
Bioenergetics of Euphausia superba and Euphausia crystallorophias in the Ross Sea




Abstract
:1. Introduction
2. Materials and Methods
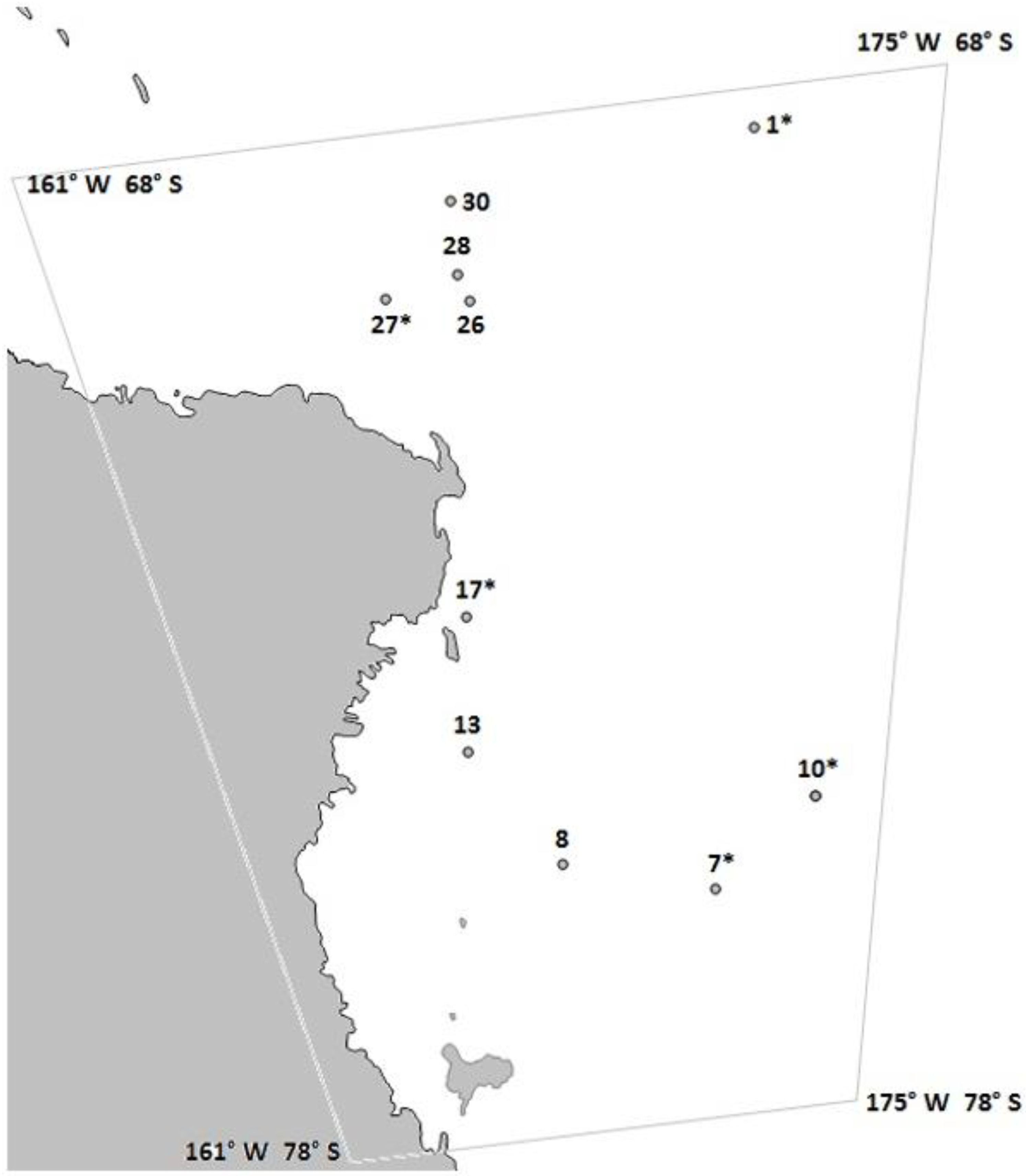
2.1. Study Area and Sampling
2.2. Biometric Measurements
2.3. Biochemical Analyses
2.4. Statistical Analysis
3. Results
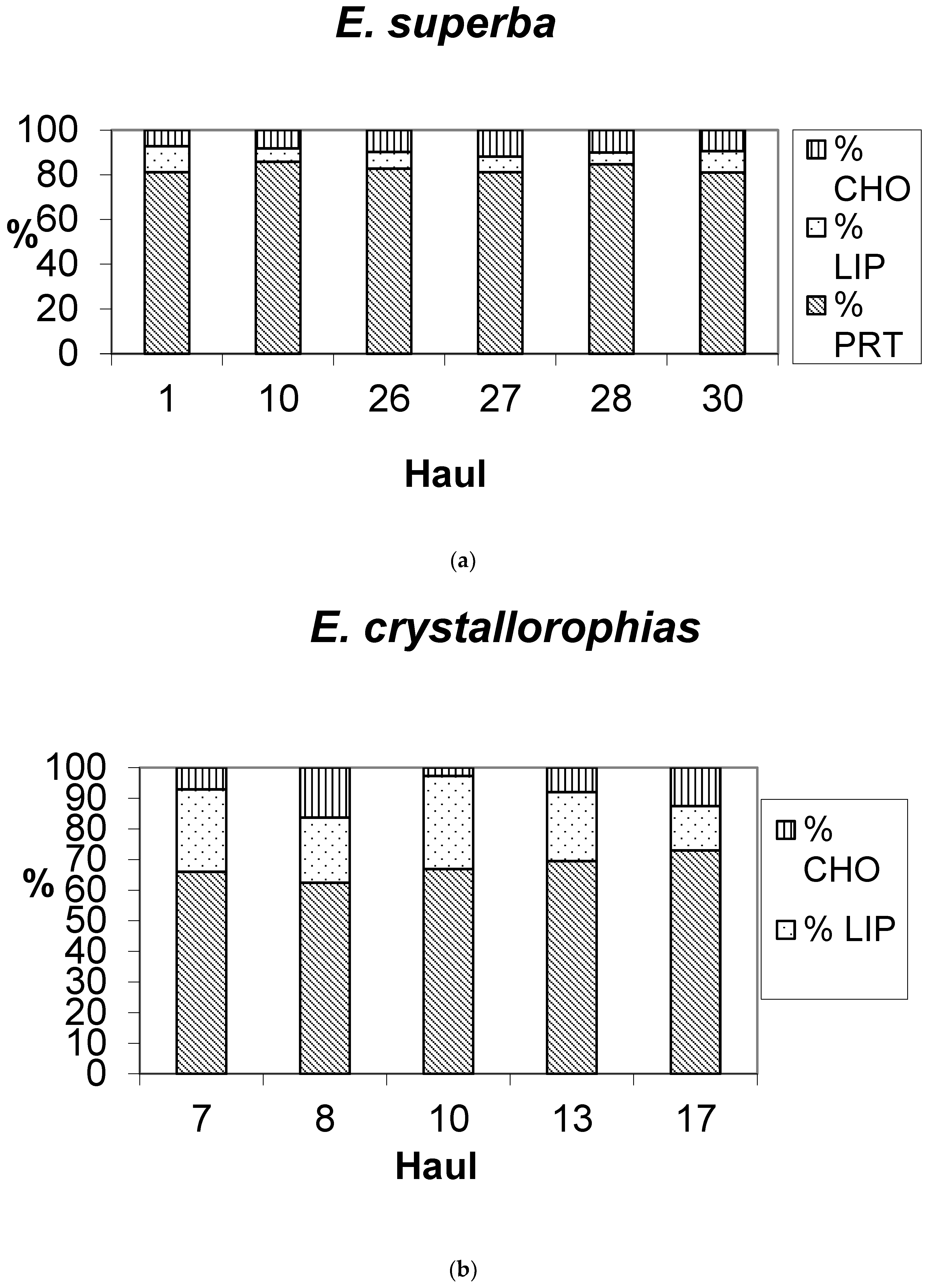
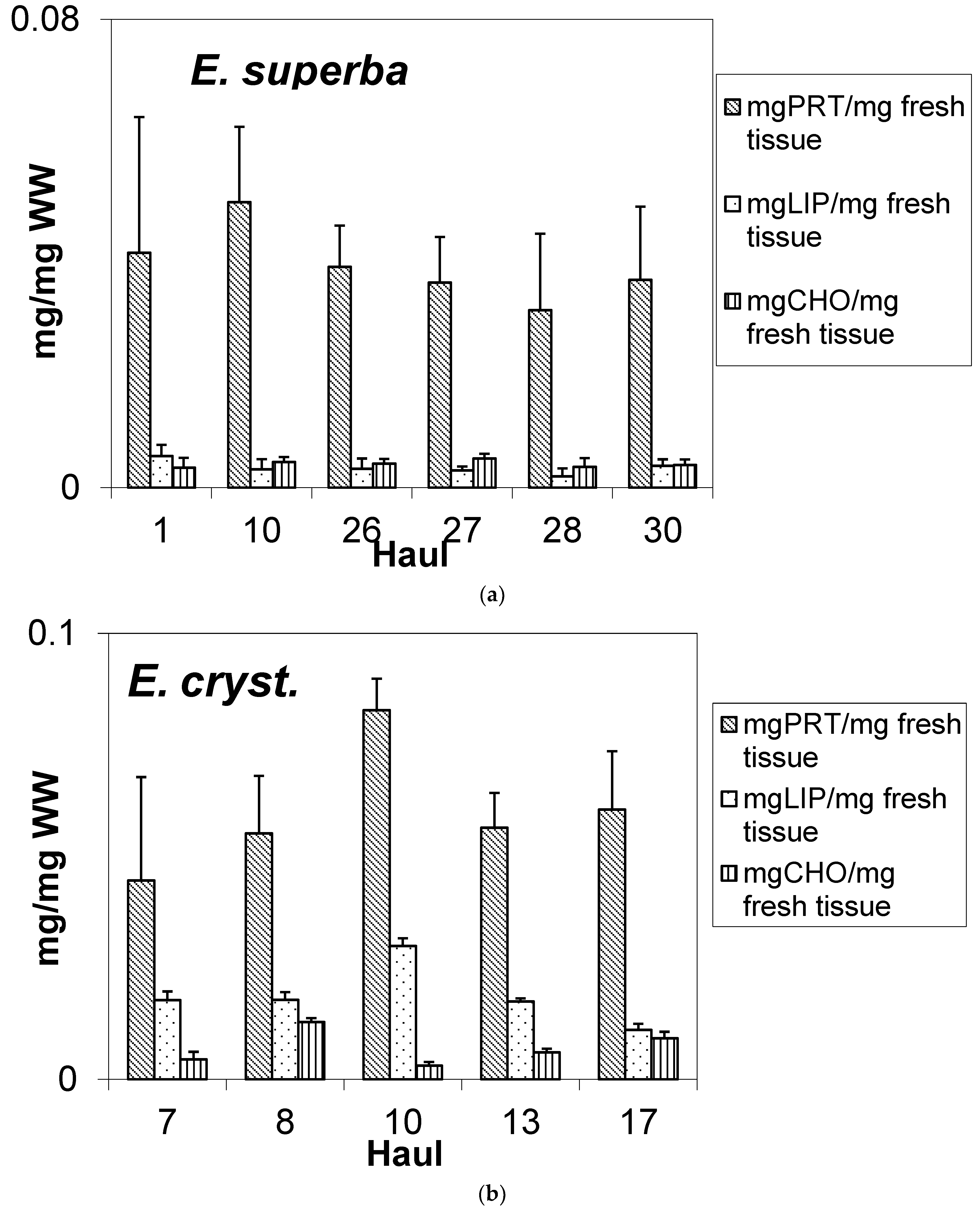
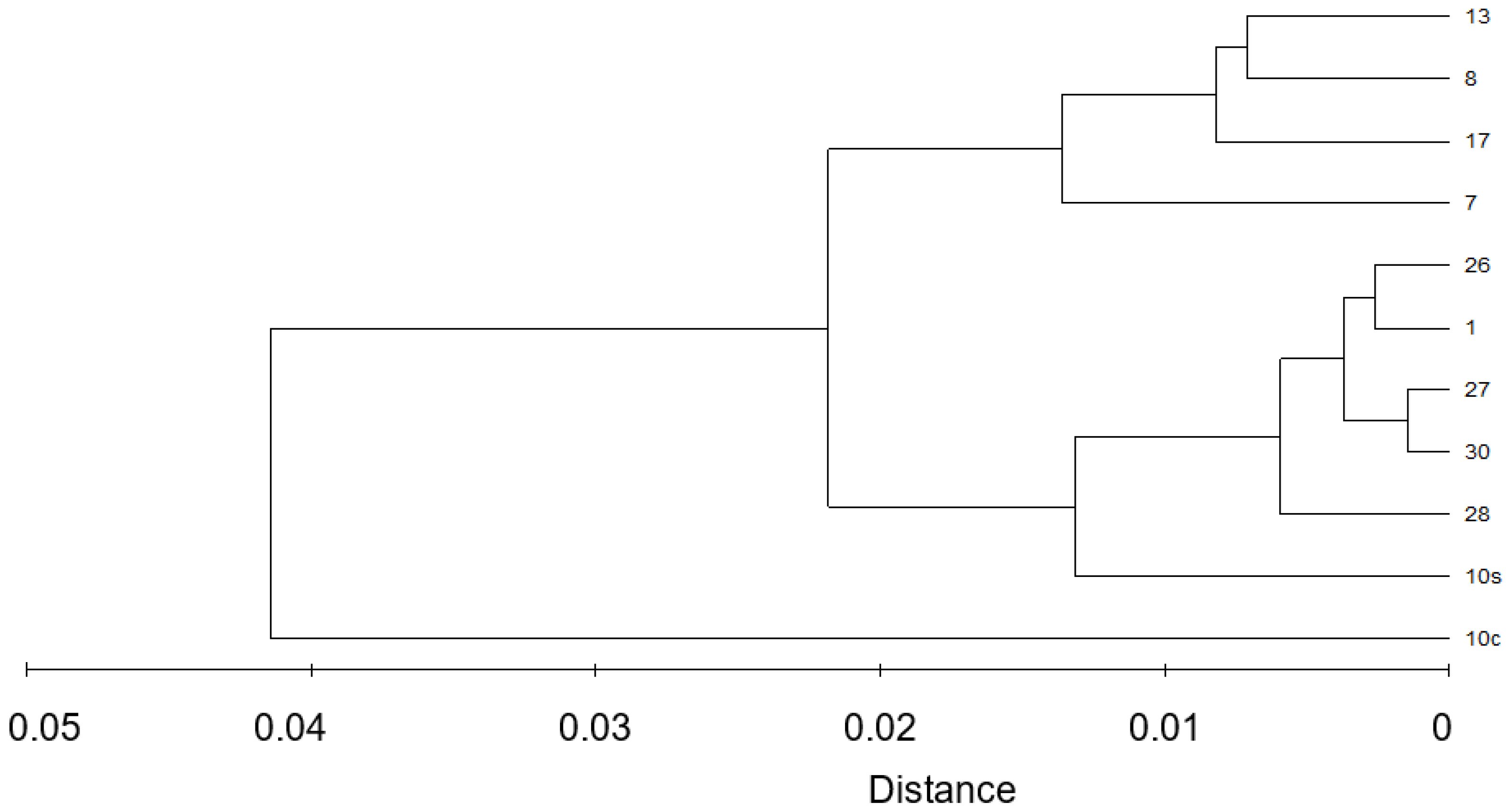
3.1. Biochemical Composition of E. superba and E. crystallorophias
3.2. Fatty Acid Composition in E. superba and E. crystallorophias
4. Discussion
4.1. Biochemical Composition of E. superba and E. crystallorophias
4.2. Fatty Acid Composition in E. superba and E. crystallorophias
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Azzali, M.; Kalinowski, J. Spatial and temporal distribution of krill Euphausia superba biomass in the Ross Sea (1989–1990 and 1994). In Ross Sea Ecology; Faranda, F., Guglielmo, L., Ianora, A., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2000; pp. 433–455. [Google Scholar]
- Leonori, I.; De Felice, A.; Canduci, G.; Costantini, I.; Biagiotti, I.; Giuliani, G.; Budillon, G. Krill distribution in relation to environmental parameters in mesoscale structures in the Ross Sea. J. Mar. Sys. 2017, 166, 159–171. [Google Scholar] [CrossRef]
- Murphy, E.J.; Cavanagh, R.D.; Hofmann, E.E.; Hill, S.L.; Constable, A.J.; Costa, D.P.; Pinkerton, M.H.; Johnston, N.M.; Trathan, P.N.; Klinck, J.M.; et al. Developing integrated models of Southern Ocean food webs: Including ecological complexity, accounting for uncertainty and the importance of scale. Prog. Ocean. 2012, 102, 74–92. [Google Scholar] [CrossRef] [Green Version]
- Hewitt, R.P.; Lipsky, J.D. Krill and other plankton. In Encyclopedia of Marine Mammals, 3rd ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 537–543. [Google Scholar]
- Cavan, E.L.; Belcher, A.; Atkinson, A.; Hill, S.L.; Kawaguchi, S.; McCormack, S.; Meyer, B.; Nicol, S.; Ratnarajah, L.; Schmidt, K.; et al. The importance of Antarctic krill in biogeochemical cycles. Nat. Comm. 2019, 10, 4742. [Google Scholar] [CrossRef] [Green Version]
- Nicol, S. Antarctic Krill Sustainability; Aker Bio Marine Antarctic: Lysaker, Norway, 2014. [Google Scholar]
- Quetin, L.B.; Ross, R.M. Environmental variability and its impact on the reproductive cycle of Antarctic krill. Amer. Zool. 2001, 41, 74–89. [Google Scholar] [CrossRef] [Green Version]
- Rakusa-Suszczewski, S.; Zdanowski, M.K. Bacteria in krill (Euphausia superba Dana) stomach. Acta Protozool. 1989, 28, 87–90. [Google Scholar]
- Flores, H.; Atkinson, A.; Kawaguchi, K.; Krafft, B.A.; Milinevsky, G.; Nicol, S.; Reiss, C.; Tarling, G.A.; Werner, R.; Bravo Rebolledo, E.; et al. Impact of climate change on Antarctic krill. Mar. Ecol. Prog. Ser. 2012, 458, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Veytia, D.; Corney, S.; Meiners, K.M.; Kawaguchi, S.; Murphy, E.J.; Bestley, S. Circumpolar projections of Antarctic krill growth potential. Nat. Clim. Change 2020, 10, 568–575. [Google Scholar] [CrossRef]
- Perissinotto, R.; Gurney, L.; Pakhomov, E.A. Contribution of heterotrophic material to diet and energy budget of Antarctic krill, Euphausia superba. Mar. Biol. 2000, 136, 129–135. [Google Scholar] [CrossRef]
- Schmidt, K.; Atkinson, A.; Petzke, K.; Voss, M.; Pond, D.W. Protozoans as a food source for Antarctic krill, Euphausia superba: Complementary insights from stomach content, fatty acids, and stable isotopes. Limn. Ocean. 2006, 51, 2409–2427. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, T.; Dixon, P. Observations on moulting in Antarctic krill (Euphausia superba Dana). Aust. J. Mar. Freshw. Res. 1982, 33, 71–76. [Google Scholar] [CrossRef]
- Hagen, W.; Auel, H. Seasonal adaptations and the role of lipids in oceanic zooplankton. Zoology 2001, 104, 313–326. [Google Scholar] [CrossRef]
- Falk-Petersen, S.; Hagen, W.; Kattner, G.; Clarke, A.; Sargent, J. Lipids, trophic relationships, and biodiversity in Arctic and Antarctic krill. Can. J. Fish. Aquat. Sci. 2000, 57, 178–191. [Google Scholar] [CrossRef]
- Schaafsma, F.L.; Kohlbach, D.; David, C.; Lange, B.A.; Graeve, M.; Flores, H.; van Franeker, J.A. Spatio-temporal variability in the winter diet of larval and juvenile Antarctic krill, Euphausia superba, in ice-covered waters. Mar. Ecol. Prog. Ser. 2017, 580, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Hagen, W.; Van Vleet, E.S.; Kattner, G. Seasonal storage as overwintering strategy of Antarctic krill. Mar. Ecol. Prog. Ser. 1996, 134, 85–89. [Google Scholar] [CrossRef]
- Nicol, S.; Virtue, P.; King, R.; Davenport, S.R.; McGaffin, A.F.; Nichols, P. Condition of Euphausia crystallorophias off East Antarctica in winter in comparison to other seasons. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2004, 51, 2215–2224. [Google Scholar] [CrossRef]
- Lee, R.F.; Hagen, W.; Kattner, G. Lipid storage in marine zooplankton. Mar. Ecol. Prog. Ser. 2006, 307, 273–306. [Google Scholar] [CrossRef]
- Pond, D.W.; Tarling, G.A. Phase transitions of wax esters adjust buoyancy in diapausing Calanoides acutus. Limn. Ocean. 2011, 56, 1310–1318. [Google Scholar] [CrossRef]
- Pond, D.W. The physical properties of lipids and their role in controlling the distribution of zooplankton in the oceans. J. Plank. Res. 2012, 34, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Hellessey, N.; Ericson, J.A.; Nichols, P.D.; Kawaguchi, S.; Nicol, S.; Hoem, N.; Virtue, P. Regional diet in Antarctic krill (Euphausia superba) as determined by lipid, fatty acid, and sterol composition. Polar Biol. 2022, 45, 1077–1092. [Google Scholar] [CrossRef]
- Ju, S.J.; Harvey, H.R. Lipids as markers of nutritional condition and diet in the Antarctic krill Euphausia superba and Euphausia crystallorophias during austral winter. Deep-Sea Res. II 2004, 51, 2199–2214. [Google Scholar] [CrossRef]
- De Felice, A.; Biagiotti, I.; Canduci, G.; Costantini, I.; Malavolti, S.; Giuliani, G.; Leonori, I. Is It the Same Every Summer for the Euphausiids of the Ross Sea? Diversity 2022, 14, 433. [Google Scholar] [CrossRef]
- Azzali, M.; Russo, A.; Sala, A.; De Felice, A.; Catalano, B. Preliminary results of a survey on krill, environment and predators in CCAMLR Division 88.1 carried out in December 2003 and in January 2004 (Project 8.4). In Proceedings of the CCAMLR WG-EMM-04/71, Siena, Italy, 12–23 July 2004. [Google Scholar]
- Hartree, E.F. Determination of protein: A modification of the Lowry method that gives a linear photometric response. Anal. Biochem. 1972, 48, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Perrone, F.M.; Della Croce, N.; Dell’Anno, A. Biochemical composition and trophic strategies of the amphipod Eurythenes grillus at hadal depths (Atacama Trench, South Pacific). Chem. Ecol. 2003, 19, 441–449. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin-Phenol reagents. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Dubois, N.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Analyt. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Gerchakov, S.M.; Hatcher, P.G. Improved technique for analysis of carbohydrates in sediments. Limn. Ocean 1972, 17, 938–943. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane-Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Christie, W.W. Preparation of derivatives of fatty acids. In Lipid Analysis, 3rd ed.; The Oily Press: Bridgwater, UK, 2003; p. 416. [Google Scholar]
- De Felice, A. Stock e Bioenergetica di due Specie di Eufausiacei (E. superba ed E. crystallorophias) nel Mare di Ross. Ph.D. Thesis, Università Politecnica delle Marche, Ancona, Italy, 9 February 2010. [Google Scholar]
- Sala, A.; Azzali, M.; Russo, A. Krill of the Ross Sea: Distribution, abundance and demography of Euphausia superba and Euphausia crystallorophias during the Italian Antarctic Expedition (January–February 2000). Sci. Mar. 2002, 66, 123–133. [Google Scholar] [CrossRef]
- Watkins, J.L. Variations in the size of Antarctic krill, Euphausia superba Dana, in small swarms. Mar. Ecol. Prog. Ser. 1986, 31, 67–73. [Google Scholar] [CrossRef]
- Azzali, M.; Leonori, I.; De Felice, A.; Russo, A. Spatial-temporal relationships between two euphausiid species in the Ross Sea. Chem. Ecol. 2006, 22, S219–S233. [Google Scholar] [CrossRef]
- Murase, H.; Kitakado, T.; Hakamada, T.; Matsuoka, K.; Nishiwaki, S.; Naganobu, M. Spatial distribution of Antarctic minke whales (Balaenoptera bonaerensis) in relation to spatial distributions of krill in the Ross Sea, Antarctica. Fish. Ocean 2013, 22, 154–173. [Google Scholar] [CrossRef]
- Sologub, D.O.; Remeslo, A.V. Distribution and size-age composition of Antarctic krill (Euphausia superba) in the South Orkney Islands region (CCAMLR subarea 48.2). CCAMLR Sci. 2011, 18, 123–134. [Google Scholar]
- Azzali, M.; Leonori, I.; Lanciani, G. A hybrid approach to acoustical classification and length estimation of krill. CCAMLR Sci. 2004, 11, 33–58. [Google Scholar]
- Sargent, J.R.; Henderson, R.J. Lipids. In The Biological Chemistry of Marine Copepods; Corner, E.D.S., O’Hara, S.C.M., Eds.; Clarendon Press: Oxford, UK, 1986; pp. 59–108. [Google Scholar]
- Danovaro, R.; Pusceddu, A.; Covazzi Harriague, A.; Marrale, D.; Dell’Anno, A.; Petrillo, M.; Albertelli, G.; Della Croce, N. Community experiments using benthic chambers: Microbial significance in highly organic enriched sediments. Chem. Ecol. 1999, 16, 7–30. [Google Scholar] [CrossRef]
- Virtue, P.; Johannes, R.E.; Nichols, P.D.; Young, J.W. Biochemical composition of Nyctiphanes australia and its possible use as an aquaculture feed source: Lipids, pigments and fluoride content. Mar. Biol. 1995, 122, 121–128. [Google Scholar] [CrossRef]
- Clarke, A.; Peck, L.S. The physiology of polar marine zooplankton. Polar Res. 1991, 10, 355–369. [Google Scholar] [CrossRef] [Green Version]
- Bamstedt, U. Studies on the deep-water pelagic community of Korsfjorden, western Norway. Ecological aspects of individual variations in weight and protein and lipid content of Euchaeta norvegica (Copepoda). Sarsia 1976, 61, 15–30. [Google Scholar]
- Bamstedt, U. Studies on the deep-water pelagic community of Korsfjorden, western Norway. Seasonal variation in weight and biochemical composition of Chiridius armatus (Copepoda), Boreomysis arctica (Mysidacea), and Eukrohnia hamata (Chaetognata) in relation to their biology. Sarsia 1978, 63, 145–154. [Google Scholar] [CrossRef]
- Miliou, H.; Moraitou-Apostolopoulou, M. Effects of seven diets on the population dynamics of laboratory cultured Tisbeholothuriae Humes (Copepoda, Harpacticoida). Helgol. Meeresunters 1991, 45, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Lehtonen, K.K. Ecophysiology of the benthic amphipod Monoporeia affinis in an open-sea area of the northern Baltic Sea: Seasonal and interannual variations in body composition, with bioenergetic considerations. Mar. Ecol. Prog. Ser. 1996, 14, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Ericson, J.A.; Hellessey, N.; Nichols, P.D.; Nicol, S.; Kawaguchi, S.; Hoem, N.; Virtue, P. New insights into the seasonal diet of Antarctic krill using triacylglycerol and phospholipid fatty acids, and sterol composition. Pol. Biol. 2019, 42, 1985–1996. [Google Scholar] [CrossRef]
- Phleger, C.F.; Nelson, M.M.; Mooney, B.D.; Nichols, P.D. Interannual and between species comparison of the lipids, fatty acids and sterols of Antarctic krill from the US AMLR Elephant Island survey area. Comp. Biochem. Physiol. Part B 2002, 131, 733–747. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Li, C.; Wang, Y. Fatty acid composition of Euphausia superba, Thysanoessa macrura and Euphausia crystallorophias collected from Pridz Bay, Antarctica. J. Ocean. Univ. China 2016, 15, 297–302. [Google Scholar] [CrossRef]
- Ko, A.R.; Yang, E.J.; Kim, M.S.; Ju, S.-J. Trophodynamics of euphausiids in the Amundsen Sea during the austral summer by fatty acid and stable isotopic signatures. Deep-Sea Res. II 2016, 123, 78–85. [Google Scholar] [CrossRef]
- Hagen, W.; Kattner, G.; Terbruggen, A.; Van Vleet, E.S. Lipid metabolism of the Antarctic krill Euphausia superba and its ecological implications. Mar. Biol. 2001, 139, 95–104. [Google Scholar] [CrossRef]
- Kattner, G.; Hagen, W. Lipid metabolism of the Antarctic euphausiid Euphausia crystallorophias and its ecological implication. Mar. Ecol. Prog. Ser. 1998, 170, 203–213. [Google Scholar] [CrossRef]
- Stübing, D.; Hagen, W. Fatty acid biomarker ratios-suitable trophic indicators in Antarctic euphausiids? Pol. Biol. 2003, 26, 774–782. [Google Scholar] [CrossRef]
- Cripps, G.C.; Watkins, J.L.; Hill, H.J.; Atkinson, A. Fatty acid content of Antarctic krill Euphausia superba at South Georgia related to regional populations and variations in diet. Mar. Ecol. Prog. Ser. 1999, 181, 177–188. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Haul | Lat. | Long. | Water Depth (m) | Net Depth (m) | Ice Cover (%) |
|---|---|---|---|---|---|
| 1 | 68°42.3′ S | 179°53.1′ W | 3438.5 | 120 | 30 |
| 7 | 75°56.4′ S | 178°30.9′ E | 540.0 | 100 | 20 |
| 8 | 75°36.0′ S | 172°39.8′ E | 555.5 | 100 | 0 |
| 10 | 75°6.59′ S | 177°24.5′ W | 450.9 | 37 | 10 |
| 13 | 74°27.1′ S | 170°0.35′ E | 578.3 | 125 | 0 |
| 17 | 73°8.8′ S | 170°44.4′ E | 512.6 | 75 | 0 |
| 26 | 70°10.6′ S | 171°56.8′ E | 1479.0 | 40 | 0 |
| 27 | 70°2.69′ S | 169°54.0′ E | 2662.5 | 21 | 0 |
| 28 | 69°54.1′ S | 172°0.95′ E | 1863.0 | 35 | 0 |
| 30 | 69°11.7′ S | 171°47.8′ E | 3043.5 | 64 | 0 |
| Species | Haul | No. | Mean TL (mm) | σTL (mm) | Mean WW (g) | σWW (g) | TOM (%) | Ash (%) |
|---|---|---|---|---|---|---|---|---|
| E. superba | 1 | 20 | 42.94 | 3.72 | 0.1113 | 0.0283 | na | na |
| E. superba | 10 | 20 | 39.92 | 4.90 | 0.1156 | 0.0441 | na | na |
| E. superba | 26 | 20 | 41.75 | 4.01 | 0.1277 | 0.0481 | 84.9 | 15.1 |
| E. superba | 27 | 20 | 41.82 | 2.46 | 0.1026 | 0.0268 | 81.8 | 18.2 |
| E. superba | 28 | 20 | 40.56 | 2.29 | 0.1115 | 0.0348 | 88.4 | 11.6 |
| E. superba | 30 | 20 | 42.48 | 2.50 | 0.1233 | 0.0383 | 82.9 | 17.1 |
| E. cryst. | 7 | 20 | 24.12 | 1.82 | 0.0281 | 0.0171 | 86.4 | 13.6 |
| E. cryst. | 8 | 20 | 22.35 | 2.22 | 0.0167 | 0.0093 | 86.0 | 14.0 |
| E. cryst. | 10 | 20 | 24.85 | 3.04 | 0.0224 | 0.0087 | na | na |
| E. cryst. | 13 | 20 | 22.84 | 3.30 | 0.0258 | 0.0157 | 88.1 | 11.9 |
| E. cryst. | 17 | 20 | 23.26 | 3.28 | 0.0138 | 0.0055 | 82.0 | 18.0 |
| Species and Life Stage | Haul | Specimen No. | Mean TL (mm) | σTL (mm) | SFAs (%) | MUFAs (%) | PUFAs (%) | PUFAs/SFAs |
|---|---|---|---|---|---|---|---|---|
| E. superba adults | 1 | 5 | 44.70 | 1.70 | 28.6 | 30.0 | 41.4 | 1.4 |
| E. superba subadults | 1 | 8 | 29.60 | 10.15 | 27.4 | 36.6 | 36.0 | 1.3 |
| E. superba adults | 10 | 13 | 40.85 | 4.82 | 36.3 | 30.3 | 33.4 | 0.9 |
| E. superba adults | 27 | 11 | 41.20 | 2.67 | 33.8 | 29.2 | 37.0 | 1.1 |
| E. superba subadults | 27 | 9 | 31.85 | 3.55 | 31.8 | 29.2 | 39.0 | 1.2 |
| E. crystallorophias adults | 7 | 25 | 24.85 | 2.58 | 16.4 | 49.3 | 34.3 | 2.1 |
| E. crystallorophias adults | 17 | 32 | 24.88 | 2.37 | 20.7 | 42.4 | 36.9 | 1.8 |
| Fatty Acid | Haul 1 (E. s. Adults) | Haul 1 (E. s. Subadults) | Haul 10 (E. s. Adults) | Haul 27 (E. s. Adults) | Haul 27 (E. s. Subadults) |
|---|---|---|---|---|---|
| C14:0 | 6.0 | 5.6 | 10.4 | 8.7 | 9.2 |
| C15:0 | 0.4 | 0.3 | 0.2 | 0.1 | 0.1 |
| C16:0 | 21.0 | 19.7 | 23.7 | 23.1 | 20.8 |
| C16:1ω7 | 3.4 | 3.3 | 5.3 | 3.9 | 4.1 |
| C16:2 | 1.4 | 1.2 | 1.4 | 1.2 | 1.2 |
| C18:0 | 1.3 | 1.8 | 2.0 | 1.9 | 1.7 |
| C18:1ω9C | 16.7 | 25.8 | 16.2 | 16.8 | 16.2 |
| C18:1ω7C | 8.5 | 6.1 | 7.1 | 7.2 | 7.4 |
| C18:2ω6 | 3.4 | 3.8 | 2.4 | 2.1 | 1.8 |
| C18:3ω6 | 0.1 | 0.2 | 0.4 | 0.6 | 0.7 |
| C18:3ω3 | 0.7 | 0.5 | 0.8 | 0.8 | 0.8 |
| C18:4ω3 | 2.6 | 2.4 | 5.5 | 5.3 | 6.2 |
| C20:1ω9 | 1.1 | 1.1 | 1.3 | 0.9 | 1.2 |
| C20:3ω6 | 0.5 | 0.5 | 0.6 | 0.4 | 0.3 |
| C20:4ω6 | 0.8 | 0.7 | 0.4 | 0.6 | 0.6 |
| C20:5ω3 | 16.7 | 13.6 | 11.8 | 12.5 | 12.8 |
| C22:1ω9 | 0.2 | 0.2 | 0.4 | 0.4 | 0.4 |
| C22:6ω3 | 15.2 | 13.1 | 10.0 | 13.5 | 14.5 |
| PUFA/SFA | 1.4 | 1.3 | 0.9 | 1.1 | 1.2 |
| DHA/EPA | 0.9 | 1.0 | 0.9 | 1.1 | 1.1 |
| C18:4ω3/C16:1ω7 | 0.8 | 0.7 | 1.0 | 1.4 | 1.5 |
| C18:1ω9C/C18:1ω7C | 2.0 | 4.2 | 2.3 | 2.3 | 2.2 |
| 18PUFA/16PUFA | 4.9 | 5.8 | 6.5 | 7.3 | 7.9 |
| Fatty Acid | Haul 7 (E. c. Adults) | Haul 17 (E. c. Adults) |
|---|---|---|
| C14:0 | 2.0 | 3.7 |
| C15:0 | 0.1 | 0.1 |
| C16:0 | 13.2 | 15.9 |
| C16:1ω7 | 4.1 | 4.5 |
| C16:2 | 0.4 | 0.4 |
| C18:0 | 1.0 | 1.0 |
| C18:1ω9C | 32.5 | 26.9 |
| C18:1ω7C | 12.0 | 10.1 |
| C18:2ω6 | 2.0 | 1.9 |
| C18:3ω6 | 0.4 | 0.4 |
| C18:3ω3 | 0.3 | 0.4 |
| C18:4ω3 | 2.4 | 2.0 |
| C20:1ω9 | 0.6 | 0.8 |
| C20:3ω6 | 0.2 | 0.3 |
| C20:4ω6 | 0.6 | 0.7 |
| C20:5ω3 | 15.7 | 16.4 |
| C22:1ω9 | 0.1 | 0.1 |
| C22:6ω3 | 12.3 | 14.4 |
| PUFA/SFA | 2.1 | 1.8 |
| DHA/EPA | 0.8 | 0.9 |
| C18:4ω3/C16:1ω7 | 0.6 | 0.4 |
| C18:1ω9C/C18:1ω7C | 2.7 | 2.7 |
| 18PUFA/16PUFA | 12.8 | 11.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Felice, A.; Manini, E.; Biagiotti, I.; Leonori, I. Bioenergetics of Euphausia superba and Euphausia crystallorophias in the Ross Sea. Diversity 2023, 15, 480. https://doi.org/10.3390/d15040480
De Felice A, Manini E, Biagiotti I, Leonori I. Bioenergetics of Euphausia superba and Euphausia crystallorophias in the Ross Sea. Diversity. 2023; 15(4):480. https://doi.org/10.3390/d15040480
Chicago/Turabian StyleDe Felice, Andrea, Elena Manini, Ilaria Biagiotti, and Iole Leonori. 2023. "Bioenergetics of Euphausia superba and Euphausia crystallorophias in the Ross Sea" Diversity 15, no. 4: 480. https://doi.org/10.3390/d15040480