example, frugivore and nectarivore birds can be influenced by vegetation structure and specific plant species [
70,
71].
Recent studies have explored the functional and taxonomic diversity of birds in urban green spaces [
72,
73], showing different situations. The canopy heterogeneity affected FD [
73] and the presence of large trees enhanced taxonomic diversity and functional diversity [
74]. A study conducted in Austria revealed that sealed areas (e.g., roads and buildings) had a considerable negative impact on bird species richness and functional diversity [
73]. However, another study showed that tree coverage and site size had a positive correlation with richness, while none of the measured land use characteristics were associated with FD, which means functional diversity was unrelated to measured landscape structure variables [
72]. We found that the average patch area of forest had significantly positive impacts on bird richness, the average patch area of constructed land had negative impacts on bird richness, and the area proportion of forest had significantly positive impacts on functional diversity.
Edge density (ED) is a tool to quantify how many marginal habitats are next to other types of land cover. The ED at the class level is the entire length of the patch’s edge per unit area for a certain class [
75]. The characteristic of a patch edge for a class or an entire landscape is crucial to multiple ecological processes, especially for wildlife. We found that the ED of construction land and forest had negative impacts on bird richness and functional diversity. The ED of construction land reflects the characteristics of its shape, and an increase in ED shows the increasing negative effects on those species sensitive to human disturbance. A higher ED for forestland with the same percentage of forestland area suggests more habitat exposure to human disturbance, which will be detrimental to sensitive species [
76,
77,
78].
Previous studies have demonstrated the significance of forest leaf litter for insectivorous birds because it provides greater habitat for litter-dependent invertebrates [
79,
80,
81]. Many birds like to forage among the ground’s dead leaves, such as
Turdus merula and
Turdus dissimilis. Protection of woodland insectivorous birds is essential since they are in global decline [
82,
83] and depend on specific forest structures, including large trees, dense undergrowth, and thick leaf litter [
80]. Studies indicate that the degree of park management may play a significant role in influencing the bird community structure in urban green space [
84,
85], and successful management should avoid excessive understory cleanup. In this study, although the litter layer did not appear in the average models, we obtained the VIP scores of the litter layer by PLSR for bird richness and FD greater than 1 (
Table A3 and
Table A4), which means the litter layer area was an important habitat attribute. In addition, our study focused on a general analysis of anthropogenic impact, but further research could expand on these findings by incorporating additional variables such as human population density and building type.
We suggest that urban riverside green space should be planned and managed at multiple scales, including landscape-scale management (landscape composition and configuration) and patch-scale management (vegetation structure and litter layer). Multi-scale management may be beneficial for the conservation of bird species richness and FD in urban riverside green space. It is critical to increase the total and mean area of the forest while decreasing the mean area and ED of constructed land [
13,
86,
87] at the landscape scale, as well as maintain high vegetation structure [
51,
88], vertical vegetation heterogeneity [
47], and low-intensity management that preserves the litter layer at the patch scale.



{kind=link}
{kind=link}
{kind=link}


