

Distribution of Non-Indigenous Crayfish Species in Estonia and Their Impacts on Noble Crayfish (Astacus astacus L.) Populations

Abstract
:1. Introduction
2. Materials and Methods
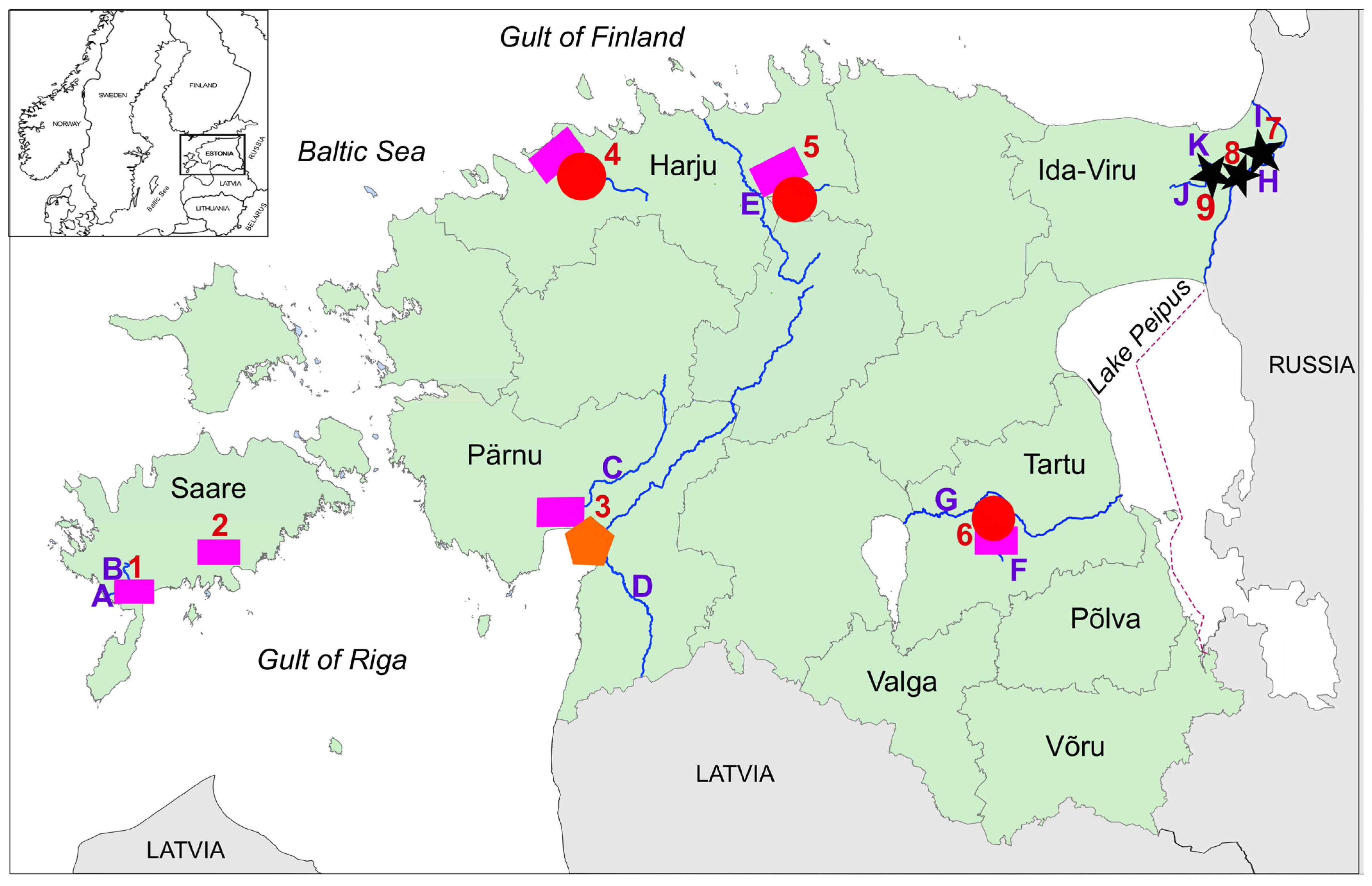
2.1. Study Area
2.2. Test Trapping of Crayfish
2.3. Data Analysis
3. Results and Discussion
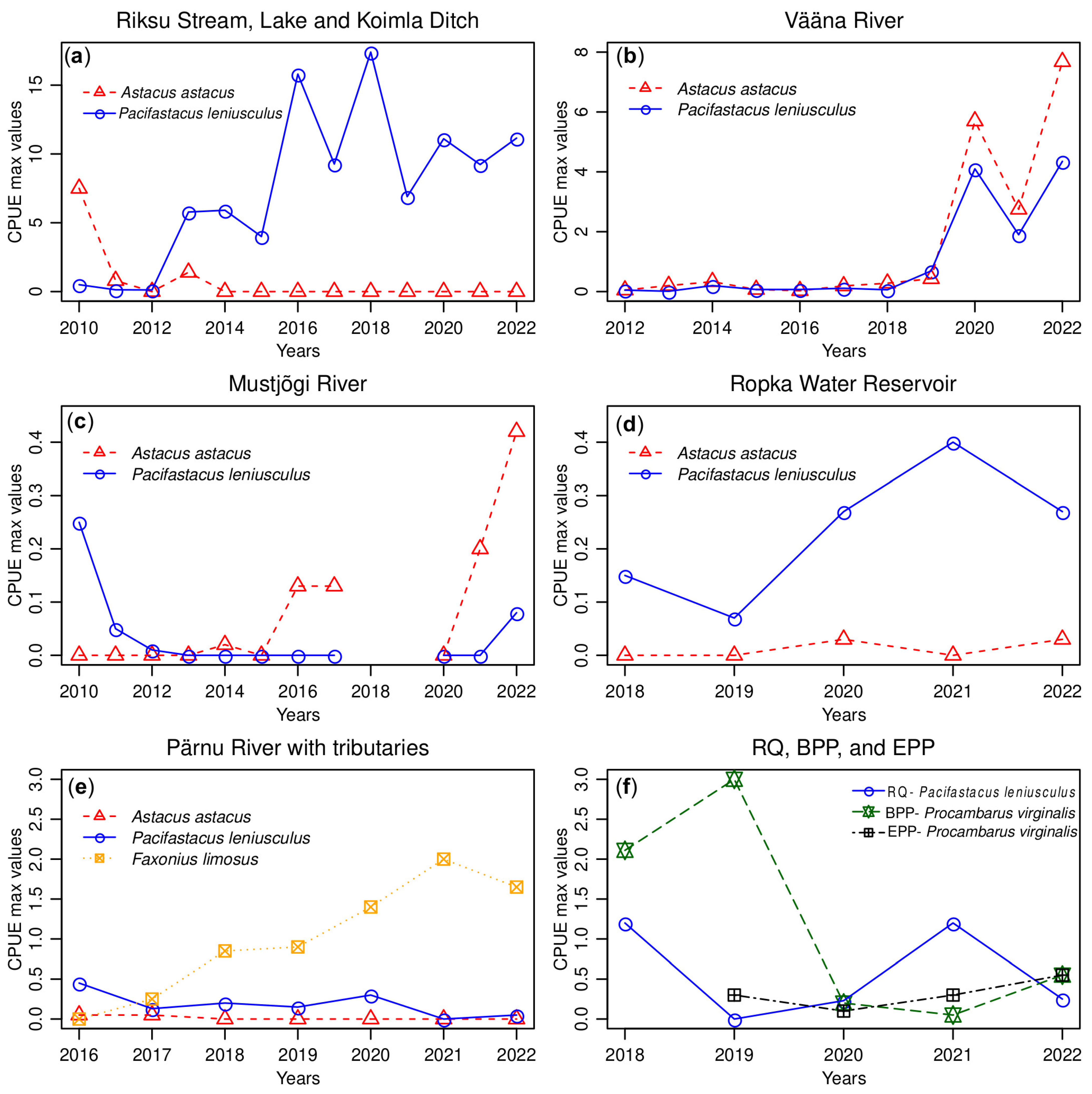
3.1. Riksu Stream
3.2. Vääna River
3.3. Mustjõgi River
3.4. Ropka Water Reservoir
3.5. Pärnu River
3.6. Reo Quarry
3.7. Estonian and Baltic Power Plant Outflow Channels
3.8. General Observations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gross, R.; Palm, S.; Kõiv, K.; Prestegaard, T.; Jussila, J.; Paaver, T.; Geist, J.; Kokko, H.; Karjalainen, A.; Edsman, L. Microsatellite Markers Reveal Clear Geographic Structuring among Threatened Noble Crayfish (Astacus astacus) Populations in Northern and Central Europe. Conserv. Genet. 2013, 14, 809–821. [Google Scholar] [CrossRef]
- Hurt, M.; Kivistik, M. Tegevuskava Rakendamine Jõevähi Varude Kasutamiseks Ja Kaitseks 2021.a.—Keskkonnaameti Tellitud (Riigihanke Viitenumber 236237) Ja Eesti Maaülikooli Teostatud Projekti Aruanne.107 Lk. 2022. Available online: https://envir.ee/media/4550/download (accessed on 7 February 2023).
- Tulonen, J.; Erkamo, E.; Järvenpää, T.; Westman, K.; Savolainen, R.; Mannonen, A. Rapuvedet Tuottaviksi; Riistan-ja kalantutkimuslaitos: Helsinki, Finland, 1998; 152p. [Google Scholar]
- Tulonen, J.; Erkamo, E.; Jussila, J.; Mannonen, A. The Effects of Minimum Size Regulations and Exploitation on Population Dynamics of Noble Crayfish (Astacus astacus (Linnaeus)) in a Small Lake in Central Finland: A Seven Year Study. Freshw. Crayfish 2008, 16, 7–14. [Google Scholar]
- Tuusti, J.; Taugbøl, T.; Skurdal, J.; Kukk, L. Freshwater Crayfish in Estonia. I: Action Plan for Crayfish Management. II: Crayfish Status Report; Østlandsforskning: Oppland, Finland, 1998. [Google Scholar]
- Reynolds, J.D. A Review of Ecological Interactions between Crayfish and Fish, Indigenous and Introduced. Knowl. Manag. Aquat. Ecosyst. 2011, 401, 10p1–10p21. [Google Scholar] [CrossRef] [Green Version]
- Souty-Grosset, C.; Holdich, D.M.; Noel, P.Y.; Reynolds, J.D.; Haffner, P. Atlas of Crayfish in Europe; Muséum National d´Histoire naturelle: Paris, France, 2006; pp. 60–123. [Google Scholar]
- Holdich, D.M.; Reynolds, J.D.; Souty-Grosset, C.; Sibley, P.J. A Review of the Ever Increasing Threat to European Crayfish from Non-Indigenous Crayfish Species. Knowl. Manag. Aquat. Ecosyst. 2009, 394–395, 11p1–11p46. [Google Scholar] [CrossRef] [Green Version]
- Diéguez-Uribeondo, J.; Söderhäll, K. Procambarus clarkii Girard as a Vector for the Crayfish Plague Fungus, Aphanomyces astaci Schikora. Aquac. Res. 2008, 24, 761–765. [Google Scholar] [CrossRef]
- Kozubíková, E.; Viljamaa-Dirks, S.; Heinikainen, S.; Petrusek, A. Spiny-Cheek Crayfish Orconectes limosus Carry a Novel Genotype of the Crayfish Plague Pathogen Aphanomyces astaci. J. Invertebr. Pathol. 2011, 108, 214–216. [Google Scholar] [CrossRef]
- Keller, N.S.; Pfeiffer, M.; Roessink, I.; Schulz, R.; Schrimpf, A. First Evidence of Crayfish Plague Agent in Populations of the Marbled Crayfish (Procambarus fallax Forma virginalis). Knowl. Manag. Aquat. Ecosyst. 2014, 414, 15p1–15p8. [Google Scholar]
- Alderman, D.J. Crayfish Plague in Britain, the First Twelve Years. Freshw. Crayfish 1993, 9, 266–272. [Google Scholar]
- Alderman, D.J. History of the Spread of Crayfish Plague in Europe, in Crustaceans: Bacterial and Fungal Diseases. QIE Sci. Technol. Rev. 1997, 15, 15–23. [Google Scholar]
- Taugbøl, T.; Skurdal, J.; Håstein, T. Crayfish Plague and Management Strategies in Norway. Biol. Conserv. 1993, 63, 75–82. [Google Scholar] [CrossRef]
- Diéguez-Uribeondo, J.; Temiño, C.; Mùzquiz, J.L. The Crayfish Plague Fungus (Aphanomyces astaci) in Spain. Bull. Fr. Pêche Piscic. 1997, 347, 753–763. [Google Scholar] [CrossRef] [Green Version]
- Mirimin, L.; Brady, D.; Gammell, M.; Lally, H.; Minto, C.; Graham, C.T.; Slattery, O.; Cheslett, D.; Morrissey, T.; Reynolds, J.; et al. Investigation of the First Recent Crayfish Plague Outbreak in Ireland and Its Subsequent Spread in the Bruskey River and Surrounding Areas. Knowl. Manag. Aquat. Ecosyst. 2022, 423, 13. [Google Scholar] [CrossRef]
- Kaldre, K.; Paaver, T.; Hurt, M.; Grandjean, F. First Records of the Non-Indigenous Signal Crayfish (Pacifastacus leniusculus) and Its Threat to Noble Crayfish (Astacus astacus) Populations in Estonia. Biol. Invasions 2017, 19, 2771–2776. [Google Scholar] [CrossRef]
- Ercoli, F.; Kaldre, K.; Paaver, T.; Gross, R. First Record of an Established Marbled Crayfish Procambarus virginalis (Lyko, 2017) Population in Estonia. Bioinvasions Rec. 2019, 8, 675–683. [Google Scholar] [CrossRef]
- Kaldre, K.; Paaver, T.; Hurt, M.; Gross, R. Continuing Expansion of Non-Indigenous Crayfish Species in Northern Europe: First Established Spiny-Cheek Crayfish Faxonius limosus (Refinesque, 1817) Population in Estonia. BioInvasions Rec. 2020, 9, 127–132. [Google Scholar] [CrossRef]
- Martin, P.; Kohlmann, K.; Scholtz, G. The Parthenogenetic Marmorkrebs (Marbled Crayfish) Produces Genetically Uniform Offspring. Naturwissenschaften 2007, 94, 843–846. [Google Scholar] [CrossRef] [PubMed]
- Kaldre, K.; Meženin, A.; Paaver, T.; Kawai, T. A Preliminary Study on the Tolerance of Marble Crayfish Procambarus fallax f. virginalis to Low Temperature in Nordic Climate. In Freshwater Crayfish: A Global Overview; Taylor & Francis: Boca Raton, FL, USA, 2015; pp. 54–62. [Google Scholar]
- Lipták, B.; Veselý, L.; Ercoli, F.; Bláha, M.; Buřič, M.; Ruokonen, T.; Kouba, A. Trophic Role of Marbled Crayfish in a Lentic Freshwater Ecosystem. Aquat. Invasions 2019, 14, 299–309. [Google Scholar] [CrossRef] [Green Version]
- Kozák, P.; Buřič, M.; Policar, T. The Fecundity, Time of Egg Development and Juvenile Production in Spiny-Cheek Crayfish (Orconectes limosus) under Controlled Conditions. Bull. Fr. Pêche Piscic. 2006, 380–381, 1171–1182. [Google Scholar] [CrossRef] [Green Version]
- Jaszczołt, J.; Szaniawska, A. The Spiny-Cheek Crayfish Orconectes limosus (Rafinesque, 1817) as an Inhabitant of the Baltic Sea—Experimental Evidences for Its Invasion of Brackish Waters. Oceanol. Hydrobiol. Stud. 2011, 40, 52–60. [Google Scholar] [CrossRef]
- Bohmann, K.; Evans, A.; Gilbert, M.T.P.; Carvalho, G.R.; Creer, S.; Knapp, M.; Yu, D.W.; de Bruyn, M. Environmental DNA for Wildlife Biology and Biodiversity Monitoring. Trends Ecol. Evol. 2014, 29, 358–367. [Google Scholar] [CrossRef]
- Strand, D.A.; Jussila, J.; Johnsen, S.I.; Viljamaa-Dirks, S.; Edsman, L.; Wiik-Nielsen, J.; Viljugrein, H.; Engdahl, F.; Vrålstad, T. Detection of Crayfish Plague Spores in Large Freshwater Systems. J. Appl. Ecol. 2014, 51, 544–553. [Google Scholar] [CrossRef] [Green Version]
- Dougherty, M.M.; Larson, E.R.; Renshaw, M.A.; Gantz, C.A.; Egan, S.P.; Erickson, D.M.; Lodge, D.M. Environmental DNA (EDNA) Detects the Invasive Rusty Crayfish Orconectes rusticus at Low Abundances. J. Appl. Ecol. 2016, 53, 722–732. [Google Scholar] [CrossRef] [Green Version]
- Wittwer, C.; Stoll, S.; Strand, D.; Vrålstad, T.; Nowak, C.; Thines, M. EDNA-Based Crayfish Plague Monitoring Is Superior to Conventional Trap-Based Assessments in Year-Round Detection Probability. Hydrobiologia 2018, 807, 87–97. [Google Scholar]
- Strand, D.A.; Johnsen, S.I.; Rusch, J.C.; Agersnap, S.; Larsen, W.B.; Knudsen, S.W.; Møller, P.R.; Vrålstad, T. Monitoring a Norwegian freshwater crayfish tragedy: eDNA snapshots of invasion, infection and extinction. J. Appl. Ecol. 2019, 56, 1661–1673. [Google Scholar] [CrossRef]
- Rusch, J.C.; Mojžišová, M.; Strand, D.A.; Svobodová, J.; Vrålstad, T.; Petrusek, A. Simultaneous Detection of Native and Invasive Crayfish and Aphanomyces astaci from Environmental DNA Samples in a Wide Range of Habitats in Central Europe. NeoBiota 2020, 58, 1–32. [Google Scholar] [CrossRef]
- Grandjean, F.; Roques, J.; Delaunay, C.; Petrusek, E.; Becking, T.; Collas, M. Status of Pacifastacus leniusculus and Its Role in Recent Crayfish Plague Outbreaks in France: Improving Distribution and Crayfish Plague Infection Patterns. Aquat. Invasions 2017, 12, 541–549. [Google Scholar] [CrossRef]
- Dragičević, P.; Faller, M.; Kutlesa, P.; Hudina, S. Update on the Signal Crayfish, Pacifastacus leniusculus (Dana, 1852) Range Expansion in Croatia: A 10-Year Report. BioInvasions Rec. 2020, 9, 793–807. [Google Scholar] [CrossRef]
- Hudina, S.; Žganec, K.; Lucić, A.; Trgovčić, K.; Maguire, I. Recent Invasion of the Karstic River Systems in Croatia through Illegal Introductions of the Signal Crayfish. Freshw. Crayfish 2013, 19, 21–27. [Google Scholar] [CrossRef]
- Hudina, S.; Kutleša, P.; Trgovčić, K. Duplić Dynamics of Range Expansion of the Signal Crayfish (Pacifastacus leniusculus) in a Recently Invaded Region in Croatia. Aquat. Invasions 2017, 12, 67–75. [Google Scholar] [CrossRef]
- Maguire, I.; Jelić, M.; Klobučar, G. Update on the Distribution of Freshwater Crayfish in Croatia. Knowl. Manag. Aquat. Ecosyst. 2011, 401, 31p1–31p10. [Google Scholar] [CrossRef]
- Dobrzycka-Krahel, A.; Skóra, M.E.; Raczyński, M.; Szaniawska, A. The Signal Crayfish Pacifastacus leniusculus—Distribution and Invasion in the Southern Baltic Coastal River. Pol. J. Ecol. 2017, 65, 445–452. [Google Scholar]
- Weinländer, M.; Füreder, L. The Continuing Spread of Pacifastacus leniusculus in Carinthia (Austria). Knowl. Manag. Aquat. Ecosyst. 2009, 394–395, 17p1–17p10. [Google Scholar]
- Birzaks, J.; Skute, A. Alien Crayfish Species in Latvian Inland Waters. Environ. Exp. Biol. 2019, 17, 21–25. [Google Scholar]
- Hein, C.L.; Vander Zanden, M.J.; Magnuson, J.J. Intensive Trapping and Increased Fish Predation Cause Massive Population Decline of an Invasive Crayfish. Freshw. Biol. 2007, 52, 1134–1146. [Google Scholar] [CrossRef]
- Moorhouse, T.P.; Poole, A.E.; Evans, L.C.; Bradley, D.C.; Macdonald, D.W. Intensive Removal of Signal Crayfish (Pacifastacus leniusculus) from Rivers Increases Numbers and Taxon Richness of Macroinvertebrate Species. Ecol. Evol. 2014, 4, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Moorhouse, T.P.; Macdonald, D.W. The Effect of Manual Removal on Movement Distances in Populations of Signal Crayfish (Pacifastacus leniusculus). Freshw. Biol. 2011, 56, 2370–2377. [Google Scholar] [CrossRef]
- Stancliffe-Vaughan, A.E. Sampling UK Pacifastacus leniusculus (Dana, 1852): The Effect of Trapping on Population Structure. Master’s Thesis, Anglia Ruskin University, Cambridge, UK, 2015. [Google Scholar]
- Loureiro, T.G.; Anastácio, P.M.; Bueno, S.L.d.S.; Araujo, P.B. Management of Invasive Populations of the Freshwater Crayfish Procambarus clarkii (Decapoda, Cambaridae): Test of a Population-Control Method and Proposal of a Standard Monitoring Approach. Environ. Monit. Assess. 2018, 190, 559. [Google Scholar] [CrossRef]
- Skov, C.; Aarestrup, K.; Sivebæk, F.; Pedersen, S.; Vrålstad, T.; Berg, S. Non-Indigenous Signal Crayfish Pacifastacus leniusculus Are Now Common in Danish Streams: Preliminary Status for National Distribution and Protective Actions. Biol. Invasions 2011, 13, 1269–1274. [Google Scholar] [CrossRef]
- Hudina, S.; Hock, K.; Radović, A.; Klobučar, G.; Petković, J.; Jelić, M.; Maguire, I. Species-Specific Differences in Dynamics of Agonistic Interactions May Contribute to the Competitive Advantage of the Invasive Signal Crayfish (Pacifastacus leniusculus) over the Native Narrow-Clawed Crayfish (Astacus leptodactylus). Mar. Freshw. Behav. Physiol. 2016, 49, 147–157. [Google Scholar] [CrossRef]
- Estonian Nature Information System Display of Water Body Data. Available online: https://eelis.ee/default.aspx?state=15;572247461;est;eelisand;;&comp=objresult=veekogu&obj_id=268239962 (accessed on 31 January 2023).
- Edsman, L.; Söderbäck, B. Standardised Sampling Methodology for Crayfish—The Swedish Protocol. Freshw. Crayfish 1999, 12, 705–713. [Google Scholar]
- Cheng, Z.; Folkins, M.H.; Huang, L.; Li, Y. Comparing Catch Efficiency of Crayfish (Procambarus clarkii) Traps with Different Entrance Numbers. J. Mar. Sci. Eng. 2022, 10, 1708. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 7 February 2023).
- Manfrin, C.; Souty-Grosset, C.; Anastácio, P.M.; Reynolds, J.; Giulianini, P.G. Detection and Control of Invasive Freshwater Crayfish: From Traditional to Innovative Methods. Diversity 2019, 11, 5. [Google Scholar] [CrossRef] [Green Version]
- Hudina, S.; Maguire, I.; Dragičević, P.; Galic, N. Evaluating the Efficacy of Approaches to Control Invasive Populations: A Conceptual Model Development for the Signal Crayfish. Ecologies 2022, 3, 78–95. [Google Scholar] [CrossRef]
- Eesti Energia Environmental Report. Available online: https://www.energia.ee/c/document_library/get_file?uuid=e704c22c-c190-469b-9e3e-f2a9bf95b777&groupId=10187 (accessed on 3 February 2023).
- Olsson, K.; Stenroth, P.; Nyström, P.; Granéli, W. Invasions and Niche Width: Does Niche Width of an Introduced Crayfish Differ from a Native Crayfish? Freshw. Biol. 2009, 54, 1731–1740. [Google Scholar] [CrossRef]
- Westman, K.; Savolainen, R. Long Term Study of Competition between Two Co-Occurring Crayfish Species, the Native Astacus astacus L. and the Introduced Pacifastacus leniusculus Dana, in a Finnish Lake. Bull. Fr. Pêche Piscic. 2001, 361, 613–627. [Google Scholar] [CrossRef] [Green Version]
- Ercoli, F.; Ghia, D.; Gruppuso, L.; Fea, G.; Bo, T.; Ruokonen, T.J. Diet and Trophic Niche of the Invasive Signal Crayfish in the First Invaded Italian Stream Ecosystem. Sci. Rep. 2021, 11, 8704. [Google Scholar] [CrossRef]
- Johnson, M.F.; Rice, S.P.; Reid, I. The Activity of Signal Crayfish (Pacifastacus leniusculus) in Relation to Thermal and Hydraulic Dynamics of an Alluvial Stream, UK. Hydrobiologia 2014, 724, 41–54. [Google Scholar] [CrossRef] [Green Version]
- Mathers, K.L.; White, J.C.; Fornaroli, R.; Chadd, R. Flow Regimes Control the Establishment of Invasive Crayfish and Alter Their Effects on Lotic Macroinvertebrate Communities. J. Appl. Ecol. 2020, 57, 886–902. [Google Scholar] [CrossRef]
- Larson, C.E.; Bo, T.; Candiotto, A.; Fenoglio, S.; Doretto, A. Predicting Invasive Signal Crayfish (Pacifastacus leniusculus) Spread Using a Traditional Survey and River Network Simulation. River Res. Appl. 2022, 38, 1424–1435. [Google Scholar] [CrossRef]
- García-de-Lomas, J.; Dana, E.D.; González, R. Traps and Netting, Better Together than Alone: An Innovative Approach to Improve Procambarus clarkii Management. Knowl. Manag. Aquat. Ecosyst. 2020, 421, 39. [Google Scholar] [CrossRef]
- Nyström, P.; Stenroth, P.; Holmqvist, N.; Berglund, O.; Larsson, P.; Granéli, W. Crayfish in Lakes and Streams: Individual and Population Responses to Predation, Productivity and Substratum Availability. Freshw. Biol. 2006, 51, 2096–2113. [Google Scholar] [CrossRef]
- Schrimpf, A.; Maiwald, T.; Vrålstad, T.; Schulz, H.K.; Śmietana, P.; Schulz, R. Absence of the Crayfish Plague Pathogen (Aphanomyces astaci) Facilitates Coexistence of European and American Crayfish in Central Europe. Freshw. Biol. 2013, 58, 1116–1125. [Google Scholar] [CrossRef]
- James, J.; Nutbeam-Tuffs, S.; Cable, J.; Mrugała, A.; Viñuela-Rodriguez, N.; Petrusek, A.; Oidtmann, B. The Prevalence of Aphanomyces astaci in Invasive Signal Crayfish from the UK and Implications for Native Crayfish Conservation. Parasitology 2017, 144, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Vanni, M.J.; Duncan, J.M.; González, M.J.; Horgan, M.J. Competition Among Aquatic Organisms. In Encyclopedia of Inland Waters; Likens, G.E., Ed.; Academic Press: Oxford, UK, 2009; pp. 395–404. [Google Scholar]
- Söderbäck, B. Interactions among Juveniles of Two Freshwater Crayfish Species and a Predatory Fish. Oecologia 1994, 100, 229–235. [Google Scholar] [CrossRef]
- Schulz, H.; Śmietana, P.; Maiwald, T.; Oidtmann, B.; Schulz, R. Case Studies on the Co-Occurrence of Astacus astacus (L.) and Orconectes limosus (Raf.): Snapshots of a Slow Displacement. Freshw. Crayfish 2006, 15, 212–219. [Google Scholar]
- Paaver, T.; Hurt, M. Status and Management of Noble Crayfish Astacus astacus in Estonia. Knowl. Manag. Aquat. Ecosyst. 2009, 394–395, 18. [Google Scholar] [CrossRef] [Green Version]
- Jussila, J.; Edsman, L. Relaxed Attitude towards Spreading of Alien Crayfish Species Affects Protection of Native Crayfish Species: Case Studies and Lessons Learnt from a Fennoscandian Viewpoint. Freshw. Crayfish 2020, 25, 39–46. [Google Scholar] [CrossRef]
- Krzywosz, T. Evaluation of Changes in the Abundance of Three Catchable Crayfish Species in Lake Pobłędzie (Northern Poland). Fish. Aquat. Life 2006, 14, 131–140. [Google Scholar]
- Gherardi, F.; Aquiloni, L.; Diéguez-Uribeondo, J.; Tricarico, E. Managing Invasive Crayfish: Is There a Hope? Aquat. Sci. 2011, 73, 185–200. [Google Scholar] [CrossRef]
- Jimenez, S.A.; Faulkes, Z. Establishment and Care of a Colony of Parthenogenetic Marbled Crayfish, Marmorkrebs. Invertebr. Rearing 2010, 1, 10–18. [Google Scholar]
- Linzmaier, S.M.; Goebel, L.S.; Ruland, F.; Jeschke, J.M. Behavioral Differences in an Over-Invasion Scenario: Marbled vs. Spiny-Cheek Crayfish. Ecosphere 2018, 9, e02385. [Google Scholar] [CrossRef]
- Vogt, G. The Marbled Crayfish: A New Model Organism for Research on Development, Epigenetics and Evolutionary Biology. J. Zool. 2008, 276, 1–13. [Google Scholar] [CrossRef]
- Sibley, P.; Noel, P. Control and Management of Alien Crayfish. Bull. Fr. Pêche Piscic. 2002, 367, 881–886. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| No | Sampling Locations | Descriptions * |
|---|---|---|
| 1 | Riksu Stream (with Riksu Lake and Koimla Ditch) |
|
| 2 | Reo Quarry (RQ) |
|
| 3 | Pärnu River with tributaries |
|
| 4 | Vääna River |
|
| 5 | Mustjõgi River |
|
| 6 | Ropka Water Reservoir |
|
| 7 | Baltic Power Plant (BPP) outflow channel and Narva Water Reservoir |
|
| 8 | Estonian Power Plant (EPP) outflow channel and Narva River |
|
| 9 | Narva Quarry (NQ) with outlet and Metsküla Stream |
|
| No | Sampling Locations | Number of Trap Nights per Year | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | ||
| 1 | Riksu Stream | 465 | 1430 | 440 | 754 | 238 | 514 | 310 | 250 | 205 | 230 | 232 | 310 | 3000 |
| 2 | Reo Quarry | NA | NA | NA | NA | NA | NA | NA | NA | 10 | NA | 30 | 5 | 2600 |
| 3 | Pärnu River | NA | NA | NA | NA | NA | NA | 120 | 240 | 460 | 405 | 340 | 160 | 620 |
| 4 | Vääna River | NA | NA | 20 | 240 | 160 | 180 | 160 | 80 | 80 | 80 | 140 | 80 | 960 |
| 5 | Mustjõgi River | 40 | 320 | 200 | 310 | 100 | 80 | 40 | 40 | NA | NA | 40 | 40 | 460 |
| 6 | Ropka Water Reservoir | NA | NA | NA | NA | NA | NA | NA | NA | 70 | 60 | 110 | 40 | 1120 |
| 7 | BPP outlet | NA | NA | NA | NA | NA | NA | NA | NA | 197 | 292 | 260 | 200 | 400 |
| 8 | EPP outlet | NA | NA | NA | NA | NA | NA | NA | NA | NA | 150 | 200 | 80 | 440 |
| 9 | NQ | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | 130 | 110 |
| Crayfish Species per Sampling Location | CPUE Values per Year | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | p-Value | |
| Riksu Stream, Lake and Koimla Ditch | ||||||||||||||
| A. astacus | <0.1 | <0.1 | 0 | <0.1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| P. leniusculus | 0.1 | <0.1 | 0.1 | 0.2 | 1.1 | 1.4 | 3.3 | 3.1 | 6.3 | 4.0 | 6.9 | 2.8 | 1.9 | *** |
| Vääna River | ||||||||||||||
| A. astacus | NA | NA | 0.1 | 0.1 | 0.1 | <0.1 | <0.1 | 0.2 | 0.3 | 0.4 | 2.0 | 2.2 | 1.8 | |
| P. leniusculus | NA | NA | 0.1 | <0.1 | 0.1 | <0.1 | <0.1 | 0.1 | 0.1 | 0.7 | 0.7 | 0.4 | 1.6 | n.s. |
| Mustjõgi River | ||||||||||||||
| A. astacus | 0 | 0 | 0 | 0 | <0.1 | 0 | 0.1 | 0.1 | NA | NA | 0 | 0.1 | 0.1 | |
| P. leniusculus | 0.1 | <0.1 | <0.1 | 0 | 0 | 0 | 0 | 0 | NA | NA | 0 | 0 | <0.1 | n.s. |
| Ropka Water Reservoir | ||||||||||||||
| A. astacus | NA | NA | NA | NA | NA | NA | NA | NA | 0 | 0 | <0.1 | 0 | <0.1 | |
| P. leniusculus | NA | NA | NA | NA | NA | NA | NA | NA | 0.1 | <0.1 | 0.1 | 0.3 | 0.1 | ** |
| Pärnu River with tributaries | ||||||||||||||
| A. astacus | NA | NA | NA | NA | NA | NA | <0.1 | <0.1 | 0 | 0 | 0 | 0 | 0 | |
| P. leniusculus | NA | NA | NA | NA | NA | NA | 0.1 | <0.1 | <0.1 | <0.1 | <0.1 | 0 | <0.1 | ** |
| F. limosus | NA | NA | NA | NA | NA | NA | 0 | <0.1 | 0.1 | 0.2 | 0.4 | 0.5 | 0.2 | ** |
| Reo Quarry | ||||||||||||||
| P. leniusculus | NA | NA | NA | NA | NA | NA | NA | NA | 1.2 | 0 | 0.2 | 1.2 | 0.1 | - |
| Baltic Power Plant outflow channel and Narva Water Reservoir | ||||||||||||||
| Procambarus virginalis | NA | NA | NA | NA | NA | NA | NA | NA | 0.5 | 0.9 | <0.1 | <0.1 | 0.1 | - |
| Estonian Power Plant outflow channel and Narva River | ||||||||||||||
| Procambarus virginalis | NA | NA | NA | NA | NA | NA | NA | NA | NA | <0.1 | <0.1 | 0.2 | 0.1 | - |
| Narva Quarry with outlet and Metsküla Stream | ||||||||||||||
| A. astacus | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | 0.1 | 0 | - |
| Procambarus virginalis | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | <0.1 | <0.1 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aluma, M.O.; Pukk, L.; Hurt, M.; Kaldre, K. Distribution of Non-Indigenous Crayfish Species in Estonia and Their Impacts on Noble Crayfish (Astacus astacus L.) Populations. Diversity 2023, 15, 474. https://doi.org/10.3390/d15040474
Aluma MO, Pukk L, Hurt M, Kaldre K. Distribution of Non-Indigenous Crayfish Species in Estonia and Their Impacts on Noble Crayfish (Astacus astacus L.) Populations. Diversity. 2023; 15(4):474. https://doi.org/10.3390/d15040474
Chicago/Turabian StyleAluma, Michael Oliewo, Lilian Pukk, Margo Hurt, and Katrin Kaldre. 2023. "Distribution of Non-Indigenous Crayfish Species in Estonia and Their Impacts on Noble Crayfish (Astacus astacus L.) Populations" Diversity 15, no. 4: 474. https://doi.org/10.3390/d15040474