
How an Ecological Race Is Forming: Morphological and Genetic Disparity among Thermal and Non-Thermal Populations of Aquatic Lymnaeid Snails (Gastropoda: Lymnaeidae)
 , , , and
, , , and
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subclass: Family | Species Name | Distribution | Reference |
|---|---|---|---|
| Neritimorpha: Neritidae | Theodoxus prevostianus (C. Pfeiffer, 1828) | Eastern Europe (Austria, Hungary) | [50] |
| Th. fluviatilis fluviatilis (Linnaeus, 1758); Th. f. thermalis (Dupuy, 1851) | England (Derbyshire); France (Pyrenees) | [51,52] | |
| Caenogastropoda: Thiaridae | Melanoides tuberculata (O.F. Müller, 1774) | Central Europe (Austria, Hungary, Romania *), Indonesia (Sulawesi and Sumatra Islands), USA (Wyoming, Yellowstone National Park, Grand Teton National Park) *; New Zealand, Jordan * | [53,54,55,56,57,58,59]; our data |
| Melanoides pamirica Lindholm, 1930 M. shahdaraensis Starobogatov & Izzatullaev, 1980 | Tajikistan (Pamir Mts); West Himalaya (M. pamiricus) | [60] | |
| M. kainarensis Starobogatov & Izzatullaev, 1980 | Turkmenistan; North Afghanistan | ||
| Caenogastropoda: Bithyniidae | Bithynia tentaculata (Linnaeus, 1758) | Austria, Hungary, England (Derbishire) | [44,51,61,62] |
| ‘Bythinia’ rumelica Wohlberedt, 1911 | Bulgaria | [63] | |
| ‘Bithynia thermalis’ | Italy (Lucca) | [64] | |
| Boreoelona contortrix (Lindholm, 1909) | Russia (Siberia, Baikal area) | [65] | |
| B. sibirica (Westerlund, 1886) | Russia (Siberia, Baikal area) | [66] | |
| Caenogastropoda: Cochliopidae | Kainarella minima Starobogatov, 1972 | Turkmenistan | [67] |
| Lobogenes michaelis Pilsbry &Bequaert, 1927 | Zaire | [68] | |
| Pseudocaspia starostini Starobogatov, 1972 | Turkmenistan | [67] | |
| Semisalsa aponensis (von Martens, 1858) | Italy | [69] | |
| S. foxianensis (De Stefani, 1883) | Italy | ||
| Tryonia monitorae Hershler, 1999 | USA (Nevada) | [70] | |
| Tryonia chuviscarae Hershler, Liu & Landye, 2011 T. julimesensis Hershler, Liu & Landye, 2011 T. minckleyi Hershler, Liu & Landye, 2011 | Mexico (Chichuachua) | [71] | |
| Caenogastropoda: Tateidae | Austropyrgus nanus Clark, Miller & Ponder 2003 Austropyrgus solitarius Clark, Miller & Ponder 2003 | Australia (Tasmania) | [72] |
| ‘Hydrobia jenkinsi’ = Potamopyrgus antipodarum (Gray, 1843) | England (Derbishire) | [51] | |
| Potamopyrgus sp. | New Zealand | [73] | |
| Caenogastropoda: Tomichiidae | Tomichia hendrickxi (Verdcourt, 1950) | Zaire | [68] |
| Caenogastropoda: Bythinellidae | Bythinella rubiginosa (Boubée, 1833) | France | [74] |
| B. schmidtii (Küster, 1852) | Austria | [54] | |
| B. thermophila Glöer, Varga & Mrkvicka, 2015 | Hungary | [75] | |
| Caenogastropoda: Lithoglyphidae | Fluminicola seminalis (Hinds, 1842) | USA (California) | [76] |
| Caenogastropoda: Hydrobiidae | Belgrandia thermalis (Linnaeus, 1767) | Italy (Pisa) | [1,77] |
| Belgrandiella mimula Haase, 1996 | Austria | [78] | |
| Pseudamnicola letourneuxiana (Bourguignat, 1862) | Algeria | [79] | |
| Pyrgulopsis bruneauensis Hershler, 1990 | USA (Idaho) | [80] | |
| P. bryantwalkeri Hershler, 1994 = Fluminicola nevadensis (Walker, 1916 P. nevadensis (Stearns, 1883) | USA (Nevada) | [76] | |
| P. thermalis (D.W. Taylor, 1987) | USA (New Mexico) | [81] | |
| Hadziella thermalis Bole, 1992 | Slovenia | [82] | |
| ‘Iglica’ veklovrhi De Mattia, 2007 | [83] | ||
| Radomaniola bulgarica Glöer & Georgiev, 2009 | Bulgaria | [84] | |
| Caenogastropoda: Stenothyridae | Stenothyra thermaecola Kuroda, 1962 | Japan (Kyushu Island) | [85] |
| Caenogastropoda: Semisulcospiridae | Juga plicifera (I. Lea, 1838) J. nigrina (I. Lea, 1856) | USA (California) | [76] |
| Caenogastropoda: Melanopsidae | Melanopsis etrusca (Brot, 1862) | Italy (Tuscany Region) | [86] |
| M. daudebartii thermalis (Brot, 1868) | Austria, Hungary | ||
| M. parreyssii (Philippi, 1847) | Bulgaria, Romania | [63,87] | |
| Melanopsis sp. | Cyprus | own observations | |
| Caenogastropoda: Viviparidae | Angulyagra costata (Quoy & Gaimard, 1834) | Indonesia (Sulawesi) | [53] |
| Viviparus contectus (Millet, 1813) | Austria | [54] | |
| Heterobranchia: Acroloxidae | Acroloxus lacustris (Linnaeus, 1758) | Hungary | [62] |
| Heterobranchia: Chilinidae | Chilina patagonica Sowerby, 1874 | Patagonia | [9] |
| Heterobranchia: Lymnaeidae | Ampullaceana balthica (Linnaeus, 1758) **. (Referred to as Lymnaea peregra (Müller) and Radix pereger var. ovata (Draparnaud)) | Iceland, French Pyrenees | [2,7,43] |
| A. lagotis (Schrank, 1803) [referred to as Lymnaea zazurnensis; L. ovata, L. fontinalis] | Northeast European Russia (Nenets Autonomous Region); Siberia (Baikal area); East Kazakhstan | [6,66,88], this study | |
| Austropeplea tomentosa (L. Pfeiffer, 1855) | New Zealand | [73] | |
| Galba bowelli (Preston, 1909) | Russia (Siberia, Baikal area) | [66] | |
| G. sibirica (Westerlund, 1885) | Russia (Siberia, Baikal area) | [66] | |
| G. truncatula (O.F. Müller, 1774) | Austria, Iceland, Hungary, Tajikistan | [54,62,89]; own observations | |
| Kamtschaticana kamtschatica (Middendorff, 1850) | Russian Far East and Eastern Siberia | [19,47] | |
| Ladislavella catascopium (Say, 1817) | USA (Yellowstone Park), Russia (Kamchatka Peninsula), Canada (British Columbia and Yukon Provinces) | [16,46]; this study | |
| Ladislavella elodes (Say, 1821) | USA (Nevada, Yellowstone National Park) | [76,90] | |
| Lanx patelloides (I. Lea, 1856) | USA (California) | [76] | |
| Lymnaea stagnalis (Linnaeus, 1758) | Austria | [54] | |
| Peregriana peregra (O.F. Müller, 1774) | Croatia, Ireland | [91,92] | |
| Pseudosuccinea columella (Say, 1817) | Austria * | [54] | |
| Radix alticola (Izzatullaev, Kruglov & Starobogatov, 1983) | Tajikistan, Nepal | [19,89] | |
| R. auricularia (Linnaeus, 1758) | Europe (Austria), Russia (Baikal area, Kamchatka), Afghanistan; Tibet | [9,47,48,54,93,94] | |
| R. ‘persica’ (Issel, 1865) | Iran | [95] | |
| R. rubiginosa (Michelin, 1831) | Indonesia (Sulawesi, Flores and Sumatra) | [53]; our data | |
| R. rufescens (Gray, 1822) [referred to as Lymnaea acuminata] | India (Himalaya) | [49] | |
| Stagnicola palustris (O.F. Müller, 1774) | Austria, Hungary | [54,62] | |
| Tibetoradix kozlovi Vinarski, Bolotov & Aksenova, 2018 | China (Tibetan Plateau) | [96] | |
| Heterobranchia: Physidae | ‘Physa fontinalis’ (Linnaeus, 1758) | New Zealand | [73] |
| ‘Physa thermalis’ Locard, 1899 = Paraplexa cornea (Massot, 1845) | France | [97] | |
| Physella acuta (Draparnaud, 1805), including Ph. acuta var. thermalis (Boettger, 1913) | USA (Virginia, Utah, Yellowstone National Park, Alaska), Europe (Poland, Slovakia) *; Abkhazia; Russia (North Caucasus) ** | [5,46,54,97]; our data | |
| Ph. cooperi (Tryon, 1865) | USA (California) | [76,90] | |
| Ph. johnsoni (Clench, 1926) | Canada (Alberta) | ||
| Ph. osculans (Haldeman, 1841) | USA (California) | ||
| Ph. propinqua (Tryon, 1865) | USA (Idaho) | ||
| Ph. smithiana Baker, 1920 | USA (Nevada) | ||
| Ph. spelunca Turner & Clench, 1974 | USA (Wyoming) | [5] | |
| Ph. virginea (Gould, 1847) | USA (New Mexico, Nevada, Utah) | [90] | |
| Ph. wrighti Te & Clarke, 1985 | Canada (British Columbia) | [4] | |
| Ph. johnsoni (Clench, 1926) | USA | ||
| Heterobranchia: Planorbidae | Ancylus fluviatilis O.F. Müller, 1774 | Austria | [54] |
| Anisus spirorbis (Linnaeus, 1758) | Hungary | [62] | |
| A. vorticulus (Troschel, 1834) | |||
| Armiger annandalei (Germain, 1918) | East Kazakhstan; Tajikistan (Pamir Mts.) | [98]; our data | |
| Bathyomphalus contortus (Linnaeus, 1758) | Austria; Russia (Siberia, Baikal area) | [54] | |
| Biomphalaria costata (Biese, 1951) B. thermalus (Biese, 1951) | Chile | [99] | |
| Ferrissia fragilis (Tryon, 1863) ** | Austria, Hungary | [54,62] | |
| Gyraulus acronicus (Férussac, 1807) | Russia (Kamchatka) | [14] | |
| G. albus (O.F. Müller, 1774) | Austria | [54] | |
| G. baicalicus (Dybowski, 1913) | Russia (Siberia, Baikal area) | [66] | |
| G. borealis Lovén in Westerlund, 1875 | Russia (Siberia, Baikal area) | [66] | |
| G. chereshnevi (Prozorova & Starobogatov, 1997) | Russia (Chukchi Peninsula) | [100] | |
| G. convexiusculus (Hutton, 1849) | Afghanistan | [94] | |
| G. cf. ignotellus (Dybowski, 1913) | Russia (Siberia, Baikal area) | [66] | |
| G. iwaotakii (Mori, 1938) | Japan (Honshu Island) | [20,101] | |
| G. ladacensis (Nevill, 1878) | Tajikistan (Pamir Mts.) | [98] | |
| G. laevis (Alder, 1838)–described as Planorbis thermalis Westerlund, 1885 | Austria, Northeast European Russia (Nenets Autonomous Region) | [14,102]; our data | |
| G. pankongensis (Martens, 1882) | Tajikistan (Pamir Mts.) | [89] | |
| G. parvus (Say, 1817) | Iceland | [14] | |
| G. takhteevi Sitnikova & Peretolchina, 2018 | Russia (Siberia, Baikal area) | [10] | |
| G. thermochukchensis (Prozorova & Starobogatov, 1997) | Russia (Chukchi Peninsula) | [100] | |
| Helisoma anceps (Menke, 1830) | USA (Nevada) | [76] | |
| H. trivolvis (Say, 1817) | Austria * | [54] | |
| Planorbarius corneus (Linnaeus, 1758) | Hungary | [62] | |
| Planorbella duryi (Wetherby, 1879) | Hungary | [55] | |
| P. subcrenata (Carpenter, 1857) | USA (Nevada) | [76] | |
| Planorbis planorbis (Linnaeus, 1758) | Austria, Hungary | [44,54] | |
| P. sieversi Mousson, 1873 | Afghanistan | [94] | |
| Vorticifex effusus (I. Lea, 1856) | USA (California, Nevada) | [76] | |
| Heterobranchia: Valvatidae | Valvata cristata O.F. Müller, 1774 | Hungary | [62] |
| V. pamirensis Starobogatov, 1972 | Tajikistan (Pamir Mts.) | [67] | |
| V. sibirica (Middendorff, 1851) | Russia (Siberia, Baikal area) | [65] |
2. Material & Methods
2.1. Material Examined
2.2. Morphological Study and Statistical Analysis
2.3. DNA Analysis, Sequence Alignment and Phylogeography
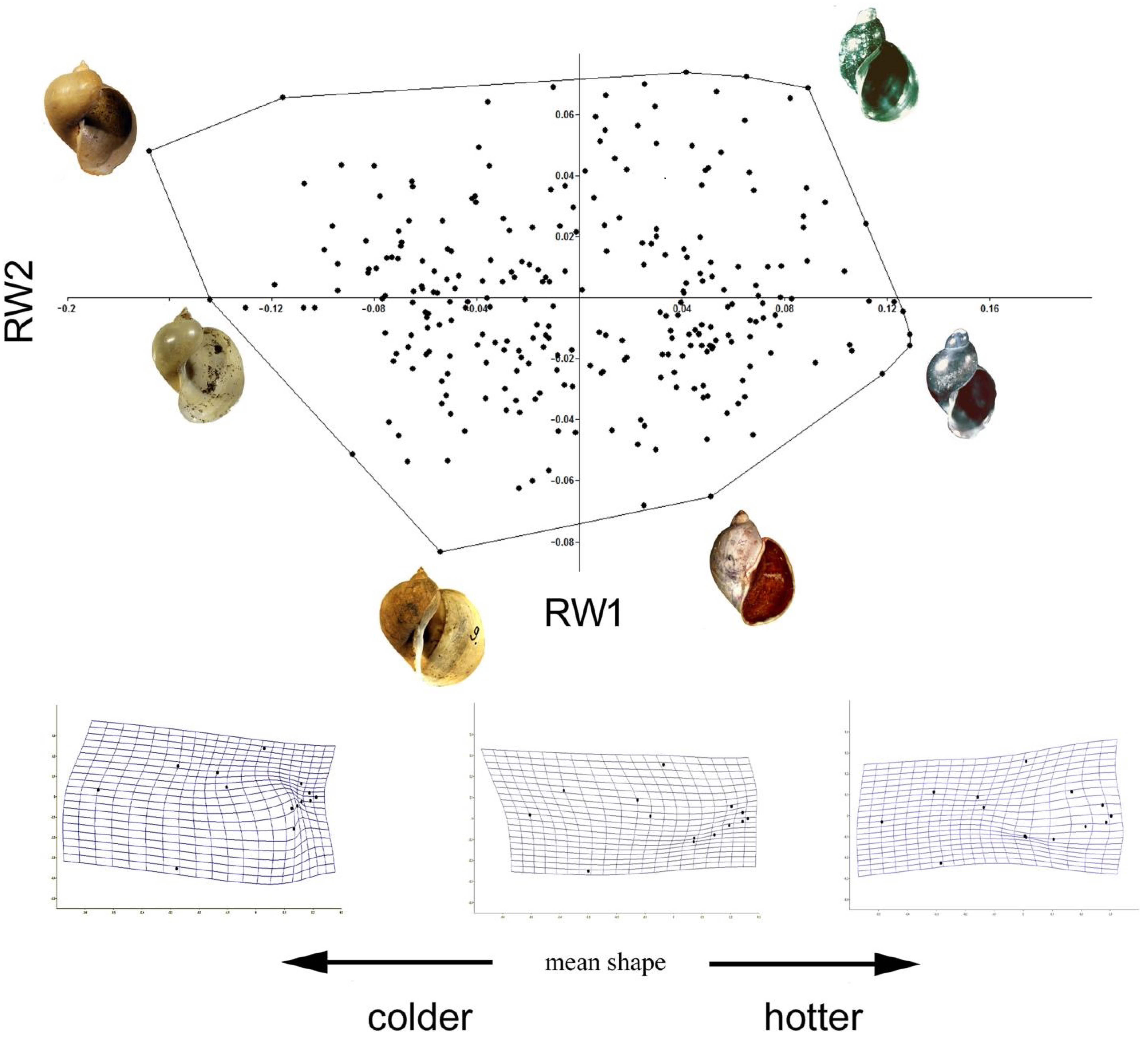
3. Results
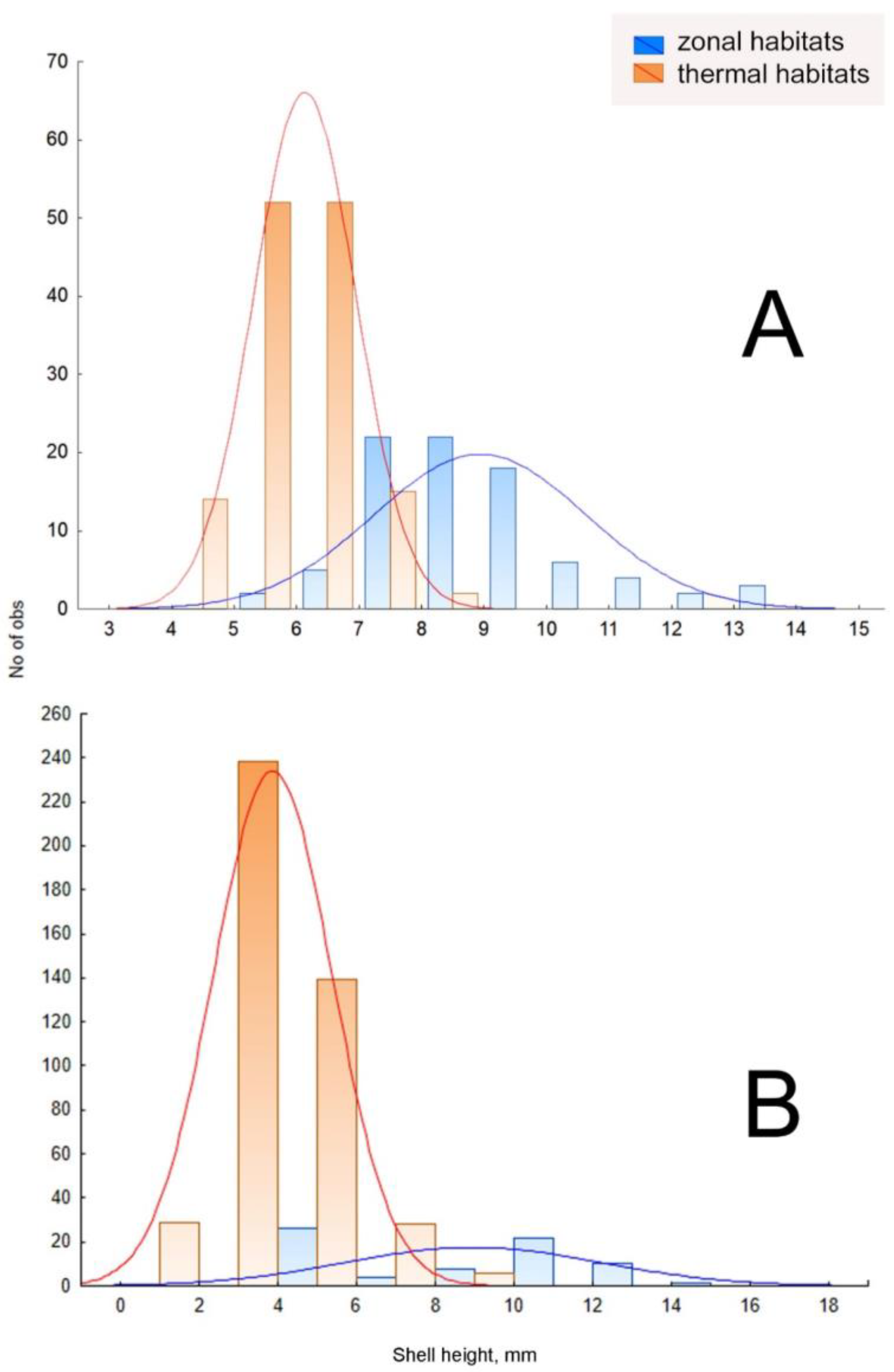
3.1. Variation in Shell Size and Proportions
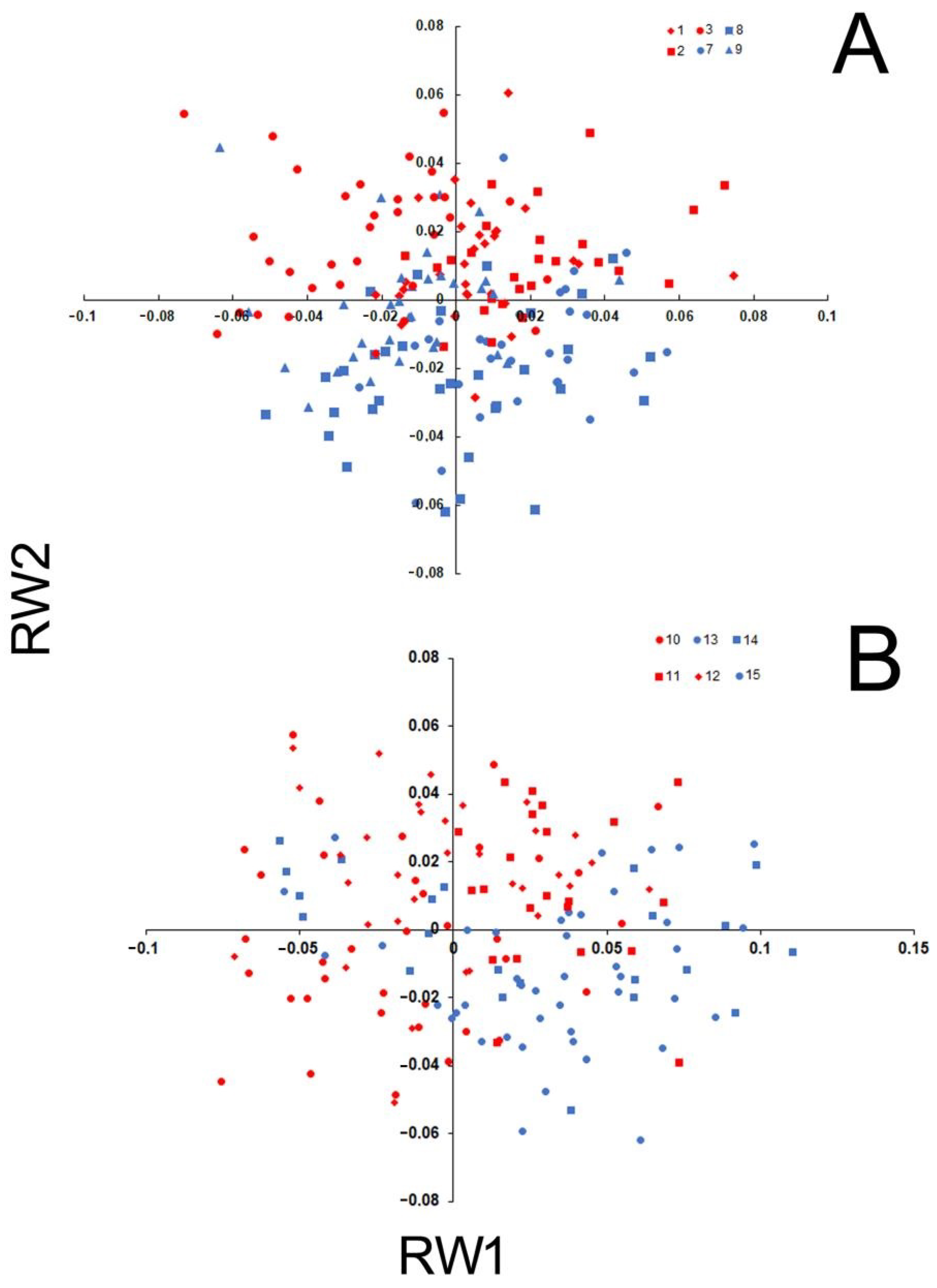
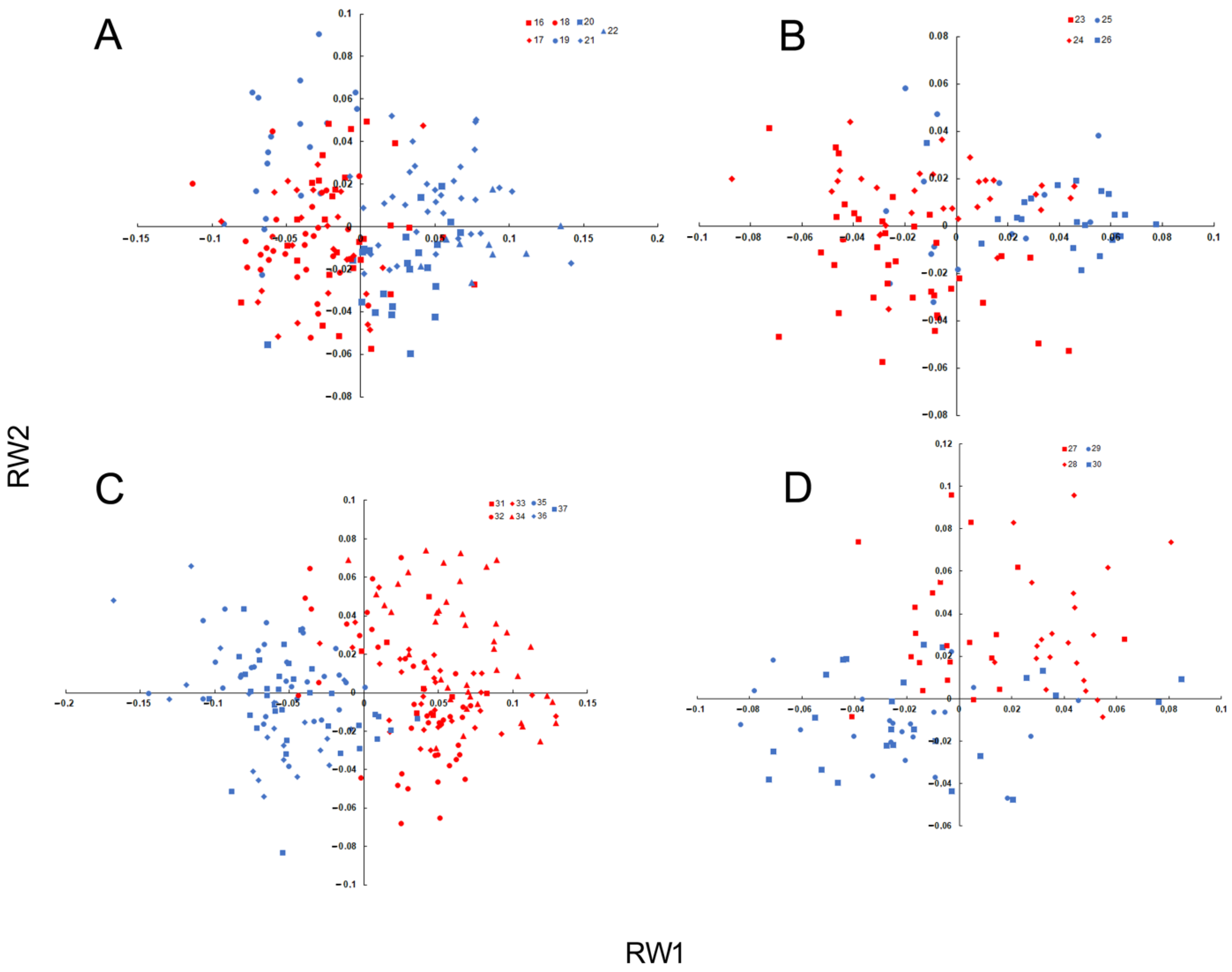
3.2. Anatomical Variation
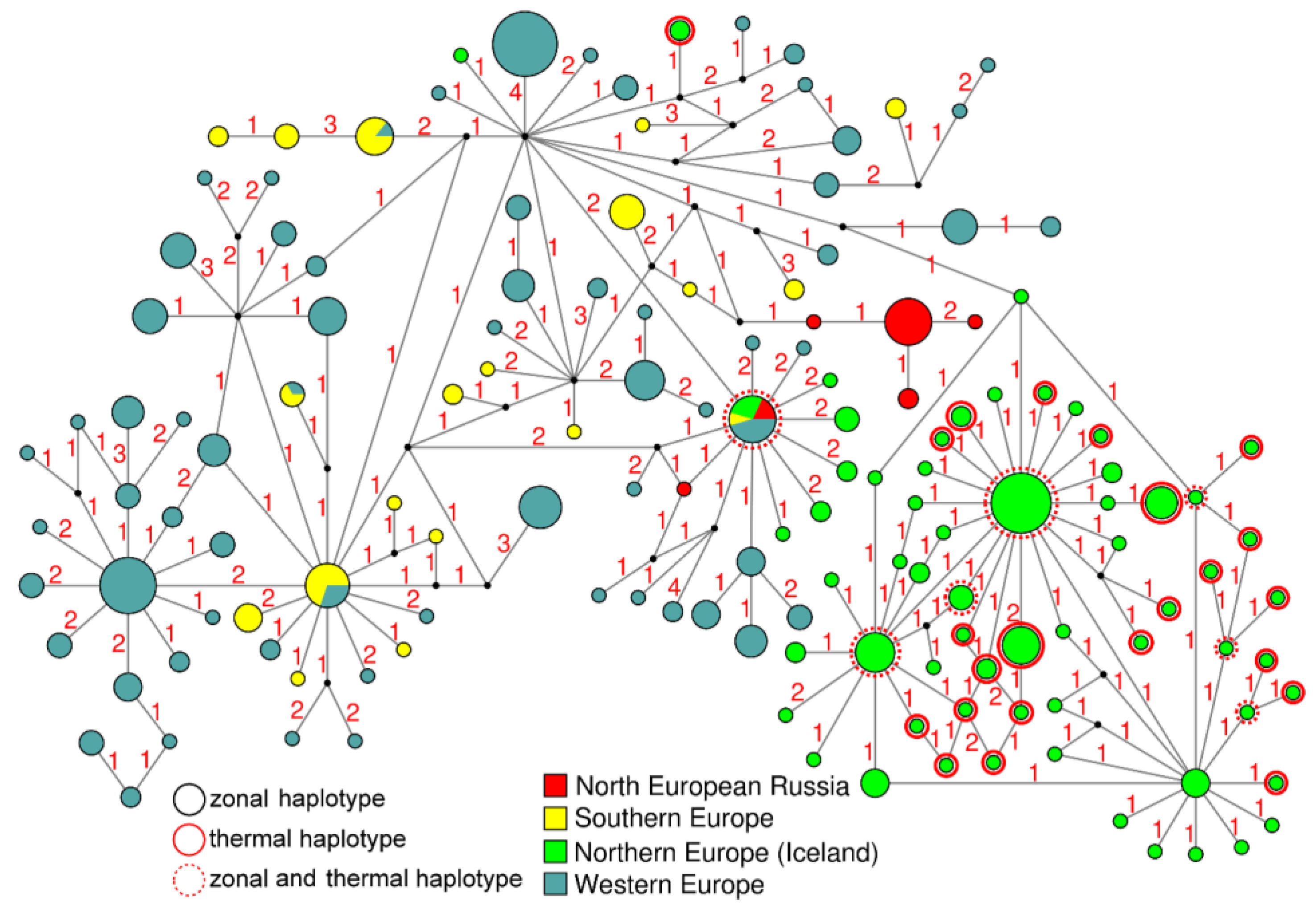
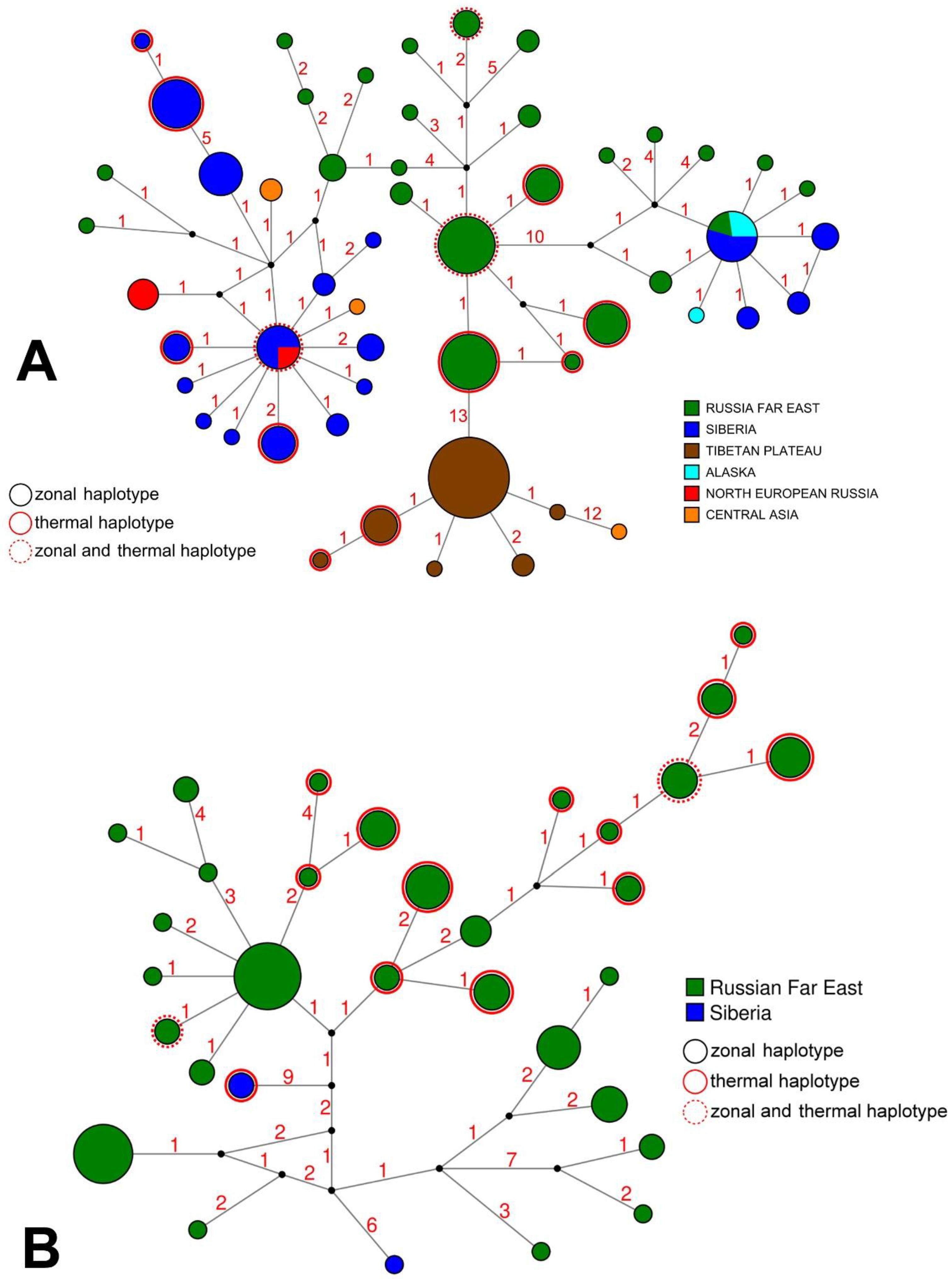
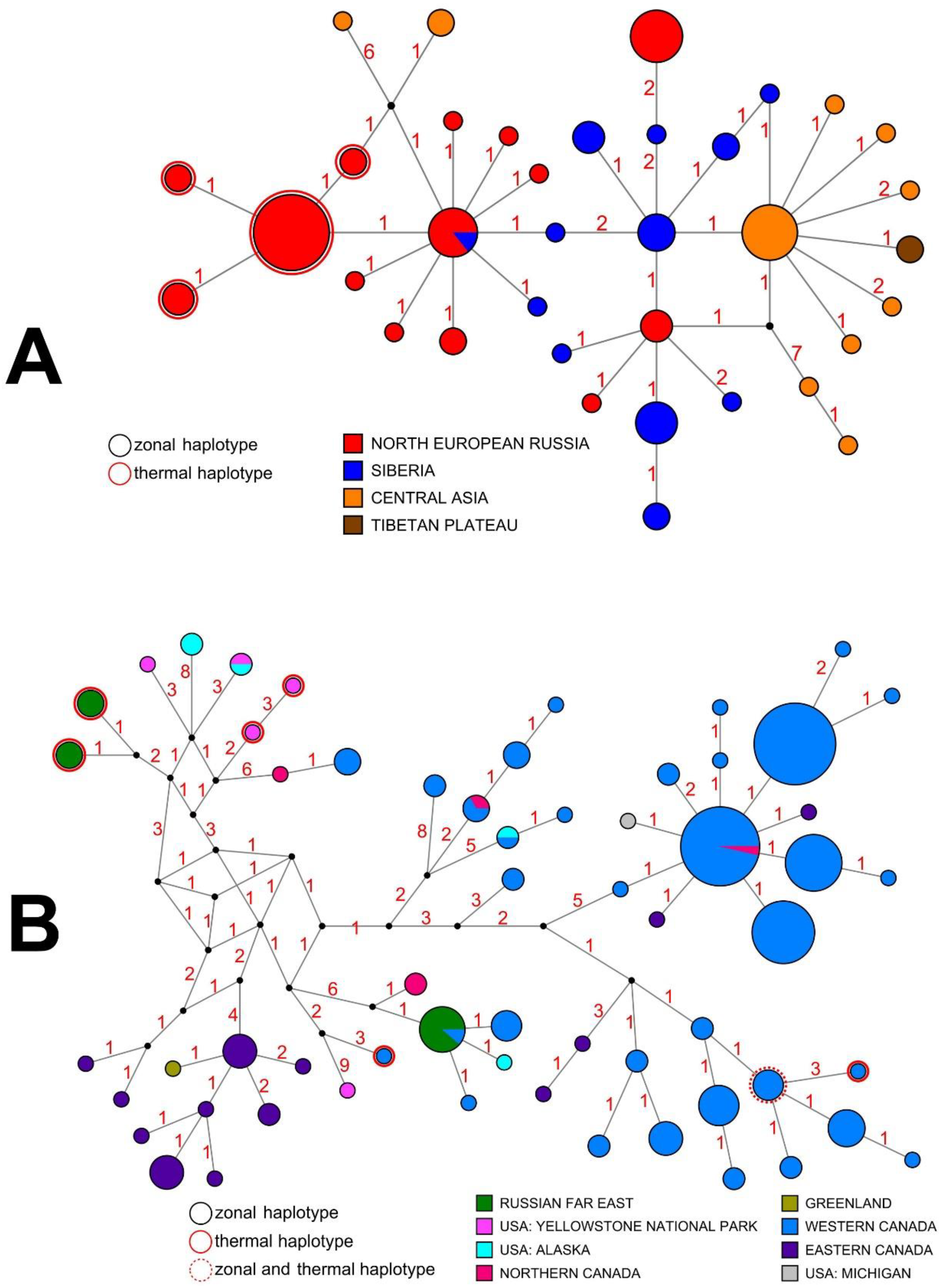
3.3. Intraspecific Genetic Diversity and Phylogeography
4. Discussion
4.1. Origin and Evolution of Thermal Lymnaeid Populations
4.2. The Probable Causes of Miniaturization in the Thermal Lymnaeids
- (a)
- A direct (modifying) influence of external conditions;
- (b)
- An outcome of some selective forces acting during the thermal acclimation;
- (c)
- Miniaturization as an inadaptive by-product of other adaptations that, probably, occur at physiological or biochemical levels.
4.3. ‘Ecological Race’ vs. ‘Incipient Speciation’ (a Conclusion)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Linnaeus, C. Systema Naturae per Regna Tria Naturae: Secundum Classes, Ordines, Genera, Species, Cum Characteribus, Differentiis, Synonymis, Locis, 12th ed.; 1. Regnum Animale. Pars 2.; Laurentius Salvius: Stockholm, Sweden, 1767; pp. 533–1327. [Google Scholar]
- Boubee, S.S.N. Nouveaux gisemens, en France, de Mollusques et Zoophytes. In Bulletin D’histoire Naturelle de France: Pour Servir à la Statistique, à la Géographie Naturelle et à L’étude Géologique de Cette Contrée; Food and Agriculture Organization of the United Nations: Roma, Italy, 1833; Volume 1, pp. 3–24. [Google Scholar]
- Dupuy, D. Histoire Naturelle des Mollusques Terrestres et d’eau Douce qui Vivent en France; Fascicule 5. V.; Masson: Paris, France, 1851; pp. 459–594. [Google Scholar]
- Remigio, E.A.; Lepitzki, D.A.W.; Lee, J.S.; Hebert, P.D.N. Molecular systematic relationships and evidence for a recent origin of the thermal spring endemic snails Physella johnsoni and Physella wrighti (Pulmonata: Physidae). Can. J. Zool. 2001, 79, 1941–1951. [Google Scholar] [CrossRef]
- Wethington, A.R.; Guralnick, R. Are populations of physids from different hot springs distinctive lineages? Am. Malacol. Bull. 2004, 19, 135–144. [Google Scholar]
- Bolotov, I.N.; Bespalaya, Y.V.; Usacheva, O.V. Ecology and evolution of hydrobionts in hot springs of the Subarctic and Arctic: Formation of similar assemblages, adaptation of species, and microevolutionary processes. Biol. Bull. Rev. 2012, 2, 340–348. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Aksenova, O.V.; Bespalaya, Y.V.; Gofarov, M.Y.; Kondakov, A.V.; Paltser, I.S.; Stefansson, A.; Travina, O.V.; Vinarski, M.V. Origin of a divergent mtDNA lineage of a freshwater snail species, Radix balthica, in Iceland: Cryptic glacial refugia or a postglacial founder event? Hydrobiologia 2017, 787, 73–98. [Google Scholar] [CrossRef]
- Johansson, M.P.; Ermold, F.; Kristjánsson, B.K.; Laurila, A. Divergence of gastropod life history in contrasting thermal environments in a geothermal lake. J. Evol. Biol. 2016, 29, 2043–2053. [Google Scholar] [CrossRef] [Green Version]
- von Oheimb, P.V.; Landler, L.; von Oheimb, C.M. Cold snails in hot springs: Observations from Patagonia and the Tibetan plateau. Malacologia 2016, 59, 313–320. [Google Scholar] [CrossRef]
- Takhteev, V.V.; Eropova, I.O.; Egorova, I.N.; Kobanova, G.I.; Krivenko, D.A.; Lishtva, A.V.; Ilin, G.D.; Pomazkova, G.I.; Okuneva, G.L.; Sitnikova, T.Y.; et al. Structure of hydrobiocenoses in mineral and thermal springs of the Lake Baikal region: A review. Contemp. Probl. Ecol. 2019, 12, 126–142. [Google Scholar] [CrossRef]
- Vinarski, M.V.; Bolotov, I.N.; Aksenova, O.V.; Babushkin, E.S.; Bespalaya, Y.V.; Makhrov, A.A.; Nekhaev, I.O.; Vikhrev, I.V. Freshwater Mollusca of the Circumpolar Arctic: A review on their taxonomy, diversity and biogeography. Hydrobiologia 2021, 848, 2891–2918. [Google Scholar] [CrossRef]
- Quenta-Herrera, E.; Daza, A.; Lazzaro, X.; Jacobsen, D.; Dangles, O.; Cauvy-Fraunié, S. Aquatic biota responses to temperature in a high Andean geothermal stream. Freshw. Biol. 2021, 66, 1889–1900. [Google Scholar] [CrossRef]
- Hubendick, B. Recent Lymnaeidae. Their Variation, Morphology, Taxonomy, Nomenclature and Distribution. In Kungliga Svenska Vetenskapsakademiens Handlingar; Fjärde Serien; Almqvist & Wiksell: Stockholm, Sweden, 1951; Volume 3, pp. 1–223. [Google Scholar]
- Meier-Brook, C. Taxonomic studies on Gyraulus (Gastropoda: Planorbidae). Malacologia 1983, 24, 1–113. [Google Scholar]
- Bolotov, I.; Bespalaya, Y.; Aksenova, O.; Aksenov, A.; Bolotov, N.; Gofarov, M.; Kondakov, A.; Paltser, I.; Vikhrev, I. A taxonomic revision of two local endemic Radix spp. (Gastropoda: Lymnaeidae) from Khodutka geothermal area, Kamchatka, Russian Far East. Zootaxa 2014, 3869, 585–593. [Google Scholar] [CrossRef] [Green Version]
- Vinarski, M.V.; Aksenova, O.V.; Bespalaya, Y.V.; Bolotov, I.N.; Gofarov, M.Y.; Kondakov, A.V. Ladislavella tumrokensis: The first molecular evidence of a Nearctic clade of lymnaeid snails inhabiting Eurasia. Syst. Biodiv. 2016, 14, 276–287. [Google Scholar] [CrossRef]
- Vinarski, M.V.; Aksenova, O.V.; Bolotov, I.N. Taxonomic assessment of genetically-delineated species of radicine snails (Mollusca, Gastropoda, Lymnaeidae). Zoosystematics Evol. 2020, 96, 577–608. [Google Scholar] [CrossRef]
- Aksenova, O.; Vinarski, M.; Bolotov, I.; Kondakov, A.; Bespalaya, Y.; Tomilova, A.; Paltser, I.; Gofarov, M. Two Radix spp. (Gastropoda: Lymnaeidae) endemic to thermal springs around Lake Baikal represent ecotypes of the widespread Radix auricularia. J. Zool. Syst. Evol. Res. 2017, 55, 298–309. [Google Scholar] [CrossRef]
- Aksenova, O.V.; Bolotov, I.N.; Gofarov, M.Y.; Kondakov, A.V.; Vinarski, M.V.; Bespalaya, Y.V.; Kolosova, Y.S.; Palatov, D.M.; Sokolova, S.E.; Spitsyn, V.M.; et al. Species richness, molecular taxonomy and biogeography of the radicine pond snails (Gastropoda: Lymnaeidae) in the Old World. Sci. Rep. 2018, 8, 11199. [Google Scholar] [CrossRef] [Green Version]
- Saito, T.; Chiba, S.; Fukuda, H. Type materials of the species of the Planorbidae (Mollusca, Gastropoda, Hygrophila) described by Shuichi Mori. Molluscan Res. 2020, 40, 169–182. [Google Scholar] [CrossRef]
- Sitnikova, T.Y.; Peretolchina, T.E. Description of a new species of Gyraulus (Pulmonata: Planorbidae) from the land thermal spring Khakusy of Lake Baikal. ZooKeys 2018, 762, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Borisov, S.N. Dragonflies (Odonata) of thermal springs in Central Asia. Entomol. Rev. 2015, 95, 1203–1211. [Google Scholar] [CrossRef]
- Aksenova, O.V.; Potapov, G.S.; Bespalaya, Y.V.; Kolosova, Y.S.; Vikhrev, I.V.; Kondakov, A.V.; Gofarov, M.Y.; Bolotov, I.N. Dragonflies from hot springs in Russia with a country-level checklist of species known to occur in geothermal environments. Ecol. Montenegrina 2020, 34, 49–63. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Aksenova, O.V.; Bespalaya, Y.V.; Spitsyn, V.M. Endemism of freshwater fish fauna in the geothermal regions: A review of the molecular and biogeographic studies. In Vestn. Sev. (Arkticheskogo) Fed. Univ. Seriya Estestv. Nauk; 2016; Volume 1, pp. 29–50. (In Russian) [Google Scholar]
- Novoselov, A.P.; Aksenova, O.V.; Bolotov, I.N.; Skyutte, N.G.; Anufriev, V.V.; Surso, M.V. Species composition, distribution and ecological features of ichthyofauna in the Pymvashor Geothermal Valley (Bolshezemelskaya tundra, Nenets Autonomous Okrug). J. Ichthyol. 2020, 60, 578–584. [Google Scholar] [CrossRef]
- Hofmann, S. Population genetic structure and geographic differentiation in the hot spring snake Thermophis baileyi (Serpentes, Colubridae): Indications for glacial refuges in southern-central Tibet. Mol. Phylogenetics Evol. 2012, 63, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Shain, D.H.; Iakovenko, N.S.; Cridge, A.G.; Novis, P.M.; Plášek, V.; Dearden, P.K. Microinvertebrate colonization of New Zealand’s thermally extreme environments. Evol. Biol. 2022, 49, 414–423. [Google Scholar] [CrossRef]
- Mitchell, R. The evolution of thermophyly in hot springs. Q. Rev. Biol. 1974, 49, 229–242. [Google Scholar] [CrossRef]
- Arendt, J.; Reznick, D. Convergence and parallelism reconsidered: What have we learned about the genetics of adaptation? Trends Ecol. Evol. 2008, 23, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Losos, J.B. Convergence, adaptation, and constraint. Evolution 2011, 65, 1827–1840. [Google Scholar] [CrossRef] [PubMed]
- Wake, D.B.; Wake, M.H.; Specht, C.D. Homoplasy: From detecting pattern to determining process. Science 2011, 331, 1032–1035. [Google Scholar] [CrossRef] [Green Version]
- Stayton, C.T. The definition, recognition, and interpretation of convergent evolution, and two new measures for quantifying and assessing the significance of convergence. Evolution 2015, 69, 2140–2153. [Google Scholar] [CrossRef]
- Agrawal, A.A. Toward a predictive framework for convergent evolution: Integrating natural history, genetic mechanisms, and consequences for the diversity of life. Am. Nat. 2017, 190, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, C.; Wilke, T.; Kuhn, K.; Streit, B. Convergent evolution of shell shape in freshwater limpets: The African genus Burnupia. Zool. J. Linn. Soc. 2004, 140, 577–588. [Google Scholar] [CrossRef]
- Adams, C.E.; Wilson, A.J.; Ferguson, M.M. Parallel divergence of sympatric genetic and body size forms of Arctic charr, Salvelinus alpinus, from two Scottish lakes. Biol. J. Linn. Soc. 2008, 95, 748–757. [Google Scholar] [CrossRef] [Green Version]
- Muschik, M.; Indermaur, A.; Salzburger, W. Convergent evolution within an adaptive radiation of cichlid fishes. Curr. Biol. 2012, 22, 2362–2368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vermeij, G. How convergent are Lake Tanganyika’s gastropods to marine ones? Comparative ecology and adaptive morphology. Biol. J. Linn. Soc. 2019, 127, 508–517. [Google Scholar] [CrossRef]
- Oke, K.B.; Rolshausen, G.; LeBlond, C.; Hendry, A.P. How parallel is parallel evolution? A comparative analysis in fishes. Am. Nat. 2017, 190, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Vinarski, M.V.; Clewing, C.; Albrecht, C. Family Lymnaeidae. In Freshwater Mollusks of the World: A Distribution Atlas; Lydeard, C., Cummings, K.S., Eds.; John Hopkins University Press: Baltimore, MD, USA, 2019; pp. 158–162. [Google Scholar]
- Hanken, J.; Wake, D.B. Miniaturization of body size: Organismal consequences and evolutionary significance. Ann. Rev. Ecol. Syst. 1993, 24, 501–519. [Google Scholar] [CrossRef]
- Polilov, A.A. Small is beautiful: Features of the smallest insects and limits to miniaturization. Annu. Rev. Entomol. 2015, 60, 103–121. [Google Scholar] [CrossRef]
- Moquin-Tandon, A. Histoire Naturelle des Mollusques Terrestres et Fluviatiles de France Contenant des Études Générales sur Leur Anatomie et Leur Physiologie et la Description Particulière des Genres, les Espèces et des Variétés; J.-B. Bailliére: Paris, France, 1856; Volume 2, pp. 177–646. [Google Scholar]
- Starmühlner, F. Zur Individuendichte und Formänderung von Lymnaea peregra Müller in isländischen Thermalbiotopen. Ergebnisse der Österreichischen Island-Expedition 1955. Sitz. Der Akad. Der Wiss. Wien Math.-Nat. Kl. (Abt. 1) 1957, 166, 331–384. [Google Scholar]
- Hazay, J. Die Molluskenfauna von Budapest. Malakozool. Blätter 1881, 3, 1–192. [Google Scholar]
- Clessin, S. Eine isländische Limnäe. Malakozool. Blätter 1878, 25, 79–80. [Google Scholar]
- Brues, C.T. Animal life in hot springs. Q. Rev. Biol. 1927, 2, 181–203. [Google Scholar] [CrossRef]
- Khmeleva, N.N.; Golubev, A.P.; Laenko, T.M. The ecology of gastropod mollusks from geothermal springs in Kamchatka. Zh Obsch. Biol. 1985, 46, 230–240. (In Russian) [Google Scholar]
- Aksenova, O.V.; Vinarski, M.V.; Bolotov, I.N.; Beslapaya, Y.V.; Kondakov, A.V.; Paltser, I.S. An overview of Radix species of the Kamchatka Peninsula (Gastropoda: Lymnaeidae). Bull. Russ. Far East. Malacol. Soc. 2016, 20, 5–27. [Google Scholar]
- Chatterjee, N.; Gupta, A.K.; Tiwari, S.K.; Clemens, S.C.; Sharma, K. Small size gastropod fauna from the Matli Geothermal Spring, Bhagirathi Valley, Garhwal Himalaya, Uttarakhand: Ecological implications. J. Geol. Soc. India 2022, 98, 47–52. [Google Scholar] [CrossRef]
- Fehér, Z.; Zettler, M.; Bozsó, M.; Szabó, K. An attempt to reveal the systematic relationship between Theodoxus prevostianus (C. Pfeiffer, 1828) and Theodoxus danubialis (C. Pfeiffer, 1828) (Mollusca, Gastropoda, Neritidae). Mollusca 2009, 27, 95–107. [Google Scholar]
- Warwick, T. Molluscs in thermal pools at Matlock Bath. J. Conchol. 1947, 22, 273. [Google Scholar]
- Falkner, G.; Bank, R.A.; von Proschwitz, T. Check-list of the non-marine molluscan species-group taxa of the states of Northern, Atlantic and Central Europe (CLECOM I). Heldia 2001, 4, 1–76. [Google Scholar]
- Brues, C.T. Studies on the fauna of some thermal springs in the Dutch East Indies. Proc. Am. Acad. Arts Sci. 1939, 73, 71–95. [Google Scholar] [CrossRef]
- Hafner, W.; Honsig-Erlemburg, W.; Mildner, P. Faunistischer Bericht über die Thermen in Warmbad Villach; Carinthia II (176./96. Jghrgan); Naturwissenschaftlicher Verein für Kärnten: Klagenfurt am Wörthersee, Austria, 1986; pp. 231–239. [Google Scholar]
- Domokos, T. New records of freshwater mollusks in the Bükk Mts. Abstr. Bot. 1992, 16, 129–138. [Google Scholar]
- Elkarmi, A.Z.; Ismail, N.S. Growth models and shell morphometrics of two populations of Melanoides tuberculata (Thiaridae) living in hot springs and freshwater pools. J. Limnol. 2007, 66, 90–96. [Google Scholar] [CrossRef] [Green Version]
- Sîrbu, I.; Benedek, A.-M. Native relict versus alien molluscs in thermal waters of North-Western Romania, including the first record of Melanoides tuberculata (O.F. Müller, 1774) (Gastropoda: Thiaridae) from Romania. Acta Zool. Bulg. 2017, 69, 31–36. [Google Scholar]
- Hotchkiss, E.R.; Hall, R.O., Jr. Linking exotic snails to carbon cycling in Kelly Warm Springs, Grand Teton National Park. University of Wyoming National Park Service Research Center. Annu. Rep. 2006, 30, 2. Available online: https://journals.uwyo.edu/index.php/uwnpsrc/article/view/3633 (accessed on 21 January 2023).
- Duggan, I.C.; Boothroyd, I.K.G.; Speirs, D.A. Factors affecting the distribution of stream macroinvertebrates in geothermal areas: Taupo Volcanic Zone, New Zealand. Hydrobiologia 2007, 592, 235–247. [Google Scholar] [CrossRef]
- Starobogatov, Y.I.; Izzatullaev, Z. Mollusks of the family Melanoididae (Gastropoda, Pectinibranchia) of Middle Asia and adjacent territories. Zool. Zhurnal 1980, 59, 23–31. (In Russian) [Google Scholar]
- Westerlund, C.A. Fauna der in der Paläarktischen Region (Europa, Kaukasien, Sibirien, Turan, Persien, Kurdistan, Armenien, Mesopotamien, Kleinasien, Syrien, Arabien, Egypten, Tripolis, Tunisien, Algerien und Marocco) Lebenden Binnenconchylien; VI. Fam; Ampullariidae, Paludinidae, Hydrobiidae, Melaniidae, Valvatidae & Neritidae; Ohlsson: Lund, Sweden, 1886; pp. 1–156. [Google Scholar]
- Pinter, I. A Hévízi-tó és a környező berek csigafaunája. Allatt. Közlemények 1979, 66, 117–130. [Google Scholar]
- Georgiev, D. In search of the thermal springs species Bithynia rumelica and Melanopsis parreyssi in Bulgaria. Tentacle 2010, 18, 16–17. [Google Scholar]
- Kobelt, W. Fauna der Nassauischen Mollusken; J. Niedner: Wiesbaden, Germany, 1871; pp. 1–286. [Google Scholar]
- Eropova, I.O.; Takhteev, V.V.; Sitnikova, T.Y.; Khadeeva, E.R.; Lopatovskaya, O.G. Macrozoobenthos of some thermal springs of Northern Baikal region in early springtime. Baikalskiy Zool. Zhurnal 2016, 2, 54–60. (In Russian) [Google Scholar]
- Takhteev, V.V.; Sitnikova, T.Y. The Gastropoda of thermomineral springs and accompanying waterbodies. In Biota Vodoemov Baikal’skoy Riftovoy Zony (The Biota of the Baikal Rift Zone Waterbodies); Pleshanov, A.S., Ed.; Irkutsk State University Press: Irkutsk, Russia, 2009; pp. 131–140. (In Russian) [Google Scholar]
- Starobogatov, Y.I. New species of gastropods from springs and subterranean waters of Middle Asia. Proc. Zool. Inst. Acad. Sci. USSR 1972, 50, 165–172. (In Russian) [Google Scholar]
- Brown, D.S. Freshwater Snails of Africa and Their Medical Importance; Taylor & Francis: London, UK, 1994; pp. 1–673. [Google Scholar]
- Kroll, O.; Hershler, R.; Albrecht, C.; Terrazas, E.M.; Apaza, R.; Fuentealba, C.; Wolff, C.; Wilke, T. The endemic gastropod fauna of Lake Titicaca: Correlation between molecular evolution and hydrographic history. Ecol. Evol. 2012, 2, 1517–1530. [Google Scholar] [CrossRef]
- Hershler, R. A systematic review of the hydrobiid snails (Gastropoda: Rissooidea) of the Great Basin, western United States. Part II. Genera Colligyrus, Eremopyrgus, Fluminicola, Pristinicola and Tryonia. Veliger 1999, 42, 306–337. [Google Scholar]
- Hershler, R.; Liu, H.P.; Landye, J.J. New species and records of springsnails (Caenogastropoda: Cochliopidae: Tryonia) from the Chihuahuan Desert (Mexico and United States), an imperiled biodiversity hotspot. Zootaxa 2011, 3001, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Clark, S.A.; Miller, A.C.; Ponder, W.F. Revision of the Snail Genus Austropyrgus (Gastropoda: Hydrobiidae): A Morphostatic Radiation of Freshwater Gastropods in Southeastern Australia; Records of the Australian Museum, Supplement; Australian Museum: Darlinghurst, Australia, 2003; Volume 28, pp. 1–109. [Google Scholar]
- Winterbourn, M.J. The faunas of thermal waters in New Zealand. Tuatara 1968, 16, 111–122. [Google Scholar]
- Bichain, J.M.; Gaubert, P.; Samadi, S.; Boisselier-Dubayle, M.C. A gleam in the dark: Phylogenetic species delimitation in the confusing spring-snail genus Bythinella Moquin-Tandon, 1856 (Gastropoda: Rissooidea: Amnicolidae). Mol. Phylogenetics Evol. 2007, 45, 927–941. [Google Scholar] [CrossRef] [PubMed]
- Glöer, P.; Varga, A.; Mrkvicka, A.C. Enigmatic Bythinella species in Bükk Mountains with the description of Bythinella thermophila n. sp. (Gastropoda: Amnicolidae). Ecol. Montenegrina 2015, 3, 40–45. Available online: https://www.biotaxa.org/em/article/view/em.2015.3.6/0 (accessed on 16 January 2023). [CrossRef]
- Brues, C.T. Further studies of the fauna of North American hot springs. Proc. Am. Acad. Arts Sci. 1932, 67, 185–303. [Google Scholar] [CrossRef]
- Haase, M. A revision of the genus Belgrandia, with the description of a new species from France (Caenogastropoda: Hydrobiidae). Malacologia 2000, 42, 171–201. [Google Scholar]
- Moog, O.; Reischütz, A.; Reischütz, P.L.; Winkler, G. Neue Fundorte von Thermal- und Quellschnecken in Bad Fischau-Brunn (Niederösterreich). Biodiversität Nat. Ostösterreich—BCBEA 2021, 6, 12–23. [Google Scholar]
- Glöer, P.; Bouzid, S.; Boeters, H.D. Revision of the genera Pseudamnicola Paulucci 1878 and Mercuria Boeters 1971 from Algeria with particular emphasis on museum collections (Gastropoda: Prosobranchia: Hydrobiidae). Arch. für Molluskenkd. 2010, 139, 1–22. [Google Scholar] [CrossRef]
- Mladenka, G.C.; Minshall, G.W. Variation in the life history and abundance of three populations of Bruneau hot springsnails (Pyrgulopsis bruneauensis). West. N. Am. Nat. 2001, 61, 204–212. [Google Scholar]
- Johnson, P.D.; Bogan, A.E.; Brown, K.M.; Burkhead, N.M.; Cordeiro, J.R.; Garner, J.T.; Hartfield, P.D.; Lepitzki, D.A.W.; Mackie, G.L.; Pip, E.; et al. Conservation status of freshwater gastropods of Canada and the United States. Fisheries 2013, 38, 247–282. [Google Scholar] [CrossRef]
- Bole, J. Neue Arten der unterirdischen schnecken Westbalkans. Slov. Akad. Znan. Umet. Razpr. 1992, 33, 3–20. [Google Scholar]
- De Mattia, W. A new subterranean hydrobiid from a thermal spring in eastern Slovenia: “Iglica” velkovrhi n. sp. (Gastropoda: Prosobranchia: Hydrobiidae). Mollusca 2007, 25, 27–31. [Google Scholar]
- Glöer, P.; Georgiev, D. New Rissooidea from Bulgaria (Gastropoda: Rissooidea). Mollusca 2009, 27, 123–136. [Google Scholar]
- Kuroda, T. Notes on the Stenothyridae (Aquatic Gastropoda) from Japan and adjacent regions. Venus. Jpn. J. Malacol. 1962, 22, 59–69. [Google Scholar]
- Neiber, M.T.; Cianfanelli, S.; Bartolini, F.; Glaubrecht, M. Not a marginal loss: Genetic diversity of the endangered freshwater snail Melanopsis etrusca (Brot, 1862) from thermal springs in Tuscany, Italy. Conserv. Genet. 2020, 21, 199–216. [Google Scholar] [CrossRef]
- Neubauer, T.A.; Harzhauser, M.; Georgopoulou, E.; Wrozyna, C. Population bottleneck triggering millennial-scale morphospace shifts in endemic thermal-spring melanopsids. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 414, 116–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bespalaya, Y.V.; Bolotov, I.N.; Usacheva, O.V. Mollusk population of a Subarctic hydrothermal system in winter. Zool. Zhurnal 2011, 90, 1311–1324. (In Russian) [Google Scholar]
- Izzatullaev, Z. Molluscs of the Aquatic Ecosystems of Middle Asia; Lessonpress: Tashkent, Uzbekistan, 2018; pp. 1–232. (In Russian) [Google Scholar]
- Brues, C.T. Studies on the fauna of hot springs in the Western United States and the biology of thermophilous animals. Proc. Am. Acad. Arts Sci. 1928, 63, 139–228. [Google Scholar] [CrossRef]
- Krkać, N. Temperature and reproductive cycle relations in Radix peregra O.F. Müller. Malacologia 1979, 18, 227–232. [Google Scholar]
- Fahy, E. Fauna and flora of a thermal spring at Innfield (Enfield). Ir. Nat. J. 1974, 18, 9–12. [Google Scholar]
- Weber, A. Über Binnenmollusken aus Ost-Turkestan, Kaschmir und West-Tibet. Zool. Jahrb. Abt. Syst. Okologie Geogr. Tiere 1910, 29, 299–312. [Google Scholar]
- Likharev, I.M.; Starobogatov, Y.I. On the molluscan fauna of Afghanistan. Proc. Zool. Inst. Acad. Sci. USSR 1967, 42, 159–197. (In Russian) [Google Scholar]
- Issel, A. Catalogo dei molluschi raccolti dalla missione italiana in Persia aggiuntavi la descrizione delle specie nuove o poco note. In Memorie della Reale Accademia delle Scienze di Torino; series 2; Stamperia Reale: Torino, Italy, 1865; Volume 23, pp. 387–439. [Google Scholar]
- Vinarski, M.V.; von Oheimb, P.V.; Aksenova, O.V.; Gofarov, M.Y.; Kondakov, A.V.; Nekhaev, I.O.; Bolotov, I.N. Trapped on the Roof of the World: Taxonomic diversity and evolutionary patterns of Tibetan Plateau endemic freshwater snails (Gastropoda: Lymnaeidae: Tibetoradix). Integr. Zool. 2022, 17, 825–848. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.W. Introduction to Physidae (Gastropoda: Hygrophila); biogeography, classification, morphology. Rev. Biol. Trop. 2003, 51 (Suppl. S1), 1–287. [Google Scholar] [PubMed]
- Yankovskaya, A.I. Fauna of warm springs of the East Pamir. Proc. Zool. Inst. Acad. Sci. USSR 1965, 35, 43–55. (In Russian) [Google Scholar]
- Biese, W.A. Revision de los moluscos terrestres y de agua dulce provistos de concha de Chile IV. Bol. Mus. Nac. Chile 1951, 25, 115–137. [Google Scholar]
- Prozorova, L.A.; Starobogatov, Y.I. New species of the subgenus Gyraulus of the genus Anisus and Gyraulus-similar forms from Asiatic Russia (with notes on classification of the genera Anisus and Choanomphalus) (Gastropoda Planorbidae). Ruthenica 1997, 7, 39–50. [Google Scholar]
- Mori, S. Classification of the Japanese Planorbidae. In Memoirs of the College of Science, Kyoto Imperial University, Series B (Biology); College of Science, Kyoto Imperial University: Kyoto, Japan, 1938; Volume 14, pp. 279–300. [Google Scholar]
- Klemm, W. Kreis: Mollusca. In Catalogus Faunae Austriae; Springer: Wien, Austria, 1960; Volume VIIa, pp. 1–60. [Google Scholar]
- Rohlf, F.J. tpsDig Version 2.11; State University of New York: Stony Brook, NY, USA, 2017. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 20 December 2022).
- Rohlf, F.J. tpsRelw Version 1.67; State University of New York: Stony Brook, NY, USA, 2017. [Google Scholar]
- Kruglov, N.D. Lymnaeid Snails of Europe and Northern Asia; Smolensk State Pedagogical University Press: Smolensk, Russia, 2005; pp. 1–508. (In Russian) [Google Scholar]
- Kruglov, N.D.; Starobogatov, Y.I. The volume of the subgenus Galba and of other similar subgenera of the genus Lymnaea (Gastropoda, Pulmonata). Zool. Zhurnal 1985, 64, 24–35. (In Russian) [Google Scholar]
- Kruglov, N.D.; Starobogatov, Y.I. Morphology and taxonomy of the molluscs from the subgenus Radix of the genus Lymnaea (Gastropoda, Pulmonata, Lymnaeidae) from Siberia and Far East of the USSR. Zool. Zhurnal 1989, 68, 17–30. (In Russian) [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7. 0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Middendorff, A.T. Die Thierwelt Sibiriens. Des eingeborenen Sibiriens. In Reise in Den Äussersten Norden und Osten Sibiriens. Band IV. Übersicht Natur Nord- und Ost-Sibiriens; Middendorff, A.T., Ed.; Kaiserlische Akademie der Wissenschaften: Saint-Pétersburg, Russia, 1867; pp. 785–1094. [Google Scholar]
- Økland, J. Lakes and Snails: Environment and Gastropoda in 1500 Norwegian Lakes, Ponds and Rivers; Universal Book Services/W; Backhuys: Oegstgeest, The Netherlands, 1990; pp. 1–516. [Google Scholar]
- Nekhaev, I.O. Surviving at the edge of land: Finding of the limnetic snail Sibirenauta sibirica (Gastropoda: Physidae) on the coast of the Laptev Sea (Eastern Siberia). Bull. Russ. Far East Malacol. Soc. 2015, 19, 25–30. [Google Scholar]
- Vinarski, M.V.; Palatov, D.M.; Novichkova, A.A. The first freshwater mollusks from Wrangel Island, Arctic Russia. Polar Res. 2015, 34, 23889. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.M.; Buckley, L.B.; Sheldon, K.S.; Vickers, M.; Pörtner, H.O.; Dowd, W.W.; Gunderson, A.R.; Marshall, K.E.; Stillman, J.H. Biological impacts of thermal extremes: Mechanisms and costs of functional responses matter. Integr. Comp. Biol. 2016, 56, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Hochachka, P.W.; Somero, G.N. Biochemical Adaptation: Mechanism and Process in Physiological Evolution; Oxford University Press: New York, NY, USA, 2002; pp. 1–466. [Google Scholar]
- Janzen, D.H. Why mountain passes are higher in the tropics. Am. Nat. 1967, 101, 233–249. [Google Scholar] [CrossRef]
- Deutsch, C.A.; Tewksbury, J.J.; Huey, R.B.; Sheldon, K.J.; Ghalambor, C.K.; Haak, D.C.; Martin, P.R. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl. Acad. Sci. USA 2008, 105, 6668–6672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, N.L.; Smith, J.A. An alternative explanation for global trends in thermal tolerance. Ecol. Lett. 2017, 20, 70–77. [Google Scholar] [CrossRef] [Green Version]
- Schmalhausen, I.I. The problem of stability of organic forms (onthogeneses) in their historical development. Zh. Obsch. Biol. 1945, 6, 3–25. (In Russian) [Google Scholar]
- Makhrov, A.A. Decreased evolutionary plasticity as a result of phylogenetic immobilization and its ecological significance. Contemp. Probl. Ecol. 2019, 12, 405–417. [Google Scholar] [CrossRef]
- Bohlen, J.; Freyhof, J.; Nolte, A. Sex ratio and body size in Cobitis elongatoides and Sabanejewia balcanica (Cypriniformes, Cobitidae) from a thermal spring. Folia Zool. 2008, 57, 191–197. [Google Scholar]
- Esin, E.V.; Sorokin, Y.V. Residential form of white-spotted char Salvelinus leucomaenis inhabiting the warm stream discharging into Semlyachikskii Firth (Kronotskii natural reserve, Kamchatka). J. Ichthyol. 2012, 52, 172–179. [Google Scholar] [CrossRef]
- Lema, S.C.; Bock, S.L.; Malley, M.M.; Elkins, E.A. Warming waters beget smaller fish: Evidence for reduced size and altered morphology in a desert fish following anthropogenic temperature change. Biol. Lett. 2019, 15, 20190518. [Google Scholar] [CrossRef]
- Schmalhausen, I.I. Faktory Evolyutsii [The Factors of Evolution]; Nauka: Moscow, Russia, 1968; pp. 1–452. [Google Scholar]
- Pakhnevich, A.V. Reasons of micromorphism in modern or fossil brachiopods. Paleontol. J. 2009, 43, 1458–1468. [Google Scholar] [CrossRef]
- MacColl, A.D.C.; Nagar, A.E.; de Roij, J. The evolutionary ecology of dwarfism in three-spined sticklebacks. J. Anim. Ecol. 2013, 82, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Passow, C.N.; Greenway, R.; Arias-Rodriquez, L.; Jeyasingh, P.D.; Tobler, M. Reduction of energetic demands through modification of body size and routine metabolic rates in extremophile fish. Physiol. Biochem. Zool. 2015, 88, 371–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passow, C.N.; Arias-Rodriguez, L.; Tobler, M. Convergent evolution of reduced energy demands in extremophile fish. PLoS ONE 2017, 12, e0186935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGrath, C. Big surprises from the World’s smallest fish. Genome Biol. Evol. 2018, 10, 1104–1105. [Google Scholar] [CrossRef] [Green Version]
- Esin, E.V.; Markevich, G.N.; Shkil, F.N. Rapid miniaturization of Salvelinus fish as an adaptation to the volcanic impact. Hydrobiologia 2020, 847, 2947–2962. [Google Scholar] [CrossRef]
- Niu, J.; Huss, M.; Vasemägi, A.; Gårdmark, A. Decades of warming alters maturation and reproductive investment in fish. Ecosphere 2023, 14, e4381. [Google Scholar] [CrossRef]
- Geyer, D. Zwergschnecken. Arch. Molluskenkd. 1923, 55, 231–235. [Google Scholar]
- Geyer, D. Die Schlammschnecke Limnaea und ihre Anpassungsformen im Bodensee. Natur und Museum. 1929, 59, 336–353. [Google Scholar]
- Walter, H.J. Illustrated biomorphology of the “angulata” lake form of the basommatophoran snail Lymnaea catascopium Say. Malacol. Rev. 1969, 2, 1–102. [Google Scholar]
- Hu, J.; Barrett, R.D.H. Epigenetics in natural animal populations. J. Evol. Biol. 2017, 30, 1612–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pimpinelli, S.; Piacentini, L. Environmental change and the evolution of genomes: Transposable elements as translators of phenotypic plasticity into genotypic variability. Funct. Ecol. 2020, 34, 428–441. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, D. Temperature and organism size: A biological law for ectotherms. Adv. Ecol. Res. 1994, 25, 1–58. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Huey, R.B. Size, temperature, and fitness: Three rules. Evol. Ecol. Res. 2008, 10, 251–268. [Google Scholar]
- Lindmark, M.; Ohlberger, J.; Gårdmark, A. Optimum growth temperature declines with body size within fish species. Glob. Chang. Biol. 2022, 28, 2259–2271. [Google Scholar] [CrossRef] [PubMed]
- Hoefnagel, K.N.; Verberk, W.C.E.P. Is the temperature-size rule mediated by oxygen in aquatic ectotherms? J. Therm. Biol. 2015, 54, 56–65. [Google Scholar] [CrossRef]
- Audzijonyte, A.; Barneche, D.R.; Baudron, A.R.; Belmaker, J.; Clark, T.D.; Marshall, C.T.; Morrongiello, J.R.; van Rijn, I. Is oxygen limitation in warming waters a valid mechanism to explain decreased body sizes in aquatic ectotherms? Glob. Ecol. Biogeogr. 2018, 28, 2259–2271. [Google Scholar] [CrossRef] [Green Version]
- Rollinson, N.; Rowe, L. Temperature-dependent oxygen limitation and the rise of Bergmann’s rule in species with aquatic respiration. Evolution 2018, 72, 977–988. [Google Scholar] [CrossRef]
- Boettger, C.R. Beiträge zur Kenntnis der Molluskenfauna Schlesiens. Nachr. bl. dtsch. Malak.-Ges. 1913, 45, 153–163. [Google Scholar]
- Backhuys, W. Land & Fresh-Water Molluscs of the Azores; Backhuys & Meesters: Amsterdam, The Netherlands, 1975; pp. 1–350. [Google Scholar]
- Van Bocxlaer, B.; Ortiz-Sepulveda, C.M.; Gurdebeke, P.R.; Vekemans, X. Adaptive divergence in shell morphology in an ongoing gastropod radiation from Lake Malawi. BMC Evol. Biol. 2020, 20, 5. [Google Scholar] [CrossRef]
- Di Santo, V.; Lobel, P.S. Body size and thermal tolerance in tropical gobies. J. Exp. Mar. Biol. Ecol. 2017, 487, 11–17. [Google Scholar] [CrossRef]
- Siepielski, A.M.; Morrissey, M.B.; Carlson, S.M.; Francis, C.D.; Kingsolver, J.G.; Whitney, K.D.; Kruuk, L.E.B. No evidence that warmer temperatures are associated with selection for smaller body sizes. Proc. R. Soc. B 2019, 286, 20191332. [Google Scholar] [CrossRef]
- Vermeij, G. Gastropod shell growth rate, allometry, and adult size: Environmental implications. In Skeletal Growth of Aquatic Organisms: Biological Records of Environmental Change; Rhoads, D.C., Lutz, K.A., Eds.; Plenum Press: New York, NY, USA, 1980; pp. 379–394. [Google Scholar]
- Calow, P. On the regulatory nature of individual growth: Some observations from freshwater snails. J. Zool. 1972, 170, 415–428. [Google Scholar] [CrossRef]
- Calow, P. Adaptational aspects of growth and reproduction in Lymnaea peregra (Gastropoda: Pulmonata) from exposed and sheltered aquatic habitats. Malacologia 1981, 21, 5–13. [Google Scholar]
- Brown, K.M. Intraspecific life history variation in a pond snail: The roles of population divergence and phenotypic plasticity. Evolution 1985, 39, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, D.; Sibly, R.M. Why are organisms usually bigger in colder environments? Making sense of a life history puzzle. Trends Ecol. Evol. 1997, 12, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.M. Do life history tactics exist at the intra-specific level? Data from freshwater snails. Am. Nat. 1983, 121, 871–879. [Google Scholar] [CrossRef]
- Lam, P.K.S.; Calow, P. Intraspecific life-history variation in Lymnaea peregra (Gastropoda: Pulmonata). I. Field study. J. Anim. Ecol. 1989, 58, 571–588. [Google Scholar] [CrossRef]
- Aziz, M.D.A.; Raut, S.K. Thermal effect on the life-cycle parameters of the medically important freshwater snail species Lymnaea (Radix) luteola (Lamarck). Memórias Do Inst. Oswaldo Cruz 1996, 97, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Carreira, B.M.; Segurado, P.; Laurila, A.; Rebelo, R. Heat waves trigger swift changes in the diet and life-history of a freshwater snail. Hydrobiologia 2020, 847, 999–1011. [Google Scholar] [CrossRef]
- Nätscher, P.S.; Dera, G.; Reddin, C.J.; Rita, P.; De Baets, K. Morphological response accompanying size reduction of belemnites during an Early Jurassic hyperthermal event modulated by life history. Sci. Rep. 2021, 11, 14480. [Google Scholar] [CrossRef] [PubMed]
- Walczynska, A.; Franch-Gras, L.; Serra, M. Empirical evidence for fast temperature-dependent body size evolution in rotifers. Hydrobiologia 2017, 796, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Arakelova, E.S. Effect of temperature on the duration of embryonic development in freshwater pulmonates. Zh. Obsch. Biol. 1986, 47, 128–135. (In Russian) [Google Scholar]
- Gustafson, K.D.; Kensinger, B.J.; Bolek, M.G.; Luttbeg, B. Distinct snail (Physa) morphotypes from different habitats converge in shell shape and size under common garden conditions. Evol. Ecol. Res. 2014, 16, 77–89. [Google Scholar]
- McNamara, K.J. A guide to the nomenclature of heterochrony. J. Paleontol. 1986, 60, 4–13. [Google Scholar] [CrossRef]
- Schmalhausen, I.I. Problemy Darvinizma [The Problems of Darwinism]; Nauka: Moscow, Russia, 1969; pp. 1–493. (In Russian) [Google Scholar]
- Gould, S.J. Ontogeny and Phylogeny; The Belknap Press of Harvard University Press: Cambridge, MA, USA; London, UK, 1977; pp. 1–501. [Google Scholar]
- Vermeij, G.J. Evolution and Escalation: An Ecological History of Life; Princeton University Press: Princeton, NJ, USA, 1987; pp. 1–527. [Google Scholar]
- Geary, D. Heterochrony in gastropods: A paleontological view. In Heterochrony in Evolution; McKinney, M.L., Ed.; Plenum Press: New York, NY, USA, 1988; pp. 183–196. [Google Scholar]
- Lindberg, D. Heterochrony in gastropods: A neontological view. In Heterochrony in Evolution; McKinney, M.L., Ed.; Plenum Press: New York, NY, USA, 1988; pp. 197–216. [Google Scholar]
- Takhteev, V.V.; Sudakova, E.A.; Egorova, I.N.; Shamanova, S.I.; Sitnikova, T.Y.; Kravtsova, L.S.; Rozhkova, N.A.; Pomazkova, G.I.; Arbuzov, A.V.; Ambrosova, E.V.; et al. Characteristic of aquatic and terrestrial biocenoses in the abruption places of thermal springs in Eastern Cis-Baikal region. In Gidrobiologiya Vodoemov Yuga Vostochnoi Sibiri (Hydrobiology of Southern Reservoirs of Eastern Siberia); Irkutsk State University Press: Irkutsk, Russia, 2006; pp. 111–136. (In Russian) [Google Scholar]
- Glöer, P. Die Süßwassergastropoden Nord- und Mitteleuropas: Bestimmungsschlüssel, Lebensweise, Verbreitung; Conchbooks: Hackenheim, Germany, 2002; pp. 1–327. [Google Scholar]
- Aksenova, O.V.; Vinarski, M.V.; Itagaki, T.; Ohari, Y.; Oshida, Y.; Kim, S.K.; Lee, J.H.; Kondakov, A.V.; Khrebtova, I.S.; Tomilova, A.A.; et al. Taxonomy and trans-Beringian biogeography of the pond snails (Gastropoda: Lymnaeidae) of East Asia: An integrative view. Sci. Rep. 2023. submitted. [Google Scholar]
- Buckley, L.B.; Huey, R.B. How extreme temperatures impact organisms and the evolution of their thermal tolerance. Integr. Comp. Biol. 2016, 56, 98–109. [Google Scholar] [CrossRef] [Green Version]
- Ladd, H.L.; Rogowski, D.L. Egg predation and parasite prevalence in the invasive freshwater snail, Melanoides tuberculata (Müller, 1774) in a west Texas spring system. Aquat. Invasions 2012, 7, 287–290. [Google Scholar] [CrossRef]
- Mehler, K.; Acharya, K. Size distribution and nutrient excretion of Melanoides tuberculata in a southern Nevada spring ecosystem. West. N. Am. Nat. 2014, 74, 386–395. [Google Scholar] [CrossRef]












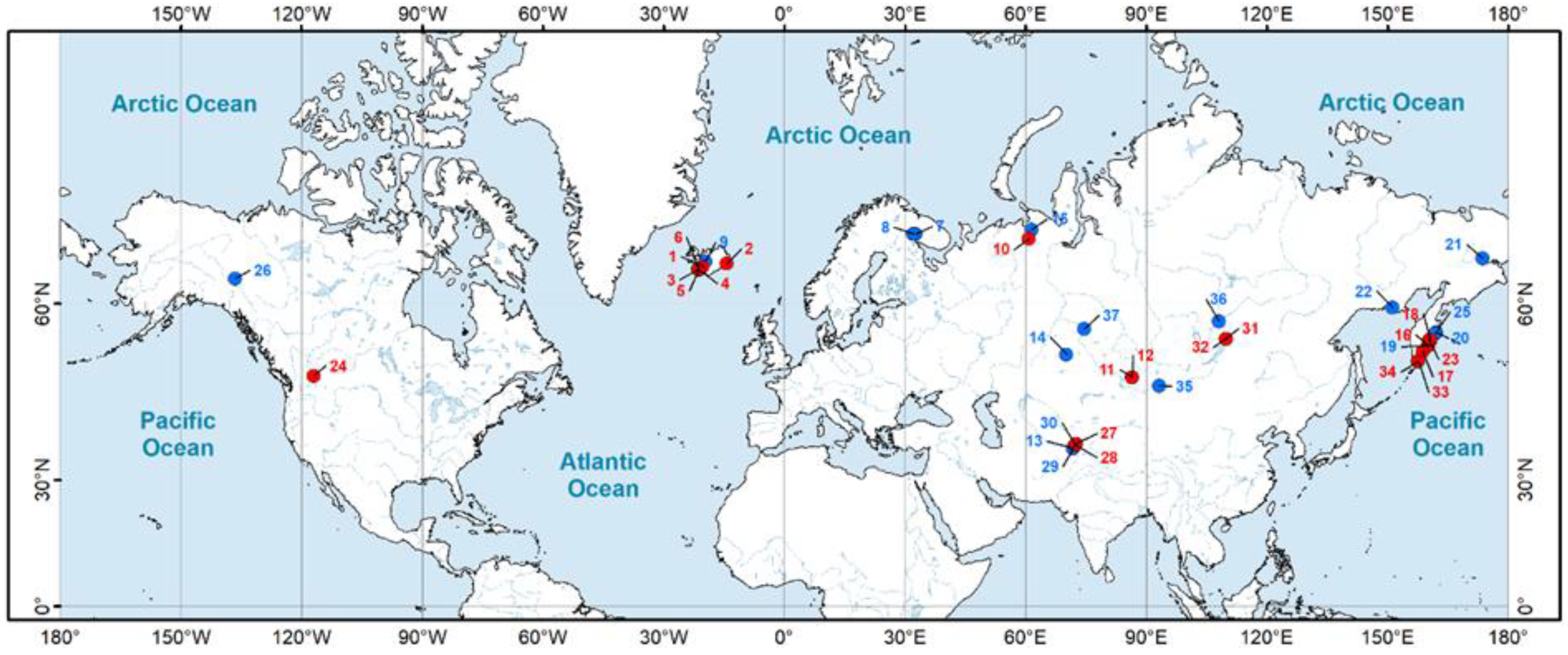
| No. | Species | Sampling Locality, Sampling Date (Month, Year) | Type of Habitat | Latitude | Longitude | N/n * | Museum Accession Number |
|---|---|---|---|---|---|---|---|
| 1 | Ampullaceana balthica | Iceland, Geysir Hot Spring area, a thermal pool, July 2013 | GT | 64.3144 | −20.2991 | 80/33 | RMBH, MLym-195–MLym 198 |
| 2 | A. balthica | Iceland, hot spring in Djúpivogur, July, 2015 | GT | 64.6568 | −14.2847 | 26/26 | LMBI (not catalogized) |
| 3 | A. balthica | Iceland, thermal springs near Hengill volcano, July 2013 | GT | 64.0538 | −21.2894 | 32/32 | RMBH, Mlym-190 |
| 4 | A. balthica | Iceland, a hot brook near Rejkamork settlement, July 2013 | GT | 64.0058 | −21.1811 | 105/32 | RMBH, MLym-184 |
| 5 | A. balthica | Iceland, hot springs of the Graendalsa River valley, near Rejkakot settlement, July 2013 | GT | 64.0268 | −21.1988 | 153/30 | RMBH, MLym-183 |
| 6 | A. balthica | Iceland, a hot brook of the Graendalsa River valley, July 2013 | GT | 64.0241 | −21.2036 | 36/30 | RMBH, MLym-189 |
| 7 | A. balthica | Russia, Kola Peninsula, Chunozero Lake, August 2012 | NG | 67.6367 | 32.6330 | 32/32 | LMBI (not catalogized) |
| 8 | A. balthica | Russia, Kola Peninsula, Pirenga Lake, August 2012 | NG | 67.6276 | 32.1206 | 32/32 | LMBI (not catalogized) |
| 9 | A. balthica | Iceland, a brook near Hveravellir, July 2015 | NG | 64.8675 | −19.5456 | 32/32 | LMBI (not catalogized) |
| 10 | Ampullaceana lagotis | Russia, Nenets Autonomous Region, Pymvashor hot springs, July–August 2009; December 2009; August – September 2010; December 2011 | GT | 67.1893 | 60.8617 | 336/36 | RMBH (not catalogized) |
| 11 | A. lagotis | Kazakhstan, East Kazakhstan Region, a thermal pool on the shore of Lake Rakhmanovskoye, July 2018 | GT | 49.5338 | 86.5165 | 33/33 | LMBI, 15-3765 |
| 12 | A. lagotis | Kazakhstan, East Kazakhstan Region, a warm brook on the shore of Lake Rakhmanovskoye, July 2018 | GT | 49.5309 | 86.5157 | 22/22 | LMBI, 15-3721 |
| 13 | A. lagotis | Tajikistan, a wetland in vicinity of Zumagd village, July 2016 | NG | 36.6853 | 71.8318 | 19/19 | LMBI, 15-3303 |
| 14 | A. lagotis | Kazakhstan, Kustanay Region, Lake Maibalyk, July 2016 | NG | 53.1138 | 70.1808 | 22/22 | LMBI, 15-895 |
| 15 | A. lagotis | Russia, Nenets Autonomous Region, Vashutkiny Lakes, July 2009 | NG | 68.0201 | 61.5391 | 26/26 | RMBH (not catalogized) |
| 16 | Kamtschaticana kamtschatica | Russia, Kamchatka, Teremkovaya hot area, July 2014 | GT | 54.4341 | 160.1380 | 30/30 | LMBI, 15-3205 |
| 17 | K. kamtschatica | Russia, Kamchatka, Nalychevskiye hot springs, July 2014 | GT | 53.5055 | 158.7633 | 24/24 | LMBI, 15-2970 |
| 18 | K. kamtschatica | Russia, Kamchatka, Storozhevskiye hot springs, August 1988 | GT | 55.1880 | 160.6580 | 29/29 | LMBI, 15-3222 |
| 19 | K. kamtschatica | Russia, Kamchatka, Troinoye Lake, July 2014 | NG | 54.4313 | 160.1297 | 19/19 | LMBI, 15-3215 |
| 20 | K. kamtschatica | Russia, Kamchatka, Krasikovskoye Lake, August 2019 | NG | 56.2575 | 162.0429 | 23/23 | RMBH, MLym-894 |
| 21 | K. kamtschatica | Russia, Chukchi Peninsula, Utesinskoye Lake, August 2019 | NG | 65.2132 | 173.6357 | 35/35 | RMBH, MLym-876 |
| 22 | K. kamtschatica | Russia, Magadan Region, a lake near Magadan City, September 2019 | NG | 59.5726 | 151.2531 | 9/9 | LMBI, 15-3779 |
| 23 | Ladislavella catascopium | Russia, Kamchatka, Tumrok hot springs (paratypes of Lymnaea tumrokensis Kruglov et Starobogatov, 1985) **, 1963 | GT | 55.2049 | 160.3991 | 38/38 | ZIN, No. 2 |
| 24 | L. catascopium | Canada, British Columbia, Ainsworth hot springs, July 2017 | GT | 49.7356 | −116.9105 | 30/30 | RMBH, MLym-815 |
| 25 | L. catascopium | Russia, Kamchatka, Lake Azabachye, August 2019 | NG | 56.1140 | 161.8204 | 30/30 | RMBH, MLym-896 |
| 26 | L. catascopium | Canada, Yukon Territory, unnamed lake, July 2017 | NG | 63.0196 | −136.4683 | 22/22 | RMBH, MLym-660 |
| 27 | Radix alticola | Tajikistan, hot spring in Djelandy village, July 2016 | GT | 37.5773 | 72.5764 | 60/34 | LMBI, 15-3298, 15-3299 |
| 28 | R. alticola | Tajikistan, hot spring near Djavshangoz village, July 2016 | GT | 37.3588 | 72.4168 | 34/34 | LMBI, 15-3300 |
| 29 | R. alticola | Tajikistan, a warm brook inflowing into Shakhdara River (middle course), July 2016 | NG | 37.3588 | 72.4168 | 44/32 | LMBI, 15-3349 |
| 30 | R. alticola | Tajikistan, a warm brook inflowing into Shakhdara River (mouth), July 2016 | NG | 37.3588 | 72.4168 | 22/22 | LMBI, 15-3326, 15-3569 |
| 31 | Radix auricularia | Russia, Republic of Buryatia, Khakusy hot springs (paratypes of Lymnaea khakusyensis Kruglov et Starobogatov, 1989) ***, September 1976 | GT | 55.3595 | 109.8274 | 8/8 | ZIN, Nos 2, 3 |
| 32 | R. auricularia | Russia, Republic of Buryatia, Khakusy hot springs (paratypes of Lymnaea thermobaicalica Kruglov et Starobogatov, 1989) ***, September 1976 | GT | 55.3595 | 109.8274 | 42/42 | ZIN, Nos 2, 4 |
| 33 | R. auricularia | Russia, Kamchatka, Khodutka hot springs (paratypes of Lymnaea hodutkae Kruglov et Starobogatov, 1989) ***, September 1979 | GT | 52.1073 | 157.6573 | 30/30 | ZIN, Nos 2, 3 |
| 34 | R. auricularia | Russia, Kamchatka, Khodutka hot springs (paratypes of Lymnaea thermokamchatica Kruglov et Starobogatov, 1989) ***,September 1979 | GT | 52.1073 | 157.6573 | 43/43 | ZIN, Nos 2, 3 |
| 35 | R. auricularia | Mongolia, Khar-Nuur Lake, July 2012 | NG | 48.1333 | 93.2500 | 32/32 | LMBI, 15-2481 |
| 36 | R. auricularia | Russia, Irkutsk Region, Kirenga River in Kirensk Town, July 2003 | NG | 57.7757 | 108.1108 | 21/21 | LMBI, 15-1335 |
| 37 | R. auricularia | Russia, Omsk Region, Krivoye Lake, August 2002 | NG | 56.7720 | 74.6133 | 35/35 | LMBI, 15-412 |
| Species | Mean SH, mm (±σ) | Statistical Significance of the Difference (Z Adjusted; p-Value) | |
|---|---|---|---|
| Thermal Populations | Non-Thermal Populations | ||
| Ampullaceana balthica, all samples A. balthica, Iceland populations only | 4.0 ± 1.6 4.0 ± 1.6 | 14.3 ± 2.3 8.9 ± 3.2 | 11.80; 0.0011.28; 0.00 |
| A. lagotis | 7.7 ± 1.6 | 11.0 ± 2.6 | –7.84; 0.00 |
| Kamtschaticana kamtschatica | 6.6 ± 0.9 | 8.3 ± 1.2 | –9.54; 0.00 |
| Ladislavella catascopium | 9.7 ± 1.8 | 17.1 ± 2.5 | 8.29; 0.00 |
| Radix alticola | 8.5 ± 0.6 | 11.9 ± 1.3 | –10.51; 0.00 |
| R. auricularia | 9.1 ± 2.0 | 21.6 ± 3.2 | –12.5; 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vinarski, M.V.; Aksenova, O.V.; Bespalaya, Y.V.; Gofarov, M.Y.; Kondakov, A.V.; Khrebtova, I.S.; Makhrov, A.A.; Bolotov, I.N. How an Ecological Race Is Forming: Morphological and Genetic Disparity among Thermal and Non-Thermal Populations of Aquatic Lymnaeid Snails (Gastropoda: Lymnaeidae). Diversity 2023, 15, 548. https://doi.org/10.3390/d15040548
Vinarski MV, Aksenova OV, Bespalaya YV, Gofarov MY, Kondakov AV, Khrebtova IS, Makhrov AA, Bolotov IN. How an Ecological Race Is Forming: Morphological and Genetic Disparity among Thermal and Non-Thermal Populations of Aquatic Lymnaeid Snails (Gastropoda: Lymnaeidae). Diversity. 2023; 15(4):548. https://doi.org/10.3390/d15040548
Chicago/Turabian StyleVinarski, Maxim V., Olga V. Aksenova, Yulia V. Bespalaya, Mikhail Yu. Gofarov, Alexander V. Kondakov, Irina S. Khrebtova, Alexander A. Makhrov, and Ivan N. Bolotov. 2023. "How an Ecological Race Is Forming: Morphological and Genetic Disparity among Thermal and Non-Thermal Populations of Aquatic Lymnaeid Snails (Gastropoda: Lymnaeidae)" Diversity 15, no. 4: 548. https://doi.org/10.3390/d15040548