
Trophic Partitioning among Three Mesopredatory Shark Species Inhabiting the Northwestern Adriatic Sea

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Stomach Content Analysis
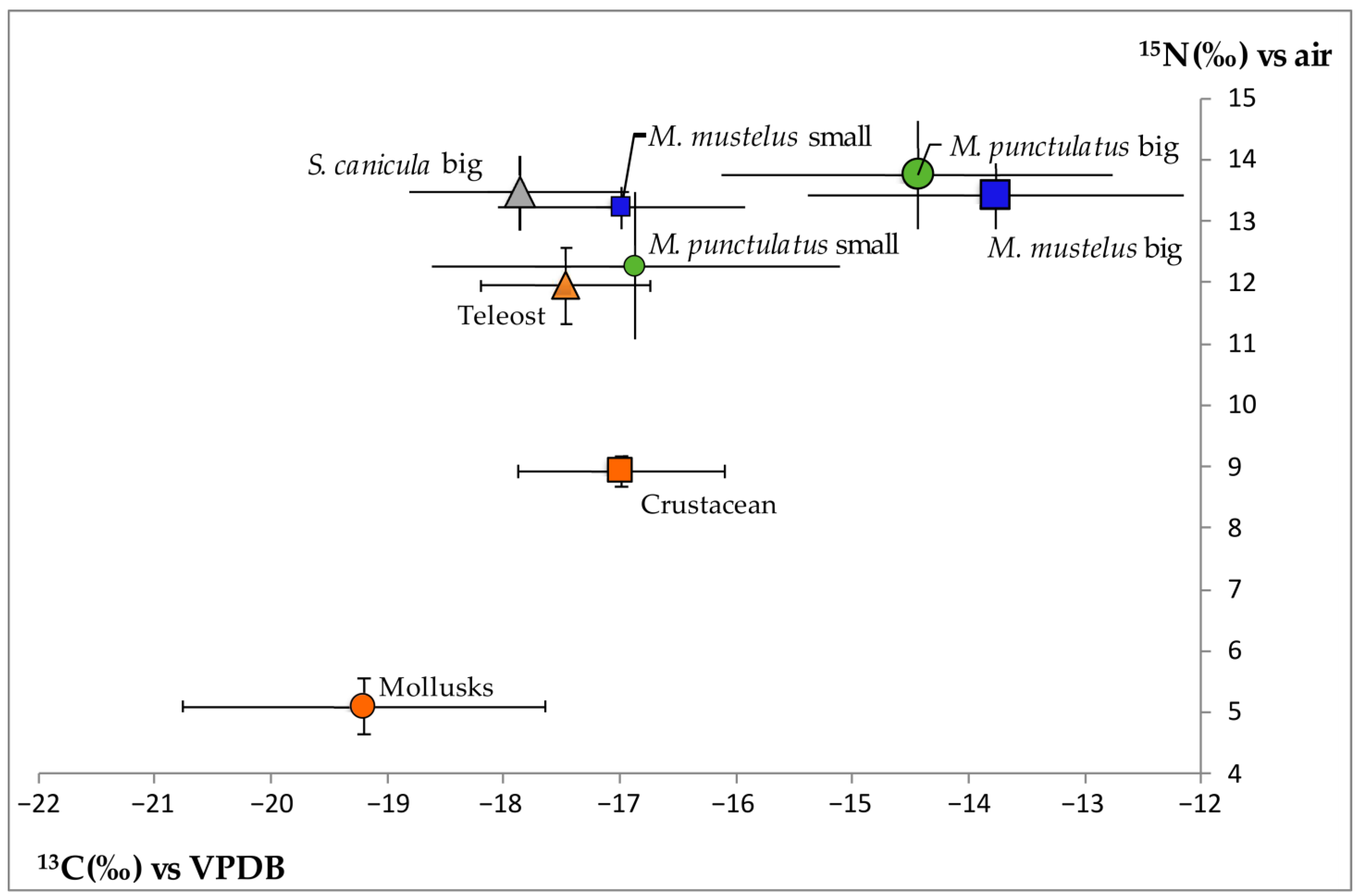
2.2. Stable Isotope Analysis
2.3. Data Analysis
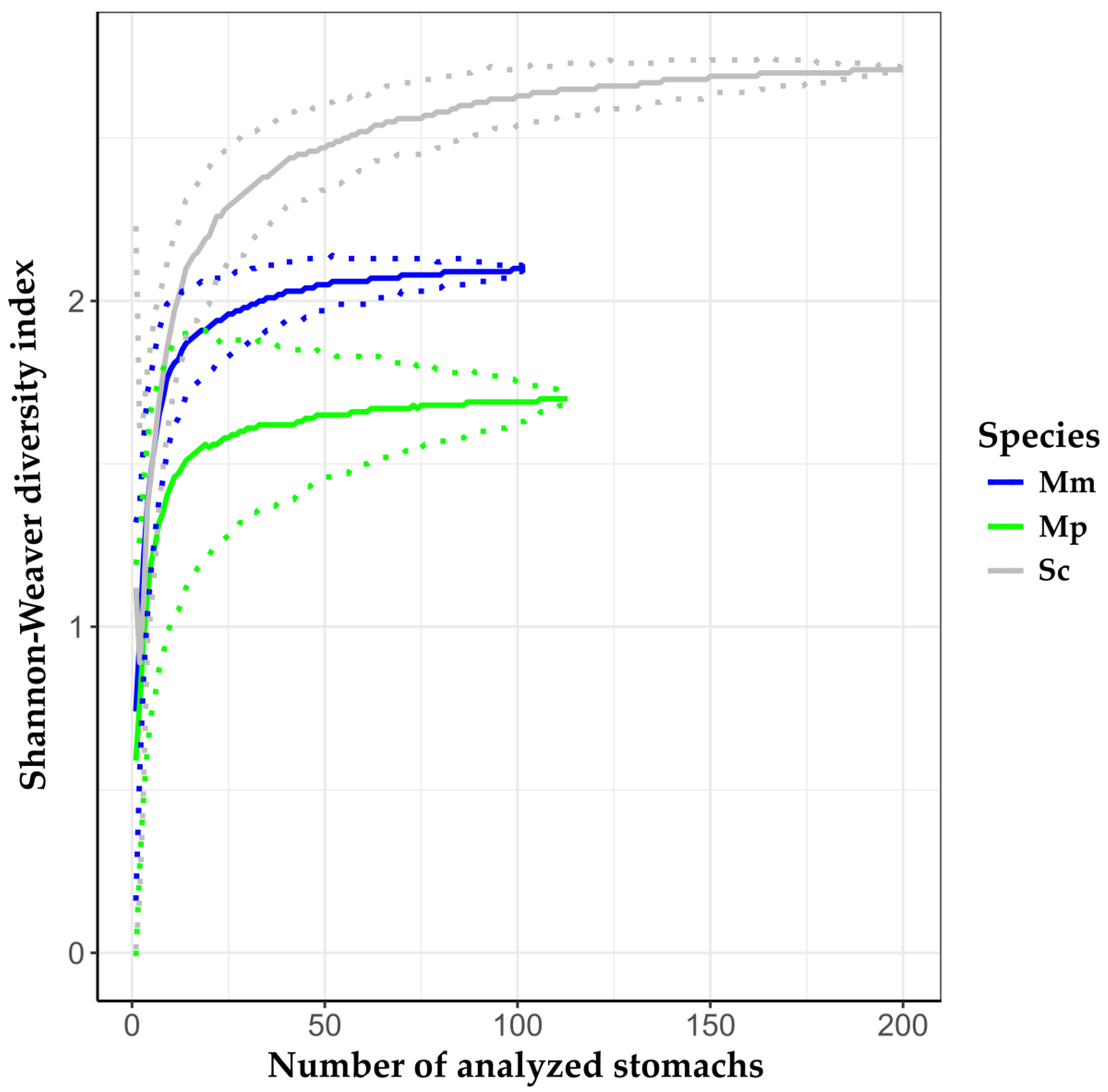
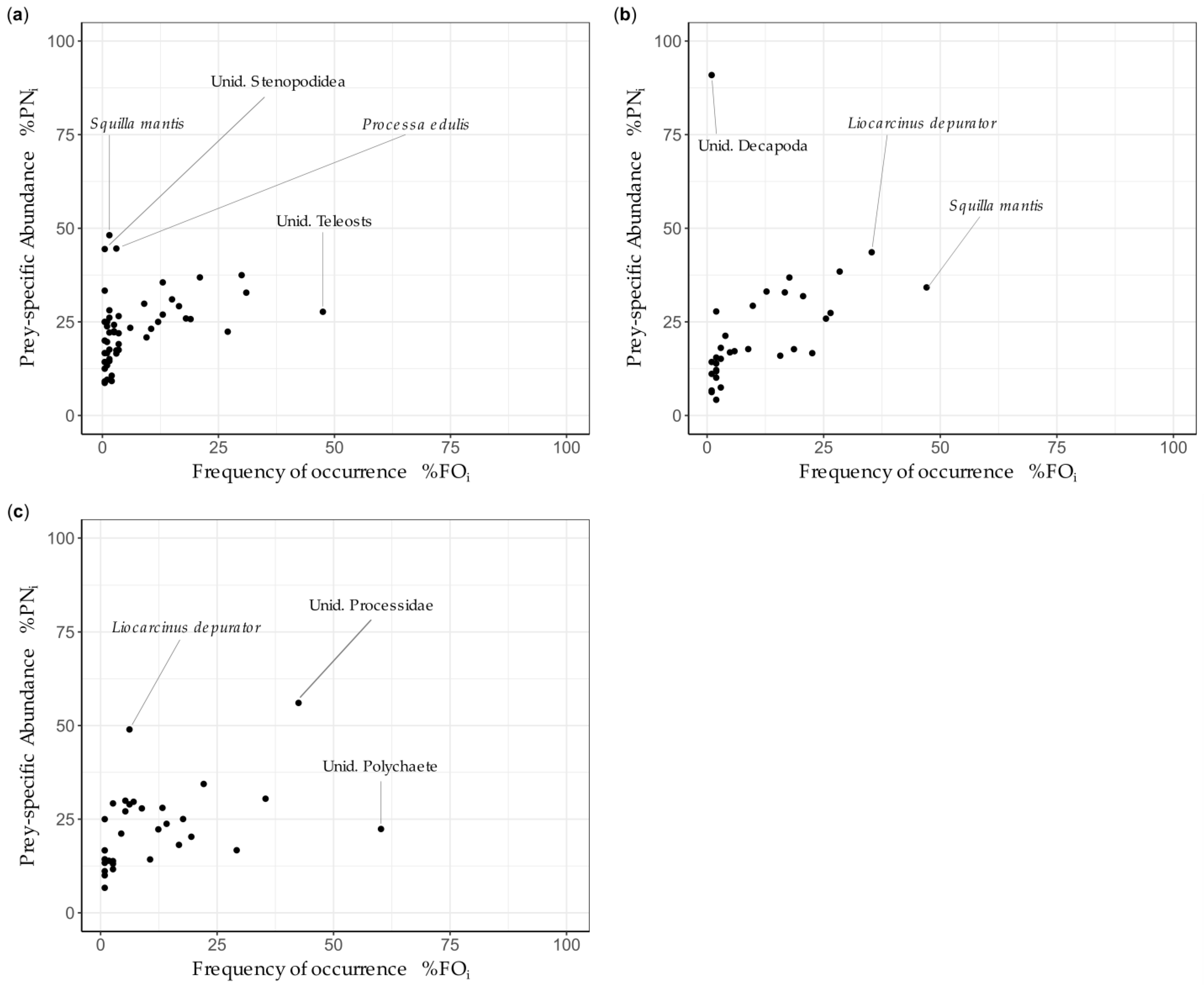
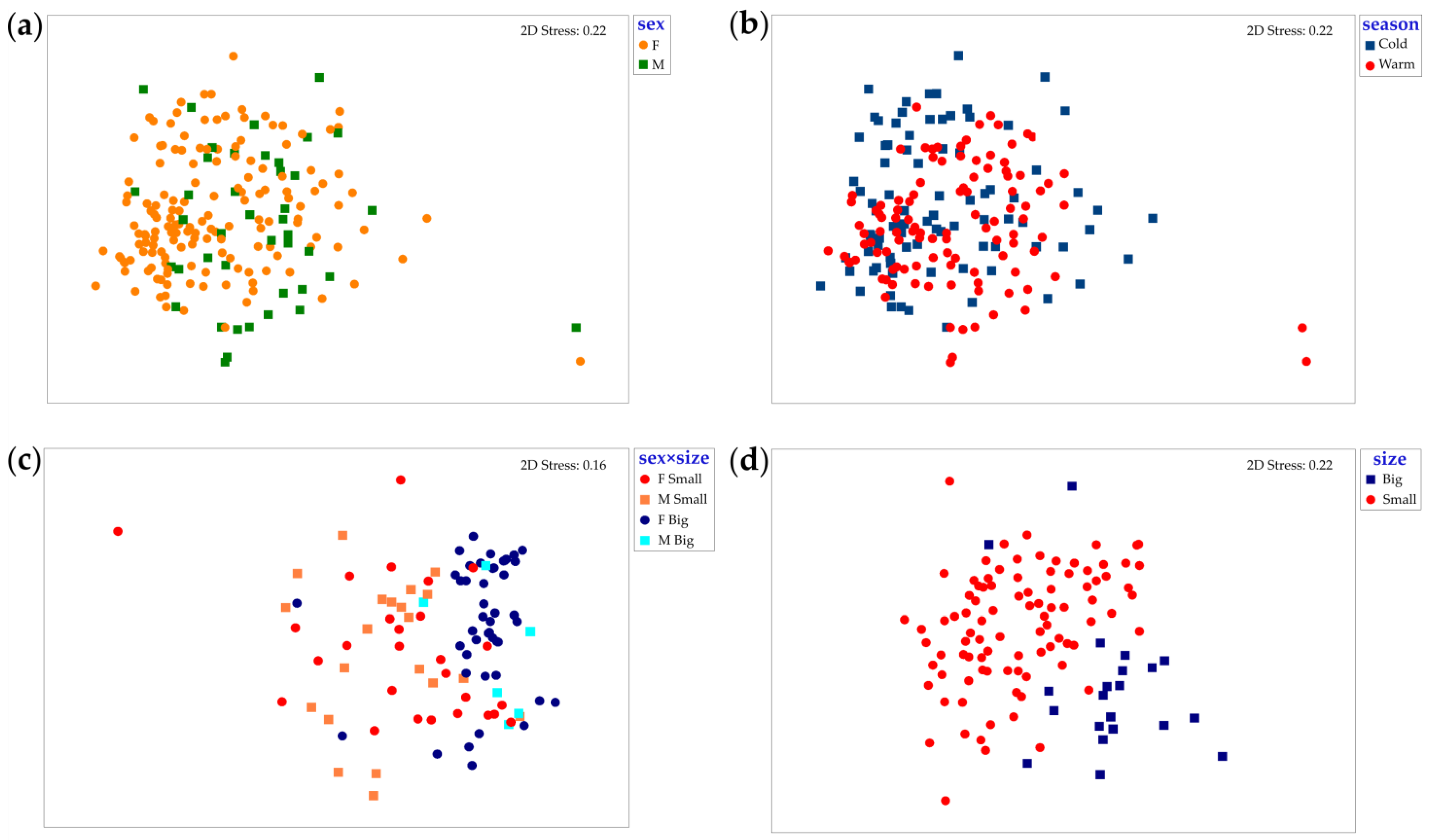
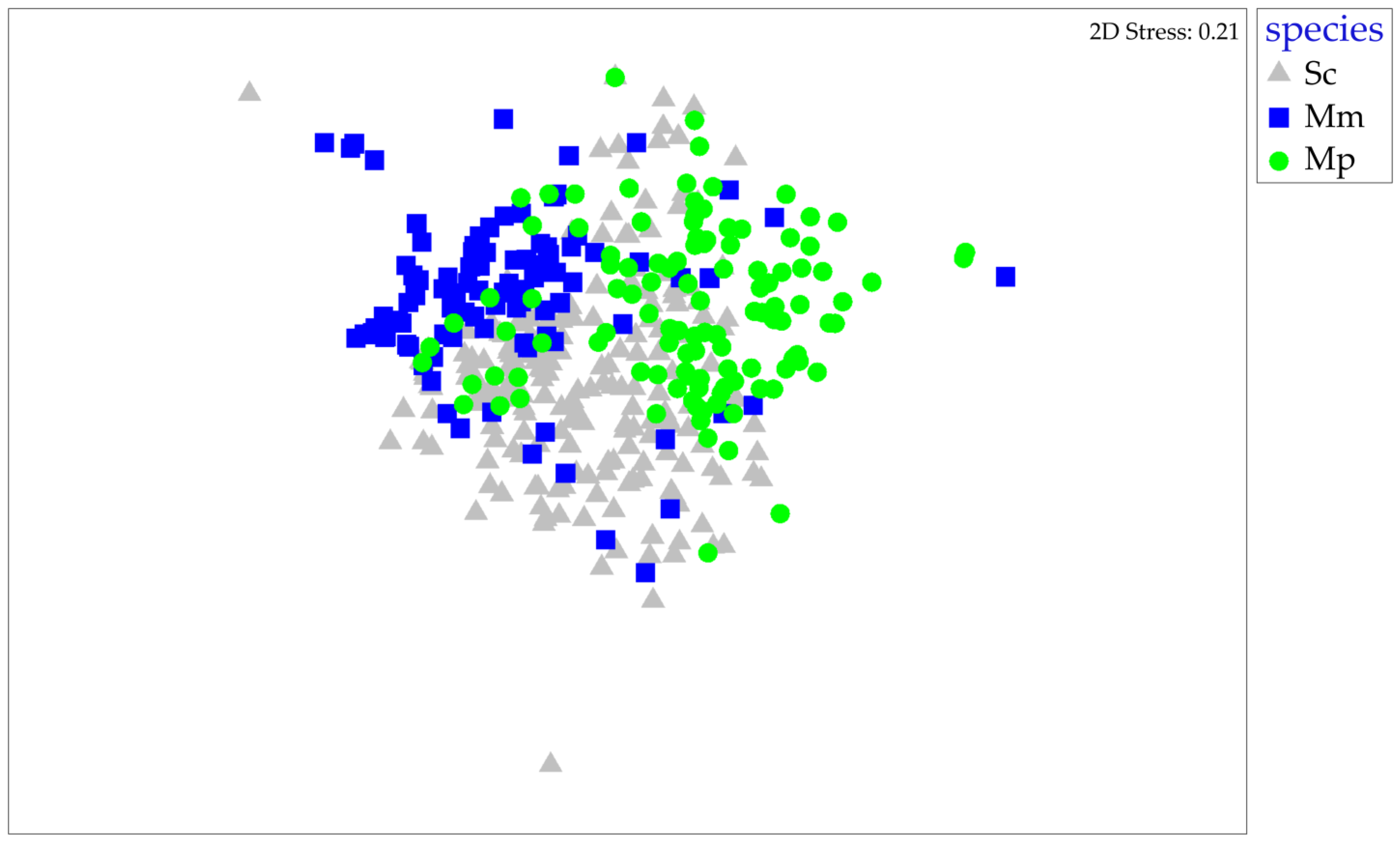
3. Results
3.1. Intraspecific Analysis
3.1.1. Scyliorhinus canicula
3.1.2. Mustelus mustelus
3.1.3. Mustelus punctulatus
3.2. Interspecific Comparison
3.3. Stable Isotope
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krebs, C.J. Ecology; Benjamin Cummings: San Francisco, CA, USA, 2001. [Google Scholar]
- Langeland, A.; L’abée-lund, J.H.; Jonsson, B.; Jonsson, N. Resource partitioning and niche shift in Arctic charr Savelinus alpinus and brown trout Salmo trutta. J. Anim. Ecol. 1991, 60, 895–912. [Google Scholar] [CrossRef]
- Ross, S.T. Resource partitioning in fish assemblages: A review of field studies. Copeia 1986, 1986, 352–388. [Google Scholar] [CrossRef]
- Sibbing, F.A.; Nagelkerke, L.A.J. Resource partitioning by Lake Tana barbs predicted from fish morphometrics and prey characteristics. Rev. Fish Biol. Fish. 2001, 10, 393–437. [Google Scholar] [CrossRef]
- Fanelli, E.; Badalamenti, F.; D’Anna, G.; Pipitone, C.; Riginella, E.; Azzurro, E. Food partitioning and diet temporal variation in two coexisting sparids, Pagellus erythrinus and Pagellus acarne. J. Fish Biol. 2011, 78, 869–900. [Google Scholar] [CrossRef] [PubMed]
- Bornatowski, H.; Navia, A.F.; Braga, R.R.; Abilhoa, V.; Corrêa, M.F.M. Ecological importance of sharks and rays in a structural foodweb analysis in southern Brazil. ICES J. Mar. Sci. 2014, 71, 1586–1592. [Google Scholar] [CrossRef]
- Papastamatiou, Y.P.; Wetherbee, B.M.; Lowe, C.J.; Crow, G.L. Distribution and diet of four species of carcharhinid shark in the Hawaiian Islands: Evidence for resource partitioning and competitive exclusion. Mar. Ecol. Prog. Ser. 2006, 320, 239–251. [Google Scholar] [CrossRef]
- White, W.T.; Platell, M.E.; Potter, I.C. Comparisons between the diets of four abundant species of elasmobranchs in a subtropical embayment: Implications for resource partitioning. Mar. Biol. 2004, 144, 439–448. [Google Scholar] [CrossRef]
- Bizzarro, J.J.; Carlisle, A.B.; Smith, W.D.; Cortés, E. Diet composition and trophic ecology of Northeast Pacific Ocean sharks. Adv. Mar. Biol. 2017, 77, 111–148. [Google Scholar]
- Navia, A.F.; Cortés, E.; Mejía-Falla, P. Topological analysis of the ecological importance of elasmobranch fishes: A food web study on the Gulf of Tortugas, Colombia. Ecol. Model. 2010, 221, 2918–2926. [Google Scholar] [CrossRef]
- Cortés, E. Standardized diet compositions and trophic levels of sharks. J. Mar. Sci. 1999, 56, 707–717. [Google Scholar] [CrossRef]
- Heithaus, M.R. Predator–prey interactions. In Biology of Sharks and Their Relatives; Carrier, C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Lucas, Z.; Stobo, W.T. Shark inflicted mortality on a population of harbour seals (Phoca vitulina) at Sable Island, Nova Scotia. J. Zool. 2000, 252, 405–414. [Google Scholar] [CrossRef]
- Ellis, J.R.; Pawson, M.G.; Shackley, S.E. The comparative feeding ecology of six species of shark and four species of ray (Elasmobranchii) in the North-East Atlantic. J. Mar. Biol. Assoc. UK 1996, 76, 89–106. [Google Scholar] [CrossRef]
- Ferretti, F.; Worm, B.; Britten, G.L.; Heithaus, M.R.; Lotze, H.K. Patterns and ecosystem consequences of shark declines in the ocean. Ecol. Lett. 2010, 13, 1055–1071. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, E.G.; Johnson, C.N. Predator interactions, mesopredator release and biodiversity conservation. Ecol. Lett. 2009, 12, 982–998. [Google Scholar] [CrossRef]
- Jacobsen, I.P.; Bennett, M.B. A comparative analysis of feeding and trophic level ecology in stingrays (Rajiformes: Myliobatoidei) and electric rays (Rajiformes: Torpedinoidei). PLoS ONE 2013, 8, e71348. [Google Scholar] [CrossRef]
- Ebert, D.A.; Cowley, P.D. Diet, feeding behaviour and habitat utilisation of the blue stingray Dasyatis chrysonata (Smith, 1828) in south African waters. Mar. Freshw. Res. 2003, 54, 957–965. [Google Scholar] [CrossRef]
- Munroe, S.E.M.; Heupel, M.R.; Aaron, T.; Fisk, A.; Simpfendorfer, C.A. Geographic and temporal variation in the trophic ecology of a small bodied shark: Evidence of resilience to environmental change. Can. J. Fish. Aquat. Sci. 2015, 72, 343–351. [Google Scholar] [CrossRef]
- Valls, M.; Quetglas, A.; Ordines, F.; Moranta, J. Feeding ecology of demersal elasmobranchs from the shelf and slope off the Balearic Sea (western Mediterranean). Sci. Mar. 2011, 75, 633–639. [Google Scholar] [CrossRef]
- Chipps, S.R.; Garvey, J.E. Assessment of food habits and feeding patterns. In Analysis and Interpretation of Freshwater Fisheries Data; Guy, C.S., Brown, M.L., Eds.; American Fisheries Society: Bethesda, MD, USA, 2007. [Google Scholar]
- Shiffman, D.S.; Gallagher, A.J.; Boyle, M.D.; Hammerschlag-Peyer, C.M.; Hammerschlag, N. Stable isotope analysis as a tool for elasmobranch conservation research: A primer for non-specialists. Mar. Freshw. Res. 2012, 63, 635–643. [Google Scholar] [CrossRef]
- Barría, C.; Coll, M.; Navarro, J. Unravelling the ecological role and trophic relationships of uncommon and threatened elasmobranchs in the western Mediterranean Sea. Mar. Ecol. Prog. Ser. 2015, 539, 225–240. [Google Scholar] [CrossRef]
- Fonda Umani, S. Pelagic production and biomass in the Adriatic Sea. Sci. Mar. 1996, 60, 6–77. [Google Scholar]
- Giani, M.; Djakovac, T.; Degobbis, D.; Cozzi, S.; Solidoro, C.; Fonda Umani, S. Recent changes in the marine ecosystems of the northern Adriatic Sea. Estuar. Coast. Shelf Sci. 2012, 115, 1–13. [Google Scholar] [CrossRef]
- Barausse, A.; Correale, V.; Curkovic, A.; Finotto, L.; Riginella, E.; Visentin, E.; Mazzoldi, C. The role of fisheries and the environment in driving the decline of elasmobranchs in the Northern Adriatic Sea. ICES J. Mar. Sci. 2014, 71, 1593–1603. [Google Scholar] [CrossRef]
- Fortibuoni, T.; Giovanardi, O.; Pranovi, F.; Raicevich, S.; Solidoro, C.; Libralato, S. Analysis of long-term changes in a Mediterranean marine ecosystem based on fishery landings. Front. Mar. Sci. 2017, 4, 33. [Google Scholar] [CrossRef]
- Relini, G.; Mannini, A.; De Ranieri, S.; Bitetto, I.; Follesa, M.C.; Gancitano, V.; Manfredi, C.; Casciaro, L.; Sion, L. Chondrichthyes caught during the MEDITS surveys in Italian waters. Biol. Mar. Mediterr. 2010, 17, 186–204. [Google Scholar]
- Ferretti, F.; Osio, G.C.; Jenkins, C.J.; Rosenberg, A.A.; Lotze, H.K. Long-term change in a meso-predator community in response to prolonged and heterogeneous human impact. Sci. Rep. Cetacean Res. 2013, 3, 1057. [Google Scholar] [CrossRef] [PubMed]
- Compagno, L.J.V. Sharks of the World: An Annotated and Illustrated Catalogue of Shark Species Known to Date, Part 2, Carcharhiniformes; FAO: Rome, Italy, 1984. [Google Scholar]
- Di Lorenzo, M.; Vizzini, S.; Signa, G.; Andolina, C.; Palo, G.B.; Gristina, M.; Mazzoldi, C.; Colloca, F. Ontogenetic trophic segregation between two threatened smoothhound sharks in the Central Mediterranean Sea. Sci. Rep. 2020, 10, 11011. [Google Scholar] [CrossRef]
- Kousteni, V.; Karachle, P.K.; Megalofonou, P. Diet of the small-spotted catshark Scyliorhinus canicula in the Aegean Sea (eastern Mediterranean). Mar. Biol. Res. 2017, 13, 161–173. [Google Scholar] [CrossRef]
- Finotto, L.; Gristina, M.; Garofalo, G.; Riginella, E.; Mazzoldi, C. Contrasting life history and reproductive traits in two populations of Scyliorhinus canicula. Mar. Biol. 2015, 162, 1175–1186. [Google Scholar] [CrossRef]
- Rodríguez-Cabello, C.; Sánchez, F.; Olaso, I. Distribution patterns and sexual segregations of Scyliorhinus canicula (L.) in the Cantabrian Sea. J. Fish Biol. 2007, 70, 1568–1586. [Google Scholar] [CrossRef]
- Sims, D.W. Differences in habitat selection and reproductive strategies of male and female sharks. In Sexual Segregation in Vertebrates: Ecology of the Two Sexes; Ruckstuhl, K.E., Neuhaus, P., Eds.; Cambridge University Press: Cambridge, UK, 2005; p. 488. [Google Scholar]
- Riginella, E.; Correale, V.; Marino, I.A.M.; Rasotto, M.B.; Vrbatovic, A.; Zane, L.; Mazzoldi, C. Contrasting life-history traits of two sympatric smooth-hound species: Implication for vulnerability. J. Fish Biol. 2020, 96, 853–857. [Google Scholar] [CrossRef]
- Marino, I.A.M.; Finotto, L.; Colloca, F.; Di Lorenzo, M.; Gristina, M.; Farrell, E.D.; Zane, L.; Mazzoldi, C. Resolving the ambiguities in the identification of two smooth-hound sharks (Mustelus mustelus and Mustelus punctulatus) using genetics and morphology. Mar. Biodivers. 2018, 48, 1551–1562. [Google Scholar] [CrossRef]
- Filiz, H. Diet composition of smooth-hound, Mustelus mustelus (Linnaeus, 1758), in Aegean Sea, Turkey. Belg. J. Zool. 2009, 139, 81–84. [Google Scholar]
- Gravino, F.; Dimech, M.; Schembri, P.J. Feeding Habits of the Small-Spotted Catshark Scyliorhinus Canicula (L. 1758) in the Central Mediterranean. Paper presented at the Rapport du Congrès de la Commission Internationale pour l’Exploration Scientifique de la Mer Méditerranée. CIESM Meet. Rapp. Comm. Int. Mer Médit. 2010, 39, 1. [Google Scholar]
- Saïdi, B.; Bradaï, M.N.; Bouaïn, A. Reproductive biology and diet of Mustelus punctulatus (Risso, 1826) (Chondrichthyes: Triakidae) from the Gulf of Gabès, central Mediterranean Sea. Sci. Mar. 2009, 73, 249–258. [Google Scholar] [CrossRef]
- Saïdi, B.; Enajjar, S.; Bradaï, M.N.; Bouaïn, A. Diet composition of smooth-hound shark, Mustelus mustelus (Linnaeus, 1758), in the Gulf of Gabès, southern Tunisia. J. Appl. Ichthyol. 2009, 25, 113–118. [Google Scholar] [CrossRef]
- Gračan, R.; Mladineo, I.; Lazar, B. Insight into the diet composition and gastrointestinal parasite community of the common smooth-hound, Mustelus mustelus (Carcharhiniformes: Triakidae), in the northern Adriatic Sea. Nat. Croat. 2014, 23, 35–44. [Google Scholar]
- Jardas, I.; Šantić, M.; Nerlović, N.; Pallaoro, A. Diet composition of blackspotted smooth-hound, Mustelus punctulatus (Risso, 1826), in the eastern Adriatic Sea. J. Appl. Ichthyol. 2007, 23, 279–281. [Google Scholar] [CrossRef]
- Jardas, I.; Šantić, M.; Nerlović, N.; Pallaoro, A. Diet of the smooth-hound, Mustelus mustelus (Chondrichthyes: Triakidae), in the eastern Adriatic Sea. Cybium Int. J. Ichthyol. 2007, 31, 459–464. [Google Scholar]
- Lipej, L.; Mavrič, B.; Rešek, S.; Chérif, M.; Capapé, C. Food and feeding habits of the blackspotted smooth-hound, Mustelus punctulatus (Elasmobranchii: Carcharhiniformes: Triakidae), from the northern Adriatic. Acta Ichthyol. Piscat. 2011, 41, 171–177. [Google Scholar] [CrossRef]
- Šantić, M.; Rađa, B.; Pallaoro, A. Feeding habits of small-spotted catshark (Scyliorhinus canicula Linnaeus, 1758) from the eastern central Adriatic Sea. Mar. Biol. Res. 2012, 8, 1003–1011. [Google Scholar] [CrossRef]
- Barausse, A.; Michieli, A.; Riginella, E.; Palmeri, L.; Mazzoldi, C. Long-term changes in community composition and lifehistory traits in a highly exploited basin (northern Adriatic Sea): The role of environment and anthropogenic pressures. J. Fish Biol. 2011, 79, 1453–1486. [Google Scholar] [CrossRef]
- AFORO. Available online: http://aforo.cmima.csic.es/index.jsp (accessed on 14 July 2023).
- Falciai, L.; Minervini, R. Guida dei Crostacei Decapodi d’Europa; Franco Muzzio Editore: Padova, Italy, 1992; p. 282. [Google Scholar]
- Fischer, W.; Bauchot, M.L.; Schneider, M. Fiches FAO D’identification des Espèces Pour les Besoins de la Pêche. Méditerranée et Mer Noire. Zone de Pêche 37 (Vol. Volume I. Végétaux et Invertébrés); FAO: Rome, Italy, 1987. [Google Scholar]
- Louisy, P. Guida all’identificazione dei Pesci Marini d’Europa e del Mediterraneo. (Ed. italiana a cura di Trainito E.); Il Castello: Perugia, Italy, 2006. [Google Scholar]
- Riedl, R. Fauna e Flora del Mediterraneo; Franco Muzzio Editore: Padova, Italy, 1991. [Google Scholar]
- Šoljan, T. I pesci dell’Adriatico; Arnoldo Mondadori Editore: Verona, Italy, 1975. [Google Scholar]
- Tortonese, E.; Lanza, B. Piccola Fauna Italiana: Pesci, Anfibi e Rettili; Martello Editore: Milano, Italy, 1968. [Google Scholar]
- Whitehead, P.J.P.; Bauchot, M.L.; Hureau, J.C.; Nielsen, J.; Tortonese, E. Fishes of the North-Eastern Atlantic and Mediterranean (Vol. 1, 2, 3); UNESCO: Paris, France, 1986. [Google Scholar]
- Colwell, R.K.; EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. Version 9. Available online: http://purl.oclc.org/estimates (accessed on 14 July 2023).
- Bizzarro, J.J.; Robison, H.J.; Rinewalt, C.S.; Ebert, D.A. Comparative feeding ecology of four sympatric skate species off central California, USA. Environ. Biol. Fishes 2007, 80, 197–220. [Google Scholar] [CrossRef]
- Ferry, L.A.; Cailliet, G.M. Sample Size and Data Analysis: Are We Characterizing and Comparing Diet Properly? In Proceedings of the Feeding Ecology and Nutrition in Fish, Symposium Proceedings, San Francisco, CA, USA, 14–18 July 1996. [Google Scholar]
- Hyslop, E.J. Stomach contents analysis—A review of methods and their application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; Urbans, University of Illionis Press: Champaign, IL, USA, 1949. [Google Scholar]
- Fanelli, E.; Badalamenti, F.; D’Anna, G.; Pipitone, C.; Romano, C. Trophodynamic effects of trawling on the feeding ecology of pandora, Pagellus erythrinus, off the northern Sicily coast (Mediterranean Sea). Mar. Freshw. Res. 2010, 61, 408–417. [Google Scholar] [CrossRef]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Brown, S.C.; Bizzarro, J.J.; Cailliet, G.M.; Ebert, D.A. Breaking with tradition: Redefining measures for diet description with a case study of the Aleutian skate Bathyraja aleutica (Gilbert 1896). Environ. Biol. Fishes 2012, 95, 3–20. [Google Scholar] [CrossRef]
- Amundsen, P.A.; Gabler, H.M.; Staldvik, F.J. A new approach to graphical analysis of feeding strategy from stomach contents data—Modification of the Costello (1990) method. J. Fish Biol. 1996, 48, 607–614. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecological Methodology; Harper & Row Publishers: New York, NY, USA, 1989. [Google Scholar]
- Langton, R.S. Diet overlap between the Atlantic cod, Gadus morhua, silver hake, Merluccius bilinearis and fifteen other northwest Atlantic finfish. Fish. Bull.-Natl. Ocean. Atmos. Adm. 1982, 80, 745–759. [Google Scholar]
- Ehrich, D.; Tarroux, A.; Stien, J.; Lecomte, N.; Killengreen, S.; Berteaux, D.; Yoccoz, N.G. Stable isotope analysis: Modelling lipid normalization for muscle and eggs from arctic mammals and birds. Methods Ecol. Evol. 2011, 2, 66–76. [Google Scholar] [CrossRef]
- Fortibuoni, T.; Noventa, S.; Rampazzo, F.; Gion, C.; Formalewicz, M.; Berto, D.; Raicevich, S. Evidences of butyltins biomagnification along the Northern Adriatic food web (Mediterranean Sea) elucidated by stable isotopes ratios. Environ. Sci. Technol. 2013, 47, 3370–3377. [Google Scholar] [CrossRef] [PubMed]
- Simpfendorfer, C.A.; Goodreid, A.B.; McAuley, R.B. Size, sex and geographic variation in the diet of the tiger shark, Galeocerdo cuvier, from Western Australian waters. Environ. Biol. Fishes 2001, 61, 37–46. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Ellison, A.M. A Primer of Ecological Statistics, 2nd ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2004. [Google Scholar]
- Yemisken, E.; Forero, M.G.; Megalofonou, P.; Eryilmaz, L.; Navarro, J. Feeding habits of three Batoids in the Levantine Sea (north-eastern Mediterranean Sea) based on stomach content and isotopic data. J. Mar. Biol. Assoc. UK 2017, 98, 89–96. [Google Scholar] [CrossRef]
- Pagliarini, C.D.; da Silva Riberiro, C.; Spada, L.; Delariva, R.L.; Araújo Chagas, J.M.; dos Anjos, L.A.; Ramos, I.P. Trophic ecology and metabolism of two species of nonnative freshwater stingrays (Chondrichthyes: Potamotrygonidae). Hydrobiologia 2020, 847, 2895–2908. [Google Scholar] [CrossRef]
- Gonzalez-Pestana, A.; Silva-Garay, L.; Quiñones, J.; Mayaute, L.; Manrique, M.; Segura-Cobeña, E.; Espinoza, P.; Moscoso, V.; Vélez-Zuazo, X.; Alfaro-Shigueto, J.; et al. Geographic and ontogenetic variation in the diet of two commonly exploited batoids (Chilean eagle ray and Pacific guitarfish) off Peru: Evidence of trophic plasticity. Environ. Biol. Fishes 2021, 104, 1525–1540. [Google Scholar] [CrossRef]
- Fanelli, E.; Da Ros, Z.; Martino, I.; Azzurro, E.; Bargione, G.; Donato, F.; Lucchetti, A. Crowding in the middle of marine food webs: A focus on Raja asterias and other mediterranean batoids. Mar. Environ. Res. 2022, 183, 105830. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R.V. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef]
- Lenth, R.V. Emmeans: Estimated Marginal Means, aka Least-Squares Means. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 14 July 2023).
- Wieczorek, A.M.; Power, A.M.; Browne, P.; Graham, C.T. Stable-isotope analysis reveals the importance of soft-bodied prey in the diet of lesser spotted dogfish Scyliorhinus canicula. J. Fish Biol. 2017, 93, 685–693. [Google Scholar] [CrossRef]
- Olaso, I.; Sánchez, F.; Rodríguez-Cabello, C.; Velasco, F. The feeding behaviour of some demersal fish species in response to artificial discarding. Sci. Mar. 2002, 66, 301–311. [Google Scholar] [CrossRef]
- Eales, N.B. The food of the dogfish, Scyliorhinus caniculus L. J. Mar. Biol. Assoc. UK 1949, 28, 791–793. [Google Scholar] [CrossRef]
- Henderson, A.C.; Dunne, J. Food of the lesser-spotted dogfish Scyliorhinus canicula (L.) in Galway Bay. Ir. Nat. J. 1999, 26, 191–194. [Google Scholar]
- Martinho, F.; Sá, C.; Falcão, J.; Cabral, H.N.; Pardal, M.A. Comparative feeding ecology of two elasmobranch species, Squalus blainville and Scyliorhinus canicula, off the coast of Portugal. Fish. Bull. 2012, 110, 71–84. [Google Scholar]
- Olaso, I.; Velasco, F.; Pérez, N. Importance of discarded blue whiting (Micromesistius poutassou) in the diet of lesser spotted dogfish (Scyliorhinus canicula) in the Cantabrian Sea. ICES J. Mar. Sci. 1998, 55, 331–341. [Google Scholar] [CrossRef]
- Olaso, I.; Velasco, F.; Sánchez, F.; Serrano, A.; Rodríguez-Cabello, C.; Cendrero, D. Trophic relations of lesser-spotted catshark (Scyliorhinus canicula) and blackmouth catshark (Galeus melastomus) in the Cantabrian Sea. J. Northwest Atl. Fish. Sci. 2005, 36, 481–494. [Google Scholar] [CrossRef]
- Saldanha, L.; Almeida, A.J.; Andrade, F.; Guerreiro, J. Observations on the diet of some slope dwelling fishes of southern Portugal. Int. Rev. Der Gesamten Hydrobiol. Hydrogr. 1995, 80, 217–234. [Google Scholar] [CrossRef]
- Serrano, A.; Velasco, F.; Olaso, I.; Sánchez, F. Macrobenthic crustaceans in the diet of demersal fish in the Bay of Biscay in relation to abundance in the environment. Sarsia 2003, 88, 36–48. [Google Scholar] [CrossRef]
- Smale, M.J.; Compagno, L.J.V. Life history and diet of two southern African smoothhound sharks, Mustelus mustelus (Linnaeus, 1758) and Mustelus palumbes Smith, 1957 (Pisces: Triakidae). S. Afr. J. Mar. Sci. 1997, 18, 229–248. [Google Scholar] [CrossRef]
- Kara, A.F.; Al Hajaji, M.; Ghmati, H.; Shakman, E.A. Food and feeding habits of Mustelus mustelus (Linnaeus, 1758) (Chondrichthyes: Triakidae) along the Western coast of Libya. Ann. Ser. Hist. Nat. 2019, 29, 197–204. [Google Scholar] [CrossRef]
- Karachle, P.K.; Stergiou, K.I. Food and Feeding Habits of Nine Elasmobranch Species in the Aegean Sea. Rapp. Comm. Int. Mer. Médit. 2010, 39, 553. [Google Scholar]
- Jardas, I. Supplement to the knowledge of ecology of some Adriatic cartilaginous fishes (Chondrichthyes) with special reference to nutrition. Act. Adriat. 1972, 14, 1–60. [Google Scholar]
- Boscolo Palo, G.; Di Lorenzo, M.; Gancitano, S.; Ragonese, S.; Mazzoldi, C.; Colloca, F. Sharks do not always grow slowly: Tagging data reveal a different pattern of growth, longevity and maturity for threatened smooth-hounds in the central Mediterranean Sea. J. Mar. Sci. Eng. 2022, 10, 1647. [Google Scholar] [CrossRef]
- Ivory, P.; Jeal, F.; Nolan, C.P. Age determination, growth and reproduction in the lesser-spotted dogfish, Scyliorhinus canicula (L.). J. Northwest Atl. Fish. Sci. 2004, 35, 89–106. [Google Scholar] [CrossRef]
- Chabot, D.; Steffensen, J.F.; Farrell, A.P. The determination of standard metabolic rate in fishes. J. Fish Biol. 2016, 88, 81–121. [Google Scholar] [CrossRef]
- Whitney, N.M.; Lear, K.O.; Gaskins, L.C.; Gleiss, A.C. The effects of temperature and swimming speed on the metabolic rate of the nurse shark (Ginglymostoma cirratum, Bonaterre). J. Exp. Mar. Biol. Ecol. 2016, 477, 40–46. [Google Scholar] [CrossRef]
- Ellis, J.R.; Shackley, S.E. Ontogenic changes and sexual dimorphism in the head, mouth and teeth of the lesser spotted dogfish. J. Fish Biol. 1995, 47, 155–164. [Google Scholar] [CrossRef]
- Wilga, C.D.; Motta, P.J.; Sanford, C.P. Evolution and ecology of feeding in elasmobranchs. Int. Comp. Biol. 2007, 47, 55–69. [Google Scholar] [CrossRef]
- Kolmann, M.A.; Huber, D.R.; Motta, P.J.; Grubbs, R.D. Feeding biomechanics of the cownose ray, Rhinoptera bonasus, over ontogeny. J. Anat. 2015, 227, 341–351. [Google Scholar] [CrossRef]
- Kemper, J.M.; Bizzarro, J.J.; Ebert, D.A. Dietary variability in two common Alaskan skates (Bathyraja interrupta and Raja rhina). Mar. Biol. 2017, 164, 52. [Google Scholar] [CrossRef]
- Colloca, F.; Enea, M.; Ragonese, S.; Di Lorenzo, M. A century of fishery data documenting the collapse of smooth-hounds (Mustelus spp.) in the Mediterranean Sea. Aquat. Conserv. Mar. Freshw. 2017, 27, 1145–1155. [Google Scholar] [CrossRef]
- Herrel, A.; Gibb, A.C. Ontogeny of performance in vertebrates. Physiol. Biochem. Zool. 2006, 79, 1–6. [Google Scholar] [CrossRef]
- Bressan, G. Studio Morfologico Della Pinna Caudale di Trentatré Specie di Squali (Elasmobranchii). Bachelor’s Thesis, University of Padova, Padova, Italy, 2019. [Google Scholar]
- Scarpa, A. Differenze Morfologiche fra i Palombi Mustelus Mustelus (Linnaeus, 1758) e Mustelus Punctulatus Risso, 1827 (Elasmobranchii, Triakidae). Bachelor’s Thesis, University of Padova, Padova, Italy, 2017. [Google Scholar]
- Deynat, P. Les Requins. Identification des Nageoires. Guide Pratique; Editions Quae: Versailles, France, 2010. [Google Scholar]
- Irschick, D.J.; Fu, A.; Lauder, G.V.; Wilga, C.D.; Kuo, C.Y.; Hammerschlag, N. A comparative morphological analysis of body and fin shape for eight shark species. Biol. J. Linn. Soc. 2017, 122, 589–604. [Google Scholar] [CrossRef]
- Lipej, L.; De Maddalena, A.; Soldo, A. Sharks of the Adriatic Sea; Knjiznica Annales Majora: Koper, Slovenia, 2004. [Google Scholar]
- Lauder, G.V.; Wainwright, D.K.; Domel, A.G.; Weaver, J.C.; Wen, L.; Bertoldi, K. Structure, biomimetics, and fluid dynamics of fish skin surfaces. Physic. Rev. Fluids 2016, 1, 060502. [Google Scholar] [CrossRef]
- Raschi, W.; Tabit, C. Functional aspects of placoid scales: A review and update. Aust. J. Mar. Freshw. Res. 1992, 43, 123–147. [Google Scholar] [CrossRef]
- Rodríguez-Mendoza, R.; Muñoz, M.; Saborido-Rey, F. Ontogenetic allometry of the bluemouth, Helicolenus dactylopterus dactylopterus (Teleostei: Scorpaenidae), in the Northeast Atlantic and Mediterranean based on geometric morphometrics. Hydrobiologia 2011, 670, 5–22. [Google Scholar] [CrossRef]
- Wetherbee, B.M.; Cortés, E.; Bizzarro, J.J. Food consumption and feeding habits. In Biology of Sharks and their Relatives; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 239–263. [Google Scholar]
- Baje, L.; Chin, A.; White, W.T.; Simpfendorfer, C.A. Dietary overlap of carcharhinid sharks in the Gulf of Papua. Mar. Freshw. Res. 2022, 73, 605–614. [Google Scholar] [CrossRef]
- Ross, S.T. Patterns of resource partitioning in searobins (Pisces: Triglidae). Copeia 1977, 1977, 561–571. [Google Scholar] [CrossRef]
- Espinoza, M.; Munroe, S.E.M.; Clarke, T.M.; Fisk, A.T.; Wehrtmann, I.S. Feeding ecology of common demersal elasmobranch species in the Pacific coast of Costa Rica inferred from stable isotope and stomach content analyses. J. Exp. Mar. Biol. Ecol. 2015, 470, 12–25. [Google Scholar] [CrossRef]
- Post, D.M. Using Stable Isotopes to Estimate Trophic Position: Models, Methods, and Assumptions. Ecology 2002, 83, 703–771. [Google Scholar] [CrossRef]
- Demirhan, S.A.; Seyhan, K.; Başusta, N. Dietary overlap in spiny dogfish (Squalus acanthias) and thornback ray (Raja clavata) in the southeastern Black Sea. Ekoloji 2007, 16, 1–8. [Google Scholar]
- Šantić, M.; Rada, B.; Pallaoro, A. Diet and feeding strategy of thornback ray Raja clavata. J. Fish Biol. 2012, 81, 1070–1084. [Google Scholar] [CrossRef] [PubMed]
- Lipej, L.; Mavrič, B.; Paliska, D.; Capapé, C. Feeding habits of the pelagic stingray Pteroplatytrygon violacea (Chondrichthyes: Dasyatidae) in the Adriatic Sea. J. Mar. Biol. Assoc. UK 2013, 93, 285–290. [Google Scholar] [CrossRef]
- Kadri, H.; Marouani, S.; Bradai, M.N.; Bouaïn, A. Diet and feeding strategy of thornback ray, Raja clavata (Chondrichthyes; Rajidae) from the Gulf of Gabes (Tunisia–Central Mediterranean Sea). J. Mar. Biol. Assoc. UK 2014, 94, 1509–1516. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stomach content analysis | Scyliorhinus canicula | |||||
| Small | Big | Total | ||||
| Cold season | Hot season | Cold season | Warm season | |||
| Males | 41.3 ± 0.4 (2) | 35.3 ± 2.5 (3) | 45.8 ± 2.7 (6) | 45.5 ± 2.2 (31) | 44.6 ± 3.5 (42) | |
| Females | 40.3 ± 1.4 (11) | 40.1 ± 1.9 (18) | 44.6 ± 1.6 (70) | 44.4 ± 1.8 (59) | 43.7 ± 2.4 (158) | |
| Total | 39.8 ± 2.2 (34) | 44.8 ± 1.9 (166) | ||||
| Mustelus mustelus | ||||||
| Small | Big | Total | ||||
| Males | 62.0 ± 12.8 (21) | 106.4 ± 10.0 (6) | 71.9 ± 22.4 (27) | |||
| Females | 63.4 ± 13.3 (28) | 118.7 ± 16.3 (47) | 98.1 ± 30.9 (75) | |||
| Total | 62.8 ± 13.0 (49) | 117.3 ± 16.2 (53) | ||||
| Mustelus punctulatus | ||||||
| Small | Big | Total | ||||
| Males | 39.0 ± 3.0 (49) | 106.9 ± 11.0 (12) | 52.4 ± 27.7 (61) | |||
| Females | 38.9 ± 5.0 (43) | 108.9 ± 11.1 (9) | 51.0 ± 27.5 (52) | |||
| Total | 39.0 ± 4.0 (92) | 107.8 ± 10.8 (21) | ||||
| Stable isotope analysis | Scyliorhinus canicula | |||||
| Small | Big | Total | ||||
| Males | / | 45.2 ± 1.8 (12) | ||||
| Females | / | 45.3 ± 1.8 (12) | ||||
| Total | / | 45.2 ± 1.8 (24) | ||||
| Mustelus mustelus | ||||||
| Small | Big | Total | ||||
| Males | 73.0 ± 9.4 (4) | 114.2 ± 14.4 (6) | 97.7 ± 24.4 (10) | |||
| Females | 69.8 ± 11.5 (6) | 125.5 (1) | 77.8 ± 23.5 (7) | |||
| Total | 71.1 ± 10.3 (10) | 115.8 ± 13.8 (7) | ||||
| Mustelus punctulatus | ||||||
| Small | Big | Total | ||||
| Males | 53.7 ± 6.4 (5) | 101.0 ± 9.6 (9) | 84.1 ± 25.0 (14) | |||
| Females | 50.5 (1) | 105.1 ± 15.5 (9) | 99.6 ± 22.6 (10) | |||
| Total | 53.2 ± 5.9 () | 103.0 ± 12.7 (18) | ||||
| Fullness% | Shannon–Wiener | Pielou | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sc | Mm | Mp | Sc | Mm | Mp | Sc | Mm | Mp | ||
| Size | F2,197 = 4.45 p < 0.001 | F3,98 = 0.44 p = 0.66 | F3,109 = 4.02 p < 0.001 | F3,196 = 0.96, p = 0.34 | F3,98 = 0.51 p = 0.61 | F3,109 = 0.88 p = 0.38 | F3,178 = 0.23 p = 0.82 | F3,90 = 0.49 p = 0.62 | F3,101 = 0.27 p = 0.79 | |
| Big | 1.90 ± 1.88% | 1.29 ± 0.72% | 0.68 ± 0.49% | 0.70 ± 0.46 | 0.73 ± 0.41 | 0.85 ± 0.52 | 0.62 ± 0.26 | 0.64 ± 0.22 | 0.66 ± 0.24 | |
| Small | 4.32 ± 3.47% | 2.08 ± 1.41% | 1.48 ± 0.92% | 0.82 ± 0.42 | 0.70 ± 0.46 | 0.89 ± 0.47 | 0.63 ± 0.22 | 0.58 ± 0.26 | 0.75 ± 0.22 | |
| Sex | F2,197 = 2.45 p = 0.02 | F3,98 =0.05 p = 0.96 | F3,109 =1.64 p = 0.10 | F3,196 = 0.62, p = 0.53 | F3,98 = 0.71 p = 0.48 | F3,109 = 0.91 p = 0.37 | F3,178 = 1.28 p = 0.20 | F3,90 = 0.33 p = 0.74 | F3,101 = 0.07 p = 0.95 | |
| Males | 1.41 ± 1.47% | 1.65 ± 1.16% | 1.32 ± 0.88% | 0.76 ± 0.45 | 0.70 ± 0.49 | 0.88 ± 0.46 | 0.72 ± 0.23 | 0.59 ± 0.28 | 0.74 ± 0.19 | |
| Females | 2.55 ± 2.54% | 1.68 ± 1.18% | 1.35 ± 0.95% | 0.71 ± 0.46 | 0.72 ± 0.41 | 0.89 ± 0.49 | 0.59 ± 0.26 | 0.61 ± 0.23 | 0.72 ± 0.26 | |
| Size × sex | F3,196 = 1.28 p = 0.20 | F3,98 = 0.16 p = 0.87 | F3,109 = 1.81 p = 0.07 | F3,196 = 0.64 p = 0.52 | F3,98 = 0.82 p = 0.41 | F3,109 = 1.03 p = 0.31 | F3,178 = 0.05 p = 0.96 | F3,90 = 0.41 p = 0.68 | F3,101 = 0.01 p = 0.99 | |
| Season | F1,198 = 2.14 p = 0.03 | / | / | F1,198 = 2.15 p = 0.03 | / | / | F1,180= 2.22 p = 0.06 | / | / | |
| Warm | 2.33 ± 2.22% | 0.84 ± 0.42 | 0.69 ± 0.21 | |||||||
| Cold | 2.28 ± 2.62% | 0.58 ± 0.47 | 0.53 ± 0.28 | |||||||
| Source | df | SS | MS | Pseudo-F | P (perm) | Unique Perms | |
|---|---|---|---|---|---|---|---|
| S. canicula | Size | 1 | 3631.9 | 3631.9 | 1.008 | 0.420 | 999 |
| Sex | 1 | 10131 | 10131 | 2.811 | 0.003 | 997 | |
| Season | 1 | 7303.9 | 7303.9 | 2.027 | 0.034 | 999 | |
| Size × Sex | 1 | 3867.8 | 3867.8 | 1.073 | 0.370 | 999 | |
| Size × Season | 1 | 4132.5 | 4132.5 | 1.147 | 0.319 | 999 | |
| Sex × Season | 1 | 3229.4 | 3229.4 | 0.896 | 0.571 | 999 | |
| Size × Sex × Season | 1 | 3468.8 | 3468.8 | 0.965 | 0.474 | 998 | |
| Residual | 192 | 6.9 × 105 | 3603.9 | ||||
| Total | 199 | 7.4 × 105 | |||||
| M. mustelus | Size | 1 | 20353 | 20353 | 8.156 | 0.001 | 999 |
| Sex | 1 | 1390.3 | 1390.3 | 0.557 | 0.737 | 999 | |
| Size × Sex | 1 | 6193.4 | 6193.4 | 2.482 | 0.039 | 999 | |
| Residual | 98 | 2.4 × 105 | 2495.5 | ||||
| Total | 101 | 2.9 × 105 | |||||
| M. punctulatus | Size | 1 | 41462 | 41462 | 13.123 | 0.001 | 997 |
| Sex | 1 | 3348.9 | 3348.9 | 1.060 | 0.391 | 999 | |
| Size × Sex | 1 | 4254.8 | 4254.8 | 1.347 | 0.204 | 996 | |
| Residual | 109 | 3.4 × 105 | 3159.5 | ||||
| Total | 112 | 3.9 × 105 |
| Species | Factor | Prey Category | Average Biomass | Average Biomass | Contribution (%) |
|---|---|---|---|---|---|
| S. canicula | Sex | Females | Males | ||
| Portunidae | 1.15 | 0.33 | 21.77 | ||
| Unid. Teleosts | 0.44 | 0.24 | 9.24 | ||
| Other Caridea | 0.14 | 0.33 | 7.45 | ||
| Unid. Crustaceans | 0.19 | 0.22 | 6.62 | ||
| Sepiolidae | 0.17 | 0.22 | 5.91 | ||
| Other Brachiura | 0.18 | 0.19 | 5.82 | ||
| Rissoides desmaresti | 0.11 | 0.14 | 4.63 | ||
| Cepola macrophthalma | 0.15 | 0.12 | 4.36 | ||
| Alpheus glaber | 0.08 | 0.17 | 4.19 | ||
| Flatfishes | 0.29 | 0.06 | 4.10 | ||
| Season | Cold season | Warm season | |||
| Portunidae | 0.95 | 0.99 | 24.02 | ||
| Unid. Teleosts | 0.46 | 0.35 | 10.70 | ||
| Flatfishes | 0.36 | 0.14 | 6.54 | ||
| Unid. Crustaceans | 0.11 | 0.26 | 6.35 | ||
| Other Brachiura | 0.16 | 0.20 | 5.46 | ||
| Sepiolidae | 0.11 | 0.23 | 4.97 | ||
| Deltentosteus quadrimaculatus | 0.22 | 0.11 | 4.62 | ||
| Other Caridea | 0.14 | 0.22 | 4.61 | ||
| Cepola macrophthalma | 0.09 | 0.19 | 4.31 | ||
| Unid. Polychaete | 0.12 | 0.13 | 4.10 | ||
| M. mustelus | Size | Small animals | Big animals | ||
| Portunidae | 1.39 | 5.83 | 39.39 | ||
| Squilla mantis | 1.40 | 2.59 | 21.43 | ||
| Other Brachiura | 0.40 | 0.69 | 7.48 | ||
| Ethusa mascarone | 0.85 | 0.00 | 6.68 | ||
| Unid. Crustaceans | 0.39 | 0.25 | 4.24 | ||
| Other Stomatopoda | 0.22 | 0.37 | 4.20 | ||
| M. punctulatus | Size | Portunidae | 0.04 | 2.20 | 25.17 |
| Squilla mantis | 0.02 | 0.85 | 9.13 | ||
| Unid. Teleosts | 0.15 | 0.69 | 7.30 | ||
| Other Polychaete | 0.40 | 0.49 | 6.41 | ||
| Pelagic fishes | 0.13 | 0.48 | 5.45 | ||
| Other Brachiura | 0.20 | 0.46 | 5.35 | ||
| Other | 0.02 | 0.48 | 5.00 | ||
| Unid. Cephalopods | 0.14 | 0.41 | 4.56 | ||
| Anomura | 0.35 | 0.07 | 4.36 | ||
| Unid. Processidae | 0.36 | 0.00 | 4.19 |
| Comparison | t | P (perm) | Unique Perms |
|---|---|---|---|
| SF, SM | 1.2579 | 0.140 | 998 |
| SF, BF | 3.3407 | 0.001 | 999 |
| SF, BM | 1.3271 | 0.120 | 998 |
| SM, BF | 3.4784 | 0.001 | 999 |
| SM, BM | 1.8145 | 0.008 | 995 |
| BF, BM | 1.2952 | 0.173 | 998 |
| PERMANOVA | ||||||
|---|---|---|---|---|---|---|
| Source | df | SS | MS | Pseudo-F | P (perm) | Unique Perms |
| Species | 2 | 1.8 × 105 | 91733 | 26.52 | 0.001 | 998 |
| Residual | 412 | 1.4 × 105 | 3459.1 | |||
| Total | 414 | 1.6 × 105 | ||||
| Pairwise comparison | ||||||
| Pair-wise comparison | t | P (perm) | Unique perms | |||
| Sc, Mm | 5.211 | 0.001 | 999 | |||
| Sc, Mp | 4.492 | 0.001 | 999 | |||
| Mm, Mp | 6.002 | 0.001 | 999 | |||
| Prey Category | Average Biomass | Average Biomass | Contribution (%) |
|---|---|---|---|
| S. canicula | M. mustelus | ||
| Portunidae | 0.97 | 3.70 | 31.01 |
| Squilla mantis | 0.01 | 2.02 | 18.80 |
| Other Brachiura | 0.18 | 0.55 | 6.75 |
| Unid. Teleosts | 0.40 | 0.27 | 5.24 |
| Ethusa mascarone | 0.00 | 0.41 | 5.17 |
| Unid. Crustaceans | 0.19 | 0.32 | 4.57 |
| S. canicula | M. punctulatus | ||
| Portunidae | 0.97 | 0.44 | 17.72 |
| Unid. Teleosts | 0.40 | 0.25 | 8.32 |
| Other Polychaete | 0.13 | 0.42 | 7.30 |
| Unid. Crustaceans | 0.19 | 0.25 | 6.60 |
| Processidae | 0.10 | 0.29 | 6.57 |
| Anomura | 0.01 | 0.30 | 6.00 |
| Other Brachiura | 0.18 | 0.25 | 5.75 |
| Other Caridea | 0.18 | 0.18 | 5.52 |
| Sepiolidae | 0.18 | 0.13 | 4.50 |
| M. mustelus | M. punctulatus | ||
| Portunidae | 3.70 | 0.44 | 30.86 |
| Squilla mantis | 2.02 | 0.17 | 18.62 |
| Other Brachiura | 0.55 | 0.25 | 6.77 |
| Ethusa mascarone | 0.41 | 0.11 | 5.52 |
| Unid. Crustaceans | 0.32 | 0.25 | 4.78 |
| Other Polychaete | 0.04 | 0.42 | 4.18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Finotto, L.; Berto, D.; Rampazzo, F.; Raicevich, S.; Bonanomi, S.; Mazzoldi, C. Trophic Partitioning among Three Mesopredatory Shark Species Inhabiting the Northwestern Adriatic Sea. Diversity 2023, 15, 1163. https://doi.org/10.3390/d15121163
Finotto L, Berto D, Rampazzo F, Raicevich S, Bonanomi S, Mazzoldi C. Trophic Partitioning among Three Mesopredatory Shark Species Inhabiting the Northwestern Adriatic Sea. Diversity. 2023; 15(12):1163. https://doi.org/10.3390/d15121163
Chicago/Turabian StyleFinotto, Licia, Daniela Berto, Federico Rampazzo, Saša Raicevich, Sara Bonanomi, and Carlotta Mazzoldi. 2023. "Trophic Partitioning among Three Mesopredatory Shark Species Inhabiting the Northwestern Adriatic Sea" Diversity 15, no. 12: 1163. https://doi.org/10.3390/d15121163