
Characteristics of Fragments of Woodland and Their Influence on the Distribution of Soil Fauna in Agricultural Landscape

Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of Study Sites
2.2. Sampling Methods
2.3. Environmental Variables
2.4. Data Analysis
3. Results
4. Discussion
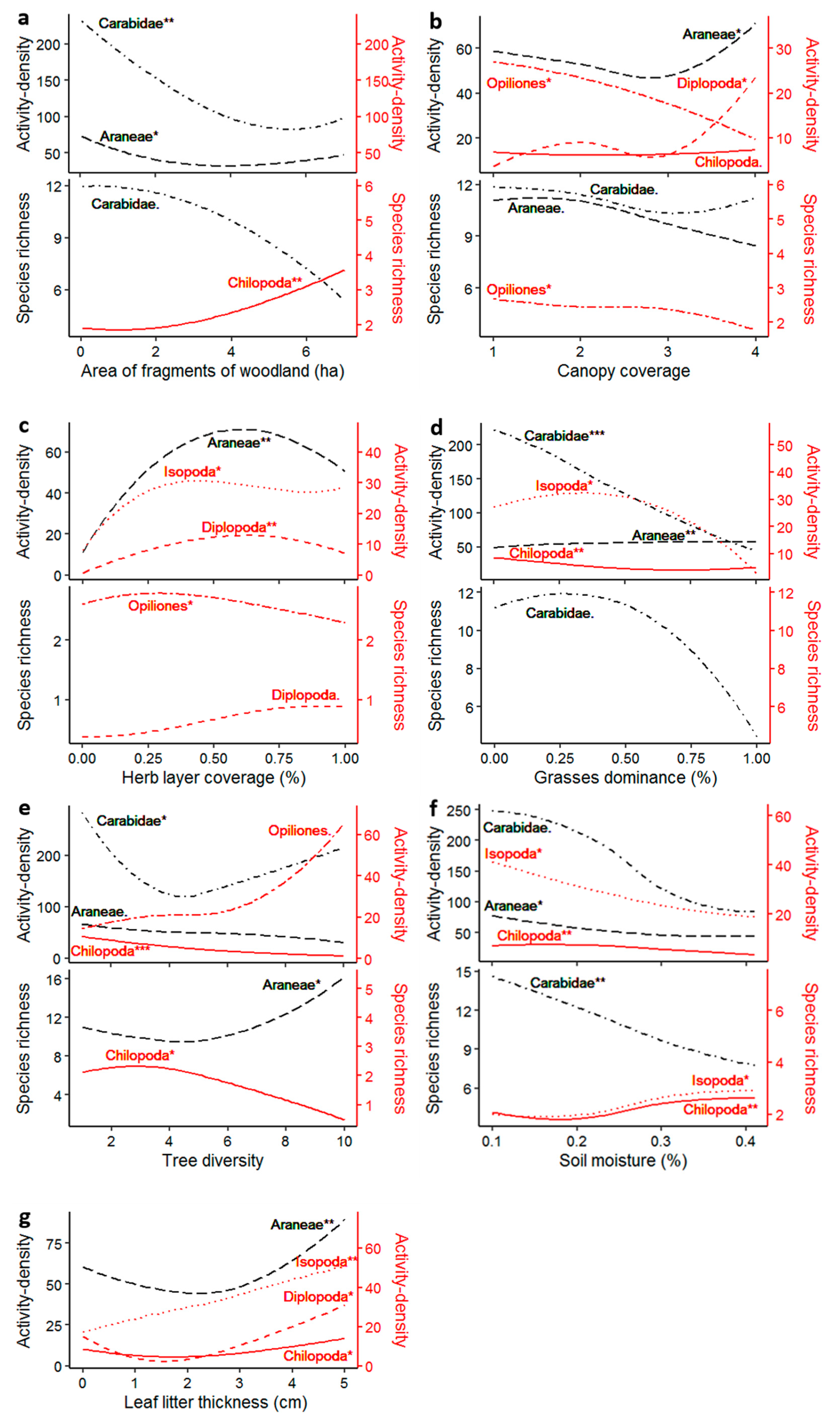
4.1. Area of Fragments of Woodland
4.2. Canopy Coverage
4.3. Herbaceous Layer Coverage and Dominance of Grasses
4.4. Tree Diversity
4.5. Soil Moisture
4.6. Leaf Litter Thickness
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duelli, P.; Obrist, M.K.; Schmatz, D.R. Biodiversity evaluation in agricultural landscapes: Above-ground insects. Invertebr. Biodivers. Bioindic. Sustain. Landsc. 1999, 74, 33–64. [Google Scholar] [CrossRef]
- Webb, N.R. Studies on the invertebrate fauna of fragmented heathland in Dorset, UK, and the implications for conservation. Biol. Conserv. 1989, 47, 153–165. [Google Scholar] [CrossRef]
- Duelli, P.; Studer, M.; Marchand, I.; Jakob, S. Population movements of arthropods between natural and cultivated areas. Biol. Conserv. 1990, 54, 193–207. [Google Scholar] [CrossRef]
- Fournier, E.; Loreau, M. Respective roles of recent hedges and forest patch remnants in the maintenance of ground-beetle (Coleopera: Carabidae) diversity in an agricultural landscape. Landsc. Ecol. 2001, 16, 17–32. [Google Scholar] [CrossRef]
- Weibull, A.C.; Östman, Ö. Species composition in agroecosystems: The effect of landscape, habitat, and farm management. Basic Appl. Ecol. 2003, 4, 349–361. [Google Scholar] [CrossRef]
- Schweiger, O.; Maelfait, J.P.; Wingerden, W.; Hendrickx, F.; Billeter, R.; Speelmans, M.; Augenstein, I.; Aukema, B.; Aviron, S.; Bailey, D.; et al. Quantifying the impact of environmental factors on arthropod communities in agricultural landscapes across organizational levels and spatial scales. J. Appl. Ecol. 2005, 42, 1129–1139. [Google Scholar] [CrossRef]
- Poschlod, P.; Braun-Reichert, R. Small natural features with large ecological roles in ancient agricultural landscapes of Central Europe–history, value, status, and conservation. Biol. Conserv. 2017, 211, 60–68. [Google Scholar] [CrossRef]
- European Commission. Green Infrastructure (GI)-Enhancing Europe’s Natural Capital. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions. Brussels, p. 11. 2013. Available online: https://ec.europa.eu/environment/nature/ecosystems/docs/green_infrastructures/1_EN_ACT_part1_v5.pdf (accessed on 14 February 2023).
- European Commission. Technical Information on Green Infrastructure (GI). Commission Staff Working Document. Brussels, p. 24. 2013. Available online: https://ec.europa.eu/environment/nature/ecosystems/docs/green_infrastructures/1_EN_autre_document_travail_service_part1_v2.pdf (accessed on 14 February 2023).
- Act No. 114/92 Coll, 1992. On Nature Conservation and Landscape Protection. Ministry of Environment of the Czech Republic. Available online: https://www.mzp.cz/www/platnalegislativa.nsf/58170589E7DC0591C125654B004E91C1/%24file/z114_1992.pdf (accessed on 14 February 2023). (In Czech).
- Burel, F. Hedgerows and Their Role in Agricultural Landscapes. Crit. Rev. Plant Sci. 1996, 15, 169–190. [Google Scholar] [CrossRef]
- Decocq, G.; Andrieu, E.; Brunet, J.; Chabrerie, O.; De Frenne, P.; De Smedt, P.; Deconchat, M.; Diekmann, M.; Ehrmann, S.; Giffard, B.; et al. Ecosystem Services from Small Forest Patches in Agricultural Landscapes. Curr. For. Rep. 2016, 2, 30–44. [Google Scholar] [CrossRef] [Green Version]
- Bentrup, G.; Hopwood, J.; Adamson, N.L.; Vaughan, M. Temperate Agroforestry Systems and Insect Pollinators: A Review. Forests 2019, 10, 981. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Gao, Z.; Li, Y.; Niu, Y.; Su, Y.; Wang, K. Erosion control of hedgerows under soils affected by disturbed soil accumulation in the slopes of loess plateau, China. Catena 2019, 181, 104079. [Google Scholar] [CrossRef]
- Montgomery, I.; Caruso, T.; Reid, N. Hedgerows as Ecosystems: Service Delivery, Management, and Restoration. Annu. Rev. Ecol. Evol. Syst. 2020, 51, 81–102. [Google Scholar] [CrossRef]
- Pollard, K.A.; Holland, J.M. Arthropods within the woody element of hedgerows and their distribution pattern. Agric. For. Entomol. 2006, 8, 203–211. [Google Scholar] [CrossRef]
- Valdés, A.; Lenoir, J.; De Frenne, P.; Andrieu, E.; Brunet, J.; Chabrerie, O.; Cousins, S.A.O.; Deconchat, M.; De Smedt, P.; Diekmann, M.; et al. High ecosystem service delivery potential of small woodlands in agricultural landscapes. J. Appl. Ecol. 2019, 57, 4–16. [Google Scholar] [CrossRef] [Green Version]
- Burel, F.; Baudry, J. Hedgerow Network Patterns and Processes in France. In Changing Landscapes: An Ecological Perspective; Zonneveld, I.S., Forman, R.T.T., Eds.; Springer: New York, NY, USA, 1990; pp. 99–120. [Google Scholar]
- Burel, F.; Baudry, J. Structural dynamic of a hedgerow network landscape in Brittany France. Landsc. Ecol. 1990, 4, 197–210. [Google Scholar] [CrossRef]
- Previati, E.; Anna Fano, E.; Leis, M. Arthropods Biodiversity in Agricultural Landscapes: Effects of Land Use and Anthropization. Ital. J. Agron. 2007, 2, 135–141. [Google Scholar] [CrossRef]
- Agger, P.; Brandt, J. Dynamics of small biotopes in Danish agricultural landscapes. Landsc. Ecol. 1988, 1, 227–240. [Google Scholar] [CrossRef] [Green Version]
- Šarapatka, B.; Štěrba, O. Optimization of agriculture in relation to the multifunctional role of the landscape. Landsc. Urban Plan. 1998, 41, 145–148. [Google Scholar] [CrossRef]
- Demková, K.; Lipský, Z. Comparison of the current state of non-forest woody vegetation in two contrasted case study areas in Central Europe. Morav. Geogr. Rep. 2017, 25, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Skokanová, H.; Netopil, P.; Havlíček, M.; Šarapatka, B. The role of traditional agricultural landscape structures in changes to green infrastructure connectivity. Agric. Ecosyst. Environ. 2020, 302, 107071. [Google Scholar] [CrossRef]
- Sklenicka, P.; Molnarova, K.; Brabec, E.; Kumble, P.; Pittnerova, B.; Pixova, K.; Salek, M. Remnants of medieval field patterns in the Czech Republic: Analysis of driving forces behind their disappearance with special attention to the role of hedgerows. Agric. Ecosyst. Environ. 2009, 129, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Stoate, C.; Boatman, N.; Borralho, R.; Carvalho, C.R.; Snoo, G.R.; Eden, P. Ecological impacts of arable intensification in Europe. J. Environ. Manag. 2001, 63, 337–365. [Google Scholar] [CrossRef] [PubMed]
- Robinson, R.A.; Sutherland, W.J. Post-war changes in arable farming and biodiversity in Great Britain. J. Appl. Ecol. 2002, 39, 157–176. [Google Scholar] [CrossRef] [Green Version]
- Diekötter, T.; Billeter, R.; Crist, T.O. Effects of landscape connectivity on the spatial distribution of insect diversity in agricultural mosaic landscapes. Basic Appl. Ecol. 2008, 9, 298–307. [Google Scholar] [CrossRef]
- Baudry, J.; Bunce, R.G.H.; Burel, F. Hedgerows: An international perspective on their origin, function and management. J. Environ. Manag. 2000, 60, 7–22. [Google Scholar] [CrossRef]
- Mazerolle, M.J.; Villard, M.A. Patch characteristics and landscape context as predictors of species presence and abundance: A review. Ecoscience 1999, 6, 117–124. [Google Scholar] [CrossRef]
- Maudsley, M.J. A review of the ecology and conservation of hedgerow invertebrates in Britain. J. Environ. Manag. 2000, 60, 65–76. [Google Scholar] [CrossRef]
- Brose, U. Bottom-up control of carabid beetle communities in early successional wetlands: Mediated by vegetation structure or plant diversity? Oecologia 2003, 135, 407–413. [Google Scholar] [CrossRef]
- Garratt, M.P.D.; Senapathi, D.; Coston, D.J.; Mortimer, S.R.; Potts, S.G. The benefits of hedgerows for pollinators and natural enemies depends on hedge quality and landscape context. Agric. Ecosyst. Environ. 2017, 247, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Sunderland, K.D.; Fraser, A.M.; Dixon, A.F.G. Field and Laboratory Studies on Money Spiders (Linyphiidae) as Predators of Cereal Aphids. J. Appl. Ecol. 1986, 23, 433–447. [Google Scholar] [CrossRef]
- Sunderland, K.; Samu, F. Effects of Agricultural Diversification on the Abundance, Distribution, and Pest Control Potential of Spiders: A Review. Entomol. Exp. Appl. 2000, 95, 1–13. [Google Scholar] [CrossRef]
- Lang, A. Intraguild interference and biocontrol effects of generalist predators in a winter wheat field. Oecologia 2003, 134, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Westerman, P.R.; Hofman, A.; Vet, L.E.M.; van der Werf, W. Relative importance of vertebrates and invertebrates in epigeaic weed seed predation in organic cereal fields. Agric. Ecosyst. Environ. 2003, 95, 417–425. [Google Scholar] [CrossRef]
- Cock, M.J.W.; Biesmajer, J.C.; Canon, R.J.C.; Gerard, P.; Gillespie, D.; Jiménez, J.; Lavelle, P.; Raina, S.K. The positive contribution of invertebrates to sustainable agriculture and food security. CAB Rev. 2012, 7, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Graham, L.; Gaulton, R.; Gerard, F.; Staley, J.T. The influence of hedgerow structural condition on wildlife habitat provision in farmed landscapes. Biol. Conserv. 2018, 220, 122–131. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton Univ. Press: Princeton, NJ, USA, 1967; p. 109. [Google Scholar]
- Bínová, L.; Culek, M.; Glos, J.; Kocián, J.; Lacina, D.; Novotný, M.; Zimová, E. Metodika Vymezování Územního Systému Ekologické Stability; Ministry of the Environment of the Czech Republic: Praha, Czech Republic, 2017; p. 186. (In Czech)
- Crawley, M.J. The R Book; John Wiley & Sons Ltd.: Chichester, UK, 2007; p. 942. [Google Scholar]
- Griffiths, G.J.K.; Winder, L.; Holland, J.M.; Thomas, C.F.G.; Williams, E. The representation and functional composition of carabid and staphylinid beetles in different field boundary types at a farm-scale. Biol. Conserv. 2007, 135, 145–152. [Google Scholar] [CrossRef]
- Knapp, M.; Řezáč, M. Even the Smallest Non-Crop Habitat Islands Could Be Beneficial: Distribution of Carabid Beetles and Spiders in Agricultural Landscape. PLoS ONE 2015, 10, e0123052. [Google Scholar] [CrossRef]
- Niemelä, J.; Haila, Y.; Halme, E.; Lahti, T.; Pajunen, T.; Punttila, P. The distribution of carabid beetles in fragments of old coniferous taiga and adjacent managed forest. Ann. Zool. Fenn. 1988, 25, 107–119. Available online: http://www.jstor.org/stable/23734516 (accessed on 14 February 2023).
- Thomas, C.F.G.; Marshall, E.J.P. Arthropod abundance and diversity in differently vegetated margins of arable fields. Agric. Ecosyst. Environ. 1999, 72, 131–144. [Google Scholar] [CrossRef]
- Ewers, R.M.; Thorpe, S.; Didham, R.K. Synergistic interactions between edge and area effects in a heavily fragmented landscape. Ecology 2007, 88, 96–106. [Google Scholar] [CrossRef]
- Lövei, G.L.; Magura, T.; Tothmeresz, B.; Kodobocz, V. The influence of matrix and edges on species richness patterns of ground beetles (Coleoptera: Carabidae) in habitat islands. Glob. Ecol. Biogeogr. 2006, 15, 283–289. [Google Scholar] [CrossRef] [Green Version]
- Bender, D.J.; Contreras, T.A.; Fahrig, L. Habitat Loss and Population Decline: A Meta-Analysis of the Patch Size Effect. Ecology 1998, 79, 517–533. [Google Scholar] [CrossRef]
- Holland, J.; Fahrig, L. Effect of woody borders on insect density and diversity in crop fields: A landscape-scale analysis. Agric. Ecosyst. Environ. 2000, 78, 115–122. [Google Scholar] [CrossRef]
- Holland, J.D.; Begbie, M.; Birkett, T.; Reynolds, C.J.M.; Thomas, C.F.G. The influence of hedgerows on coleopteran distributions: Results from a multi-field sampling study. In Hedgerows of the World: Their Ecological Functions in Different Landscapes; Barr, C., Petit, S., Eds.; IALE: Aberdeen, UK, 2001; pp. 177–186. [Google Scholar]
- Voigtländer, K. Chilopoda–Ecology. In Treatise on Zoology–Anatomy, Taxonomy, Biology. The Myriapoda; Minelli, A., Ed.; Brill: Leiden, The Netherlands, 2011; Volume 1, p. 546. [Google Scholar]
- Kubeš, J. Biocentres and corridors in a cultural landscape. A critical assessment of the “territorial system of ecological stability”. Landsc. Urban Plan. 1996, 35, 231–240. [Google Scholar] [CrossRef]
- Mackovčin, P. A multi-level ecological network in the Czech Republic: Implementating the Territorial System of Ecological Stability. GeoJournal 2000, 51, 211–220. [Google Scholar] [CrossRef]
- Bedford, S.E.; Usher, M.B. Distribution of arthropod species across the margins of farm woodlands. Agric. Ecosyst. Environ. 1994, 48, 295–305. [Google Scholar] [CrossRef]
- Maudsley, M.; Seeley, B.; Lewis, O. Spatial distribution patterns of predatory arthropods within an English hedgerow in early winter in relation to habitat variables. Agric. Ecosyst. Environ. 2002, 89, 77–89. [Google Scholar] [CrossRef]
- Stašiov, S.; Diviaková, A.; Svitok, M.; Novikmec, M.; Dovciak, M. Hedgerows support rich communities of harvestmen (Opiliones) in upland agricultural landscape. Basic Appl. Ecol. 2020, 47, 73–82. [Google Scholar] [CrossRef]
- Štrobl, M.; Saska, P.; Seidl, M.; Kocian, M.; Tajovský, K.; Řezáč, M.; Skuhrovec, J.; Marhoul, P.; Zbuzek, B.; Jakubec, P.; et al. Impact of an invasive tree on arthropod assemblages in woodlots isolated within an intensive agricultural landscape. Divers. Distrib. 2019, 25, 1800–1813. [Google Scholar] [CrossRef] [Green Version]
- Vanneste, T.; Govaert, S.; Spicher, F.; Brunet, J.; Cousins, S.A.O.; Decocq, G.; Diekmann, M.; Graae, B.J.; Hedwall, P.O.; Kapás, R.E.; et al. Contrasting microclimates among hedgerows and woodlands across temperate Europe. Agric. For. Meteorol. 2020, 281, 107818. [Google Scholar] [CrossRef]
- Kocourek, P.; Tajovský, K.; Dolejš, P. Mnohonožky České Republiky; ČSOP Vlašim: Vlasim, Czech Republic, 2017; p. 256. [Google Scholar]
- Šilhavý, V. Sekáči–Opilionidea. In Fauna ČSR, sv. 7; Nakladatelství ČSAV: Praha, Czech Republic, 1954; p. 272. [Google Scholar]
- Černecká, L.; Mihál, I.; Jarčuška, B. Response of ground-dwelling harvestman assemblages (Arachnida: Opiliones) to European beech forest canopy cover. Eur. J. Entomol. 2017, 114, 334–342. [Google Scholar] [CrossRef] [Green Version]
- Stašiov, S.; Michalková, E.; Lukáčik, I.; Čiliak, M. Harvestmen (Opiliones) communities in an arboretum: Influence of tree species. Biologia 2017, 72, 184–193. [Google Scholar] [CrossRef]
- Buchar, J.; Růžička, V. Catalogue of Spiders of Czech Republic; Peres: Praha, Czech Republic, 2002; p. 351. [Google Scholar]
- Kůrka, A.; Řezáč, M.; Macek, R.; Dolanský, J. Pavouci České Republiky; Academia: Praha, Czech Republic, 2015; p. 624. [Google Scholar]
- Chen, B.; Wise, D.H. Bottom-Up Limitation of Predaceous Arthropods in a Detritus-Based Terrestrial Food Web. Ecology 1999, 80, 761–772. [Google Scholar] [CrossRef]
- Thornhill, W.A. The distribution and probable importance of linyphiid spiders living on the soil surface of sugar-beet fields. Bull. Br. Arachnol. Soc. 1983, 6, 127–136. Available online: https://repository.rothamsted.ac.uk/item/97y07/the-distribution-and-probable-importance-of-linyphiid-spiders-living-on-the-soil-surface-of-sugar-beet-fields (accessed on 14 February 2023).
- Uetz, G.W. Habitat Structure and Spider Foraging. In Habitat Structure: The Physical Arrangement of Objects in Space; McCoy, E.D., Bell, S.S., Mushinsky, H.R., Eds.; Chapman and Hall: London, UK, 1991; pp. 325–348. [Google Scholar]
- Lövei, G.L.; Magura, T. Ground beetle (Coleoptera: Carabidae) diversity is higher in narrow hedges composed of a native compared to non-native trees in a Danish agricultural landscape. Insect Conserv. Divers. 2016, 10, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Sroka, K.; Finch, O.D. Ground beetle diversity in ancient woodland remnants in north-western Germany (Coleoptera, Carabidae). J. Insect Conserv. 2006, 10, 335–350. [Google Scholar] [CrossRef]
- Buchholz, S.; Tietze, H.; Kowarik, I.; Schirmel, J. Effects of a Major Tree Invader on Urban Woodland Arthropods. PLoS ONE 2015, 10, e0137723. [Google Scholar] [CrossRef] [Green Version]
- Scheu, S.; Poser, G. The soil macrofauna (Diplopoda, Isopoda, Lumbricidae and Chilopoda) near tree trunks in a beechwood on limestone: Indications for stemflow induced changes in community structure. Appl. Soil Ecol. 1996, 3, 115–125. [Google Scholar] [CrossRef]
- Stašiov, S.; Stašiová, A.; Svitok, M.; Michalková, E.; Slobodník, B.; Lukáčik, I. Millipede (Diplopoda) communities in an arboretum: Influence of tree species and soil properties. Biologia 2012, 67, 945–952. [Google Scholar] [CrossRef]
- Stašiov, S.; Diviaková, A.; Svitok, M.; Novikmec, M. Myriapod (Chilopoda, Diplopoda) communities in hedgerows of upland agricultural landscape. Biologia 2017, 72, 1320–1326. [Google Scholar] [CrossRef]
- Hůrka, K. Carabidae of the Czech and Slovak Republics; Kabourek: Zlín, Czech Republic, 1996; p. 565. [Google Scholar]
- Langellotto, G.A.; Denno, R.F. Responses of invertebrate natural enemies to complex-structured habitats: A meta-analytical synthesis. Oecologia 2004, 139, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Berthold, D.; Vor, T.; Beese, F. Effects of cultivating black locust (Robinia pseudoacacia L.) on soil chemical properties in Hungary. Forstarchiv 2009, 80, 307–313. [Google Scholar]
- Bååth, E.; Berg, B.; Lohm, U.; Lundgren, B.; Lundkvist, H.; Rosswall, T.; Söderström, B.; Wiren, A. Effects of experimental acidification and liming on soil organisms and decomposition in a scots pine forest. Pedobiologia 1980, 20, 85–100. [Google Scholar]
- Davey, J.S.; Vaughan, I.P.; Andrew King, R.; Bell, J.R.; Bohan, D.A.; Bruford, M.W.; Holland, J.M.; Symondson, W.O.C. Intraguild predation in winter wheat: Prey choice by a common epigeal carabid consuming spiders. J. Appl. Ecol. 2013, 50, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Uetz, G.W. Temporal and Spatial Variation in Species Diversity of Wandering Spiders (Araneae) in Deciduous Forest Litter. Environ. Entomol. 1975, 4, 719–724. [Google Scholar] [CrossRef]
- Uetz, G.W. Gradient analysis of spider communities in a streamside forest. Oecologia 1976, 22, 373–385. [Google Scholar] [CrossRef]
- Uetz, G.W. The influence of variation in litter habits on spider communities. Oecologia 1979, 40, 29–42. [Google Scholar] [CrossRef]


{kind=link}
| Carabidae | Araneae | |||||||
| a–d | s. r. | a–d | s. r. | |||||
| χ2 | p | F | p | χ2 | p | F | p | |
| area of wood fragment | 7.26 | *** | 2.96 | * | 6.57 | ** | 1.09 | n.s. |
| canopy coverage | 0.32 | n.s. | 3.06 | * | 5.92 | ** | 3.67 | * |
| herb layer coverage | 6.27 | n.s. | 1.42 | n.s. | 12.75 | *** | 0.72 | n.s. |
| dominance of grasses | 11.72 | *** | 6.53 | * | 8.71 | *** | 0.47 | n.s. |
| tree diversity | 9.71 | ** | 1.05 | n.s. | 3.36 | * | 11.95 | ** |
| soil moisture | 5.70 | * | 8.64 | *** | 5.26 | ** | 0.50 | n.s. |
| leaf litter thickness | 0.27 | n.s. | 0.64 | n.s. | 20.53 | *** | 1.17 | n.s. |
| R2 (adj.) | 0.499 | 0.451 | 0.581 | 0.325 | ||||
| Deviation explained | 64.5% | 53.4% | 74.6% | 41% | ||||
| Opiliones | Chilopoda | |||||||
| a–d | s. r. | a–d | s. r. | |||||
| χ2 | p | F | p | χ2 | p | F | P | |
| area of wood fragment | 0.17 | n.s. | 0.13 | n.s. | 0.24 | n.s. | 8.89 | *** |
| canopy coverage | 4.62 | ** | 4.93 | ** | 2.86 | * | 0.01 | n.s. |
| herb layer coverage | 1.73 | n.s. | 4.90 | ** | 0.29 | n.s. | 0.62 | n.s. |
| dominance of grasses | 1.66 | n.s. | 2.16 | n.s. | 6.64 | *** | 0.31 | n.s. |
| tree diversity | 2.73 | * | 1.97 | n.s. | 16.23 | *** | 7.34 | ** |
| soil moisture | 2.29 | n.s. | 0.26 | n.s. | 6.76 | *** | 3.60 | *** |
| leaf litter thickness | 0.54 | n.s. | 1.12 | n.s. | 11.26 | ** | 0.22 | n.s. |
| R2 (adj.) | 0.096 | 0.197 | 0.519 | 0.445 | ||||
| Deviation explained | 17.3% | 30.5% | 63.7% | 54% | ||||
| Diplopoda | Isopoda | |||||||
| a–d | s. r. | a–d | s. r. | |||||
| χ2 | p | F | p | χ2 | p | F | p | |
| area of wood fragment | 0.01 | n.s. | 1.79 | n.s. | 0.15 | n.s. | 0.28 | n.s. |
| canopy coverage | 9.65 | ** | 1.12 | n.s. | 1.93 | n.s. | 0.03 | n.s. |
| herb layer coverage | 10.79 | *** | 3.33 | * | 5.20 | ** | 0.56 | n.s. |
| dominance of grasses | 0.22 | n.s. | 0.41 | n.s. | 7.09 | ** | 0.25 | n.s. |
| tree diversity | 1.69 | n.s. | 0.25 | n.s. | 0.002 | n.s. | 0.66 | n.s. |
| soil moisture | 4.82 | n.s. | 0.20 | n.s. | 4.17 | ** | 5.31 | ** |
| leaf litter thickness | 11.08 | ** | 0.59 | n.s. | 8.09 | *** | 0.02 | n.s. |
| R2 (adj.) | −2.14 | 0.27 | -0.103 | 0.305 | ||||
| Deviation explained | 47.5% | 31% | 28% | 41.1% | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horňák, O.; Šarapatka, B.; Machač, O.; Mock, A.; Tuf, I.H. Characteristics of Fragments of Woodland and Their Influence on the Distribution of Soil Fauna in Agricultural Landscape. Diversity 2023, 15, 488. https://doi.org/10.3390/d15040488
Horňák O, Šarapatka B, Machač O, Mock A, Tuf IH. Characteristics of Fragments of Woodland and Their Influence on the Distribution of Soil Fauna in Agricultural Landscape. Diversity. 2023; 15(4):488. https://doi.org/10.3390/d15040488
Chicago/Turabian StyleHorňák, Ondřej, Bořivoj Šarapatka, Ondřej Machač, Andrej Mock, and Ivan Hadrián Tuf. 2023. "Characteristics of Fragments of Woodland and Their Influence on the Distribution of Soil Fauna in Agricultural Landscape" Diversity 15, no. 4: 488. https://doi.org/10.3390/d15040488