


Ionome of Lithuanian Populations of Purple Loosestrife (Lythrum salicaria) and Its Relation to Genetic Diversity and Environmental Variables
 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling and Element Analyses
2.3. Classification of Environment
2.4. Data Analysis
3. Results
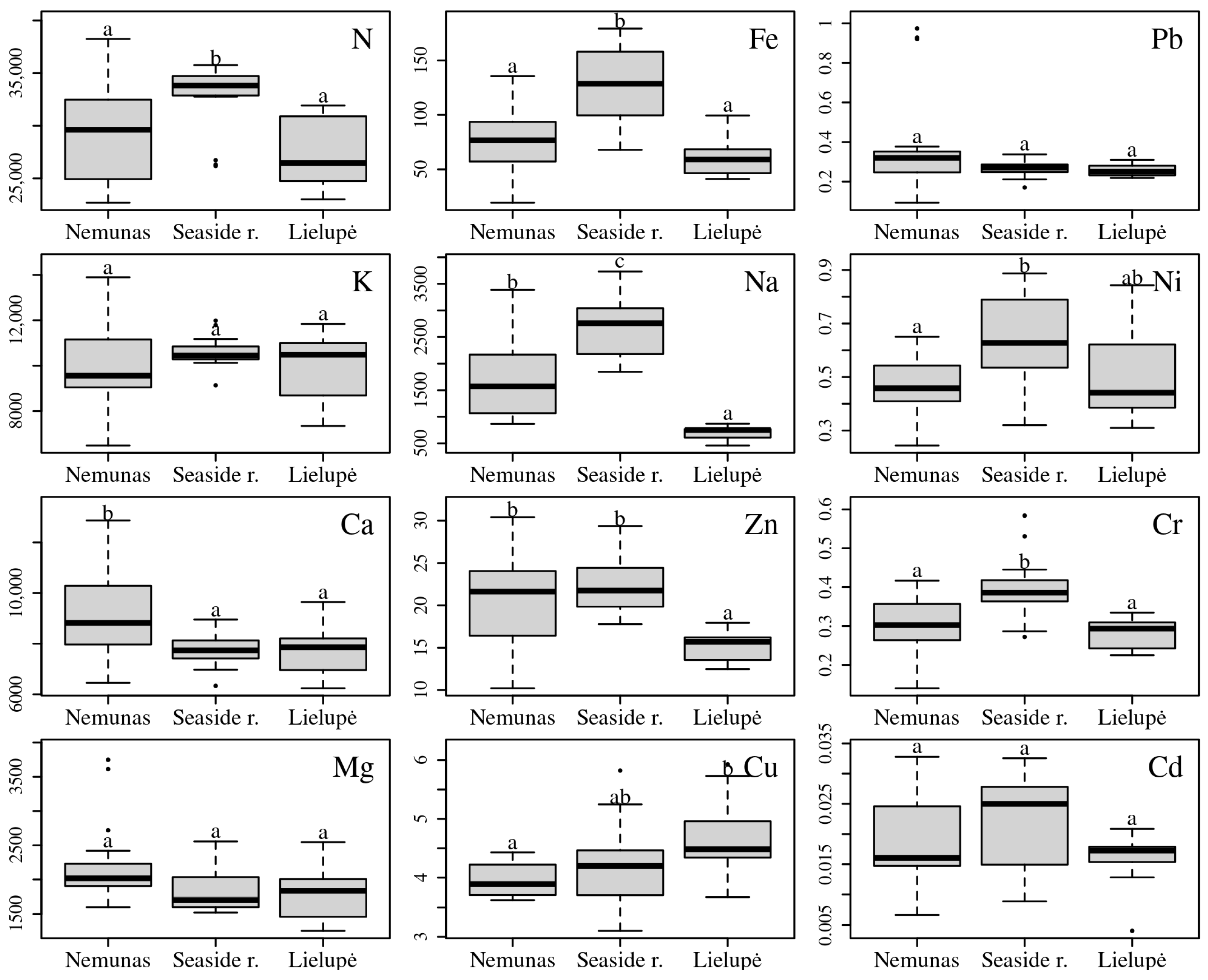
3.1. Comparison of Populations Based on Elemental Concentrations
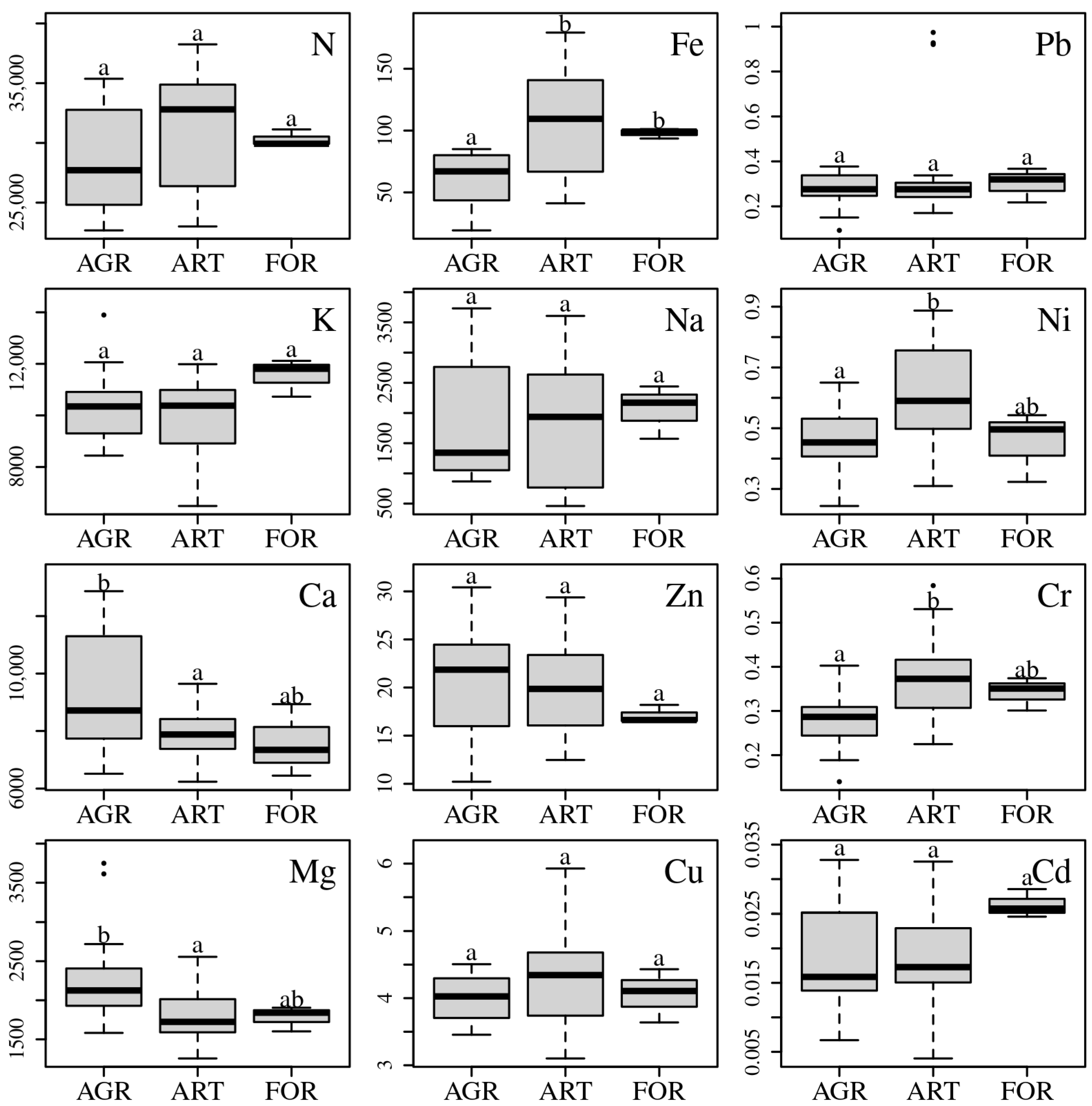
3.2. Comparison of Groups of Populations
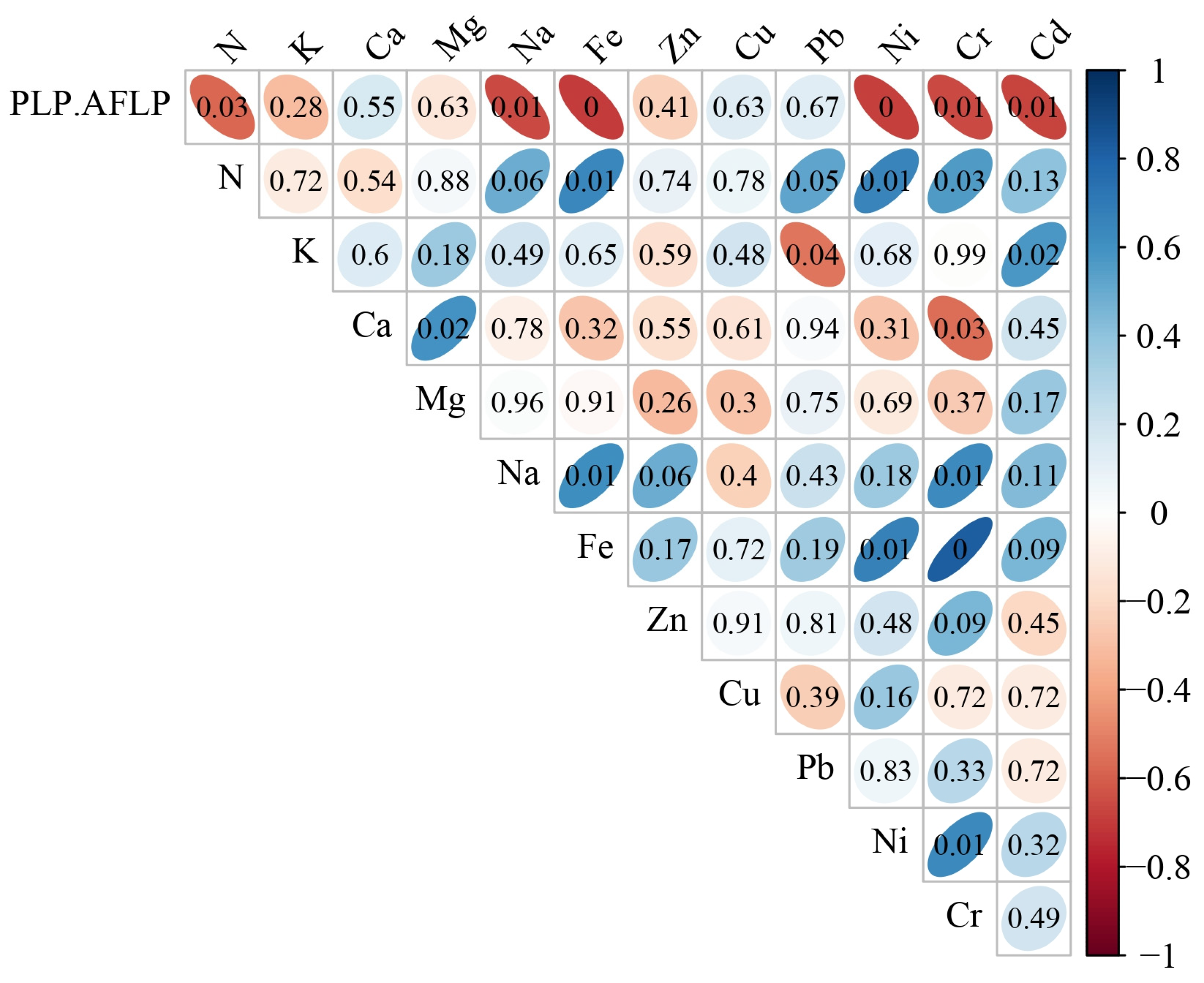
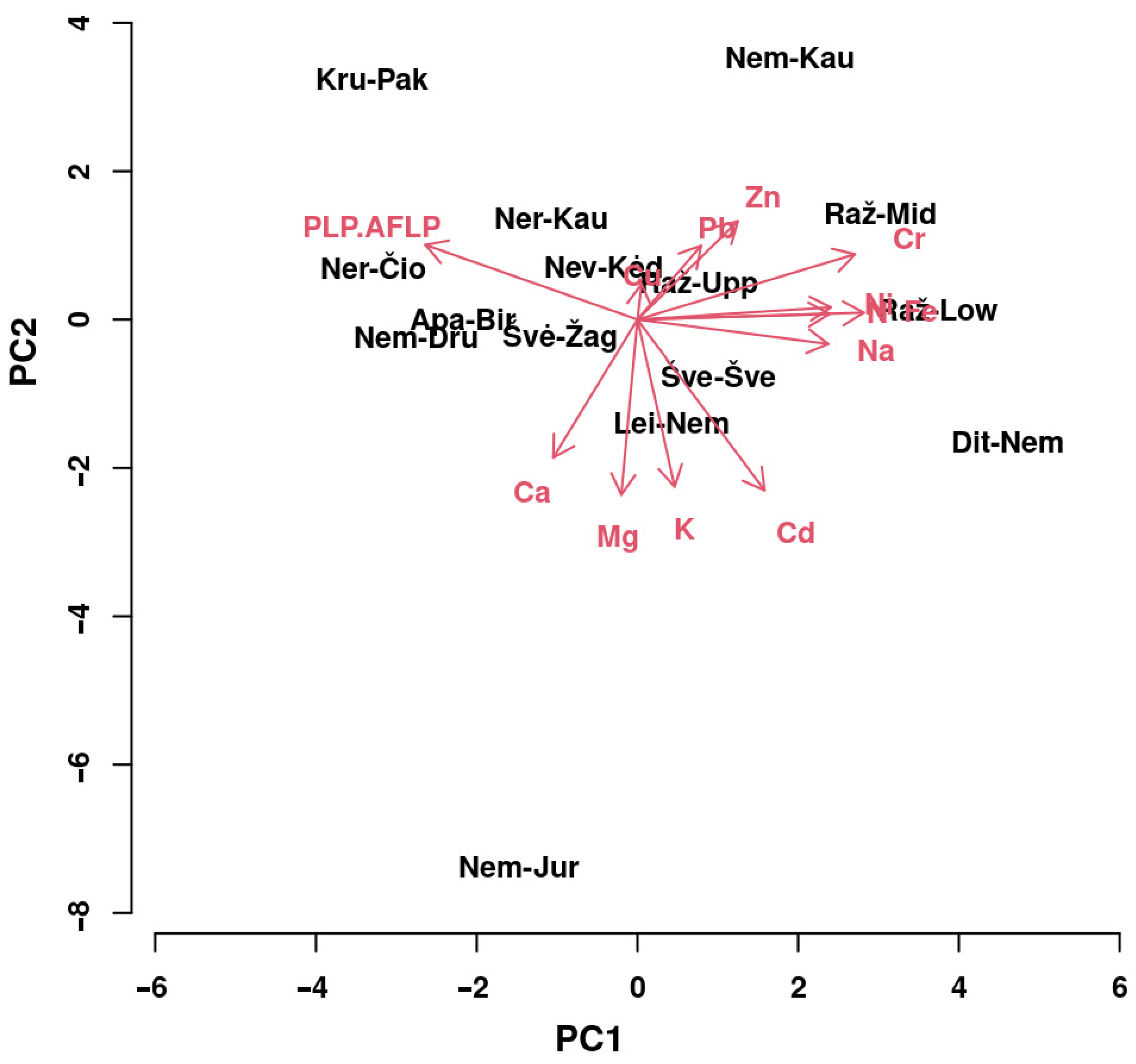
3.3. Relations between Element and Molecular Data
4. Discussion
4.1. Macroelements
4.2. Microelements
4.3. Heavy Metals
4.4. Ionome-Environment Relationships
4.5. Ionome–Genome Relationships
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Janse, J.H.; Van Dam, A.A.; Hes, E.M.; de Klein, J.J.; Finlayson, C.M.; Janssen, A.B.; van Wijk, D.; Mooij, M.W.; Verhoeven, J.T. Towards a global model for wetlands ecosystem services. Curr. Opin. Environ. Sustain. 2019, 36, 11–19. [Google Scholar] [CrossRef]
- van Kleunen, M.; Bossdorf, O.; Dawson, W. The ecology and evolution of alien plants. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 25–47. [Google Scholar] [CrossRef] [Green Version]
- Anderson, N.O. Throwing out the bathwater but keeping the baby: Lessons learned from purple loosestrife and reed canarygrass. HortTechnology 2019, 29, 539–548. [Google Scholar] [CrossRef]
- Thompson, D.Q.; Stuckey, R.L.; Thompson, E.B. Spread, impact, and control of purple loosestrife (Lythrum salicaria) in North American wetlands. U.S. Fish Wildl. Serv. 1987, 1–55. [Google Scholar]
- Lavoie, C. Should we care about purple loosestrife? The history of an invasive plant in North America. Biol. Invasions 2010, 12, 1967–1999. [Google Scholar] [CrossRef]
- Vet, R.; Artz, R.S.; Carou, S.; Shaw, M.; Ro, C.U.; Aas, W.; Baker, A.; Van Bowersox, C.; Dentener, F. A global assessment of precipitation chemistry and deposition of sulfur, nitrogen, sea salt, base cations, organic acids, acidity and pH, and phosphorus. Atmos. Environ. 2014, 93, 3–100. [Google Scholar] [CrossRef]
- O’Hare, M.T.; Baattrup-Pedersen, A.; Baumgarte, I.; Freeman, A.; Gunn, I.D.M.; Lázár, A.N.; Sinclair, R.; Wade, A.J.; Bowes, M.J. Responses of aquatic plants to eutrophication in rivers: A revised conceptual model. Front. Plant Sci. 2018, 9, 451. [Google Scholar] [CrossRef]
- Erős, T.; Kuehne, L.; Dolezsaib, A.; Sommerwerk, N.; Wolter, C. A systematic review of assessment and conservation management in large floodplain rivers–actions postponed. Ecol. Indic. 2019, 98, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Fuller, M.R.; Doyle, M.W.; Strayer, D.L. Causes and consequences of habitat fragmentation in river networks. Ann. N. Y. Acad. Sci. 2015, 1355, 31–51. [Google Scholar] [CrossRef]
- Zviedre, E.; Vītola, I.; Vizule-Kahovska, L.; Upena, I. Evaluation of Phytobenthos and Macrophytes of the Inland Surface Waters and Ecological Status Defined by Macrophyte Reference Index; Report, Part II, Rivers; Latvian Environment, Geology and Meteorology Centre: Ryga, Latvia, 2015. [Google Scholar]
- Hille, S.; Larsen, S.E.; Rubæk, G.H.; Kronvang, B.; Baattrup-Pedersen, A. Does regular harvesting increase plant diversity in buffer strips separating agricultural land and surface waters? Front. Environ. Sci. 2018, 6, 58. [Google Scholar] [CrossRef]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulißen, D. Indicator values of plants in Central Europe. Scr. Geobot. 1992, 18, 258. [Google Scholar]
- Han, W.; Fang, J.; Guo, D.; Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 2015, 168, 377–385. [Google Scholar] [CrossRef]
- Thompson, K.; Parkinson, J.A.; Band, S.R.; Spencer, R.E. A comparative study of leaf nutrient concentrations in a regional herbaceous flora. New Phytol. 1997, 136, 679–689. [Google Scholar] [CrossRef]
- Innes, J.L. Influence of air pollution on the foliar nutrition of conifers in Great Britain. Environ. Pollut. 1995, 88, 183–192. [Google Scholar] [CrossRef]
- Anderson, N.O.; Ascher, P.D. Style morph frequencies in Minnesota populations of Lythrum (Lythraceae) II. Tristylous L. salicaria L. Sex. Plant Reprod. 1995, 8, 105–112. [Google Scholar] [CrossRef]
- Caswell, W.D.; Davis, A.R. Pollen and ovule production, floral nectary structure, and nectar secretion dynamics in tristylous Lythrum salicaria L. Plant Syst. Evol. 2011, 294, 127–145. [Google Scholar] [CrossRef]
- Kubátová, B.; Trávníček, P.; Bastlová, D.; Čurn, V.; Jarolímová, V.; Suda, J. DNA ploidy-level variation in native and invasive populations of Lythrum salicaria at a large geographical scale. J. Biogeogr. 2008, 35, 167–176. [Google Scholar] [CrossRef]
- Chun, Y.J.; Nason, J.D.; Moloney, K.A. Comparison of quantitative and molecular genetic variation of native vs. invasive populations of purple loosestrife (Lythrum salicaria L., Lythraceae). Mol. Ecol. 2009, 18, 3020–3035. [Google Scholar] [CrossRef]
- Houghton-Thompson, J.; Prince, H.H.; Smith, J.J.; Hancock, J.F. Evidence of Hybridization between Lythrum salicaria (Purple Loosestrife) and L. alatum (Winged Loosestrife) in North America. Ann. Bot. 2005, 96, 877–885. [Google Scholar] [CrossRef]
- Middleton, B.A.; Travis, S.E.; Kubátová, B.; Johnson, D.; Edwards, K.R. Morphology and genetics of Lythrum salicaria from latitudinal gradients of the Northern Hemisphere grown in cold and hot common gardens. PLoS ONE 2019, 14, e0208300. [Google Scholar] [CrossRef] [Green Version]
- Jocienė, L.; Krokaitė, E.; Shakeneva, D.; Rekašius, T.; Stanys, V.; Šikšnianienė, J.B.; Žvingila, D.; Paulauskas, A.; Kupčinskienė, E. Relationship between genetic and environmental characteristics of Lithuanian populations of purple loosestrife (Lythrum salicaria). J. Environ. Eng. Landsc. Manag. 2022, 30, 81–93. [Google Scholar] [CrossRef]
- Al-Snafi, A.E. Chemical constituents and pharmacological effects of Lythrum salicaria—A review. IOSR J. Pharm. 2019, 9, 51–59. [Google Scholar]
- Brown, B.J.; Mitchell, R.J.; Graham, S.A. Competition for pollination between an invasive species (purple loosestrife) and a native congener. Ecology 2002, 83, 2328–2336. [Google Scholar] [CrossRef]
- Flanagan, R.J.; Mitchell, R.J.; Karron, J.D. Increased relative abundance of an invasive competitor for pollination, Lythrum salicaria, reduces seed number in Mimulus ringens. Oecologia 2010, 164, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Hovivk, S.M.; Bunker, D.E.; Peterson, C.J.; Carson, W.P. Purple loosestrife suppresses plant species colonization far more than broad-leaved cattail: Experimental evidence with plant community implications. J. Ecol. 2011, 99, 225–234. [Google Scholar] [CrossRef]
- Dávalos, A.; Blossey, B. Matrix habitat and plant damage influence colonization of purple loosestrife patches by specialist leaf-beetles. Environ. Entomol. 2011, 40, 1074–1080. [Google Scholar] [CrossRef] [PubMed]
- Boag, A.E.; Eckert, C.G. The effect of host abundance on the distribution and impact of biocontrol agents on purple loosestrife (Lythrum salicaria, Lythraceae). Écoscience 2013, 20, 90–99. [Google Scholar] [CrossRef]
- Rogers, J. Controlling purple loosestrife (Lythrum salicaria) along roadsides in St. Lawrence County: Monitoring and biological controls. Adirond. J. Environ. Stud. 2019, 23, 5. [Google Scholar]
- Adams, C.R.; Hovick, S.M.; Anderson, N.O.; Kettenring, K.M. We can better manage ecosystems by connecting solutions to constraints: Learning from wetland plant invasions. Front. Environ. Sci. 2021, 9, 715350. [Google Scholar] [CrossRef]
- Blossey, B.; Skinner, L.C.; Taylor, J. Impact and management of purple loosestrife (Lythrum salicaria) in North America. Biodivers. Conserv. 2001, 10, 1787–1807. [Google Scholar] [CrossRef]
- Russell-Mercierand, J.L.; Sargent, R.D. Indirect effects of herbivory on plant pollinator interactions in invasive Lythrum salicaria. Am. J. Bot. 2015, 102, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Macel, M.; Tielbörger, K.; Verhoeven, K.J. Effects of admixture in native and invasive populations of Lythrum salicaria. Biol. Invas. 2018, 20, 2381–2393. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, C. The Relative Roles of Herbivore- and Pollinator-Mediated Selection on the Evolution of Floral Display in the Invasive Plant, Lythrum salicaria. Ph.D. Thesis, Université d’Ottawa/University of Ottawa, Ottawa, ON, Canada, 2015. [Google Scholar]
- Olsson, K.; Gren, J.A. Latitudinal population differentiation in phenology, life history and flower morphology in the perennial herb Lythrum salicaria. J. Evol. Biol. 2002, 15, 983–996. [Google Scholar] [CrossRef]
- Campos, A.C.A.L.; Van Dijk, W.F.A.; Ramakrishna, P.; Giles, T.; Korte, P.; Douglas, A.; Smith, P.; Salt, D.E. 1135 ionomes reveal the global pattern of leaf and seed mineral nutrient and trace element diversity in Arabidopsis thaliana. Plant J. 2021, 106, 536–554. [Google Scholar] [CrossRef]
- Jarčuška, B.; Slezák, M.; Hrivnák, R.; Senko, D. Invasibility of alien Impatiens parviflora in temperate forest understories. Flora 2016, 224, 14–23. [Google Scholar] [CrossRef]
- Schmitz, D.; Girardi, J.; Jamin, J.; Bundschuh, M.; Geng, B.; Feldmann, R.; Rösch, V.; Riess, K.; Schirmel, J. Copper Uptake and Its Effects on Two Riparian Plant Species, the Native Urtica dioica, and the Invasive Fallopia japonica. Plants 2023, 12, 481. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.; Acreman, M.C. Wetland nutrient removal: A review of the evidence. Hydrol. Earth Syst. Sci. 2004, 8, 673–685. [Google Scholar] [CrossRef] [Green Version]
- Uveges, J.L.; Corbett, A.L.; Mal, T.K. Effects of lead contamination on the growth of Lythrum salicaria (purple loosestrife). Environ. Pollut. 2002, 120, 319–323. [Google Scholar] [CrossRef]
- Nicholls, A.M.; Mal, T.K. Effects of lead and copper exposure on growth of an invasive weed, Lythrum salicaria L. (purple loosestrife). Ohio J. Sci. 2003, 103, 129–133. [Google Scholar]
- Zhou, Q.; Zhu, H.; Bañuelos, G.; Yan, B.; Liang, Y.; Yu, J.; Li, H. Impacts of vegetation and temperature on the treatment of domestic sewage in constructed wetlands incorporated with Ferric-Carbon micro-electrolysis material. Int. J. Phytoremediation 2017, 19, 915–924. [Google Scholar] [CrossRef]
- Ma, W.; Zhao, B.; Ma, J. Comparison of heavy metal accumulation ability in rainwater by 10 sponge city plant species. Environ. Sci. Pollut. Res. 2019, 26, 26733–26747. [Google Scholar] [CrossRef]
- Zuo, X.; Zhang, H.; Yu, J. Microbial diversity for the improvement of nitrogen removal in stormwater bioretention cells with three aquatic plants. Chemosphere 2020, 244, 125626. [Google Scholar] [CrossRef] [PubMed]
- Dudek, T.; Wolański, P.; Rogut, K. The content of macro-and micro minerals in the sward of different types of semi-natural meadows of temperate climate in SE Poland. Agronomy 2020, 10, 273. [Google Scholar] [CrossRef] [Green Version]
- Muller, L.A.H.; Lambaerts, M.; Vangronsveld, J.; Colpaert, J.V. AFLP-based assessment of the effects of environmental heavy metal pollution on the genetic structure of pioneer populations of Suillus luteus. New Phytol. 2004, 164, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Muller, L.A.; Vangronsveld, J.; Colpaert, J.V. Genetic structure of Suillus luteus populations in heavy metal polluted and nonpolluted habitats. Mol. Ecol. 2007, 16, 4728–4737. [Google Scholar] [CrossRef]
- Houdegbe, A.C.; Achigan-Dako, E.G.; Sogbohossou, E.O.; Schranz, M.E.; Odindo, A.O.; Sibiya, J. Leaf elemental composition analysis in spider plant [Gynandropsis gynandra L. (Briq.)] differentiates three nutritional groups. Front. Plant Sci. 2022, 3091. [Google Scholar] [CrossRef]
- Cao, F.; Chen, F.; Sun, H.; Zhang, G.; Chen, Z.-H.; Wu, F. Genome-wide transcriptome and functional analysis of two contrasting genotypes reveals key genes for cadmium tolerance in barley. BMC Genom. 2014, 15, 611. [Google Scholar] [CrossRef] [Green Version]
- Ansari, M.K.A.; Ahmad, A.; Umar, S.; Zia, M.H.; Iqbal, M.; Owens, G. Genotypic variation in phytoremediation potential of Indian mustard exposed to nickel stress: A hydroponic study. Int. J. Phytoremediation 2015, 17, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy metal tolerance in plants: Role of transcriptomics, proteomics, metabolomics, and ionomics. Front. Plant Sci. 2016, 6, 1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, W.; Liu, C.; Cao, B.; Qin, M.; Long, D.; Xiang, Z.; Zhao, A. Genome-Wide Identification and characterization of four gene families putatively involved in cadmium uptake, translocation and sequestration in mulberry. Front. Plant Sci. 2018, 9, 879. [Google Scholar] [CrossRef]
- Angulo-Bejarano, P.I.; Puente-Rivera, J.; Cruz-Ortega, R. Metal and metalloid toxicity in plants: An overview on molecular aspects. Plants 2021, 10, 635. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.E. Analysis of vegetation and other organic materials. In Chemical Analysis of Ecological Materials, 2nd ed.; Allen, S.E., Ed.; Blackwell Scientific Publications: Oxford/London, UK, 1989; pp. 46–61. [Google Scholar]
- Kjeldahl, C. A new method for the determination of nitrogen in organic matter. J. Anal. Chem. 1883, 22, 366–372. [Google Scholar]
- Lujanienė, G.; Valiulis, D.; Byčenkienė, S.; Šakalys, J.; Povinec, P.P. Plutonium isotopes and 241Am in the atmosphere of Lithuania: A comparison of different source terms. Atmos. Environ. 2012, 61, 419–427. [Google Scholar] [CrossRef]
- Lee, S.H.; Oh, J.S.; Lee, J.M.; Lee, K.B.; Park, T.S.; Lujaniene, G.; Valiulis, D.; Šakalys, J. Distribution characteristics of 137Cs, Pu isotopes and 241Am in soil in Korea. Appl. Radiat. Isot. 2013, 81, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Lujanienė, G.; Šemčuk, S.; Lečinskytė, A.; Kulakauskaitė, I.; Mažeika, K.S.; Valiulis, D.; Pakštas, V.; Skapas, M.; Tumėnas, S. Magnetic graphene oxide based nano-composites for removal of radionuclides and metals from contaminated solutions. J. Environ. Radioact. 2017, 166, 166–174. [Google Scholar] [CrossRef] [PubMed]
- CLC. CORINE Land Cover Nomenclature Conversion to Land Cover Classification System. Available online: https://land.copernicus.eu/eagle/files/eagle-related-projects/pt_clcconversion-to-fao-lccs3_dec2010 (accessed on 30 December 2022).
- Gailiušis, B.; Jablonskis, J.; Kovalenkovienė, M. Lithuanian Rivers: Hydrography and Runoff; Lithuanian Energy Institute: Kaunas, Lithuania, 2001. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Signorell, A.; Aho, K.; Alfons, A.; Anderegg, N.; Aragon, T.; Arachchige, C.; Bolker, B. DescTools: Tools for Descriptive Statistics; R Package Version 0.99.42; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Subirana, I.; Sanz, H.; Vila, J. Building Bivariate Tables: The compareGroups Package for R. J. Stat. Soft. 2014, 57, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.; Simko, V. R Package ‘Corrplot’: Visualization of a Correlation Matrix (Version 0.90). Available online: https://github.com/taiyun/corrplot (accessed on 30 December 2022).
- Chapin, F.S. The mineral nutrition of wild plants. Ann. Rev. Ecol. Syst. 1980, 11, 233–260. [Google Scholar] [CrossRef]
- Watanabe, T.; Broadley, M.R.; Jansen, S.; White, P.J.; Takada, J.; Satake, K.; Takamatsu, T.; Tuah, S.J.; Osaki, M. Evolutionary control of leaf element composition in plants. New Phytol. 2007, 174, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Vergutz, L.; Manzoni, S.; Porporato, A.; Novais, R.F.; Jackson, R.B. Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecol. Monogr. 2012, 82, 205–220. [Google Scholar] [CrossRef] [Green Version]
- Kozma, A.; Tölgyesi, G. Plant associations of flood plains along the Middle Tisza and their agricultural utilization. Tiscia 1979, 14, 105–122. [Google Scholar]
- Tölgyesi, G.; Kozma, A. Taxonomic ans ecological comments relating macro- and micro-element concentrations in plant species of inundation area. Tiscia 1983, 18, 71–74. [Google Scholar]
- Emery, S.L.; Perry, J.A. Aboveground biomass and phosphorus concentrations of Lythrum salicaria (Purple loosestrife) and Typha spp. (Cattail) in 12 Minnesota wetlands. Am. Midl. Nat. 1995, 394–399. [Google Scholar] [CrossRef]
- Vardanyan, L.; Schmieder, K.; Sayadyan, H.; Heege, T.; Heblinski, J.; Agyemang, T.; De, J.; Breuer, J. Heavy Metal Accumulation by Certain Aquatic Macrophytes from Lake Sevan (Armenia). In Proceedings of the Taal: The 12th World Lake Conference, Jaipur, India, 28 October–2 November 2008; pp. 1028–1038. [Google Scholar]
- Hubbard, A.R. Assessment of Trace Metal Concentrations in Lythrum salicaria at Three Rhode Island Sites Using ICP-MS Analysis. Master’s Thesis, Bryant University, Smithfield, RI, USA, 2015. [Google Scholar]
- Yue, K.; Peng, Y.; Fornara, D.A.; Van Meerbeek, K.; Vesterdal, L.; Yang, W.; Peng, C.; Tan, B.; Zhou, W.; Xu, Z.; et al. Responses of nitrogen concentrations and pools to multiple environmental change drivers: A meta-analysis across terrestrial ecosystems. Glob. Ecol. Biogeogr. 2019, 28, 690–724. [Google Scholar] [CrossRef]
- Wei, C.; Tang, S.; Pan, Y.; Li, X. Plastic responses of invasive Bidens frondosa to water and nitrogen addition. Nord J. Bot. 2017, 35, 232–239. [Google Scholar] [CrossRef]
- Beumer, V.; Ohm, J.N.; van Wirdum, G.; Beltman, B.; Griffioen, J.; Verhoeven, J.T.A. Biogeochemical plant site conditions in stream valleys after winter flooding: A phytometer approach. Biogeosci. Discuss. 2008, 5, 5203–5232. [Google Scholar] [CrossRef] [Green Version]
- Shamsi, S.R.A.; Whitehead, F.H. Comparative eco-physiology of Epilobium hirsutum L. and Lythrum salicaria L. III. Mineral nutrition. J. Ecol. 1977, 65, 55–70. [Google Scholar] [CrossRef]
- Krokaitė, E.; Shakeneva, D.; Juškaitytė, E.; Rekašius, T.; Nemaniūtė-Gužienė, J.; Butkuvienė, J.; Patamsytė, J.; Rančelienė, V.; Vyšniauskienė, R.; Duchovskienė, L.; et al. Nitrogen concentration of the aquatic plant species in relation to land cover type and other variables of the environment. Zemdirbyste 2019, 106, 203–212. [Google Scholar] [CrossRef]
- Yu, H.; Wang, L.; Liu, C.; Fan, S. Coverage of native plants is key factor influencing the invasibility of freshwater ecosystems by exotic plants in China. Front. Plant Sci. 2018, 9, 250. [Google Scholar] [CrossRef] [Green Version]
- Dostal, P.; Dawson, W.; van Kleunen, M.; Keser, L.H.; Fischer, M. Central European plant species from more productive habitats are more invasive at a global scale: Habitat productivity at origin-plant invasiveness elsewhere. Global Ecol. Biogeogr. 2013, 22, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Legault, R.; Zogg, G.P.; Travis, S.E. Competitive interactions between native Spartina alterniflora and non-native Phragmites australis depend on nutrient loading and temperature. PLoS ONE 2018, 13, e0192234. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Cui, M.; Du, Y.; Ren, G.; Li, J.; Wang, C.; Li, G.; Dai, Z.; Rutherford, S.; Wan, J.S.H.; et al. Influence of multiple global change drivers on plant invasion: Additive effects are uncommon. Front. Plant Sci. 2022, 13, 1020621. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Liu, Y.; Zhao, S.; Dai, L.; Zhang, Z.; Zhang, M.; Zhang, Z. How Waterlogged Conditions Influence the Nitrogen Dynamics in a Soil–Water–Plant System: Implications for Wetland Restoration. Water 2021, 13, 2957. [Google Scholar] [CrossRef]
- Tong, X.; Wang, X.; He, X.; Sui, Y.; Shen, J.; Feng, J. Effects of antibiotics on nitrogen uptake of four wetland plant species grown under hydroponic culture. Environ. Sci. Pollut. Res. 2019, 26, 10621–10630. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Niu, S.; Yu, G. Aggravated phosphorus limitation on biomass production under increasing nitrogen loading: A meta-analysis. Global Change Biol. 2016, 22, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Sardans, J.; Peñuelas, J. Potassium control of plant functions: Ecological and agricultural implications. Plants 2021, 10, 419. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: San Diego, CA, USA, 2011. [Google Scholar]
- Istvan, P.; Benton, J. Trace Elements; Lucie Press: Boca Raton, FL, USA, 1997. [Google Scholar]
- Teuchies, J.; Jacobs, A.; Oosterlee, L.; Bervoets, L.; Meire, P. Role of plants in metal cycling in a tidal wetlands: Implications for phytoremediation. Sci. Total Environ. 2013, 445–446, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Kronzucker, H.J.; Coskun, D.; Schulze, L.M.; Wong, J.R.; Britto, D.T. Sodium as nutrient and toxicant. Plant Soil 2013, 369, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Tsonev, T.; Lidon, F.J.C. Zinc in plants—An overview. Emir. J. Food Agric. 2012, 24, 322–333. [Google Scholar]
- Aravind, P.; Prasad, M.N.V. Zinc alleviates cadmium-induced oxidative stress in Ceratophyllum demersum L.: A free floating freshwater macrophyte. Plant Physiol. Biochem. 2003, 41, 391–397. [Google Scholar] [CrossRef]
- Betül, A.K.I.N.; Bingöl, N.A. Heavy metal accumulation in wetland plants and water-sediment relationship in Köprüören-Kütahya. J. Limnol. Freshw. Fish. Res. 2019, 5, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Bingöl, N.A.; Akin, B.; Kocaçalişkan, İ.L.; Nalbantoğlu, B.; Meşeli, O. Effect of zinc on phytoremediation potential and carbonic anhydrase and polyphenol oxidase activities of Lythrum salicaria L. Turk. J. Bot. 2021, 45, 553–562. [Google Scholar] [CrossRef]
- Andresen, E.; Peiter, E.; Küpper, H. Trace metal metabolism in plants. J. Exp. Bot. 2018, 69, 909–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riyazuddin, R.; Nisha, N.; Ejaz, B.; Khan, M.I.R.; Kumar, M.; Ramteke, P.W.; Gupta, R. A comprehensive review on the heavy metal toxicity and sequestration in plants. Biomolecules 2021, 12, 43. [Google Scholar] [CrossRef]
- Sperdouli, I. Heavy metal toxicity effects on plants. Toxics 2022, 10, 715. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, D.; Krivokapic, S. The effect of Cu, Zn, Cd, and Pb accumulation on biochemical parameters (proline, chlorophyll) in the water caltrop (Trapa natans L.), Lake Skadar, Montenegro. Plants 2020, 9, 1287. [Google Scholar] [CrossRef] [PubMed]
- Čeburnis, D.; Steinnes, E. Conifer needles as biomonitors of atmospheric heavy metal deposition: Comparison with mosses and precipitation, role of the canopy. Atmos. Environ. 2000, 34, 4265–4271. [Google Scholar] [CrossRef]
- Kupcinskiene, E. Foliar elements of Scots pine across the transect from the ammonia emission source (1995–1998). Ekológia 2003, 22, 171–175. [Google Scholar]
- Kuznetsova, T.; Lukjanova, A.; Mandre, M.; Lõhmus, K. Aboveground biomass and nutrient accumulation dynamics in young black alder, silver birch and Scots pine plantations on reclaimed oil shale mining areas in Estonia. For. Ecol. Manag. 2011, 262, 56–64. [Google Scholar] [CrossRef]
- Jocienė, L.; Krokaitė, E.; Rekašius, T.; Vilčinskas, R.; Judžentienė, A.; Marozas, V.; Kupčinskienė, E. Ionomic parameters of populations of common juniper (Juniperus communis L.) depending on the habitat type. Plants 2023, 12, 961. [Google Scholar] [CrossRef] [PubMed]
- Abolina, L.; Osvalde, A.; Karlsons, A. Habitat characteristics and mineral nutrition status of Rubus chamaemorus L. in Latvia. Plants 2023, 12, 528. [Google Scholar] [CrossRef]
- Fastovetska, K.; Šlepetiene, A.; Vigricas, E.; Urbaitis, G.; Belova, O. Lead content in plant materials in the buffer zones of surface water bodies of Northwestern and Central regions of Lithuania. Zemdirbyste 2022, 109, 335–340. [Google Scholar] [CrossRef]
- Bingöl, N.A.; Akin, B.; Ozmal, V. Phytoremediation and Biosorption Potential of Lythrum salicaria L. for Nickel Removal from Aqueous Solutions. Pol. J. Environ. Stud. Vol. 2017, 26, 2479–2485. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kapoor, D.; Wang, J.; Shahzad, B.; Kumar, V.; Bali, A.S.; Jasrotia, S.; Zheng, B.; Yuan, H.; Yan, D. Chromium bioaccumulation and its impacts on plants: An overview. Plants 2020, 9, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Serrano, M.; Romero-Puertas, M.C.; Pazmino, D.M.; Testillano, P.S.; Risueño, M.C.; Del Río, L.A.; Sandalio, L.M. Cellular response of pea plants to cadmium toxicity: Cross talk between reactive oxygen species, nitric oxide, and calcium. Plant Physiol. 2009, 150, 229–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.-Y.; Salt, D.E. Plant ionomics: From elemental profiling to environmental adaptation. Mol. Plant. 2016, 9, 787–797. [Google Scholar] [CrossRef] [Green Version]
- Stein, R.J.; Höreth, S.; De Melo, J.R.F.; Syllwasschy, L.; Lee, G.; Garbin, M.L.; Clemens, S.; Krämer, U. Relationships between soil and leaf mineral composition are element-specific, environment-dependent and geographically structured in the emerging model Arabidopsis halleri. New Phytol. 2017, 213, 1274–1286. [Google Scholar] [CrossRef] [Green Version]
- Brezinová, T.; Vymazal, J. Nitrogen standing stock in Phragmites australis growing in constructed wetlands—Do we evaluate it correctly? Ecol. Eng. 2015, 74, 286–289. [Google Scholar] [CrossRef]
- Anderson, N.O.; Jocienė, L.; Krokaitė, E.; Rekašius, T.; Paulauskas, A.; Kupčinskienė, E. Genetic diversity of Phalaris arundinacea populations in relation to river regulation in the Merkys basin, Lithuania. River Res. Appl. 2018, 34, 1–10. [Google Scholar] [CrossRef]
- Butkuvienė, J.; Sinkevičienė, Z.; Naugžemys, D.; Žvingila, D. Floristic structure of two Batrachium rich plant communities in relation to habitat conditions in rivers of Lithuania. Pol. J. Ecol. 2018, 66, 1–13. [Google Scholar] [CrossRef]
- Vyšniauskienė, R.; Rančelienė, V.; Naugžemys, D.; Patamsytė, J.; Sinkevičienė, Z.; Butkuvienė, J.; Žvingila, D. Genetic diversity of populations of Bidens genera invasive and native species in Lithuania. Zemdirbyste 2018, 105, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Vyšniauskienė, R.; Rančelienė, V.; Naugžemys, D.; Rudaitytė-Lukošienė, E.; Patamsytė, J.; Butkauskas, D.; Kupčinskienė, E.; Žvingila, D. Genetic diversity of Nuphar lutea in Lithuanian river populations. Aquat. Bot. 2020, 161, 103173. [Google Scholar] [CrossRef]
- Basic, N.; Besnard, G. Gene polymorphisms for elucidating the genetic structure of the heavy-metal hyperaccumulating trait in Thlaspi caerulescens and their cross-genera amplification in Brassicaceae. J. Plant Res. 2006, 119, 479–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prus-Glowacki, W.; Chudzinska, E.; Wojnicka-Poltorak, A.; Fagiewicz, K. Effects of heavy metal pollution on genetic variation and cytological disturbances in the Pinus sylvestris L. population. J. Appl. Genet. 2006, 47, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Yan, Y.; Hu, W.; Liu, G.; Zeng, H.; Wei, Y.; Shi, H. Genome-wide association studies reveal genetic basis of ionomic variation in cassava. Plant J. 2022, 112, 1212–1223. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Population | River | River Basin | Location | Latitude (°N) | Longitude (°E) |
|---|---|---|---|---|---|
| Nem-Dru | Nemunas | Nemunas | Druskininkai | 54°01′12.9″ | 23°58′53.9″ |
| Ner-Čio | Neris | Nemunas | Čiobiškis | 54°56′55.9″ | 24°40′28.0″ |
| Ner-Kau | Neris | Nemunas | Kaunas | 54°58′46.2″ | 24°01′37.7″ |
| Nem-Kau | Nemunas | Nemunas | Kaunas | 54°53′35.9″ | 23°53′21.0″ |
| Nev-Kėd | Nevėžis | Nemunas | Kėdainiai | 55°17′58.3″ | 23°59′45.5″ |
| Nem-Jur | Nemunas | Nemunas | Jurbarkas | 55°05′35.1″ | 22°43′48.9″ |
| Lei-Nem | Leitė | Nemunas | Sausgalviai | 55°15′57.6″ | 21°27′18.4″ |
| Rąž-Upp | Rąžė | Seaside Rivers | Palanga | 55°54′39.8″ | 21°04′28.9″ |
| Raž-Mid | Rąžė | Seaside Rivers | Palanga | 55°54′59.3″ | 21°03′56.4″ |
| Rąž-Low | Rąžė | Seaside Rivers | Palanga | 55°55′14.1″ | 21°03′21.8″ |
| Dit-Nmr | Ditch | Seaside Rivers | Nemirsėta | 55°52′50.1″ | 21°03′50.2″ |
| Šve-Šve | Šventoji | Seaside Rivers | Šventoji | 56°02′02.1″ | 21°05′12.3″ |
| Švė-Žag | Švėtė | Lielupė | Žagarė | 56°21′36.6″ | 23°15′07.5″ |
| Apa-Bir | Apaščia | Lielupė | Biržai | 56°11′16.7″ | 24°45′57.8″ |
| Kru-Pak | Kruoja | Lielupė | Pakruojis | 55°58′52.2″ | 23°51′15.8″ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krokaitė, E.; Jocienė, L.; Shakeneva, D.; Rekašius, T.; Valiulis, D.; Kupčinskienė, E. Ionome of Lithuanian Populations of Purple Loosestrife (Lythrum salicaria) and Its Relation to Genetic Diversity and Environmental Variables. Diversity 2023, 15, 418. https://doi.org/10.3390/d15030418
Krokaitė E, Jocienė L, Shakeneva D, Rekašius T, Valiulis D, Kupčinskienė E. Ionome of Lithuanian Populations of Purple Loosestrife (Lythrum salicaria) and Its Relation to Genetic Diversity and Environmental Variables. Diversity. 2023; 15(3):418. https://doi.org/10.3390/d15030418
Chicago/Turabian StyleKrokaitė, Edvina, Lina Jocienė, Dinara Shakeneva, Tomas Rekašius, Darius Valiulis, and Eugenija Kupčinskienė. 2023. "Ionome of Lithuanian Populations of Purple Loosestrife (Lythrum salicaria) and Its Relation to Genetic Diversity and Environmental Variables" Diversity 15, no. 3: 418. https://doi.org/10.3390/d15030418