
Trophic Ecology during the Ontogenetic Development of the Pelagic Thresher Shark Alopias pelagicus in Baja California Sur, Mexico

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
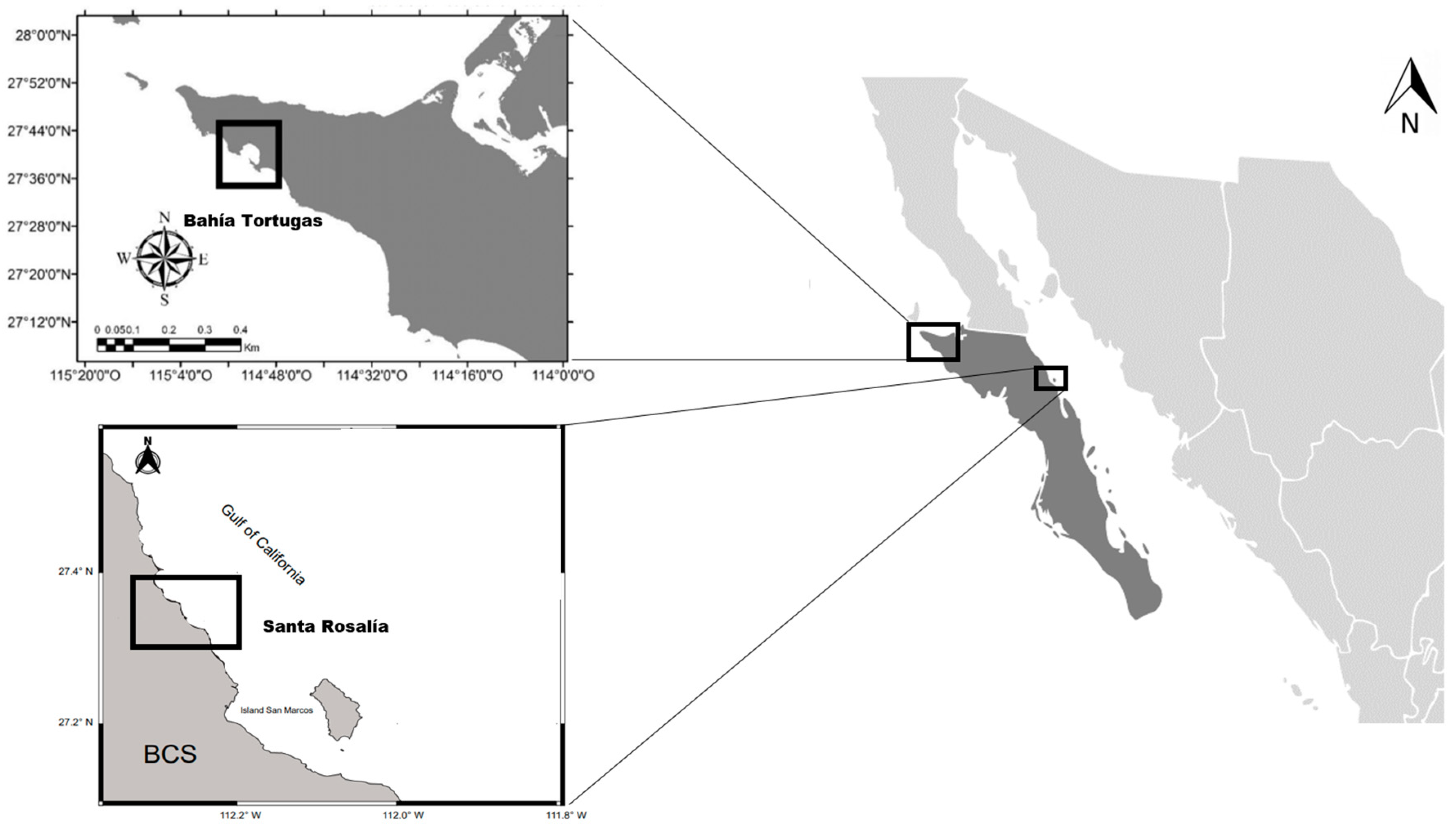
2.1. Study Area and Sample Collection
2.2. Laboratory Analysis
2.3. Data Analysis
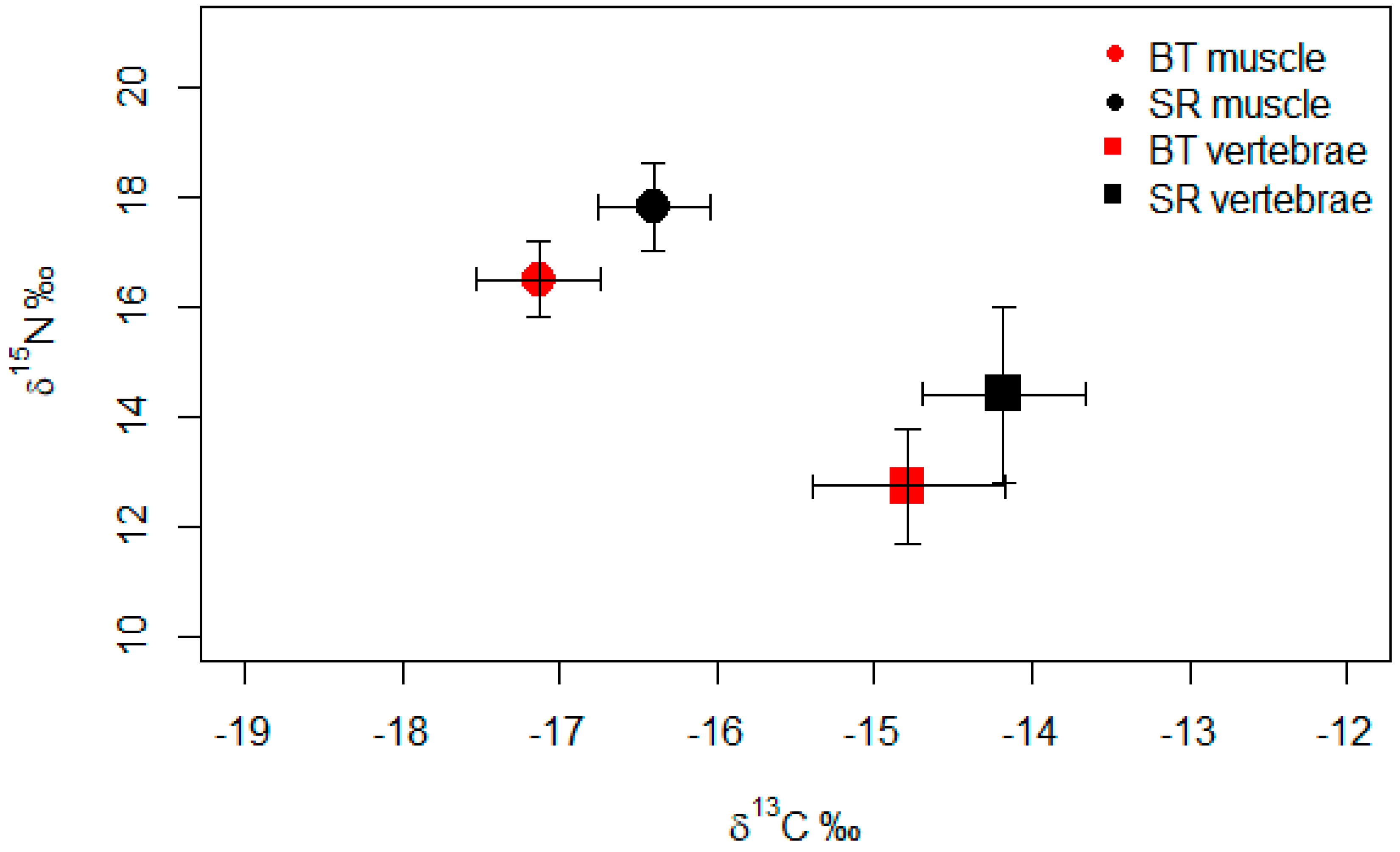
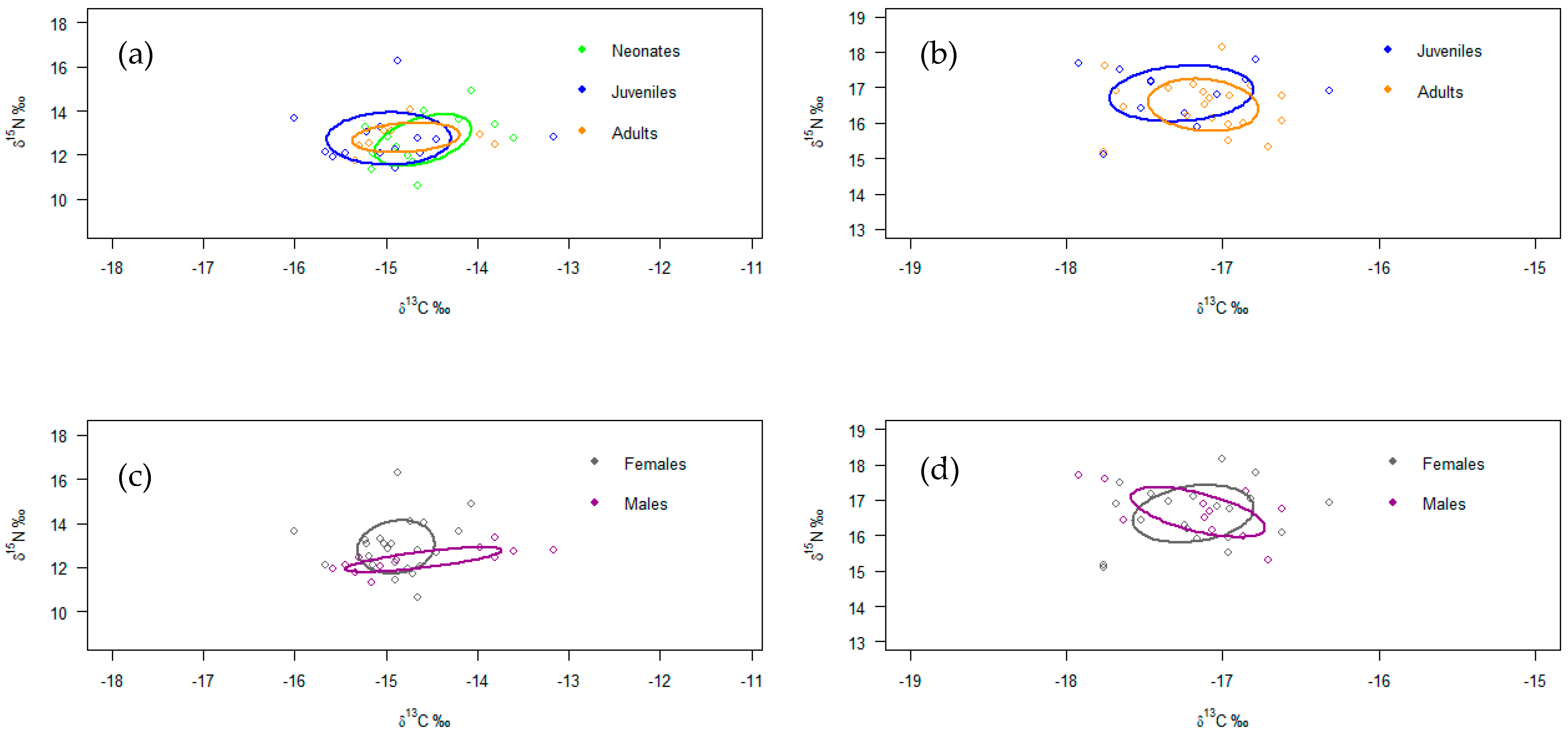
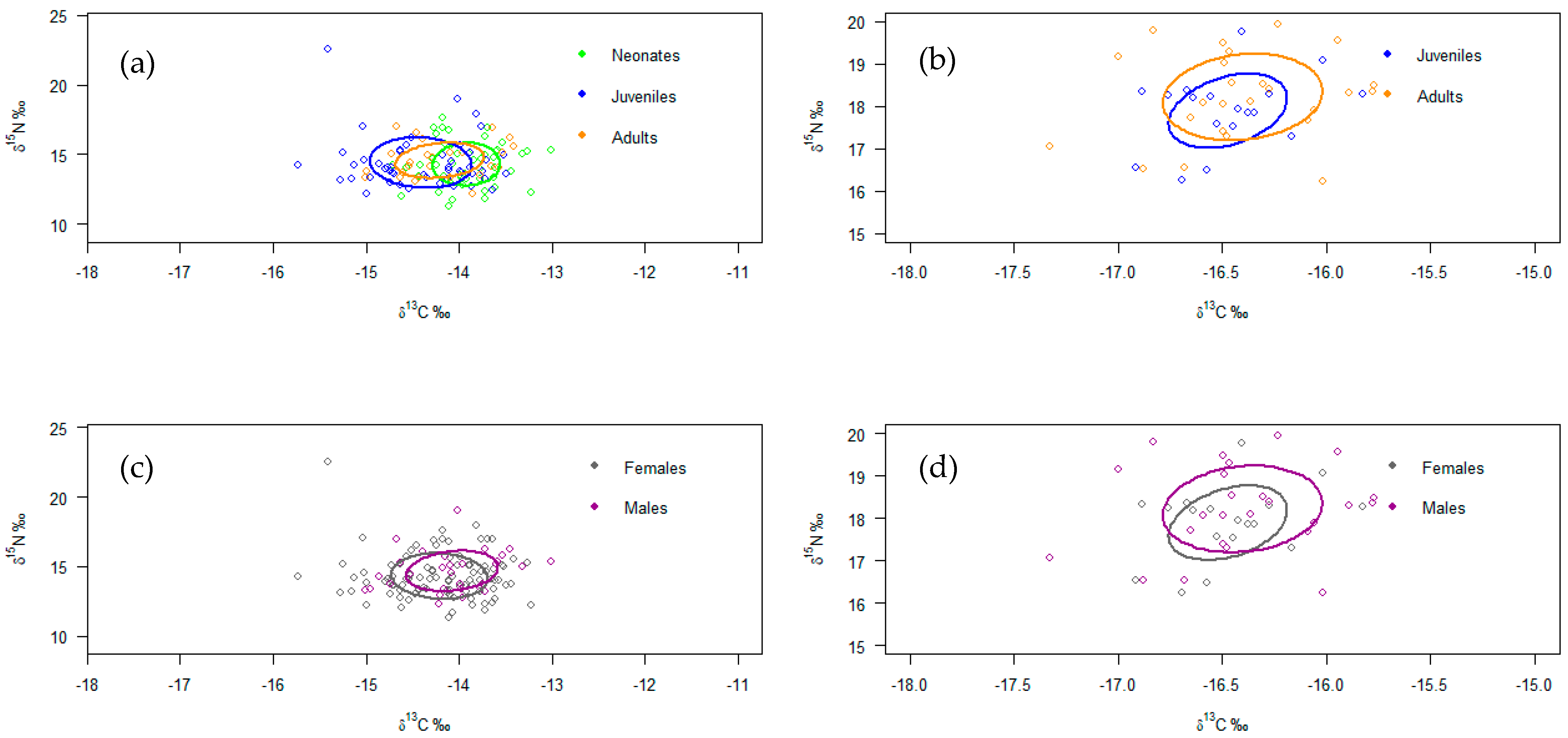
3. Results
4. Discussion
4.1. Comparison between Regions
4.2. Comparison between Tissues
4.3. Isotopic Analysis by Maturity Stages
4.4. Isotopic Analysis by Sex
4.5. Trophic Position
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weigmann, S. Annotated checklist of the living sharks, batoids and chimaeras (Chondrichthyes) of the world, with a focus on biogeographical diversity. J. Fish Biol. 2016, 88, 837–1037. [Google Scholar] [CrossRef] [PubMed]
- Romero-Caicedo, A.F.; Galván-Magaña, F.; Martinez-Ortiz, J. Reproduction of the pelagic thresher shark Alopias pelagicus in the equatorial Pacific. J. Mar. Biol. Assoc. 2014, 94, 1501–1507. [Google Scholar] [CrossRef]
- Liu, K.-M.; Chen, C.-T.; Liao, T.-H.; Joung, S.-J. Age, Growth, and Reproduction of the Pelagic Thresher Shark, Alopias pelagicus in the Northwestern Pacific. Copeia 1999, 1999, 68–74. [Google Scholar] [CrossRef]
- Smith, W.D.; Bizzarro, J.J.; Cailliet, G.M. La pesca artesanal de elasmobranquios en la costa oriental de Baja California, México: Características y consideraciones de manejo. Cienc. Mar. 2009, 35, 209–236. [Google Scholar] [CrossRef]
- Drew, M.; White, W.T.; Dharmadi; Harry, A.V.; Huveneers, C. Age, growth and maturity of the pelagic thresher Alopias pelagicus and the scalloped hammerhead Sphyrna lewini. J. Fish Biol. 2015, 86, 333–354. [Google Scholar] [CrossRef] [PubMed]
- Polo-Silva, C.; Rendón, L.; Galván-Magaña, F. Descripción de la dieta de los tiburones zorro (Alopias pelagicus) y (Alopias superciliosus) durante la época lluviosa en aguas ecuatorianas. PANAMJAS 2009, 4, 556–571. [Google Scholar]
- Calle-Morán, M.D. Ecología Trófica del Tiburón Zorro Pelágico Alopias pelagicus en Santa Rosa de Salinas, Pacífico Ecuatoriano. Master’s Thesis, Universidad Nacional Autónoma de México, Mexico City, Mexico, 2010. [Google Scholar]
- Estupiñán-Montaño, C. Ontogenia Alimentaria de Tres Especies de Tiburones Pelágicos: Alopias pelagicus, Carcharhinus falciformis y Prionace glauca en la Reserva Marina de Galápagos, Ecuador. Master’s Thesis, Instituto Politécnico Nacional, Centro Interdisciplinario de Ciencias Marinas, La Paz, Mexico, 2016. [Google Scholar]
- Lara, A.; Galván-Magaña, F.; Elorriaga-Verplancken, F.; Marmolejo-Rodríguez, A.J.; González-Armas, R.; Arreola-Mendoza, L.; Sujitha, S.B.; Jonathan, M.P. Bioaccumulation and trophic transfer of potentially toxic elements in the pelagic thresher shark Alopias pelagicus in Baja California Sur, Mexico. Mar. Pollut. Bull. 2020, 156, 111192. [Google Scholar] [CrossRef]
- García-Olvera, M.E. Ecología Trófica de los Tiburones Zorro (Alopias pelagicus) y Piloto (Carcharhinus falciformis) en los Puertos de Santa Rosalía y Punta Lobos, Baja California Sur. Bachelor’s Thesis, Universidad Autónoma de Ciudad Juarez, Ciudad Juarez, Mexico, 2023; 41p. [Google Scholar]
- Alghozali, F.A.; Salsabila, R.; Gustianto, M.W.D.; Putri, H.M.I.H.; Himawan, M.R.; Yuneni, R.R.; Hatmoro, C.K.; Rezkiani, M. Diet analyses of the pelagic thresher shark, Alopias pelagicus (Lamniformes: Alopiidae), from the Lombok Strait waters, Indonesia. Biodiversitas J. Biol. Divers. 2023, 24, 3708–3714. [Google Scholar] [CrossRef]
- Barnett, A.; Redd, K.S.; Frusher, S.D.; Stevens, J.D.; Semmens, J.M. Non-lethal method to obtain stomach samples from a large marine predator and the use of DNA analysis to improve dietary information. J. Exp. Mar. Biol. Ecol. 2010, 393, 188–192. [Google Scholar] [CrossRef]
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Hussey, N.E.; Brush, J.; McCarthy, I.D.; Fisk, A.T. Δ15N and δ13C diet–tissue discrimination factors for large sharks under semi-controlled conditions. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 155, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Peterson, B.J.; Fry, B. Stable Isotopes in Ecosystem Studies. Annu. Rev. Ecol. Syst. 1987, 18, 293–320. [Google Scholar] [CrossRef]
- Polo-Silva, C.; Newsome, S.D.; Galván-Magaña, F.; Grijalba-Bendeck, M.; Sanjuan-Muñoz, A. Trophic shift in the diet of the pelagic thresher shark based on stomach contents and stable isotope analyses. Mar. Biol. Res. 2013, 9, 958–971. [Google Scholar] [CrossRef]
- Estrada, J.A.; Rice, A.N.; Natanson, L.J.; Skomal, G.B. Use of Isotopic Analysis of Vertebrae in Reconstructing Ontogenetic Feeding Ecology in White Sharks. Ecology 2006, 87, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Grubbs, R.D. Ontogenetic shifts in movements and habitat use. In Sharks and Their Relatives II; CRC Press: Boca Raton, FL, USA, 2010; pp. 335–366. [Google Scholar]
- Lowe, C.G.; Wetherbee, B.M.; Crow, G.L.; Tester, A.L. Ontogenetic dietary shifts and feeding behavior of the tiger shark, Galeocerdo cuvier, in Hawaiian waters. Environ. Biol. Fishes 1996, 47, 203–211. [Google Scholar] [CrossRef]
- Martínez-Ayala, J.C. Ecología Trófica del Cazón Mamón Mustelus lunulatus en la Costa Occidental de Baja California Sur, México. Master’s Thesis, Instituto Politécnico Nacional, Centro Interdisciplinario de Ciencias Marinas, La Paz Mexico, 2018. [Google Scholar]
- Bizzarro, J.J.; Smith, W.D.; Hueter, R.E.; Tyminski, J.; Márquez, J.F.; Castillo, J.L.; Cailliet, G.M. El Estado Actual de los Tiburones y Rayas Sujetos a Explotación Comercial en el Golfo de California: Una Investigación Aplicada al Mejoramiento de su Manejo Pesquero y Conservación; Moss Landing Marine Laboratories Technical Publication: Moss Landing, CA, USA, 2007. [Google Scholar]
- Kim, S.L.; Koch, P.L. Methods to collect, preserve, and prepare elasmobranch tissues for stable isotope analysis. Environ. Biol. Fishes 2012, 95, 53–63. [Google Scholar] [CrossRef]
- Post, D.M.; Layman, C.A.; Arrington, D.A.; Takimoto, G.; Quattrochi, J.; Montaña, C.G. Getting to the fat of the matter: Models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia 2007, 152, 179–189. [Google Scholar] [CrossRef]
- Quezada-Romegialli, C.; Jackson, A.L.; Hayden, B.; Kahilainen, K.K.; Lopes, C.; Harrod, C. TRophicPosition, an r package for the Bayesian estimation of trophic position from consumer stable isotope ratios. Methods Ecol. Evol. 2018, 9, 1592–1599. [Google Scholar] [CrossRef]
- Aurioles-Gamboa, D.; Rodríguez-Pérez, M.Y.; Sánchez-Velasco, L.; Lavín, M.F. Habitat, trophic level, and residence of marine mammals in the Gulf of California assessed by stable isotope analysis. Mar. Ecol. Prog. Ser. 2013, 488, 275–290. [Google Scholar] [CrossRef]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER—Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef]
- Altabet, M.A.; Pilskaln, C.; Thunell, R.; Pride, C.; Sigman, D.; Chavez, F.; Francois, R. The nitrogen isotope biogeochemistry of sinking particles from the margin of the Eastern North Pacific. Deep. Sea Res. Part I Oceanogr. Res. Pap. 1999, 46, 655–679. [Google Scholar] [CrossRef]
- Graham, B.S.; Koch, P.L.; Newsome, S.D.; McMahon, K.W.; Aurioles, D. Using Isoscapes to Trace the Movements and Foraging Behavior of Top Predators in Oceanic Ecosystems. In Isoscapes: Understanding Movement, Pattern, and Process on Earth through Isotope Mapping; West, J., Bowen, G.J., Dawson, T.E., Tu, K.P., Eds.; Springer: Amsterdam, The Netherlands, 2009; pp. 299–318. [Google Scholar]
- García-Pámanes, J.; Trasviña-Castro, A.; Lara-Lara, J.R.; Bazán-Guzmán, C. Variaciones estacionales del flujo vertical de materia orgánica particulada en la región central del Golfo de California. Cienc. Mar. 2011, 37, 33–49. [Google Scholar]
- Guzman-del Proo, S.A.; Mille-Pagaza, S.R.; Campa-Guzmán, S.; Carrillo-Laguna, J.; Pereira Corona, A.; Belmar-Perez, J.; Parra-Alcocer, M.J.; Luque-Guerrero, A.C. La comunidad bentónica de los bancos de abulón (Haliotis spp. Mollusca: Gastropoda) en Bahía Tortugas, Baja California Sur, México. An. Esc. Nac. Cienc. Biológicas 1991, 36, 27–59. [Google Scholar]
- Newsome, S.D.; Rio, C.M.; Bearhop, S.; Phillips, D.L. A niche for isotopic ecology. Front. Ecol. Environ. 2007, 5, 429–436. [Google Scholar] [CrossRef]
- MacNeil, M.A.; Skomal, G.B.; Fisk, A.T. Stable isotopes from multiple tissues reveal diet switching in sharks. Mar. Ecol. Prog. Ser. 2005, 302, 199–206. [Google Scholar] [CrossRef]
- Kim, S.L.; Casper, D.R.; Galván-Magaña, F.; Ochoa-Díaz, R.; HernándezAguilar, S.B.; Koch, P.L. Carbon and nitrogen discrimination factors for elasmobranch soft tissues based on a long-term controlled feeding study. Environ. Biol. Fishes 2012, 95, 37–52. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vertebrae BT (n = 35) | Muscle BT (n = 32) | Vertebrae SR (n = 125) | Muscle SR (n = 43) | |||||
|---|---|---|---|---|---|---|---|---|
| δ15N | δ13C | δ15N | δ13C | δ15N | δ13C | δ15N | δ13C | |
| Neonates | 12.67 ± 1.17 | −14.6 ± 0.53 | 14.91 ± 1.59 | −13.84 ± 0.41 | ||||
| Juveniles | 12.74 ± 1.18 | −14.97 ± 0.68 | 16.83 ± 0.79 | −17.26 ± 0.46 | 14.47 ± 0.87 | −14.6 ± 0.42 | 17.78 ± 0.76 | −16.48 ± 0.29 |
| Adults | 12.79 ± 0.67 | −14.78 ± 0.59 | 16.51 ± 0.74 | −17.12 ± 0.35 | 14.89 ± 1.15 | −14.27 ± 0.51 | 17.83 ± 0.85 | −16.32 ± 0.4 |
| Females | 12.92 ± 1.21 | −14.89 ± 0.42 | 16.61 ± 0.81 | −17.19 ± 0.39 | 14.31 ± 1.64 | −14.21 ± 0.52 | 17.95 ± 0.77 | −16.42 ± 0.37 |
| Males | 12.34 ± 0.56 | −14.59 ± 0.85 | 16.66 ± 0.70 | −17.16 ± 0.43 | 14.67 ± 1.45 | −14.08 ± 0.49 | 17.44 ± 0.74 | −16.29 ± 0.33 |
| Mean | 12.72 ± 1.06 | −14.79 ± 0.61 | 16.63 ± 0.76 | −17.18 ± 0.39 | 14.4 ± 1.59 | −14.18 ± 0.51 | 18.08 ± 0.96 | −16.43 ± 0.34 |
| Vertebra BT | Muscle BT | Vertebra SR | Muscle SR | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| δ15N | δ13C | δ15N | δ13C | δ15N | δ13C | δ15N | δ13C | |||||||||
| F | p | X2 | p | F | p | F | p | X2 | p | X2 | p | F | p | F | p | |
| Stage | 0.22 | 0.64 | 2.97 | 0.23 | 1.59 | 0.22 | 1.19 | 0.28 | 1.08 | 0.58 | 19.42 | 6.08 × 10−5 | 2.93 | 0.09 | 0.38 | 0.54 |
| Sex | 2.37 | 0.13 | 0.09 | 0.75 | 0.02 | 0.88 | 0.007 | 0.93 | 2.21 | 0.14 | 1.11 | 0.30 | 5.68 | 0.02 | 0.042 | 0.84 |
| Vertebra BT | Muscle BT | Vertebra SR | Muscle SR | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SEAc | Overlap | SEAc | Overlap | SEAc | Overlap | SEAc | Overlap | |||
| Neonates | 1.9 | 48 | 1.79 | 49 | ||||||
| Juveniles | 2.74 | 41 | 1.24 | 47 | 3.17 | 54 | 0.78 | 48 | ||
| Adults | 1.4 | 0.86 | 1.94 | 1.27 | ||||||
| Males | 1.1 | 27 | 0.83 | 44 | 2.28 | 68 | 1.31 | 46 | ||
| Females | 1.67 | 1 | 2.68 | 0.9 | ||||||
| Overlap of neonates and adults: 41% | Overlap of neonates and adults: 55% | |||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Latorre, C.; Galván-Magaña, F.; Elorriaga-Verplancken, F.R.; Tripp-Valdez, A.; González-Armas, R.; Piñón-Gimate, A.; Delgado-Huertas, A. Trophic Ecology during the Ontogenetic Development of the Pelagic Thresher Shark Alopias pelagicus in Baja California Sur, Mexico. Diversity 2023, 15, 1057. https://doi.org/10.3390/d15101057
Sánchez-Latorre C, Galván-Magaña F, Elorriaga-Verplancken FR, Tripp-Valdez A, González-Armas R, Piñón-Gimate A, Delgado-Huertas A. Trophic Ecology during the Ontogenetic Development of the Pelagic Thresher Shark Alopias pelagicus in Baja California Sur, Mexico. Diversity. 2023; 15(10):1057. https://doi.org/10.3390/d15101057
Chicago/Turabian StyleSánchez-Latorre, Clara, Felipe Galván-Magaña, Fernando R. Elorriaga-Verplancken, Arturo Tripp-Valdez, Rogelio González-Armas, Alejandra Piñón-Gimate, and Antonio Delgado-Huertas. 2023. "Trophic Ecology during the Ontogenetic Development of the Pelagic Thresher Shark Alopias pelagicus in Baja California Sur, Mexico" Diversity 15, no. 10: 1057. https://doi.org/10.3390/d15101057