
Unrecognised Ant Megadiversity in Monsoonal Australia: The Tetramorium spininode Bolton Group in the Northern Territory

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
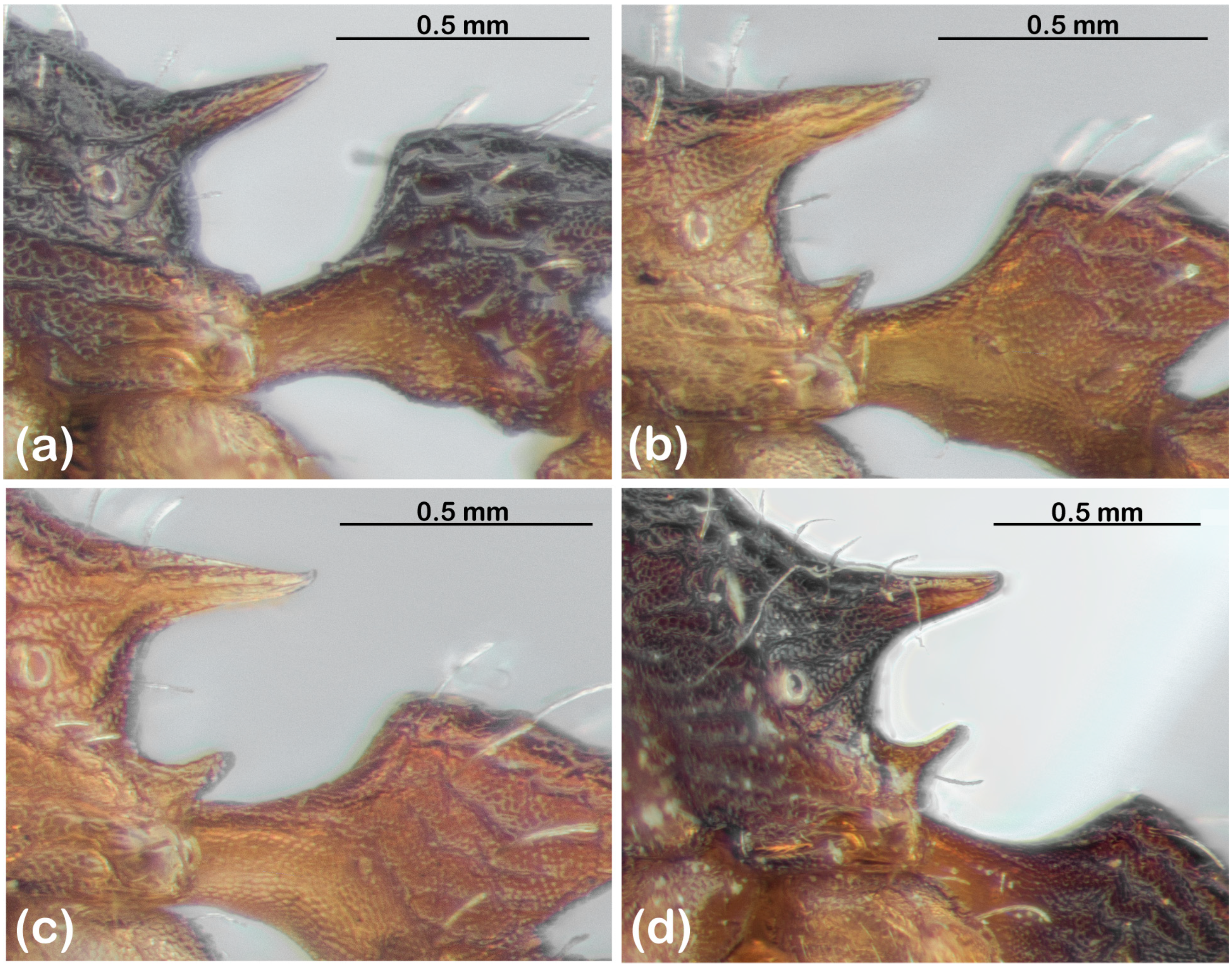
- Gastral sculpture, which ranges from finely striate (typically longitudinally, but sometimes radiating from the base, and with varying degrees of punctate background sculpture) to spectacularly costate (Figure 3);
- Rugosity of the head and mesosoma (Figure 5);
- Body colour, which ranges from uniformly yellowish or orange-brown to bicoloured, with dark brown head, mesosoma and waist, contrasting with yellowish legs and gaster (Figure 5).
2. Materials and Methods
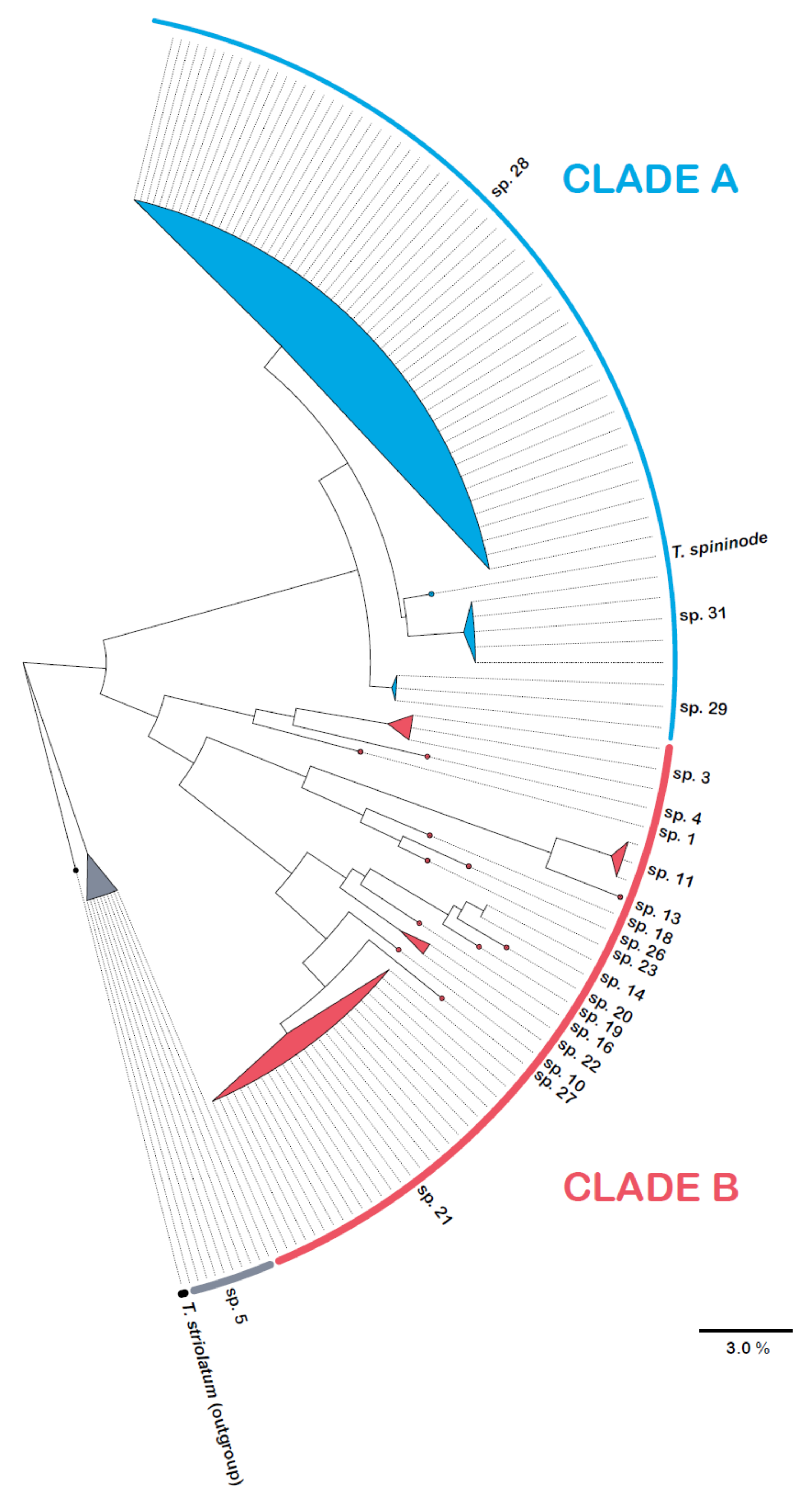
3. Results
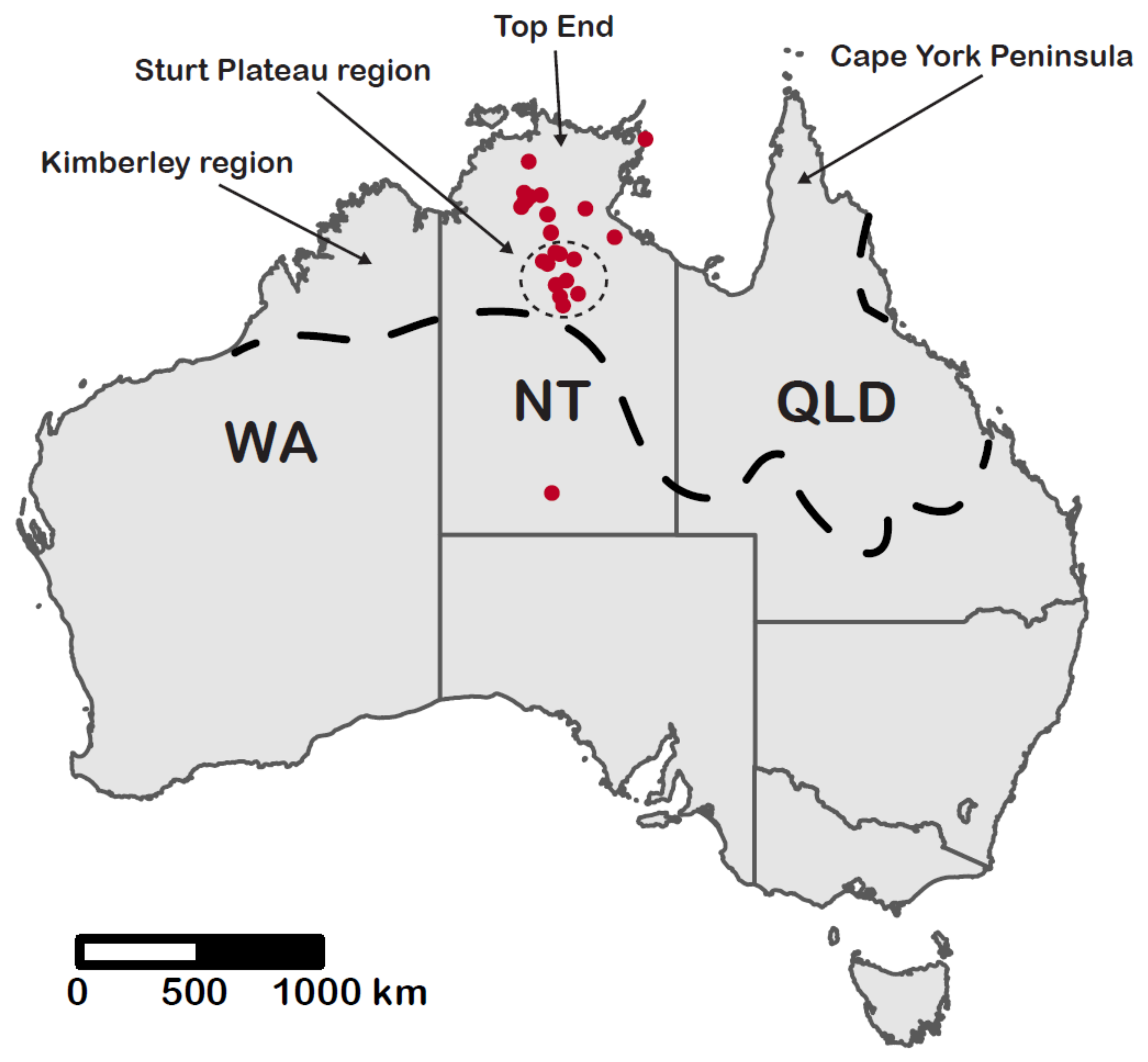
3.1. Species Richness
- First gastral tergite costate ……………………………………………………….…………………..……………..……….2First gastral tergite striate ..……………………………………………………….………………….…………..………….8
- Metapleural lobes strongly developed and dentiform or spinose .…….……………………………….……………….3Metapleural lobes weakly developed and rounded .……………………………………………………………………..6
- Metapleural spines short and broadly triangular …………………….……………………….…………….………....….4Metapleural spines long and narrow ……………………………………………………..………………………..……….5
- Body reddish brown contrasting with yellowish legs; sides of head behind eyes regularly rugose (Forest Hill and Manbulloo stns)…………………………………………………..….……..….……..….……..….……..……...….…….sp. 1Body uniformly yellowish; sides of head behind eyes irregularly sculptured (Manbulloo Stn) ....……...……....sp. 2
- Sides of head behind eyes regularly rugose; gastral costae more strongly radial anteriorly (Hayfield Shenandoah Stn) ……….………….……………………..………………..……………..……………..………………………….…….sp. 3Sides of head behind eyes irregularly sculptured; gastral costae less strongly radial anteriorly (Lorella Springs) …………………………………………………………..………………………………………………………..sp. 4
- Gastral costae transverse anteriorly (throughout Sturt Plateau) …………………………………………...………..sp. 5Gastral costae never transverse ………………………………………..…………………………………………………….7
- Small species with short petiolar and postpetiolar nodes that are not so strongly reflexed (Katherine region; Figure 5c) …...…….…………………………………….…………………………….…..…………….……………………..sp. 6Larger species with more strongly reflexed petiolar and postpetiolar nodes (Groote Eylandt, Gulf region) ..………….……………………………….…………………….…………………..………….…………………….sp. 7
- Metapleural lobes strongly developed and dentiform or spinose …….………………………………………..……….9Metapleural lobes not strongly developed, rounded or very weakly dentate …………………………………………26
- Mesosoma with a deep metanotal groove (Manyallaluk; Figure 5b) ……………………………………………….sp. 8Mesosoma with at most a weak metanotal notch ………………………..……………..……………..………………….10
- First gastral tergite with a rounded medial ridge such that in profile it is angled anteriorly, with a perpendicular anterior face ………..…………………………………………………………………………………………..…………….11First gastral tergite without a medial ridge such that its profile lacks a distinct anterior face ……..……..………..15
- Mesosoma conspicuously rugose throughout most of its lateral faces …..…………..…………..…………….………2Mesosoma without conspicuous rugae on most of its lateral faces ………….………….………….………….………..13
- Petiole and postpetiole conspicuously darker than mesosoma and gaster (Manyallaluk, Beswick; Figure 5d) ……………………………………………………………..………………………………………………...……….……..sp. 9Petiole and postpetiole concolorous with mesosoma and gaster (Nitmiluk NP) .…….……….……….…………sp. 10
- Ventral processes of postpetiole yellowish, distinctly lighter than node ……………..…...……...……...…...……….14Ventral processes of postpetiole reddish to dark brown, concolorous with node (Forest Hill and Manbulloo stns; Figure 5e) ……………………….………………………………..….……………..….……………..….…..….………..sp. 11
- First gastral tergite more strongly flanged, and lateral striations strongly radiating towards the medial line (King River; Figure 4) ………………………………………………………………………………………………………….sp. 12First gastral tergite less strongly flanged, and lateral striations less strongly radiating towards the medial line (Lakefield, Manbulloo and Mataranka stns ……………….…………….…………….…………….……………….sp. 13
- Mesosoma yellowish brown, concolorous with gaster ………………….…………….…..…………………………….16Mesosoma reddish or blackish brown, distinctly darker than gaster ………………….…………….……….………...20
- Gastral striations coarse ………………………………………………….……..………….…………….…...………..…...17Gastral striations fine, often faint …………………………………………………………….…………….…..………….18
- Metapleural spines long and narrow (Kakadu NP) .……………………….…………………………….………….sp. 14Metapleural spines short and triangular (Vermalha Stn) ………………..…………………………………..……...sp. 15
- First gastral tergite primarily punctate anteriorly, with only feeble striations (Kakadu NP) ………………..….sp. 16First gastral tergite finely but conspicuously striate throughout …………..………………………..………..…………19
- First gastral tergite with punctate background sculpture, giving it a somewhat matt appearance; rugae on frontal area finer and more closely approximated (Kakadu NP) ……………………………………………...…………....sp. 17First gastral tergite with feeble background sculpture, giving it a shiny appearance; rugae on frontal area very coarse and widely separated (Kakadu NP, Nitmiluk NP, Arnhem Land) ………………………………………...sp. 18
- Head, mesosoma and waist orange-brown; metapleural lobes spinose; first gastral tergite without striations on its posterior half medially (Gove) …………………………………………………………………………...…..………...sp. 19Head, mesosoma and waist darker reddish or blackish brown ……………..…………..……………..……………....21
- First gastral tergite conspicuously striate throughout …………………………………..……………..………………..22First gastral tergite mostly smooth and shiny medially on posterior half ……..…………..……………..……………24
- Metapleural spines especially long and narrow; first gastral tergite with feeble background sculpture, giving it a shiny appearance (Kakadu NP) ……………….………………..……………..……………..………………………...sp. 20Metapleural spines not so long and narrow; first gastral tergite with punctate background sculpture, giving it a more matt appearance ……………………………..……………..……………..……………..……..…………………….23
- Basal flanges of first gastral tergite weakly developed; in dorsal view, lateral margins of first gastral tergite only weakly curved (Sturt Plateau region, Kakadu NP) .……………..……………..……………..……………………..sp. 21Basal flanges of first gastral tergite strongly developed; in dorsal view, lateral margins of first gastral tergite strongly curved (eastern Top End) .….……………………………………..……………..……………..…………...sp. 22
- Metapleural spines short and broadly triangular (Kakadu NP) …………..…………..……………..…..………...sp. 23Metapleural spines long and narrow ………………………..…………………..……………..………………………….25
- First gastral tergite with very feeble background punctation, giving it a shiny appearance (Cobourg Peninsula, Blue Mud Bay) …………………………………..……………………..……………..……………..……………..…….sp. 24First gastral tergite with stronger background punctation, giving it a more matt appearance (Limmen NP) …………………………………………………………..……………..……………..……………………………..sp. 25
- Mesosoma dark brown, contrasting with light honey-brown legs and gaster ………………..……………..………..27Mesosoma yellowish or reddish brown, more or less concolorous with legs and gaster ……………………………………….……………………………………………..……..…………..……….………….28
- Anterior half of pronotal dorsum with regular, coarse rugae (Kimberley, WA) ……..….………….……….spininodeAnterior half of pronotal dorsum irregularly sculptured (Kakadu NP) …………………..……………..………..sp. 26
- Colour darker reddish brown, rugae on frontal area coarser ……………………………..……...………..…………..29Colour paler yellowish or orange-brown; rugae on frontal area not so coarse ..…………..……………..…………..31
- Lateral striations on first gastral tergite diverging from the base (Tandidgee Stn)………………………..……...sp. 27Striations on first gastral tergite parallel throughout ……………………...…………………..……………..…………..30
- Gastral striations finer (Sturt Plateau, Victoria River District, Gulf region) ………………..……………...……..sp. 28Gastral striations coarser (Newcastle Waters and Hidden Valley stns) ......……..……...…...…...…...…...…….sp. 29
- Striations on first gastral tergite parallel throughout (Vermhala Stn) ………..…………...…...…...…....…...…..sp. 30Lateral striations on first gastral tergite either diverging or converging from the base ……...…...…...…....….......32
- Lateral striations on first gastral tergite diverging from the base (Tandidgee, Newcastle Waters and Henbury Stations, Alice Springs, Uluru NP) ……………………..…………………………………………..………..………...….sp. 31Lateral striations on first gastral tergite converging from the basal lobes (Limmen NP) ………………………..sp. 32
3.2. Species Turnover
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andersen, A. The Ants of Northern Australia: A Guide to the Monsoonal Fauna; CSIRO Publishing: Collingwood, VIC, Australia, 2000; ISBN 0643066039. [Google Scholar]
- Oberprieler, S.K.; Andersen, A.N.; Moritz, C.C. Ants in Australia’s monsoonal tropics: CO1 barcoding reveals extensive unrecognised diversity. Diversity 2018, 10, 36. [Google Scholar] [CrossRef] [Green Version]
- Andersen, A.N.; Vasconcelos, H.L. Historical biogeography shapes functional ecology: Inter-continental contrasts in responses of savanna ant communities to stress and disturbance. J. Biogeogr. 2022, 49, 590–599. [Google Scholar] [CrossRef]
- Andersen, A.N.; Hoffmann, B.D.; Oberprieler, S.K. Megadiversity in the ant genus Melophorus: The M. rufoniger Heterick, Castalanelli and Shattuck species group in the Top End of Australia’s Northern Territory. Diversity 2020, 12, 386. [Google Scholar] [CrossRef]
- Andersen, A.N.; Brassard, F.; Hoffmann, B.D. Ant megadiversity in monsoonal Australia: Diversity and distribution in the hyperdiverse Monomorium nigrius Forel group. Diversity 2022, 14, 46. [Google Scholar] [CrossRef]
- Andersen, A.N. Ant megadiversity and its origins in arid Australia. Austral Entomol. 2016, 55, 132–147. [Google Scholar] [CrossRef]
- Bolton, B. The ant tribe Tetramoriini (Hymenoptera: Formicidae). The genus Tetramorium Mayr in the Oriental and Indo Australian regions, and in Australia. Bull. Br. Mus. (Nat. Hist.) Entomol. 1977, 36, 67–151. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, M.A.; Fisher, B.L.; Hebert, P.D.N. DNA barcoding for effective biodiversity assessment of a hyperdiverse arthropod group: The ants of Madagascar. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1825–1834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlick-Steiner, B.C.; Steiner, F.M.; Moder, K.; Seifert, B.; Sanetra, M.; Dyreson, E.; Stauffer, C.; Christian, E. A multidisciplinary approach reveals cryptic diversity in Western Palearctic Tetramorium ants (Hymenoptera: Formicidae). Mol. Phylogenetics Evol. 2006, 40, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Bickford, D.; Lohman, D.J.; Sodhi, N.S.; Ng, P.K.L.; Meier, R.; Winker, K.; Ingram, K.K.; Das, I. Cryptic species as a window on diversity and conservation. Trends Ecol. Evol. 2007, 22, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Kass, J.M.; Guénard, B.; Dudley, K.L.; Jenkins, C.N.; Azuma, F.; Fisher, B.L.; Parr, C.L.; Gibb, H.; Longino, J.Y.; Ward, P.S.; et al. The global distribution of known and undiscovered ant biodiversity. Sci. Adv. 2022, 8, eabp9908. [Google Scholar] [CrossRef] [PubMed]
- Hanisch, P.E.; Lavinia, P.D.; Suarez, A.V.; Lijtmaer, D.A.; Leponce, M.; Paris, C.I.; Tubaro, P.L. Mind the gap! Integrating taxonomic approaches to assess ant diversity at the southern extreme of the Atlantic Forest. Ecol. Evol. 2017, 7, 10451–10466. [Google Scholar] [CrossRef] [PubMed]
- Brühl, C.A.; Gunsalam, G.; Linsenmair, K.E. Stratification of ants (Hymenoptera, Formicidae) in a primary rain forest in Sabah, Borneo. J. Trop. Ecol. 1998, 14, 285–297. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andersen, A.N.; Brassard, F.; Hoffmann, B.D. Unrecognised Ant Megadiversity in Monsoonal Australia: The Tetramorium spininode Bolton Group in the Northern Territory. Diversity 2023, 15, 476. https://doi.org/10.3390/d15040476
Andersen AN, Brassard F, Hoffmann BD. Unrecognised Ant Megadiversity in Monsoonal Australia: The Tetramorium spininode Bolton Group in the Northern Territory. Diversity. 2023; 15(4):476. https://doi.org/10.3390/d15040476
Chicago/Turabian StyleAndersen, Alan N., François Brassard, and Benjamin D. Hoffmann. 2023. "Unrecognised Ant Megadiversity in Monsoonal Australia: The Tetramorium spininode Bolton Group in the Northern Territory" Diversity 15, no. 4: 476. https://doi.org/10.3390/d15040476