
Environmental Correlates to Genetic Diversity and Structure in Invasive Apple Snail (Pomacea canaliculata) Populations in China
Abstract
:1. Introduction
2. Material and Methods
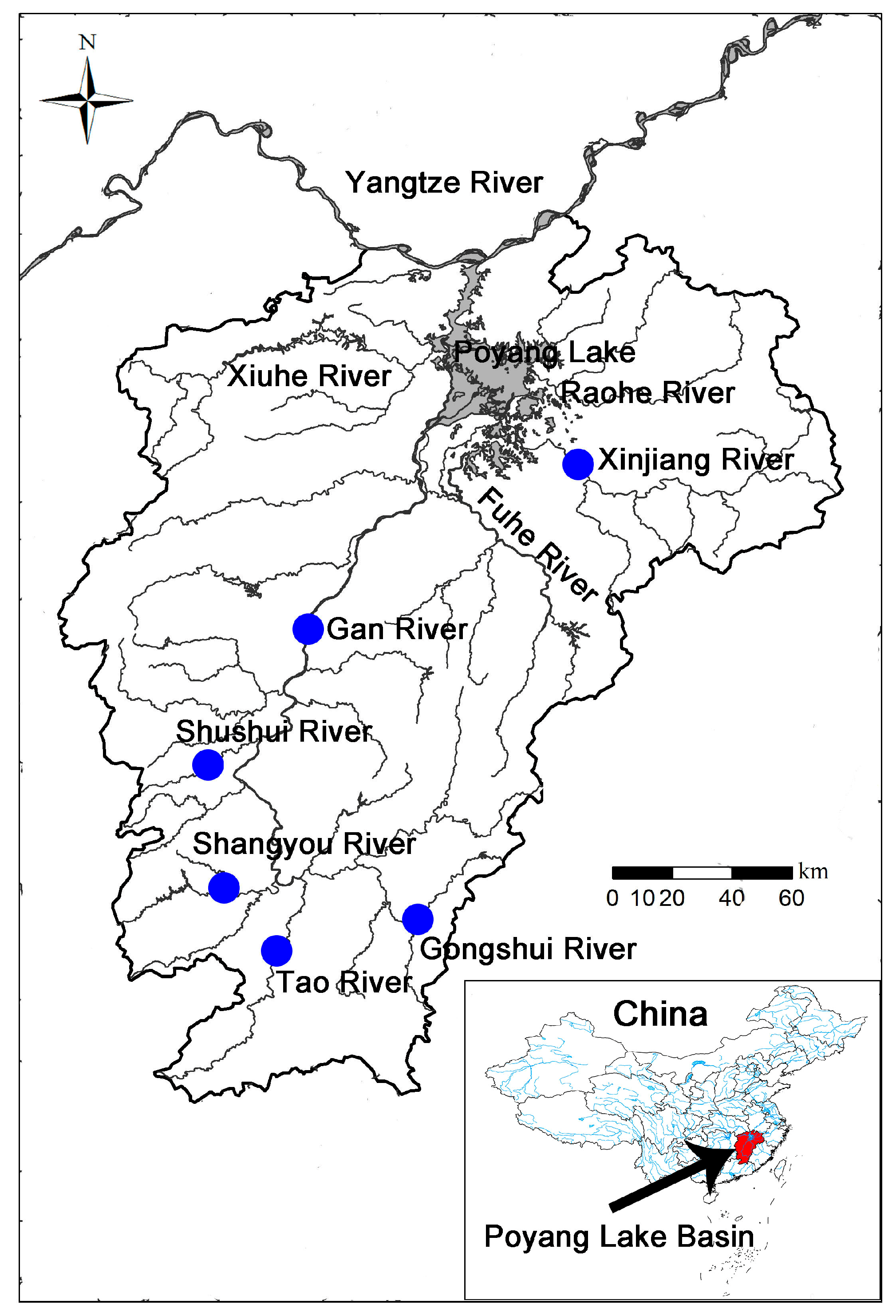
2.1. Sample Collection and DNA Extraction
2.2. PCR Amplification
2.3. Data Analysis
2.4. Correlation between Genetic Diversity and Physicochemical Measurements
3. Results
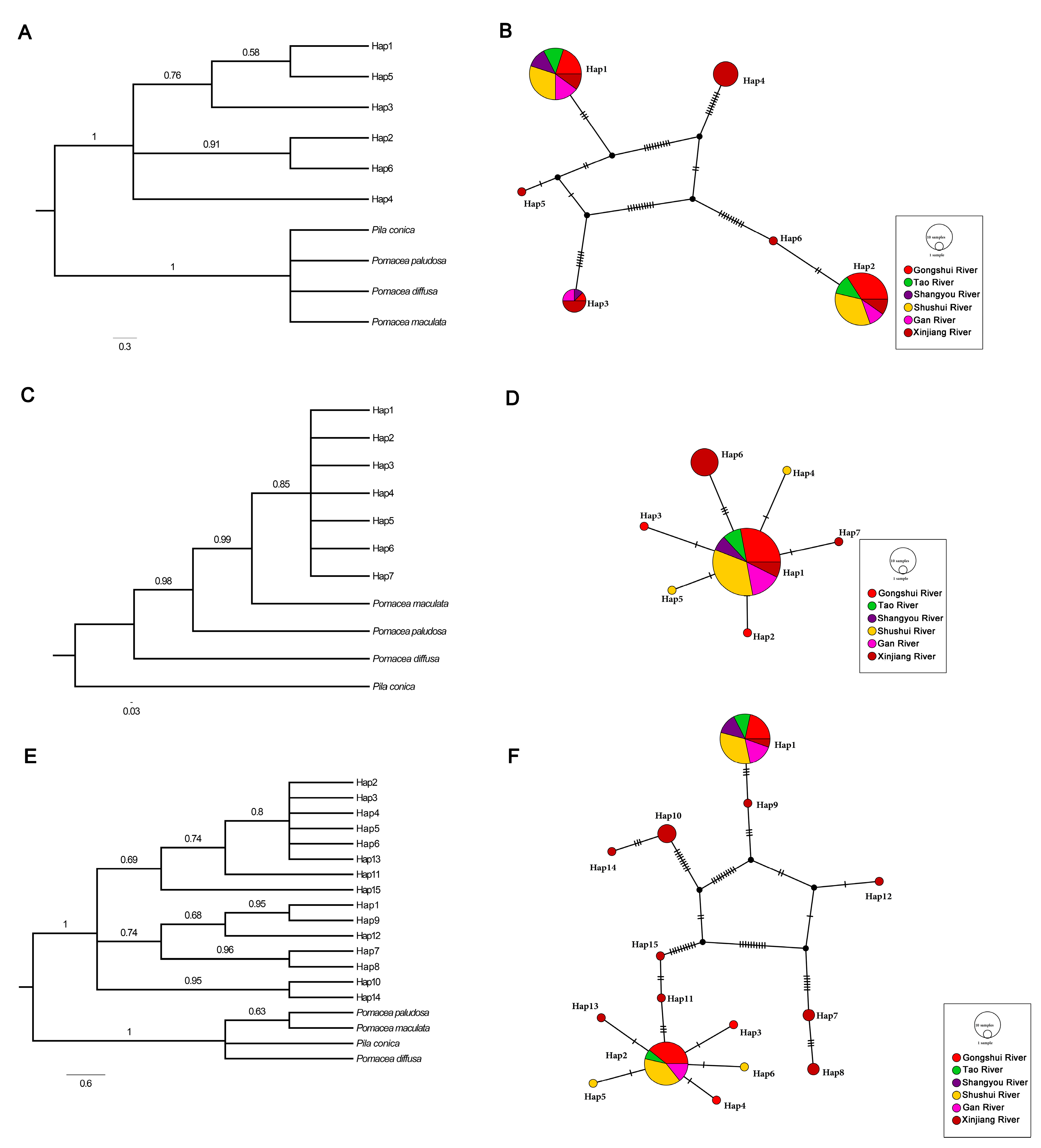
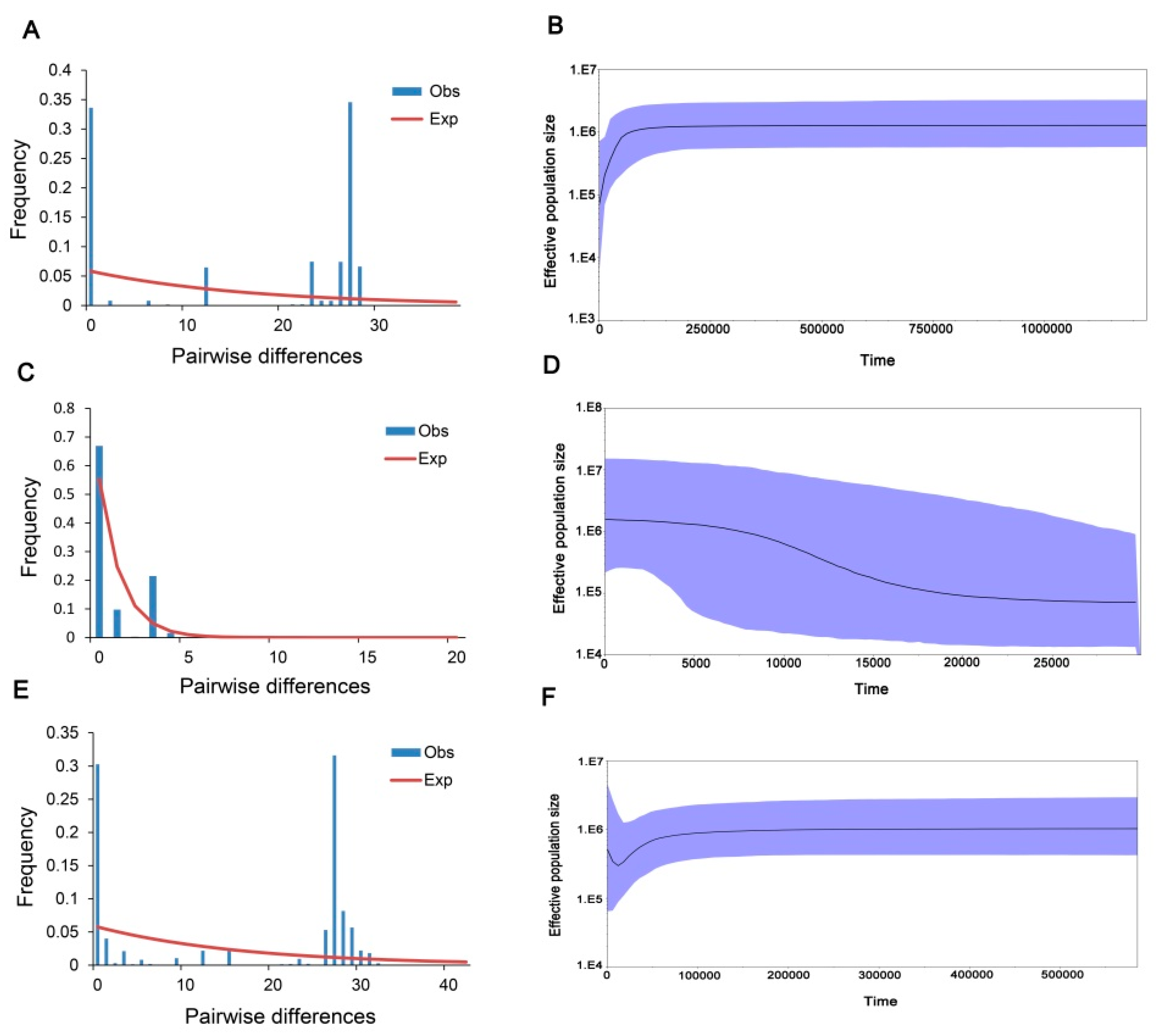
3.1. Genetic Diversity of P. canaliculata

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Collection Locations | COI Datasets | 16S Datasets | COI+16S Datasets | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genetic Diversity | Neutrality Tests | Genetic Diversity | Neutrality Tests | Genetic Diversity | Neutrality Tests | |||||||||||||
| N | H | Hd | π | Tajima’s D | Fu’s Fs | N | H | Hd | π | Tajima’s D | Fu’s Fs | N | H | Hd | π | Tajima’s D | Fu’s Fs | |
| Gongshui River | 23 | 3 | 0.530 | 0.023 | 2.11 | 17.99 | 21 | 3 | 0.186 | 0.001 | −1.51 | −1.91 | 21 | 4 | 0.605 | 0.013 | 2.64 | 13.67 |
| Tao River | 10 | 2 | 0.556 | 0.025 | 2.73 | 14.08 | 6 | 1 | 0 | 0 | 0 | 0 | 6 | 2 | 0.533 | 0.014 | 1.37 | 9.47 |
| Shangyou River | 6 | 2 | 0.333 | 0.007 | −1.45 | 4.82 | 5 | 1 | 0 | 0 | 0 | 0 | 5 | 1 | 0 | 0 | 0 | 0 |
| Shushui River | 25 | 2 | 0.520 | 0.024 | 3.58 | 23.39 | 25 | 3 | 0.157 | 0.001 | −1.51 | −2.12 | 25 | 4 | 0.597 | 0.014 | 3.16 | 15.99 |
| Gan River | 12 | 3 | 0.667 | 0.026 | 1.85 | 12.21 | 10 | 1 | 0 | 0 | 0 | 0 | 10 | 2 | 0.533 | 0.014 | 2.43 | 13.80 |
| Xinjiang River | 23 | 6 | 0.787 | 0.031 | 2.02 | 12.53 | 17 | 3 | 0.522 | 0.004 | 1.01 | 2.20 | 17 | 10 | 0.904 | 0.019 | 1.65 | 3.06 |
| Total | 100 | 6 | 0.664 | 0.028 | 84 | 7 | 0.331 | 0.002 | 84 | 15 | 0.697 | 0.016 | ||||||
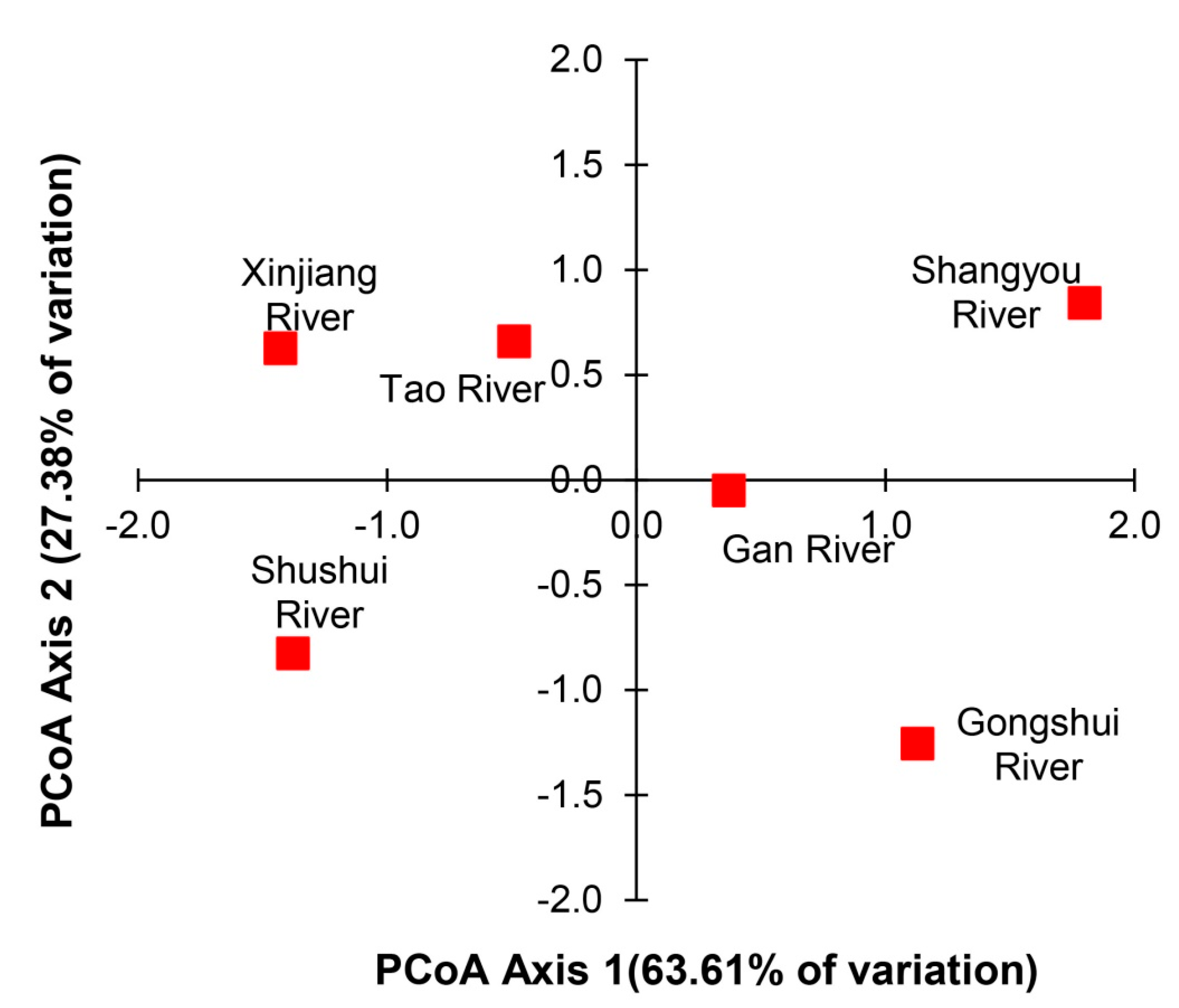
3.2. Genetic Structure of P. canaliculata
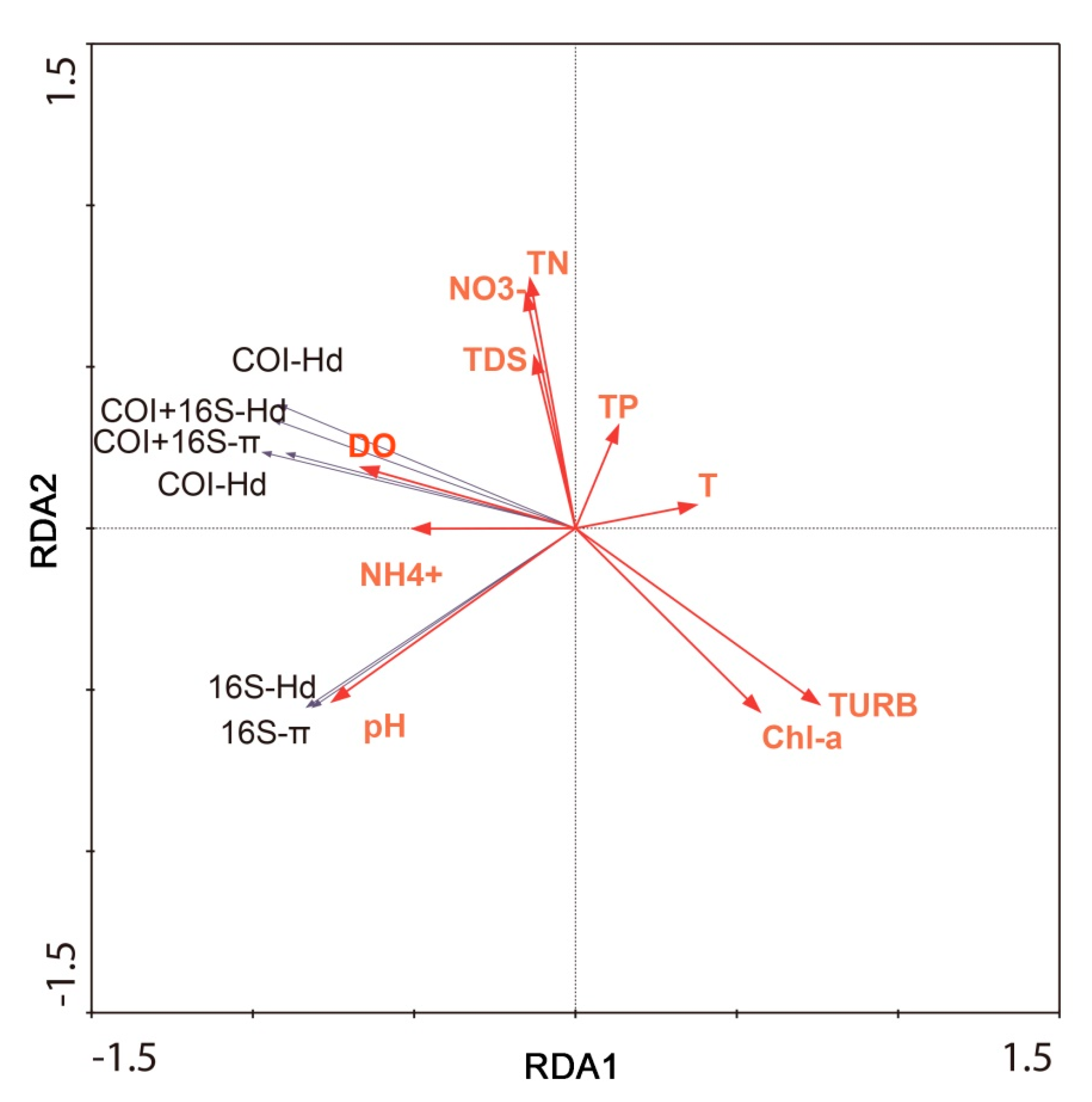
3.3. Correlation between Genetic and Physicochemical Parameters
4. Discussion
4.1. Genetic Diversity of P. canaliculata
4.2. Genetic Structure of P. canaliculata
4.3. Correlation between Genetic and Water Quality Parameters
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Havel, J.E.; Lee, C.E.; Vander Zanden, M.J. Do reservoirs facilitate invasions into landscapes? Bioscience 2005, 55, 518. [Google Scholar]
- Turak, E.; Harrison, I.; Dudgeon, D.; Abell, R.; Bush, A.; Darwall, W.; Finlayson, C.M.; Ferrier, S.; Freyhof, J.; Hermoso, V.; et al. Essential biodiversity variables for measuring change in global freshwater biodiversity. Biol. Conserv. 2016, 213, 272–279. [Google Scholar] [CrossRef]
- Sala, O.E.; Chapin, I.F.S.; Armesto, J.J.; Berlow, E.L.; Bloomfield, J.B.; Dirzo, R.H.; Huber-Sanwald, E.; Huenneke, L.; Jackson, R.B.; Kinzig, A.P. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, B.; Clavero, M.; Sanchez, M.I.; Vila, M. Global ecological impacts of invasive species in aquatic ecosystems. Global Chang. Biol. 2016, 22, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.; Knowler, D.; Leveque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status, and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Thomaz, S.M.; Kovalenko, K.E.; Havel, J.E.; Kats, L.B. Aquatic invasive species: General trends in the literature and introduction to the special issue. Hydrobiologia 2015, 746, 1–12. [Google Scholar] [CrossRef]
- Hayes, K.A.; Burks, R.L.; Castro-Vazquez, A.; Darby, P.C.; Heras, H.; Martín, P.R.; Qiu, J.W.; Thiengo, S.C.; Vega, I.A.; Wada, T.; et al. Insights from an integrated view of the biology of apple snails (Caenogastropoda: Ampullariidae). Malacologia 2015, 58, 245–302. [Google Scholar] [CrossRef]
- Nakano, D.; Baba, T.; Endo, N.; Nagayama, S.; Fujinaga, A.; Uchida, A.; Shiragane, A.; Urabe, M.; Kobayashi, T. Invasion, dispersion, population persistence and ecological impacts of a freshwater mussel (Limnoperna fortunei) in the Honshu Island of Japan. Biol. Invasions 2015, 17, 743–759. [Google Scholar] [CrossRef]
- Byers, J.E.; Mcdowell, W.G.; Dodd, S.R.; Haynie, R.S.; Pintor, L.M.; Wilde, S.B. Climate and pH predict the potential range of the invasive apple snail (Pomacea insularum) in the Southeastern United States. PLoS ONE 2013, 8, e56812. [Google Scholar] [CrossRef]
- Karraker, N.E.; Dudgeon, D. Invasive apple snails (Pomacea canaliculata) are predators of amphibians in South China. Biol. Invasions 2014, 16, 1785–1789. [Google Scholar] [CrossRef]
- Kwong, K.L.; Dudgeon, D.; Wong, P.K.; Qiu, J.W. Secondary production and diet of an invasive snail in freshwater wetlands: Implications for resource utilization and competition. Biol. Invasions 2010, 12, 1153–1164. [Google Scholar] [CrossRef]
- Cowie, R.H. Apple snails (Ampullariidae) as agricultural pests: Their biology, impacts and management. In Molluscs as Crop Pests; Barker, G.M., Ed.; CAB International: Wallingford, CT, USA, 2002; pp. 145–192. [Google Scholar]
- Ramakrishnan, V. Salinity, pH, Temperature, Desiccation and Hypoxia Tolerance in the Invasive Freshwater Snail Pomacea insularum. Ph.D. Thesis, University of Texas at Arlington, Arlington, TX, USA, 2007. [Google Scholar]
- Carlsson, N.O.L.; Brönmark, C.; Hansson, L.A. Invading herbivory: The golden apple snail alters ecosystem functioning in Asian wetlands. Ecology 2004, 85, 1575–1580. [Google Scholar] [CrossRef]
- Lv, S.; Zhang, Y.; Liu, H.X.; Hu, L.; Liu, Q.; Wei, F.R.; Guo, Y.H.; Steinmann, P.; Hu, W.; Zhou, X.N. Phylogenetic evidence for multiple and secondary introductions of invasive snails: Pomacea species in the People’s Republic of China. Divers. Distrib. 2013, 19, 147–156. [Google Scholar] [CrossRef]
- Mochida, O. Spread of freshwater Pomacea snails (Pilidae, Mollusca) from Argentina to Asia. Micronesica 1991, 3, 51–62. [Google Scholar]
- Lv, S.; Zhang, Y.; Steinmann, P.; Zhou, X.N. Emerging angiostrongyliasis in mainland China. Emerg. Infect. Dis. 2008, 14, 161–164. [Google Scholar] [CrossRef]
- Yusa, Y.; Suguira, N.; Wada, T. Predatory potential of freshwater animals on an invasive agricultural pest, the apple snail Pomacea canaliculata (Gastropoda: Ampullariidae), in southern Japan. Biol. Invasions 2006, 8, 137–147. [Google Scholar] [CrossRef]
- Rawlings, T.A.; Hayes, K.A.; Cowie, R.H.; Collins, T.M. The identity, distribution, and impacts of non-native apple snails in the continental United States. BMC Evol. Biol. 2007, 7, 97. [Google Scholar] [CrossRef]
- Pierre, S.M.; Quintana-Ascencio, P.F.; Boughton, E.H.; Jenkins, D.G. Dispersal and local environment affect the spread of an invasive apple snail (Pomacea maculata) in Florida, USA. Biol. Invasions 2017, 19, 2647–2661. [Google Scholar] [CrossRef]
- Joshi, R.C.; Parera, X.V. The rice apple snail in Spain: A review. J. Grey Syst. 2017, 59, 106–108. [Google Scholar]
- Hayes, K.A.; Joshi, R.C.; Thiengo, S.C.; Cowie, R.H. Out of South America: Multiple origins of non-native apple snails in Asia. Divers. Distrib. 2008, 14, 701–712. [Google Scholar] [CrossRef]
- Zhou, Y.; Yuan, X.Y.; Yang, Z.Y.; Liu, X.J.; Ouyang, S.; Wu, X.P. Spread pattern and potential distribution of the invasive species Pomacea spp. in China. J. Lake Sci. 2018, 30, 1379–1387. [Google Scholar]
- Lowe, S.; Browne, M.; Boudjelas, S.; DePoorter, M. 100 of the World’s Worst Invasive Alien Species, a Selection from the Global Invasive Species Database; First Published as Special Lift-Out in Aliens 12 December 2000; The Invasive Species Specialist Group (ISSG) a Specialist Group of the Species Survival Commission (SSC) of the World Conservation Union (IUCN): Rome, Italy, 2000; 12p. [Google Scholar]
- White, M.; Chejlava, M.; Fried, B.; Sherma, J. The concentration of calcium carbonate in shells of freshwater snails. Am. Malacol. Bull. 2007, 22, 139–142. [Google Scholar] [CrossRef]
- Perlman, H. Water Hardness; USGS [United States Geological Survey] Water Science School: Reston, VA, USA, 2016. [Google Scholar]
- Seuffert, M.E.; Martín, P.R. Dependence on aerial respiration and its influence on microdistribution in the invasive freshwater snail Pomacea canaliculata (Caenogastropoda, Ampullariidae). Biol. Invasions 2009, 12, 1695–1708. [Google Scholar] [CrossRef]
- Verbrugge, L.N.H.; Schipper, A.M.; Huijbregts, M.A.J.; Vander, V.G.; Leuven, R.S.E.W. Sensitivity of native and non-native mollusc species to changing river water temperature and salinity. Biol. Invasions 2012, 14, 1187–1199. [Google Scholar] [CrossRef]
- Martin, C.W.; Valentine, J.F. Tolerance of embryos and hatchlings of the invasive apple snail Pomacea maculata to estuarine conditions. Aquat. Ecol. 2014, 48, 321–326. [Google Scholar] [CrossRef]
- Andree, K.B.; López, M.A. Species identification from archived snail shells via genetic analysis: A method for DNA extraction from empty shells. Molluscan Res. 2013, 3, 1–5. [Google Scholar] [CrossRef]
- Bernatis, J.L.; Mcgaw, I.; Cross, C.L. Abiotic tolerances in different life stages of apple snails Pomacea canaliculata and Pomacea maculata and the implications for distribution. J. Shellfish Res. 2016, 35, 1013–1025. [Google Scholar] [CrossRef]
- Lv, S.; Zhang, Y.; Liu, H.X.; Hu, L.; Liu, W.; Liu, Q.; Li, S.Z.; Hu, W.; Utzinger, J.; Zhou, X.N. Population genetics of Pomacea in Mainland of China. Chin. J. Schistosomiasis Control. 2011, 23, 178–182. [Google Scholar]
- Lv, S.; Zhang, Y.; Liu, H.X.; Hu, L.; Yang, K.; Steinmann, P.; Chen, Z.; Wang, L.Y.; Utzinger, J.; Zhou, X.N. Invasive snails and an emerging infectious disease: Results from the first national survey on Angiostrongylus cantonensis in China. PLoS Neglect. Trop. Dis. 2009, 3, e368. [Google Scholar] [CrossRef]
- Hayes, K.A.; Cowie, R.H.; Thiengo, S.C.; Strong, E.E. Comparing apples with apples: Clarifying the identities of two highly invasive Neotropical Ampullariidae (Caenogastropoda). Zool. J. Linn. Soc. 2012, 166, 723–753. [Google Scholar] [CrossRef]
- Dlugosch, K.M.; Parker, I.M. Invading populations of an ornamental shrub show rapid life history evolution despite genetic bottlenecks. Ecol. Lett. 2008, 11, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Matsukura, K.; Okuda, M.; Azaniga, N.J.; Wada, T. Genetic exchange between two freshwater apple snails, Pomacea canaliculata and Pomacea maculata invading East and Southeast Asia. Biol. Invasions 2013, 15, 2039–2048. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit i from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Wei, F.R.; Lv, S.; Zhang, Y. Comparison on the methods of DNA extraction from lungs of Pomacea canaliculata used in PCR assay. Chin. J. Zool. 2009, 25, 1181–1185. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Mark, P.V.; Ayres, D.L.; Darling, A.; Hohna, S.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451. [Google Scholar] [CrossRef]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef]
- Murias dos Santos, A.; Cabezas, M.P.; Tavares, A.I.; Xavier, R.; Branco, M. tcsBU: A tool to extend TCS network layout and visualization. Bioinformatics 2016, 32, 627–628. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Rambaut, A.; Shapiro, B.; Pybus, O.G. Bayesian coalescent inference of past population dynamics from molecular sequences. Mol. Biol. Evol. 2005, 22, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Brown, W.M.; George, M.J.; Wilson, A.C. Rapid evolution of animal mitochondrial DNA. Proc. Nati. Acad. Sci. USA 1979, 76, 1967–1971. [Google Scholar] [CrossRef]
- Rogers, A.R. Genetic evidence for a Pleistocene population explosion. Evolution 1995, 49, 608–615. [Google Scholar] [CrossRef]
- Rogers, A.R.; Harpending, H. Population growth makes waves in the distribution of pairwise genetic differences. Mol. Biol. Evol. 1992, 9, 552–569. [Google Scholar]
- Li, K.; Liu, X.J.; Zhou, Y.; Xu, Y.; Lv, Q.; Ouyang, S.; Wu, X.P. Temporal and spatial changes in macrozoobenthos diversity in Poyang Lake Basin. Ecol. Evol. 2019, 9, 6353–6365. [Google Scholar] [CrossRef]
- Xing, Y.; Wu, X.P.; Ouyang, S.; Zhang, J.Q.; Xu, J.; Yin, S.L.; Xie, Z.C. Assessment of macrobenthos biodiversity and potential human-induced stressors in the Ganjiang River system. Biodivers. Sci. 2019, 27, 648–657. [Google Scholar]
- Wei, F.S.; Kou, H.R.; Hong, S.J. Methods for the Examination of Water and Wastewater; China Environmental Science Press: Beijing, China, 1989. [Google Scholar]
- Huang, X.F.; Chen, W.; Cai, Q.H. Standard Methods for Observation and Analysis in Chinese Ecosystem Research Networke Survey, Observation and Analysis of Lake Ecology; Standards Press of China: Beijing, China, 1999. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Lep, S.J.; Smilauer, P. Ultivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- ter Braak, C.J.F.; Verdonschot, P.F.M. Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquat. Sci. 1995, 57, 255–289. [Google Scholar] [CrossRef]
- Borcard, D.; Gillet, F.; Legendre, L. Numerical Ecology with R; Springer: New York, NY, USA, 2011. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Freeland, J.L.; Petersen, S.D.; Kirk, H. Molecular Ecology, 2nd ed.; Wiley-Blackwell: Oxford, UK, 2011. [Google Scholar]
- Liu, X.; Cao, Y.; Xue, T.; Wu, R.; Zhou, Y.; Zhou, C.; Zanatta, D.T.; Ouyang, S.; Wu, X. Genetic structure and diversity of Nodularia douglasiae (Bivalvia: Unionida) from the middle and lower Yangtze River drainage. PLoS ONE 2017, 12, e0189737. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.J.; Zhou, Y.; Ouyang, S.; Wu, X.P. Phylogeographic patterns and demographic history of Pomacea canaliculata and Pomacea maculata from different countries (Ampullariidae, Gastropoda, Mollusca). Nat. Conserv. 2019, 36, 71–92. [Google Scholar] [CrossRef]
- Shirk, R.Y.; Hamrick, J.L.; Zhang, C.; Qiang, S. Patterns of genetic diversity reveal multiple introductions and recurrent founder effects during range expansion in invasive populations of Geranium carolinianum (Geraniaceae). Heredity 2014, 112, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.Q.; Liu, S.W.; He, C.; Yu, X.P. Distribution and the origin of invasive apple snails, Pomacea canaliculata and P. maculata (Gastropoda: Ampullariidae) in China. Sci. Rep. 2018, 8, 1185. [Google Scholar] [CrossRef] [PubMed]
- Glass, N.H.; Darby, P.C. The effect of calcium and pH on Florida apple snail, Pomacea paludosa (Gastropoda:Ampullariidae), shell growth and crush weight. Aquat. Ecol. 2008, 43, 1085–1093. [Google Scholar] [CrossRef]
- Batzer, D.P.; Dietz-Brantley, S.E.; Taylor, B.E.; DeBiase, A.E. Evaluating regional differences in macroinvertebrate communities from forested depressional wetlands across eastern and central North America. J. N. Am. Benthol. Soc. 2005, 24, 403–414. [Google Scholar] [CrossRef]
- Bernatis, J.L. Morphology, Ecophysiology, and Impacts of Nonindigenous Pomacea in Florida. Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 2014. [Google Scholar]
- Deaton, L.E.; Schmidt, W.; Leblanc, B.; Carter, J.; Merino, S. Physiology of the invasive apple snail Pomacea maculata: Tolerance to low temperatures. J. Shellfish. Res. 2016, 35, 207–210. [Google Scholar] [CrossRef]
- Habel, J.C.; Rodder, D.; Schmitt, T.N. Global warming will affect the genetic diversity and uniqueness of Lycaena helle populations. Glob. Chang. Biol. 2011, 17, 194–205. [Google Scholar] [CrossRef]
- Row, J.R.; Wilson, P.J.; Gomez, C.; Koen, E.L.; Bowman, J.; Thornton, D.; Murray, D.L. The subtle role of climate change on population genetic structure in Canada lynx. Glob. Chang. Biol. 2014, 20, 2076–2086. [Google Scholar] [CrossRef]
- Sexton, J.P.; Hangartner, S.B.; Hoffmann, A.A. Genetic isolation by environment or distance: Which pattern of gene flow is most common? Evolution 2014, 68, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Berg, D.J. Predicting the effects of climate change on population connectivity and genetic diversity of an imperiled freshwater mussel, Cumberlandia monodonta (Bivalvia: Margaritiferidae), in riverine systems. Glob. Chang. Biol. 2016, 23, 94–107. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Lang, B.K.; Berg, D.J. Past climate change drives current genetic structure of an endangered freshwater mussel species. Mol. Ecol. 2015, 24, 1910–1926. [Google Scholar] [CrossRef] [PubMed]
- Arora, J.; Mehra, N.K. Seasonal dynamics of zooplankton in a shallow eutrophic, man-made hyposaline lake in Delhi (India): Role of environmental factors. Hydrobiologia 2009, 626, 27–40. [Google Scholar] [CrossRef]
- Chen, L.Y.; Heath, A.G.; Neves, R.J. Comparison of oxygen consumption in freshwater mussels (Unionidae) from different habitats during declining dissolved oxygen concentration. Hydrobiologia 2001, 450, 209–214. [Google Scholar] [CrossRef]
- Liu, X.J.; Wu, R.W.; Chen, X.; Zhou, Y.; Yang, L.M.; Ouyang, S.; Wu, X.P. Effects of dams and their environmental impacts on the genetic diversity and connectivity of freshwater mussel populations in Poyang Lake Basin, China. Freshw. Biol. 2020, 65, 264–277. [Google Scholar] [CrossRef]
- Quinlan, E.; Malcolm, I.A.; Gibbins, C.N. Spatio-temporal variability of dissolved oxygen within the shallow subsurface zone of a freshwater pearl mussel bed. Fundam. Appl. Limnol. 2014, 185, 281–294. [Google Scholar] [CrossRef]





| COI Datasets | |||||
|---|---|---|---|---|---|
| Source of Variance | df | Sum of Squares | Variance Components | Percentage of Variation | F-Statistics |
| Among populations | 5 | 112.32 | 0.95 | 11.46 | FST = 0.11 * |
| Within populations | 94 | 688.49 | 7.32 | 88.54 | |
| Total | 99 | 800.82 | 8.27 | 100 | |
| 16S datasets | |||||
| Among populations | 5 | 17.21 | 0.24 | 53.79 | FST = 0.54 * |
| Within populations | 78 | 16.41 | 0.21 | 46.21 | |
| Total | 83 | 33.62 | 0.45 | 100 | |
| COI+16S datasets | |||||
| Among populations | 5 | 114.13 | 1.18 | 14.01 | FST = 0.14 * |
| Within populations | 78 | 565.47 | 7.25 | 85.99 | |
| Total | 83 | 679.60 | 8.43 | 100 | |
| COI Datasets | 16S Datasets | COI+16S Datasets | ||||
|---|---|---|---|---|---|---|
| Hd | π | Hd | π | Hd | π | |
| WT | −0.278 | −0.222 | −0.075 | −0.096 | −0.195 | −0.213 |
| TDS | −0.273 | −0.177 | −0.379 | −0.379 | −0.202 | −0.197 |
| pH | 0.384 | 0.089 | 0.881 * | 0.950 * | 0.186 | 0.137 |
| DO | 0.250 | 0.379 | −0.221 | −0.231 | 0.351 | 0.369 |
| TURB | 0.396 | 0.702 | −0.352 * | −0.354 * | 0.645 | 0.692 |
| Chl-a | −0.063 | 0.159 | −0.305 | −0.300 | 0.118 | 0.156 |
| NH4+ | −0.190 | −0.042 | −0.103 | −0.166 | −0.048 | −0.028 |
| NO3− | −0.048 | 0.110 | −0.308 | −0.329 | 0.087 | 0.087 |
| TP | −0.132 | −0.153 | 0.185 | 0.246 | −0.128 | −0.121 |
| TN | 0.016 | 0.239 | −0.287 | −0.275 | 0.237 | 0.245 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Zhou, Y.; Ouyang, S.; Wu, X. Environmental Correlates to Genetic Diversity and Structure in Invasive Apple Snail (Pomacea canaliculata) Populations in China. Diversity 2023, 15, 1048. https://doi.org/10.3390/d15101048
Liu X, Zhou Y, Ouyang S, Wu X. Environmental Correlates to Genetic Diversity and Structure in Invasive Apple Snail (Pomacea canaliculata) Populations in China. Diversity. 2023; 15(10):1048. https://doi.org/10.3390/d15101048
Chicago/Turabian StyleLiu, Xiongjun, Yu Zhou, Shan Ouyang, and Xiaoping Wu. 2023. "Environmental Correlates to Genetic Diversity and Structure in Invasive Apple Snail (Pomacea canaliculata) Populations in China" Diversity 15, no. 10: 1048. https://doi.org/10.3390/d15101048