
Haplosporidium pinnae Detection from the Faeces of Pinna nobilis: A Quick and Noninvasive Tool to Monitor the Presence of Pathogen in Early-Stage or during Fan Mussel Mass Mortalities
 , ,
, ,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples Collection, DNA Extraction, and Amplification
2.2. Primer Design
2.3. Limit of Detection and Inhibition Test
3. Results
3.1. Haplosporidium Pinnae Assay
3.2. Amplification of H. pinnae DNA from Faeces and Pseudofaeces
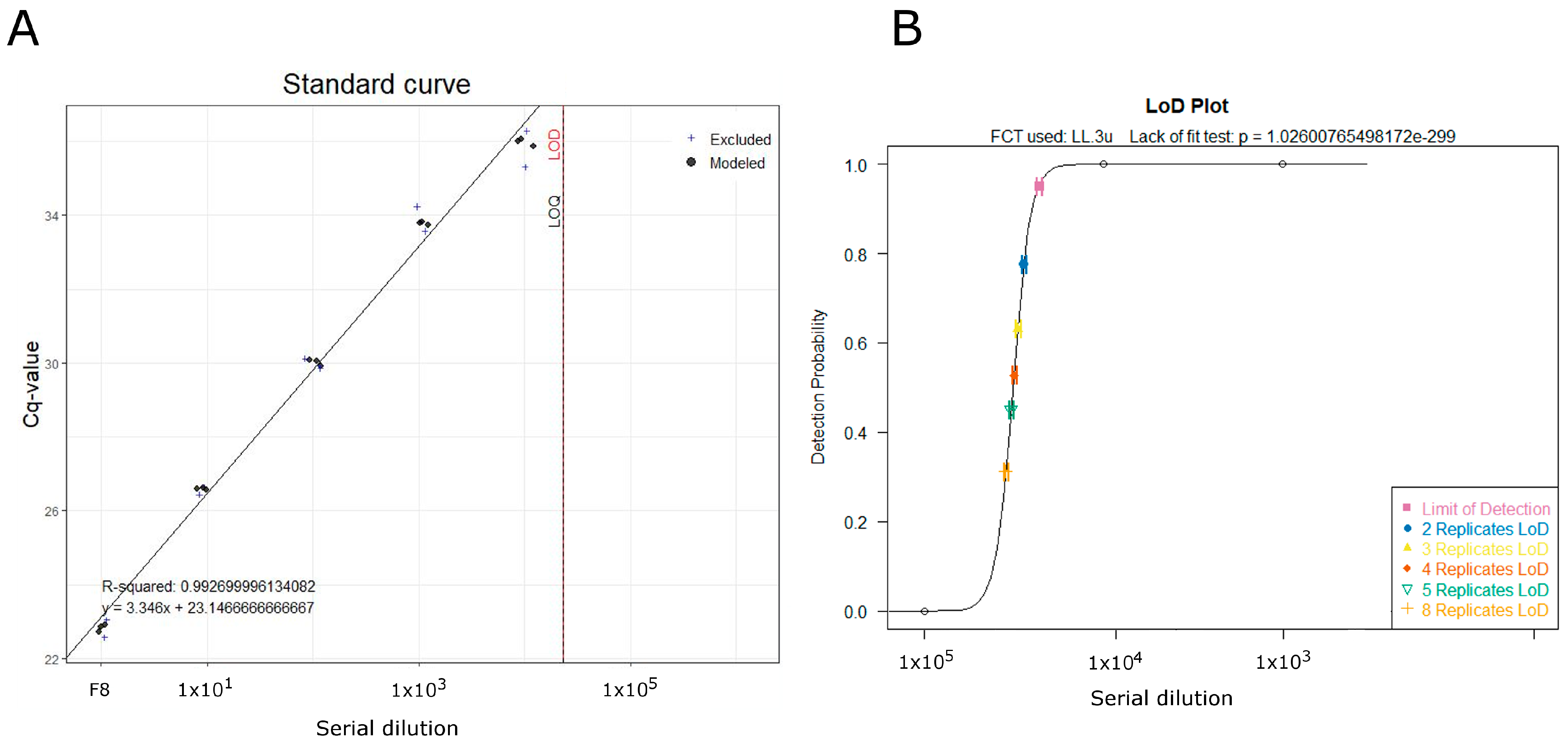
3.3. Limit of Detection and Inhibition Test
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Gozlan, R.E.; Peeler, E.J.; Longshaw, M.; St-Hilaire, S.; Feist, S.W. Effect of microbial pathogens on the diversity of aquatic populations, notably in Europe. Microbes Infect. 2006, 8, 1358–1364. [Google Scholar] [CrossRef] [PubMed]
- Harvell, D.; Aronson, R.; Baron, N.; Connell, J.; Dobson, A.; Ellner, S.; Gerber, L.; Kim, K.; Kuris, A.; McCallum, H.; et al. The rising tide of ocean diseases: Unsolved problems and research priorities. Front. Ecol. Environ. 2004, 2, 375–382. [Google Scholar] [CrossRef]
- Harvell, C.D.; Kim, K.; Burkholder, J.M.; Colwell, R.R.; Epstein, P.R.; Grimes, D.J.; Hofmann, E.E.; Lipp, E.K.; Osterhaus, A.D.M.E.; Overstreet, R.M.; et al. Emerging marine diseases—Climate links and anthropogenic factors. Science 1999, 285, 1505–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Öndes, F.; Kaiser, M.J.; Güçlüsoy, H. Human impacts on the endangered fan mussel, Pinna nobilis. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 31–41. [Google Scholar] [CrossRef]
- Basso, L.; Vázquez-Luis, M.; García-March, J.R.; Deudero, S.; Alvarez, E.; Vicente, N.; Duarte, C.M.; Hendriks, I.E. The Pen Shell, Pinna nobilis: A Review of Population Status and Recommended Research Priorities in the Mediterranean Sea. Adv. Mar. Biol. 2015, 71, 109–160. [Google Scholar] [CrossRef]
- Maeder, F. Sea-silk in Aquincum: First Production proof in antiquity. In Purpureae Vestes II. Textiles and Dyes in Antiquity; Alfaro, C., Karali, L., Eds.; University of Valencia: Valencia, Spain, 2008; pp. 109–118. [Google Scholar]
- Katsanevakis, S. The cryptogenic parasite Haplosporidium pinnae invades the Aegean Sea and causes the collapse of Pinna nobilis populations. Aquat. Invasions 2019, 14, 150–164. [Google Scholar] [CrossRef]
- Vázquez-Luis, M.; Álvarez, E.; Barrajón, A.; García-March, J.R.; Grau, A.; Hendriks, I.E.; Jiménez, S.; Kersting, D.; Moreno, D.; Pérez, M.; et al. S.O.S. Pinna nobilis: A mass mortality event in western Mediterranean Sea. Front. Mar. Sci. 2017, 4, 220. [Google Scholar] [CrossRef] [Green Version]
- Catanese, G.; Grau, A.; Valencia, J.M.; Garcia-March, J.R.; Vázquez-Luis, M.; Alvarez, E.; Deudero, S.; Darriba, S.; Carballal, M.J.; Villalba, A. Haplosporidium pinnae sp. nov., a haplosporidan parasite associated with mass mortalities of the fan mussel, Pinna nobilis, in the Western Mediterranean Sea. J. Invertebr. Pathol. 2018, 157, 9–24. [Google Scholar] [CrossRef]
- Darriba, S. First haplosporidan parasite reported infecting a member of the Superfamily Pinnoidea (Pinna nobilis) during a mortality event in Alicante (Spain, Western Mediterranean). J. Invertebr. Pathol. 2017, 148, 14–19. [Google Scholar] [CrossRef]
- Carella, F.; Aceto, S.; Pollaro, F.; Miccio, A.; Iaria, C.; Carrasco, N.; Prado, P.; De Vico, G. A mycobacterial disease is associated with the silent mass mortality of the pen shell Pinna nobilis along the Tyrrhenian coastline of Italy. Sci. Rep. 2019, 9, 2725. [Google Scholar] [CrossRef] [Green Version]
- Lattos, A.; Bitchava, K.; Giantsis, I.A.; Theodorou, J.A.; Batargias, C.; Michaelidis, B. The implication of vibrio bacteria in the winter mortalities of the critically endangered Pinna nobilis. Microorganisms 2021, 9, 922. [Google Scholar] [CrossRef]
- Beja-Pereira, A.; Oliveira, R.; Alves, P.C.; Schwartz, M.K.; Luikart, G. Advancing ecological understandings through technological transformations in noninvasive genetics. Mol. Ecol. Resour. 2009, 9, 1279–1301. [Google Scholar] [CrossRef]
- Sutherland, W.J.; Atkinson, P.W.; Butchart, S.H.M.; Capaja, M.; Dicks, L.V.; Fleishman, E.; Gaston, K.J.; Hails, R.S.; Hughes, A.C.; Le Anstey, B.; et al. A horizon scan of global biological conservation issues for 2022. Trends Ecol. Evol. 2022, 37, 95–104. [Google Scholar] [CrossRef]
- Ruppert, K.M.; Kline, R.J.; Rahman, M.S. Past, present, and future perspectives of environmental DNA (eDNA) metabarcoding: A systematic review in methods, monitoring, and applications of global eDNA. Glob. Ecol. Conserv. 2019, 17, e00547. [Google Scholar] [CrossRef]
- Klymus, K.E.; Merkes, C.M.; Allison, M.J.; Goldberg, C.S.; Helbing, C.C.; Hunter, M.E.; Jackson, C.A.; Lance, R.F.; Mangan, A.M.; Monroe, E.M.; et al. Reporting the limits of detection and quantification for environmental DNA assays. Environ. DNA 2019, 2, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Van Doorn, R.; Klerks, M.M.; Van Gent-Pelzer, M.P.E.; Speksnijder, A.G.C.L.; Kowalchuk, G.A.; Schoen, C.D. Accurate Quantification of Microorganisms in PCR-Inhibiting Environmental DNA Extracts by a Novel Internal Amplification Control Approach Using Biotrove OpenArrays. Appl. Environ. Microbiol. 2009, 75, 7253–7260. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, C.S.; Turner, C.R.; Deiner, K.; Klymus, K.E.; Thomsen, P.F.; Murphy, M.A.; Spear, S.F.; McKee, A.; Oyler-McCance, S.J.; Cornman, R.S.; et al. Critical considerations for the application of environmental DNA methods to detect aquatic species. Methods Ecol. Evol. 2016, 7, 1299–1307. [Google Scholar] [CrossRef]
- Harper, L.R.; Buxton, A.S.; Rees, H.C.; Bruce, K.; Brys, R.; Halfmaerten, D.; Read, D.S.; Watson, H.V.; Sayer, C.D.; Jones, E.P.; et al. Prospects and challenges of environmental DNA (eDNA) monitoring in freshwater ponds. Hydrobiologia 2019, 826, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Manfrin, C.; Ciriaco, S.; Segarich, M.; Fioravanti, P.; Tedesco, P.; Florio, D.; Carella, F.; Giulianini, P.G.; Terlizzi, A.; Pallavicini, A.; et al. First Detection of Haplosporidium pinnae in Pinna nobilis inhabiting the Gulf of Trieste. In Proceedings of the 51th Congress of the Italian Society of Marine Biology, Bari, Italy, 14–17 June 2022; Società Italiana di Biologia Marina: Trieste, Italy, 2022; p. 1. [Google Scholar]
- López-Sanmartín, M.; Catanese, G.; Grau, A.; Valencia, J.M.; García-March, J.R.; Navas, J.I. Real-Time PCR based test for the early diagnosis of Haplosporidium pinnae affecting fan mussel Pinna nobilis. PLoS ONE 2019, 14, e0212028. [Google Scholar] [CrossRef]
- Troth, C.R.; Burian, A.; Mauvisseau, Q.; Bulling, M.; Nightingale, J.; Mauvisseau, C.; Sweet, M.J. Development and application of eDNA-based tools for the conservation of white-clawed crayfish. Sci. Total Environ. 2020, 748, 141394. [Google Scholar] [CrossRef]
- Lopez-Nuñez, R.; Melendreras, E.C.; Casalduero, F.G.; Prado, P.; Lopez-Moya, F.; Lopez-Llorca, L.V. Detection of Haplosporidium pinnae from Pinna nobilis Faeces. J. Mar. Sci. Eng. 2022, 10, 276. [Google Scholar] [CrossRef]
- Beningerl, P.G.; Veniot, A.; Poussart, Y. Principles of pseudofeces rejection on the bivalve mantle: Integration in particle processing. Mar. Ecol. Prog. Ser. 1999, 178, 259–269. [Google Scholar] [CrossRef]
- Deagle, B.E.; Eveson, J.P.; Jarman, S.N. Quantification of damage in DNA recovered from highly degraded samples—A case study on DNA in faeces. Front. Zool. 2006, 3, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
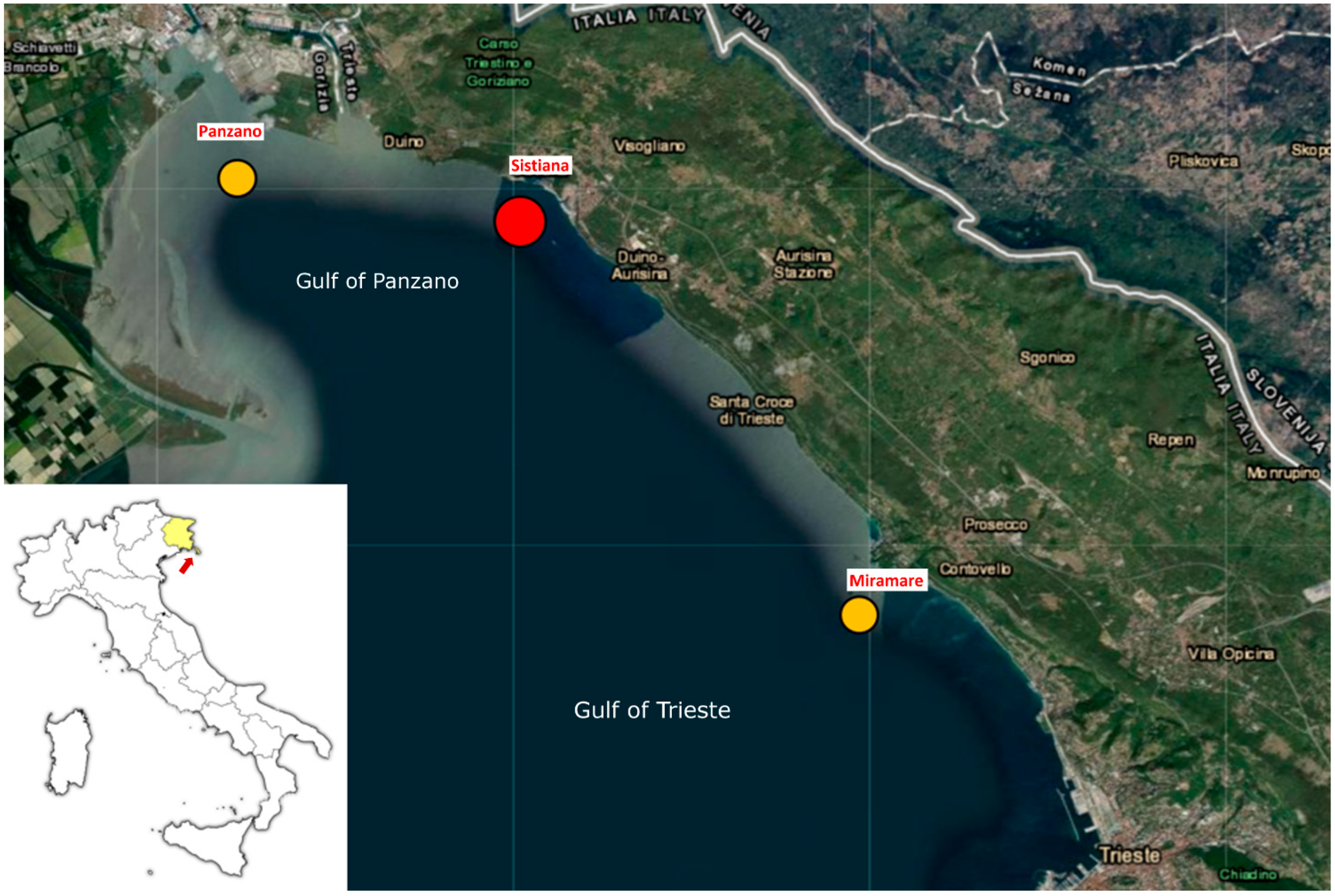
| ID Sample | Type | Collection Date | Place | Geographical Coordinates |
|---|---|---|---|---|
| F1 | Faeces | 12 July 2022 | Miramare | 45.706739 N, 13.703184 E |
| F2 | Faeces | 12 July 2022 | Miramare | 45.706739 N, 13.703184 E |
| F3 | Faeces | 12 July 2022 | Miramare | 45.706739 N, 13.703184 E |
| F4 | Faeces | 22 August 2022 | Panzano | 45.781455 N, 13.548151 E |
| F5 | Faeces | 22 August 2022 | Panzano | 45.781455 N, 13.548151 E |
| F6 | Faeces | 22 August 2022 | Panzano | 45.781455 N, 13.548151 E |
| F7 | Faeces | 5 May 2022 | Sistiana | 45.762826 N, 13.632380 E |
| F8 | Faeces | 5 May 2022 | Sistiana | 45.762826 N, 13.632380 E |
| F9 | Faeces | 5 May 2022 | Sistiana | 45.762826 N, 13.632380 E |
| F10 | Faeces | 5 May 2022 | Sistiana | 45.762826 N, 13.632380 E |
| PS1 | Pseudofaeces | 12 July 2022 | Miramare | 45.706739 N, 13.703184 E |
| PS2 | Pseudofaeces | 12 July 2022 | Miramare | 45.706739 N, 13.703184 E |
| PS3 | Pseudofaeces | 5 May 2022 | Sistiana | 45.762826 N, 13.632380 E |
| PS4 | Pseudofaeces | 22 August 2022 | Panzano | 45.781455 N, 13.548151 E |
| PS5 | Pseudofaeces | 22 August 2022 | Panzano | 45.781455 N, 13.548151 E |
| PS6 | Pseudofaeces | 22 August 2022 | Panzano | 45.781455 N, 13.548151 E |
| PS7 | Pseudofaeces | 22 August 2022 | Panzano | 45.781455 N, 13.548151 E |
| PS8 | Pseudofaeces | 22 August 2022 | Panzano | 45.781455 N, 13.548151 E |
| Name | Sequence (5′->3′) | Temp Melting (°C) |
|---|---|---|
| Hp_CM_For | TGACTCAACACGGGGAAAC | 59.51 |
| HP-CM_probe | FAM-CCAGGGCCAGACATAGCCAGGA- BHQ1 | 69.74 |
| Hp_CM_Rev | ACTAAGAACGGCCATGCAC | 58.74 |
| Name | Sequence (5′->3′) | Temp Melting (°C) |
|---|---|---|
| IAC 1_For | CCTCTGCAGCGATGTCACTA | 60.16 |
| IAC 1_probe | HEX- CCCGTCTAACACACCTGTCCA- BHQ1 | 63.26 |
| IAC 1_Rev | GCATCTGTCGGTGCGTCAAT | 64.54 |
| Sample | Replicates | |||||||
|---|---|---|---|---|---|---|---|---|
| Ct | Ct | Ct | Ct | Ct | Ct | Ct Mean | ±SD | |
| F8 | 22.01 | 22.91 | 22.87 | 22.58 | 22.72 | 23.05 | 22.69 | 0.37 |
| Positive control | 25.41 | 22.05 | 23 | 23.47 | 1.73 | |||
| IAC1 (3 × 102) | 7.61 | 7.41 | 7 | 7.34 | 0.31 | |||
| IAC1 (3 × 10) + F8 | 7.85 | 7.81 | 7.83 | 0.03 | ||||
| IAC1 (3 × 102) | 11.31 | 10.94 | 10.87 | 11.04 | 0.24 | |||
| IAC1 (3 × 102) + F8 | 11.89 | 12.04 | 11.96 | 0.11 | ||||
| IAC1 (3 × 102) + F6 | 11.85 | 11.51 | 11.68 | 0.24 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manfrin, C.; Ciriaco, S.; Segarich, M.; Aiello, A.; Florian, F.; Avian, M.; Terlizzi, A.; Giulianini, P.G.; Spoto, M.; Pallavicini, A. Haplosporidium pinnae Detection from the Faeces of Pinna nobilis: A Quick and Noninvasive Tool to Monitor the Presence of Pathogen in Early-Stage or during Fan Mussel Mass Mortalities. Diversity 2023, 15, 477. https://doi.org/10.3390/d15040477
Manfrin C, Ciriaco S, Segarich M, Aiello A, Florian F, Avian M, Terlizzi A, Giulianini PG, Spoto M, Pallavicini A. Haplosporidium pinnae Detection from the Faeces of Pinna nobilis: A Quick and Noninvasive Tool to Monitor the Presence of Pathogen in Early-Stage or during Fan Mussel Mass Mortalities. Diversity. 2023; 15(4):477. https://doi.org/10.3390/d15040477
Chicago/Turabian StyleManfrin, Chiara, Saul Ciriaco, Marco Segarich, Andrea Aiello, Fiorella Florian, Massimo Avian, Antonio Terlizzi, Piero G. Giulianini, Maurizio Spoto, and Alberto Pallavicini. 2023. "Haplosporidium pinnae Detection from the Faeces of Pinna nobilis: A Quick and Noninvasive Tool to Monitor the Presence of Pathogen in Early-Stage or during Fan Mussel Mass Mortalities" Diversity 15, no. 4: 477. https://doi.org/10.3390/d15040477