

The North Asian Genus Kolhymamnicola Starobogatov and Budnikova 1976 (Gastropoda: Amnicolidae), Its Extended Diagnosis, Distribution, and Taxonomic Relationships †
 , , and
, , and
Abstract
:1. Introduction
2. Material and Methods
3. Results
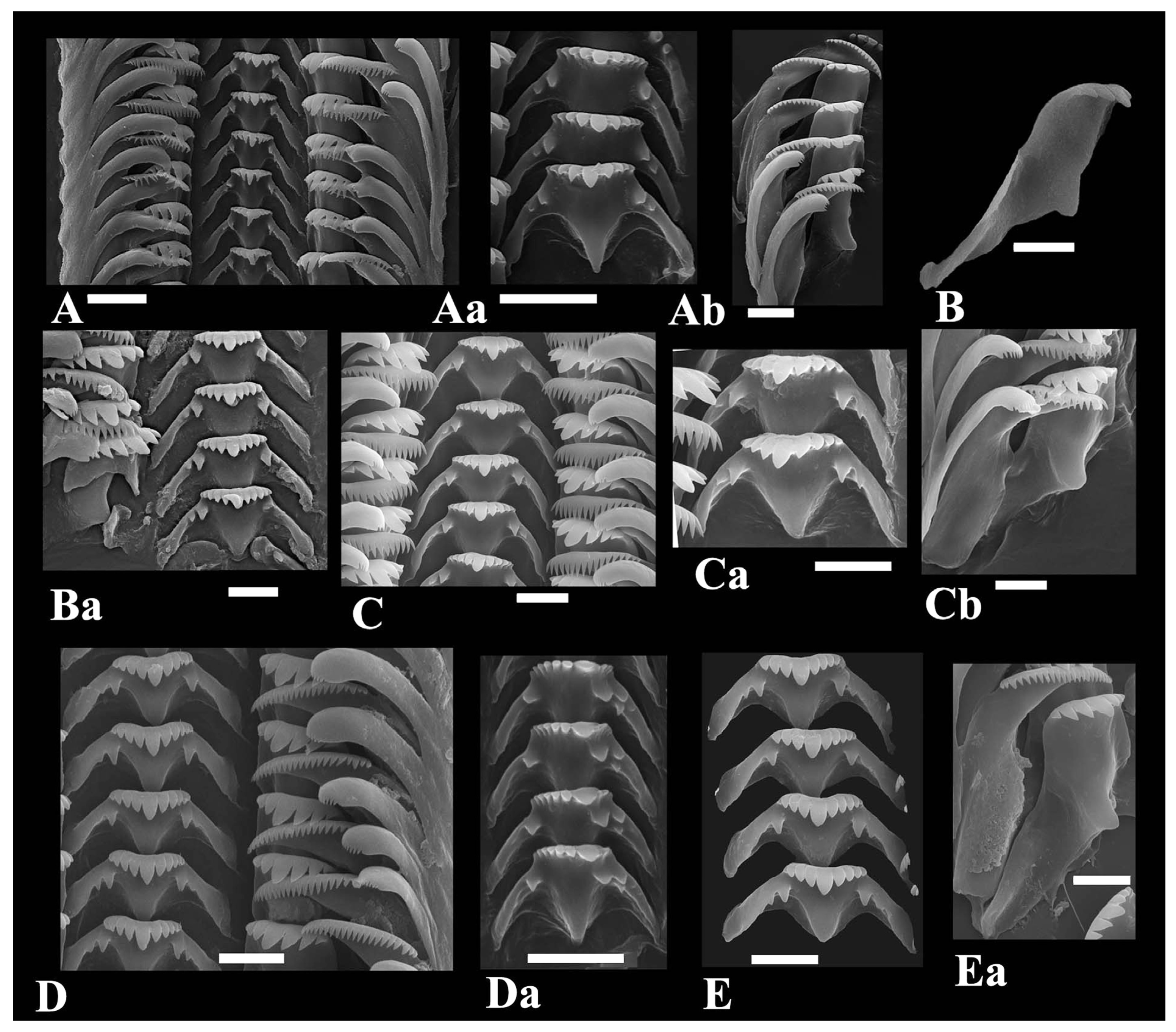
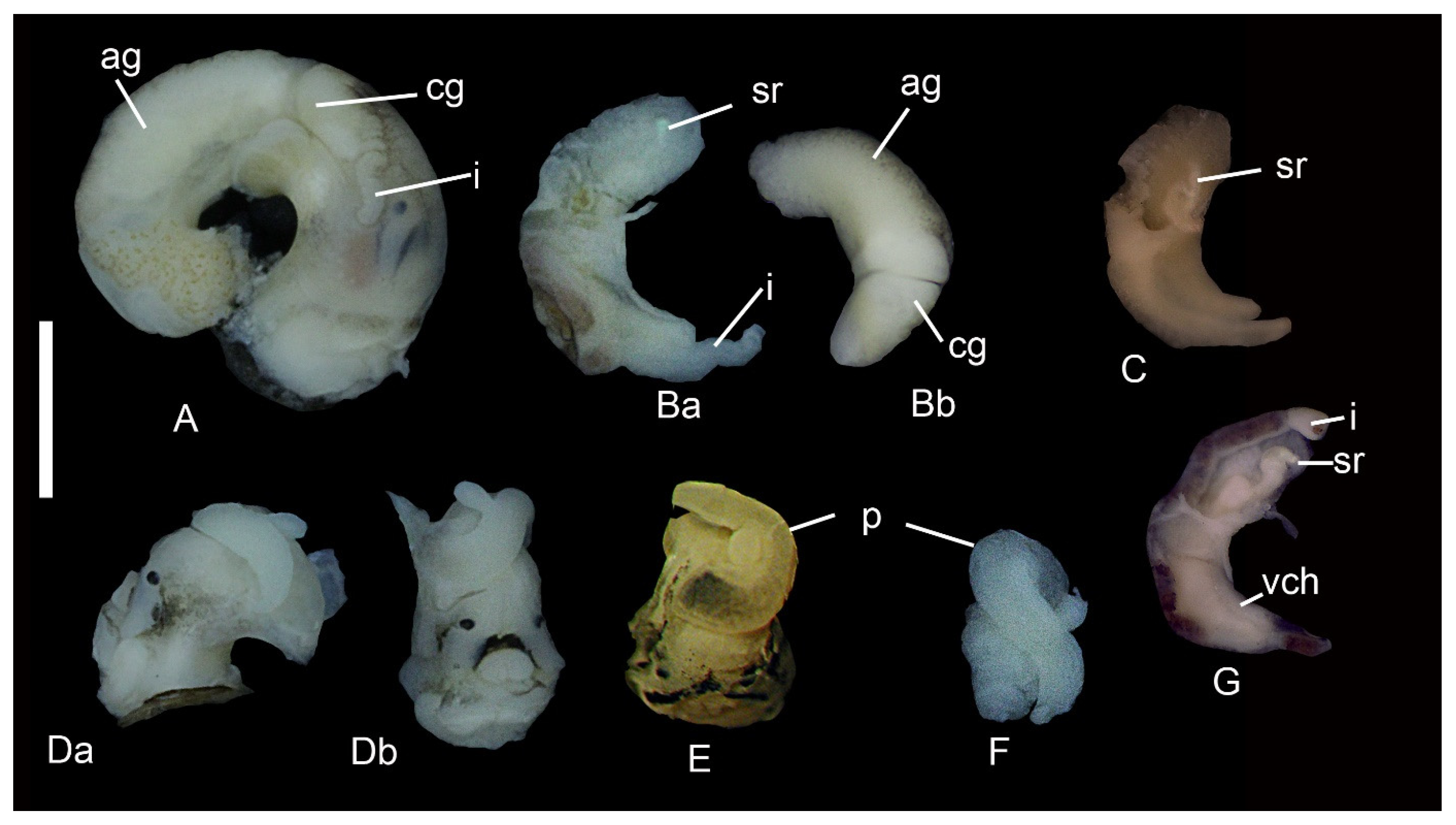
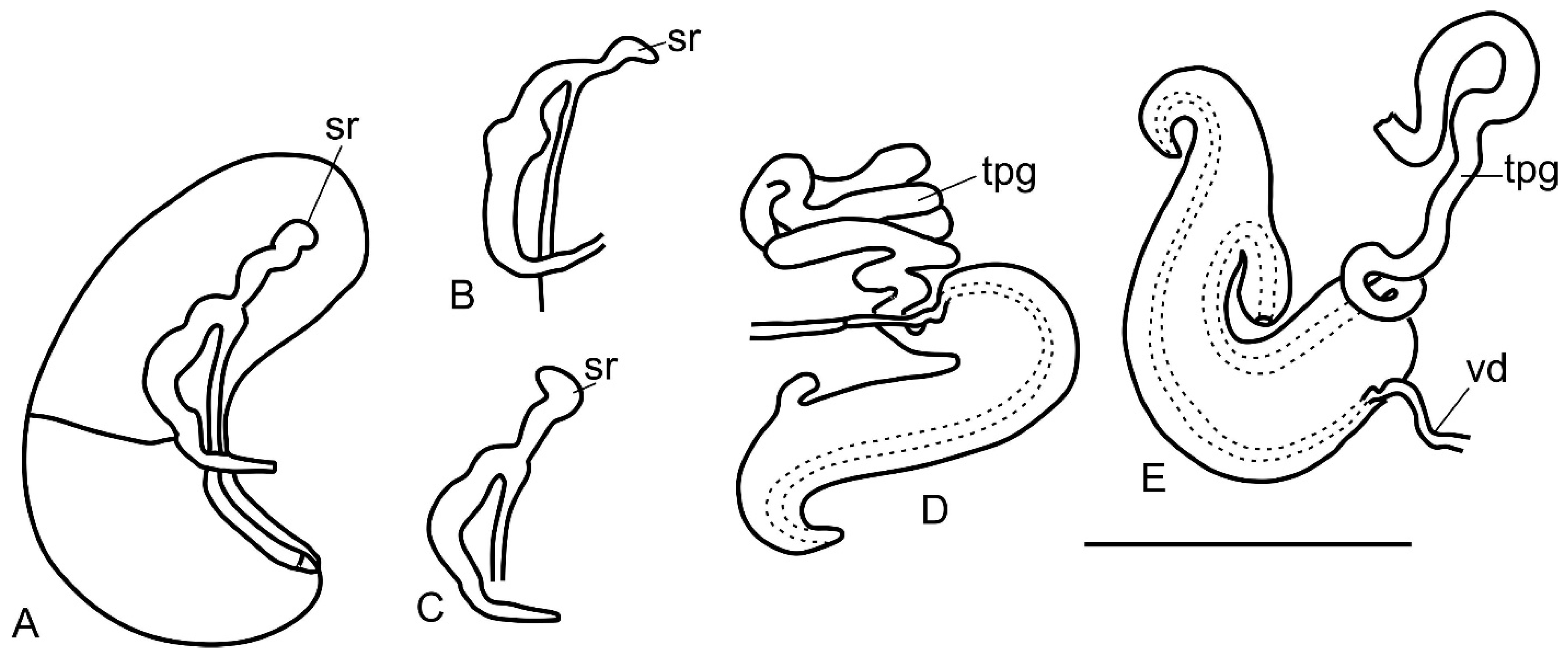
3.1. Morphology of Kolhymamnicola
3.2. A Morphological Comparison between Kolhymamnicola and Other Representatives of the Amnicolidae s. Lato
3.3. Molecular Phylogeny
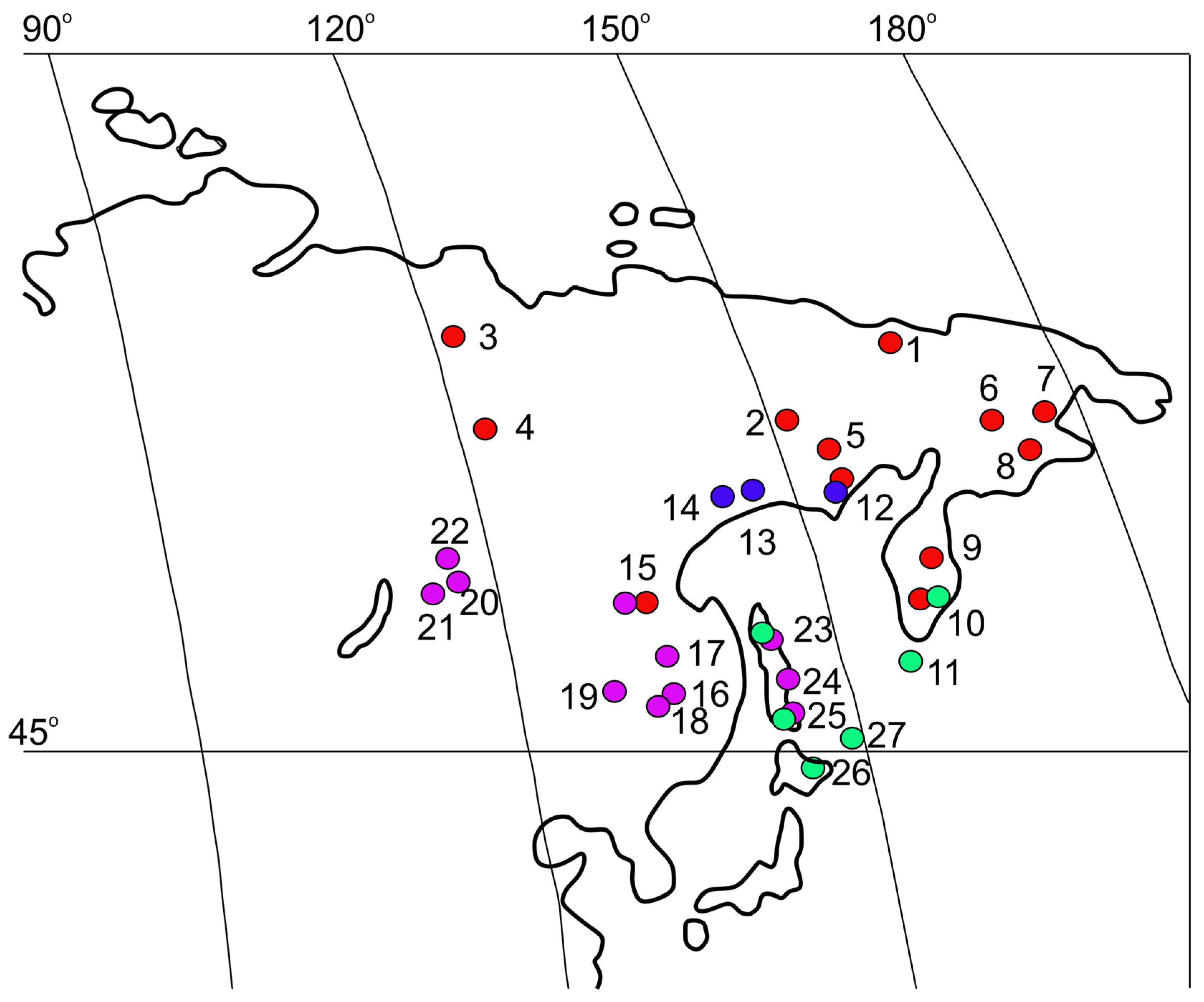
3.4. Kolhymamnicola Distribution and Habitats
4. Discussion
4.1. Species Composition and Radiation of the Genus Kolhymamnicola
4.2. The Fossil Amnicolidae s. lato and the Origin of the Kolhymamnicola
4.3. Notes on the Erhaiidae stat. nov.
4.4. The Taxonomic Status of Recent and Some Fossil Representatives of Baicaliidae
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilke, T.; Haase, M.; Hershler, R.; Liu, H.-P.; Misof, B.; Ponder, W. Pushing short DNA fragments to the limit: Phylogenetic relationships of ‘Hydrobioid’ gastropods (Caenogastropoda: Rissooidea). Mol. Phylogenet. Evol. 2013, 66, 715–736. [Google Scholar] [CrossRef] [PubMed]
- Bouchet, P.; Rocroi, J.-P.; Frýda, J.; Hausdorf, B.; Ponder, W.; Valdes, A.; Warén, A. A nomenclator and classification of gastropod family-group names. Malacologia 2005, 47, 1–368. [Google Scholar]
- Clark, S.A. Amnicolidae Tryon, pp. 90–93. In Freshwater Mollusks of the World. A distribution Atlas; Lydeard, C., Cummings, C.S., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2019; 242p. [Google Scholar]
- Sitnikova, T.Y.; Roepstorf, P.; Riedel, F. Reproduction, duration, of embryogenesis egg capsules and protoconchs of the family Baicaliidae (Gaenogastropoda) endemic to Lake Baikal. Malacologia 2001, 43, 59–85. [Google Scholar]
- Starobogatov, Y.I.; Prozorova, L.A.; Bogatov, V.V.; Saenko, E.M. Mollusks. In Key to Freshwater Invertebrates of Russia and Adjacent Lands; Tsalolikhin, S.Y., Ed.; Molluscs, polychaetes, nemerteans; Nauka: St Petersburg, Russia, 2004; Volume 6, pp. 9–491. (In Russian) [Google Scholar]
- Kantor, Y.I.; Sysoev, A.V. Catalogue of Mollusks of Russia and Adjacent Countries; KMK Scientific Press Ltd: Moscow, Russia, 2005; 627p. (In Russian) [Google Scholar]
- Vinarski, M.V.; Kantor, Y.I. Analytical Catalogue of Fresh and Brackish Water Molluscs of Russia and Adjacent Countries; A.N. Severtsov Institute of Ecology and Evolution of RAS: Moscow, Russia, 2016; 544p. [Google Scholar]
- Starobogatov, Y.I. Molluscan Fauna and Zoogeographic Zonation of Continental Freshwater Bodies of the World; Nauka: Leningrad, Russia, 1970; 372p. (In Russian) [Google Scholar]
- Gladstone, N.; Whelan, N. Pushing barcodes to their limits: Phylogenetic placement of Fontigens Pilsbry, 1933 (Caenogastropoda: Littorinimorpha: Truncatelloidea) and elevation of Fontigentidae Taylor. J. Molluscan Stud. 2022, 88, eyab038. [Google Scholar] [CrossRef]
- Starobogatov, Y.I.; Streletzkaja, E.A. Composition and Zoogeographical Characteristics of Freshwater Malacofauna of the East Siberia and Northern Part of the Far East. Mollusks and Their Role in Biocoenoses and Formation of Faunas; Trudy Zoologicheskogo Instituta AN SSSR: St Petersburg, Russia, 1967; Volume 42, pp. 221–268. (In Russian) [Google Scholar]
- Starobogatov, Y.I.; Budnikova, L.L. On the fauna of freshwater gastropod molluscs of the Far North-East of the USSR. Fresh-water fauna of the Chukchi Peninsula. Tr. Biol. Pochvennogo Inst. 1976, 36, 72–88. (In Russian) [Google Scholar]
- Zatravkin, M.N.; Bogatov, V.V. Amnicola-like gastropods from fresh waters of Far East of the USSR. Zool. Zhurnal 1988, 67, 127–129. (In Russian) [Google Scholar]
- Bogatov, V.V.; Zatravkin, M.N. Gastropod Molluscs of Fresh and Brackish Waters of the Far East of the USSR; A Guide; First Published 1990; FEB RAS: Vladivostok, Russia, 1992; 169p. (In Russian) [Google Scholar]
- Habe, T. A new amnicolid snail from the Kurile Islands. Venus 1942, 12, 25–27. [Google Scholar]
- Starobogatov, Y.I.; Sitnikova, T.Y. The system of the order Littoriniformes (Gastropoda, Pectinibranchia). Molluscs, Taxonomy, ecology and Regularities of Distribution. In Proceedings of the Seventh Meeting on the Investigation of Molluscs; Abstracts of communications; 5-7 April 1983; Nauka: Leningrad, Russia, 1983; pp. 18–22. (In Russian) [Google Scholar]
- Prozorova, L.A. Composition and biogeographic characteristics of gastropod malacofauna of the Chaun Depression. Bottom organisms of fresh waters of the Far East. In Proceedings of the Biological and Soil Institute, Far East Branch of the Russian Academy of Sciences; Far East Scientific Centre of the USSR Academy of Sciences: Vladivostok, Russia, 1986; pp. 39–47. (In Russian) [Google Scholar]
- Levanidova, I.M.; Levanidov, V.Y. Benthos of the Lake Azhabachye. Izv. TINRO 1972, 82, 51–92. (In Russian) [Google Scholar]
- Kurenkov, I.I. List of water invertebrates of Kamchatka inner water bodies. Izv. TINRO 1967, 57, 202–224. (In Russian) [Google Scholar]
- Prozorova, L.A.; Shed’ko, M.B. Molluscs of the Lake Azabachye and Their Biocenotic Importance; Trudy Kamchatskogo Filiala Tikhookeanskogo Instituta Geografii DVO RAN: 2003; pp. 120–151. (In Russian)
- Dolgin, V.N. Freshwater Mollusks of Sub-Arctic and Arctic Siberia. Ph.D. Thesis, Tomsk State University, Tomsk, Russia, 2001; 40p. (In Russian). [Google Scholar]
- Prozorova, L.A. Freshwater and terrestrial molluscs of the Tauya Bay coast. In Biological Diversity of the Okhotsk Bay. Diversity of the Tauya Bay of the Okhotsk Sea; Dalnauka: Vladivostok, Russia, 2005; pp. 252–261. (In Russian) [Google Scholar]
- Bogatov, V.V.; Sirotsky, S.E. Production of mollusks and their role in the biocenoses of floodplain lakes in the Upper Zeya River. In Hydrobiology of the Amur Basin; Lebedev, I.M., Ed.; Dalnauka: Vladivostok, Russia, 1978; pp. 116–122. (In Russian) [Google Scholar]
- Koshelev, V.N.; Kolobov, V.Y. Feeding of juvenile Kaluga and Amur sturgeon in the Amur River estuary. Vestn. ASTU: Fish. Ind. 2013, 1, 20–28. (In Russian) [Google Scholar]
- Sitnikova, T.Y.; Prozorova, L.A.; Regel, K.V. Embryonic shell, distribution, and a problem of origin of Kolhymamnicola gastropods. Zoolog. Zhurn. 2010, 89, 1190–1199. (In Russian) [Google Scholar]
- Prozorova, L.A. Gastropods and small bivalves of fresh and brackish waterbodies in the southern Kurile Islands. Annotated list of species. Bull. Russ. Far East Malacol. Soc. 1997, 1, 21–34. (In Russian) [Google Scholar]
- Prozorova, L.A.; Bogatov, V.V.; Saenko, E.M. New data on the freshwater mollusk fauna of the Sakhalin Island. In Flora and Fauna of the Sakhalin Island (Materials of the International Sakhalin Project); RAS DVO: Vladivostok, Russia, 2004; Volume 1, pp. 138–144. (In Russian) [Google Scholar]
- Pietsch, T.W.; Bogatov, V.V.; Storozhenko, S.Y.; Lelej, A.S.; Barkalov, V.Y.; Takahashi, H.; Joneson, S.L.; Kholin, S.K.; Minakawa, N.; Ohara, M.; et al. Biodiversity and biogeography of Sakhalin Islands. Materials of International Kuril and Sakhalin island Projects. In Flora and Fauna of North-West Pacific Islands; Storozhenko, S.Y., Ed.; Dalnauka: Vladivostok, Russia, 2012; pp. 11–78. (In Russian) [Google Scholar]
- Kuroda, T.; Habe, T. Troglobiontic aquatic snails from Japan. Venus 1958, 19, 183–196. [Google Scholar]
- Laforsch, C.; Tollrian, R. A new preparation technique of daphnids for Scanning Electron Microscopy using hexamethyldisilazane. Arch. Hydrob. 2000, 149, 587–596. [Google Scholar] [CrossRef]
- Radoman, P. Hydrobioidea a Superfamily of Prosobranchia (Gastropoda): Systematics; Serbian Academy of sciences and arts: Belgrade, Serbia, 1983; Volume 1, 256p. [Google Scholar]
- Szarowska, M. Molecular phylogeny, systematics and morphological character evolution in the Balkan Rissooidea (Caenogastropoda). Folia Malacol. 2006, 14, 99–168. [Google Scholar] [CrossRef]
- Hershler, R.; Frest, T.J.; Johannes, E.J.; Bower, P.A.; Thompson, F.G. Two new genera of Hydrobiid snails (Prosobranchia: Rissooidea) from the Northwestern United States. Veliger 1994, 37, 221–243. [Google Scholar]
- Hershler, R.; Liu, H.-P.; Frest, T.J.; Jonannes, E.; Clark, W. Genetic structure of the Western North American aquatic gastropod genus Taylorconcha and description of a second species. J. Molluscan Stud. 2006, 72, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Hershler, R.; Thompson, F.G. Notes on Morphology of Amnicola limosa (Say, 1817) (Gastropoda: Hydrobiidae) with Comments on Status of the Subfamily Amnicolinae. Malacol. Rev. 1988, 22, 81–92. [Google Scholar]
- Davis, G.M.; Chen, C.E.; Wu, C.; Kuang, T.F.; Xing, X.G.; Li, L.; Liu, W.J.; Yan, Y.L. The Pomatiopsidae of Hunan, China (Gastropoda: Rissoacea). Malacologia 1992, 34, 143–342. [Google Scholar]
- Davis, G.M.; Rao, S. Discovery of Erhaia Gastropoda: Pomatiopsidae) in Northern India with description of a new genus Erhaiini from China. Procced. Acad. Nat. Sci. Phil. 1997, 148, 273–299. [Google Scholar]
- Dybowski, W. Die Gastropoden-Fauna des Baikal-Sees, anatomisch und systematisch bearbeitet. Mem. de L’academie Impériale des Sci. de St. Petersbourg 1875, 22, 1–73. [Google Scholar]
- Kozhov, M.M. Mollusks of Baikal Lake. Taxonomy, distribution, ecology, some data on genesis and history. Tr. Baikal’skoi Limnol. Stantsii 1936, 8, 1–320. (In Russian) [Google Scholar]
- Kozhov, M.M. On the morphology and history of the Baikal endemic molluscs of the family Baicaliidae. Tr. Baikal’skoi Limnol. Stantsii 1951, 13, 93–119. (In Russian) [Google Scholar]
- Sitnikova, T.Y. New structure of Baikalian endemic family Baicaliidae (Mollusca: Pectinibranchia). In Morphologiya i Evolytsiya Bespozvonochnyh. Fauna of Baikal Lake; Linevich, A.A., Ed.; Nauka: Novosibirsk, Russia, 1991; pp. 281–295. (In Russian) [Google Scholar]
- Sokolov, E.P. An improved method for DNA isolation from mucopolysaccharide-rich molluscan tissues. J. Molluscan Stud. 2000, 66, 573–575. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. “BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT”. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587. [Google Scholar] [CrossRef] [Green Version]
- Schrempf, D.; Minh, B.Q.; von Haeseler, A.; Kosiol, C. Polymorphism-aware species trees with advanced mutation models, bootstrap, and rate heterogeneity. Mol. Biol. Evol. 2019, 36, 1294–1301. [Google Scholar] [CrossRef] [Green Version]
- Anisimova, M.; Gil, M.; Dufayard, J.E.; Dessimoz, C.; Gascuel, O. Survey of branch support methods demonstrates accuracy, power, and robustness of fast likelihood-based approximation schemes. Syst. Biol. 2011, 60, 685–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef] [Green Version]
- Wilke, T.; Schultheiß, R.; Albrecht, C. As time goes by: A simple fool’s guide to molecular clock approaches in invertebrates. Am. Malacol. Bull. 2009, 27, 25–45. [Google Scholar] [CrossRef]
- Stadler, T.; Kouyos, R.; von Wyl, V.; Yerly, S.; Böni, J.; Bürgisser, P.; Klimkait, T.; Joos, B.; Rieder, P.; Xie, D.; et al. Estimating the basic reproductive number from viral sequence data. Molec. Biol. Evol. 2012, 29, 347–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Davis, G.M.; Kuo, Y.H.; Hoagland, K.E.; Chen, P.L.; Yang, H.M.; Chen, D.J. Erhaia, a new genus and new species of Pomatiopsidae from China (Gastropoda: Rissoacea). Proc. Acad. Nat. Sci. Phila. 1985, 137, 48–78. [Google Scholar]
- Wilke, T.; Benke, M.; Brändle, M.; Albrecht, C.; Bichain, J.-M. The neglected side of the coin: Non-adaptive radiations in spring snails (Bythinella spp.). In Evolution in Action. Case Studies in Adaptive Radiation, Speciation and the Origin of Biodiversity; Glaubrecht, M., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 551–578. [Google Scholar]
- Bichain, J.-M.; Gaubert, P.; Samadi, S.; Boisselier-Dubayle, M.-C. A gleam in the dark: Phylogenetic species delimitation in the confusing spring-snail genus Bythinella Moquin-Tandon, 1856 (Gastropoda: Rissooidea: Amnicolidae). Mol. Phyl. Evol. 2007, 45, 927–941. [Google Scholar] [CrossRef]
- Haase, M.; Wilke, T.; Mildner, P. Identifying species of Bythinella (Caenogastropoda: Rissooidea): A plea for an integrative approach. Zootaxa 2007, 1563, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Falniowski, A.; Szarowska, M.; Sȋrbu, I. Bythinella Moquin-Tandon, 1856 (Gastropoda: Rissooidea: Bythinellidae) in Romania: Its morphology with description of four new species. Folia Malacol. 2009, 17, 21–36. [Google Scholar] [CrossRef] [Green Version]
- Falniowski, A.; Hofman, S.; Rysiewska, A. A new species of Bythinella Moquin-Tandon, 1855 (Caenogastropoda: Truncatelloidea) From Naxos Island, Greece. Folia Malacol. 2016, 24, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Falniowski, A.; Szarowska, M. Radiation and phylogeography in a spring snail Bythinella. (Mollusca: Gastropoda: Rissooidea) in continental Greece. Ann. Zool. Fennici 2011, 48, 67–90. [Google Scholar] [CrossRef]
- Szarowska, M.; Osikowski, A.; Hofman, S.; Falniowski, A. Do diversity patterns of the spring-inhabiting snail Bythinella (Gastropoda, Bythinellidae) on the Aegean Islands reflect geological history? Hydrobiologia 2016, 765, 225–243. [Google Scholar] [CrossRef] [Green Version]
- Bezverkhniy, V.L.; Pletnev, S.P.; Nabiullin, A.A. Outline of the geological structure and development of the Kuril Island System and adjacent regions. In Flora and Fauna of the Kuril Islands: Results of the International Kuril Island Project; Dalnauka: Vladivostok, Russia, 2002; pp. 9–22. (In Russian) [Google Scholar]
- Hopkins, D.M. Cenozoic history of the Bering land Bridge. Science 1959, 129, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- Gladenkov, A.Y.; Gladenkov, Y.B. The beginning of formation of interoceanic connections between the Pacific and the Arctic through the Bering Strait in the Neogene. Stratigr. Geol. Correl. 2004, 12, 72–88. (In Russian) [Google Scholar]
- Jiang, D.; Klaus, S.; Zhang, Y.-P.; Hillis, D.M.; Li, J.T. Asymmetric biotic interchange across the Bering land bridge between Eurasia and North America. Natl. Sci. Rev. 2019, 6, 739–745. [Google Scholar]
- Galanin, A.V. Beringia: The Past and The Present (Instead of Foreword). Magadan: Prospects for Beringia in 21-st Century; NESC FEB RAS: Magadan, Russia, 2004; pp. 5–19. Available online: http://ukhtoma.ru/rhythm1.htm (accessed on 17 January 2023).
- Martinson, G.G. Mesozoic and Cenozoic molluscs from continental deposits of the Siberian platform. Trans-Baikal Area and Mongolia. Proc. Baikal Limnol. st. 1961, 19, 1–332. (In Russian) [Google Scholar]
- Popova, S.M. Cenozoic Continental Malacofauna of Southern Siberia and Adjacent Areas; Nauka: Moscow, Russia, 1981; 185p. (In Russian) [Google Scholar]
- Pan, H.; Zhu, X. Early Cretaceous non-marine gastropods from the Xiazhuang Formation in North China. Cretaceous Res. 2007, 28, 215–224. [Google Scholar] [CrossRef]
- Yen, T.-C. Molluscan Fauna of the Morrison Formation. US Geol. Surv. Prof. Paper 1952, 233, 21–51. [Google Scholar]
- Yen, T.-C. Nonmarine mollusks of late cretaceous age from Wyoming, Utah and Colorado. Part 1: A fauna from western Wyoming. US Geol. Surv. Prof. Paper 1954, 254, 45–59. [Google Scholar]
- Evanof, E.; Good, S.C.; Hanley, J.H. An overview of the freshwater mollusks from the Morrison Formation (Upper Jurassic, Western Interior, USA). Mod. Geol. 1998, 22, 423–450. [Google Scholar]
- Neubauer, T.A.; Harzhauser, M.; Hartman, J.H.; Silvestro, D.; Scotese, C.R.; Czaja, A.; Vermeij, G.J.; Wilke, T. Short-term paleogeographic reorganizations and climate events shaped diversification of North American freshwater gastropods over deep time. Sci. Rep. 2022, 12, 15572. [Google Scholar] [CrossRef] [PubMed]
- Gittenberger, E.; Gyeltshen, C.; Stelbrink, B. The genus Erhaia (Gastropoda, Truncatelloidea, Amnicolidae), with a new species from Bhutan. Zookeys 2022, 1085, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Starobogatov, Y.I.; Sitnikova, T.Y. The Lists of species of animals and plants inhabiting in Baikal. Mollusca. In Lake Baikal: Evolution and Biodiversity; Kozhova, O.M., Izmest’eva, L.R., Eds.; Backuys Publishers: Leiden, The Netherlands, 1998; pp. 404–413. [Google Scholar]
- Sitnikova, T.Y.; Starobogatov, Y.I.; Shirokaya, A.A.; Shibanova, I.V.; Korobkova, N.V.; Adov, F.V. Gastropod molluscs (Gastropoda). In Index of Animal Species Inhabiting Lake Baikal and Its Catchment Area; Timoshkin, O.A., Ed.; Nauka: Novosibirsk, Russia, 2004; Volume 1, pp. 937–1002. (In Russian) [Google Scholar]
- Sherbakov, D.Y. Molecular phylogenetic studies on the origin of biodiversity in Lake Baikal. Trends Ecol. Evol. 1999, 14, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Zubakov, D.Y.; Sherbakov, D.Y.; Sitnikova, T.Y. Phylogeny of the endemic Baicaliidae molluscs inferred from partial nucleotide sequences of the CO1 mitochondrial gene. Russ. J. Mol. Biol. 1997, 31, 935–939. (In Russian) [Google Scholar]
- Fazalova, V.; Nevado, B.; Peretolchina, T.; Petunina, J.; Sherbakov, D. When environmental changes do not cause geographic separation of fauna: Differential responses of Baikalian invertebrates. BMC Evol. Biol. 2010, 10, 320. [Google Scholar] [CrossRef] [Green Version]
- Kovalenkova, M.V.; Sitnikova, T.Y.; Shcherbakov, D.Y. Genetic and morphological diversification in gastropods of the family Baicaliidae. Ecolog. Genet. 2013, 11, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Kovalenkova, M.V.; Bukin, Y.S.; Sitnikova, T.Y.; Sherbakov, D.Y. The phylogeny based on the intron of ATP synthase α-subunit gene reveals a substrate role in the origin of the gastropod species flock from Lake Baikal. Limnol. Freshw. Biol. 2020, 2, 405–411. [Google Scholar]
- Peretolchina, T.E.; Sitnikova, T.Y.; Sherbakov, D.Y. The complete mitochondrial genomes of four Baikal molluscs from the endemic family Baicaliidae (Caenogastropoda: Truncatelloida). J. Moll. Stud. 2020, 86, 201–209. [Google Scholar]
- Starobogatov, Y.I. New species of gastropods from springs and subterranean waters of Middle Asia. Proceed. Zool. Inst. 1972, 50, 165–172. (In Russian) [Google Scholar]
- Sitnikova, T.Y.; Sysoev, A.V.; Kijashko, P.V. Species of the fresh water gastropods described by Ya. I. Starobogatov: Pulmonata (Acroloxidae), Heterobranchia (Valvatidae) и Caenogastropoda (Viviparoidea, Truncatelloidea and Cerithioidea). Proc. Zool. Inst. 2017, 321, 247–299. [Google Scholar]
- Popova, S.M.; Mats, V.D.; Chernyaeva, G.P.; Shimaraeva, M.K.; Kulchitsky, A.A.; Vorobyeva, G.A.; Klimanova, V.M.; Kononov, E.E.; Kravchinsky, A.Y.; Kulagina, N.V.; et al. Paleolimnological Reconstructions Baikal Rift Zone; Nauka: Novosibirk, Russia, 1989; pp. 1–110. (In Russian) [Google Scholar]
- Mats, V. The Baikal rift: Pliocene (Miocene) Quaternary episode or product of extended development since the Late Cretaceous under various tectonic factors. A review. Geodyn. Tectonoph. 2015, 6, 467–489. [Google Scholar] [CrossRef] [Green Version]
- Sitnikova, T.; Teterina, V.; Maximova, N.; Kirilchik, S. Discordance of genetic diversification between deep-and shallow-water species of Kobeltocochlea Lindholm, 1909 (Caenogastropoda: Truncatelloidea: Benedictiidae) endemic to Lake Baikal with the description of a new species, review of the genus, and notes on its origin. J. Zool. System. Evol. Res. 2021, 59, 1775–1797. [Google Scholar]
- Martinson, G.G. Baikalian mollusks in the Tertiary deposits of Xinjiang. In Memory of Academician, L.S. Berg: Collected Essays on Geography and Biology; The USSR Academy of Sciences Press: Moscow-Leningrad, Russia, 1955; pp. 524–527. (In Russian) [Google Scholar]
- Martinson, G.G. Presnovodnyye mollyuski iz al’bskikh otlozheniy Bayan-Khongorskogo aymaka [Freshwater mollusks from Albian deposits of Bain-Chongor Ajmak]. In Fauna Mezozoya i Kaynozoya Zapadnoj Mongolii; Trofimov, B.A., Ed.; Nauka: Moskva, Russia, 1971; pp. 7–13. (In Russian) [Google Scholar]
- Martinson, G.G. Pozdnemelovyye mollyuski Mongolii: Sistematika, stratigrafiya, tafonomiya [Late Cretaceous mollusks of Mongolia: Taxonomy, stratigraphy, taphonomy]. Tr. Sovmest. Sov. Mong. Paleontol. Ekspeditsiya 1982, 17, 1–82. (In Russian) [Google Scholar]
- Martinson, G.G.; Popova, S.M. Tretichnyye mollyuski baykal’skogo tipa iz ozernykh otlozheniy yuga Zapadnoy Sibiri [Tertiary Baikal-type mollusks from lacustrine sediments in the south of Western Siberia]. Paleontol. Zh. 1959, 4, 105–109. (In Russian) [Google Scholar]
- Popova, S.M.; Devyatkin, Y.V.; Starobogatov, Y.I. Mollyuski Kyzylgirskoy Svity Gornogo Altaya [Molluscs of the Kyzylgir Formation of the Altay Mountains]; Nauka: Moskva, Russia, 1970; 95p. (In Russian) [Google Scholar]
- Yü, W.; Wang, H.-J. The late Cretaceous and Cenozoic Gastropoda from Jiangsu Province. Mem. Nan. Inst. Geol. Pal. Acad. Sin. 1977, 8, 1–90. [Google Scholar]
- Guo, F.; Wei, J.-M. Gastropoda. In Paleontological Atlas of Northwestern China, Volume Xinjiang; Geological Survey Department of Bureau of Petroleum of Xinjiang Autonomous Region, Regional Geological Survey Team, Bureau of Geology, Mineral Resources of Xinjiang Autonomous Region, Eds.; Geological Publishing House: Beijing, China, 1984; Volume 3, pp. 83–93. [Google Scholar]
- Thiele, J. Handbuch der Systematischen Weichtierkunde; Fischer: Jena, Germany, 1931; Volume 1, pp. 377–778. [Google Scholar]
- Riedel, F.; Healy, J.M.; Ropstorf, P.; Sitnikova, T. Ecology, shell morphology, anatomy and sperm ultrastructure of the Caenogastropod Pyrgula annulata, with a discussion of the relationship between the ‘Pyrgulidae’ and Caspian and Baikalian Rissooideans. Limnologica 2001, 31, 289–302. [Google Scholar] [CrossRef] [Green Version]
- Martinson, G.G. Tretichnaya fauna mollyuskov Vostochnogo Pribaykal’ya [Tertiary mollusk fauna of the Eastern Baikal region]. Tr. Baikal. Limnolog. Stan. Akad. Nauk. SSSR 1951, 13, 5–92. (In Russian) [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Palumbi, S.R. Nucleic acids II: The polymerase chain reaction. In Molecular Systematics, 2nd ed.; Hillis, D.M., Moritz, C., Mable, B.K., Eds.; Sinauer Associates Inc.: Sunderland, MA, USA, 1996; pp. 205–247. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Locality (Numbers Correspond to Names in Figure 1 | Collection, Museum Acronym, Registration Number | Examined Specimens |
|---|---|---|---|
| K. kolhymensis | (1) Lower Kolhyma River basin, near Chersky settlement, 68°44′ N 161°21′ E (type locality) | ZIN, No. 1, 4 IBSS, No. 1418 | Holotype and 2 paratypes; 5 dry shells 2 alcohol-fixed females (juvenile and mature) |
| Kolhyma River basin, lakes | ZIN, No. 6 | 2 dry shells (paratypes) | |
| (2) Middle Kolhyma River basin, an unnamed floodplain thermokarst lake, 66°39′ N 152°35′ E | IBSS, No. 2104 | 1 juvenile female | |
| (7) Chukotka Peninsula, a small lake, the Avtaatkui River basin, near Anadyr city, | IBSS, No. 1397 | 4 dry shells | |
| (12) Coast of the Okhotsk Sea, Perevolochnaya Bay, Yama River valley, an unnamed lake, 59°43′ N 149°46′ E. | IBSS, No. 2120b | 3 juvenile females, 1 male | |
| K. ochotica | (14) Okhota River basin, Lake Bolshoe Uyeginskoe, 59°20′ N 143°01′ E (type locality) | ZIN, No.1 | Holotype (dry shell) |
| (13) Kava River basin, coast of the Okhotsk Sea, 59°38′ N 149°05′ E | IBSS, No. 4325 | 1 juvenile female, 2 males | |
| (13) Yama River basin, Lake Tynerynda, 59°46′ N 149°23′ E; | IBSS, No. 2431 | 1 alcohol-fixed female, 3 dry shells | |
| K. wasiliewae | (16) Lower Amur River basin, Bolin River (backwater), vicinity of Amursk Town, 50°19′ N 136°48′ E (type locality). | ZIN, No. 1; IBSS, leg. L. Prozorova, 8 July 1980. | Holotype, 1 juvenile female and 1 male, 3 dry shells |
| (18) Bolshoy In River, a tributary of the Tunguska River, 48°36′ N 133°39′ E. | IBSS, leg. V. Makarenko & L. Prozorova | 8 juveniles (used in the molecular analysis) | |
| (20) Leprindo Lake, 56°37′ N 117°31′ E | LIN, leg. A. Matveev, 1997 | 1 male | |
| (22) Vicinity of Vitimsky Nature Reserve, lake in the Lapagar terrain, 57°15′ N 116°24′ E. | LMBI, No. 05-032 | 6 dry shells, 1 alcohol-fixed female | |
| (15) Upper Zeya River basin, 50°14′ N 127°34′ E. | IBSS, leg. L. Prozorova | 1 dry shell | |
| K. miyadii | (11) Kurile Archipelago, Shumshu Island, Lake Bolshoye, 50°45′ N 156°15′ E (type locality). | ZIN, No. 1 and 2 | Holotype, 3 paratypes |
| The same locality as above | IBSS, No. 111-97. | 4 dry shells, 3 alcohol-fixed specimens (2 females, 1 male) | |
| (23) Sakhalin Island, a floodplain lake near Tymovski settlement, the Tym River basin, 51°39′ N 142°56′ E; | LMBI, No. 05-031 | 4 fixed specimens | |
| (27) Kurile Archipelago, Zelenyi Island, Lake Sredneye, 43°29′ N 146°08′ E | IBSS, No. 518 | 5 dry shells, 5 fixed specimens (1 female, 4 males) |
| Species/Character | K. kolhymensis | K. wasiliewae | K. ochotica | K. miyadi | K. miyadi |
|---|---|---|---|---|---|
| [13] | [14] | ||||
| Shell: | |||||
| shape | elongated conical | conical | ovoid-conical | ovoid | conical |
| height | 2.75–3.25 mm | 4.0 mm | 4.0 mm | <3.25 mm | 3.1 mm |
| color | green-brown | green or yellow-green | light brown | yellow, brown, or green | corneous brown |
| whorls number | 3.74–4.25 | 3.5–4.0 | 3.5–4.0 | 3.5–4.5 | 5.0 |
| whorl shape | rounded, slightly shouldered | ovate | rounded, shouldered | rounded, slightly shouldered | conspicuously inflated |
| spire height/shell height (Sph/SH) | ~0.6 | <0.5 | <0.5 | >0.5 | >0.5 |
| Aperture: shape | ovate | ovate | ovate | rounded | oblique, rounded pyriform |
| parieto-palatal corner | rounded | rounded, slightly narrowing | rounded, slightly narrowing | ovoid, obtuse angulated | slightly angled |
| Umbilicus | wide slit | narrow slit | slit, slightly covered by columellar lip | wide slit | very narrow perforated |
| Own Data | [14] | ||||
| Species/Character | K. kolhymensis | K. wasiliewae | K. ochotica | K. miyadi | K. miyadi |
| Formulae of the central radula tooth | |||||
| Formulae for other teeth (see the text) | L (3–1–3) + MI (20) + MII (15) | L (3–1–3) + MI (~18-20) + MII (8–10) | L [3–1–2-(3)] + MI (~18) + MII (12–16) | L (3–1–3) + MI (20) + MII (10) | L (7) + M (~30) |
| Penis proximal base | slightly broad | narrow | broad | narrow | no data |
| Penis lobe and distal part of a penis | rather equal or ~2 times shorter | ~3 times shorter than distal part | ~2 times shorter than distal part | ~3–3.5 times shorter than distal part | no data |
| Penis tip | no papilla | with short papilla | no? papilla | no data | no data |
| Seminal receptacle | straight | turns right | turns left or nearly straight | turns left | |
| Species | SH (Shell Height) | SW (Shell Width) | SpH (Spire Height) | AL (Aperture Length) | AW (Aperture Width) | Whorl Number |
|---|---|---|---|---|---|---|
| K. kolhymensis | ||||||
| holotype | 2.75 | 2.2 | 1.5 | 1.05 | 3.7 | |
| (n = 6) * | 2.75 ± 0.27 (2.3–3.0) | 2.03 ± 0.14 (1.8–2.2) | 1.52 ± 0.23 (1.2–1.8) | 1.13 ± 0.14 (1.0–1.3) | 1.15 ± 0.10 (1.0–1.3) | 3.8 ± 0.3 (3.5–4.25) |
| K. ochotica | ||||||
| holotype | 4.2 | 2.6 | 2.4 | 1.8 | 1.4 | 4.0 |
| (n = 5) * | 2.78 ± 0.47 (2.2–3.5) | 2.09 ± 0.24 (1.9–2.5) | 1.59 ± 0.18 (1.0–1.3) | 1.30 ± 0.10 (1.2–1.4) | 1.14 ±0.11 (1.0–1.3) | 4.0 ± 0.3 (3.5–4.25) |
| K. miyadii | ||||||
| syntype | 3.1 | 2.2 | 1.5 | 1.1 | 5.0 | |
| (n = 12) * | 2.4 ± 0.32 (1.9–2.9) | 1.92 ± 0.20 (1.8–2.3) | 1.23 ± 0.24 (0.8–1.5) | 1.19 ± 0.12 (1.0–1.4) | 0.98 ± 0.10 (0.8–1.1) | 3.66 ± 0.31 (3.25–4.25) |
| K. wasiliewae | ||||||
| holotype | 2.7 | 2.05 | 1.3 | 1.4 | 0.9 | 4.0 |
| (n = 5) * | 2.56 ± 0.35 (2.2–3.0) | 1.94 ± 0.24 (1.6–2.2) | 1.38 ±0.18 (1.2–1.6) | 1.26 ± 0.21 (1.0–1.5) | 1.0 ± 0.14 (0.8–1.1) | 3.85 ± 0.33 (3.5–4.0) |
| Characters | Taxa, Sources of Data | ||||||
|---|---|---|---|---|---|---|---|
| Kolhymamnicola | Taylorconcha | Marstoniopsis/Parabythinella | Erhaia | Akiyoshia/Saganoa | Amnicola | Baicaliidae | |
| [12,13], Own Data | [32,33] | [31] | [35,36] | [35,53] | [34] | [4,37,40], Own Data | |
| Shell height | up to 4 mm | up to 4 mm | <6 mm | ≤3 mm | <2 mm | up to 7 mm | 2–25 mm |
| Shell shape | ovate-conical or conical | globose to ovate-conical | ovate-conical | ovate-turreted or turreted | turreted | ovate-conical | ovate-conical, globose, conical, turreted, elongated-conical etc. |
| Shell whorl number | 3.5–5 | 3.5–4.5 | 4–4.5 | <5.5 | 4.5–5.5 | 4–6 | 3.5–12.0 |
| Shell sculpture | smooth | smooth | smooth | smooth | smooth | Smooth | smooth or sculptured (with ribs, carinae, spiral threads etc.) |
| Protoconch sculpture | smooth or with a few short spiral lines | fine spiral lines | delicate spiral lines | smooth with a few spiral lines near suture or spiral microsculpture | ?/smooth | smooth, sometimes with a few fine spiral lines | with spiral lines, striae or lirae |
| Operculum, position of the nucleus | submarginal | submarginal | submarginal | significant submarginal | significant submarginal | submarginal | submarginal or close to central position |
| Size and position of osphradium along ctenidium | 1/3 of gill length, posterior to middle position | slightly anterior to middle | ? | slightly posterior | ?/middle | ¼ of gill length, posterior | from 1/3 to 1/6 of gill length, anterior to middle |
| Number of ctenidium leaflets (filaments) | ~24–32 (n = 8) | ~17 | ? | <17 | ?/11–14 | 25–35 | 20–130 (n = 19) |
| Bursa copulatrix | absent | absent | absent | present | ?/present | Present | absent |
| Seminal receptacle | present, rs2 | present, rs2 | present, rs1 | present, rs1 | ?/absent, renal loop functions as seminal receptacle | present, rs1 | absent, folded left part of the coil loop functions as seminal receptacles and bursa, some species with temporary rs1 or/and rs2 |
| Ventral channel (groove) or spermathecal duct | ventral channel | ventral channel | spermathecal duct | spermathecal duct | ?/spermathecal duct | spermathecal duct | ventral channel |
| Penis simple or with a lobe (bifid) | bifid | simple | bifid | simple | ?/simple | Bifid | simple |
| The accessory penial gland | present | absent | absent | absent | ?/absent | Present | penial gland is represented by a lamina on the penis base |
| Penis tip with or without papilla | with short or without papilla | without papilla | ? | with or without papilla | ?/with short papilla | ? | with or without papilla |
| Central radular tooth: shape | trapezoidal or broadly trapezoidal | broadly trapezoidal | broadly trapezoidal | trapezoidal | trapezoidal | broadly trapezoidal | square or rectangular |
| Shape of basal tongue | V-shaped | V-shaped | V-shaped | V-shaped | V-shaped | V-shaped | hollow, straight or slightly convex |
| Formulae of the central radular tooth | |||||||
| Distribution, typical habitats | Northern Asian mainland, Kurile islands, small lakes | Northwestern USA, rivers | Europe, Balkan peninsula, lakes | China, India, small streams | Japan, China, caves, drilled pipe wells, streams | Eastern part of the USA, creeks, swamps, lakes | Lake Baikal, various substrates and depths (from 1.5 to ~300 m) |
| Species | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Taylorconcha serpenticola | ||||||||||
| 2 | Taylorconcha insperata | 0.012 | |||||||||
| 3 | Kolhymamnicola wasiliewae | 0.113 | 0.113 | ||||||||
| 4 | Marstoniopsis graeca | 0.118 | 0.118 | 0.126 | |||||||
| 5 | Akiyoshia kobayashii | 0.148 | 0.148 | 0.155 | 0.166 | ||||||
| 6 | Erhaia jianouensis | 0.159 | 0.159 | 0.162 | 0.154 | 0.142 | |||||
| 7 | Moria kikuchii | 0.155 | 0.152 | 0.160 | 0.159 | 0.099 | 0.145 | ||||
| 8 | Baicaliidae cl. 1 | 0.142 | 0.144 | 0.145 | 0.137 | 0.161 | 0.138 | 0.162 | |||
| 9 | Baicaliidae cl. 2 | 0.141 | 0.144 | 0.152 | 0.154 | 0.149 | 0.162 | 0.154 | 0.065 | ||
| 10 | Amnicola limosa | 0.164 | 0.169 | 0.172 | 0.184 | 0.174 | 0.162 | 0.171 | 0.159 | 0.156 | |
| 11 | Amnicola dalli | 0.160 | 0.166 | 0.172 | 0.186 | 0.167 | 0.171 | 0.174 | 0.154 | 0.154 | 0.027 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sitnikova, T.; Peretolchina, T.; Prozorova, L.; Sherbakov, D.; Babushkin, E.; Vinarski, M. The North Asian Genus Kolhymamnicola Starobogatov and Budnikova 1976 (Gastropoda: Amnicolidae), Its Extended Diagnosis, Distribution, and Taxonomic Relationships. Diversity 2023, 15, 483. https://doi.org/10.3390/d15040483
Sitnikova T, Peretolchina T, Prozorova L, Sherbakov D, Babushkin E, Vinarski M. The North Asian Genus Kolhymamnicola Starobogatov and Budnikova 1976 (Gastropoda: Amnicolidae), Its Extended Diagnosis, Distribution, and Taxonomic Relationships. Diversity. 2023; 15(4):483. https://doi.org/10.3390/d15040483
Chicago/Turabian StyleSitnikova, Tatiana, Tatiana Peretolchina, Larisa Prozorova, Dmitry Sherbakov, Eugeny Babushkin, and Maxim Vinarski. 2023. "The North Asian Genus Kolhymamnicola Starobogatov and Budnikova 1976 (Gastropoda: Amnicolidae), Its Extended Diagnosis, Distribution, and Taxonomic Relationships" Diversity 15, no. 4: 483. https://doi.org/10.3390/d15040483