
Phytoplankton Community Dynamics in Ponds with Diverse Biomanipulation Approaches


Abstract
:1. Introduction
2. Materials and Methods
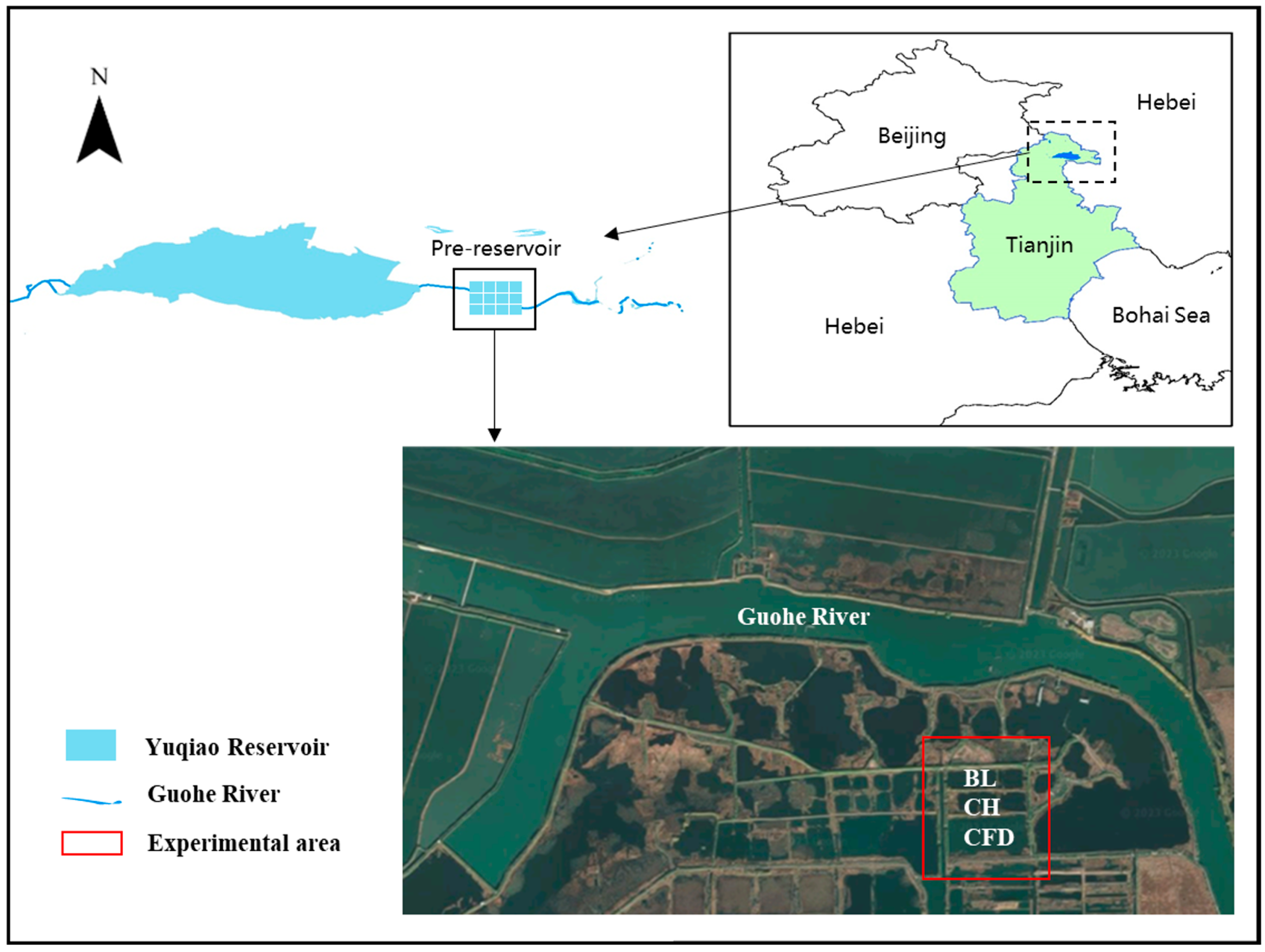
2.1. Study Area
2.2. Sampling and Analysis
2.3. Evaluation Indicators
2.4. Statistical Analysis
3. Results
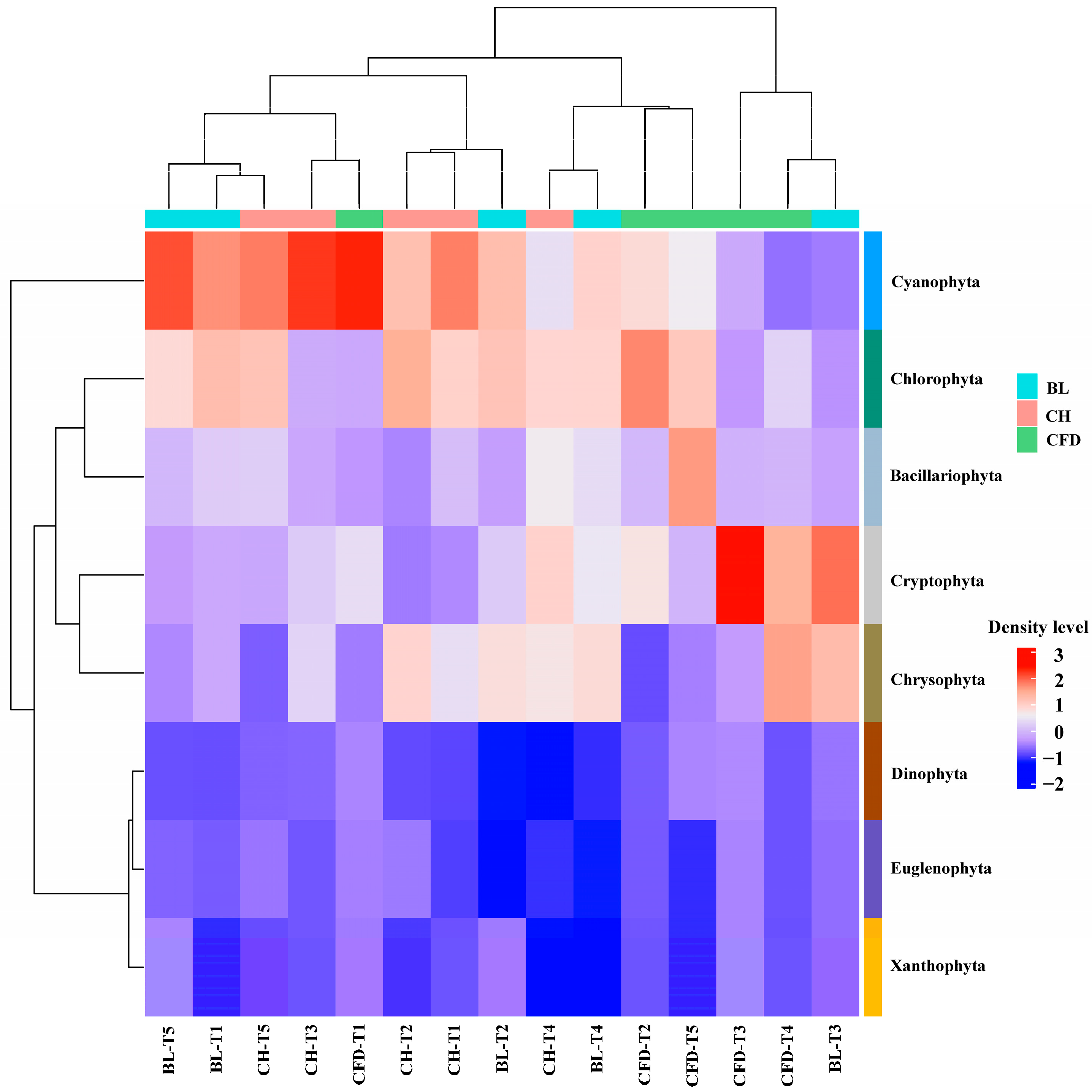
3.1. Phytoplankton Species Composition and Dominant Species
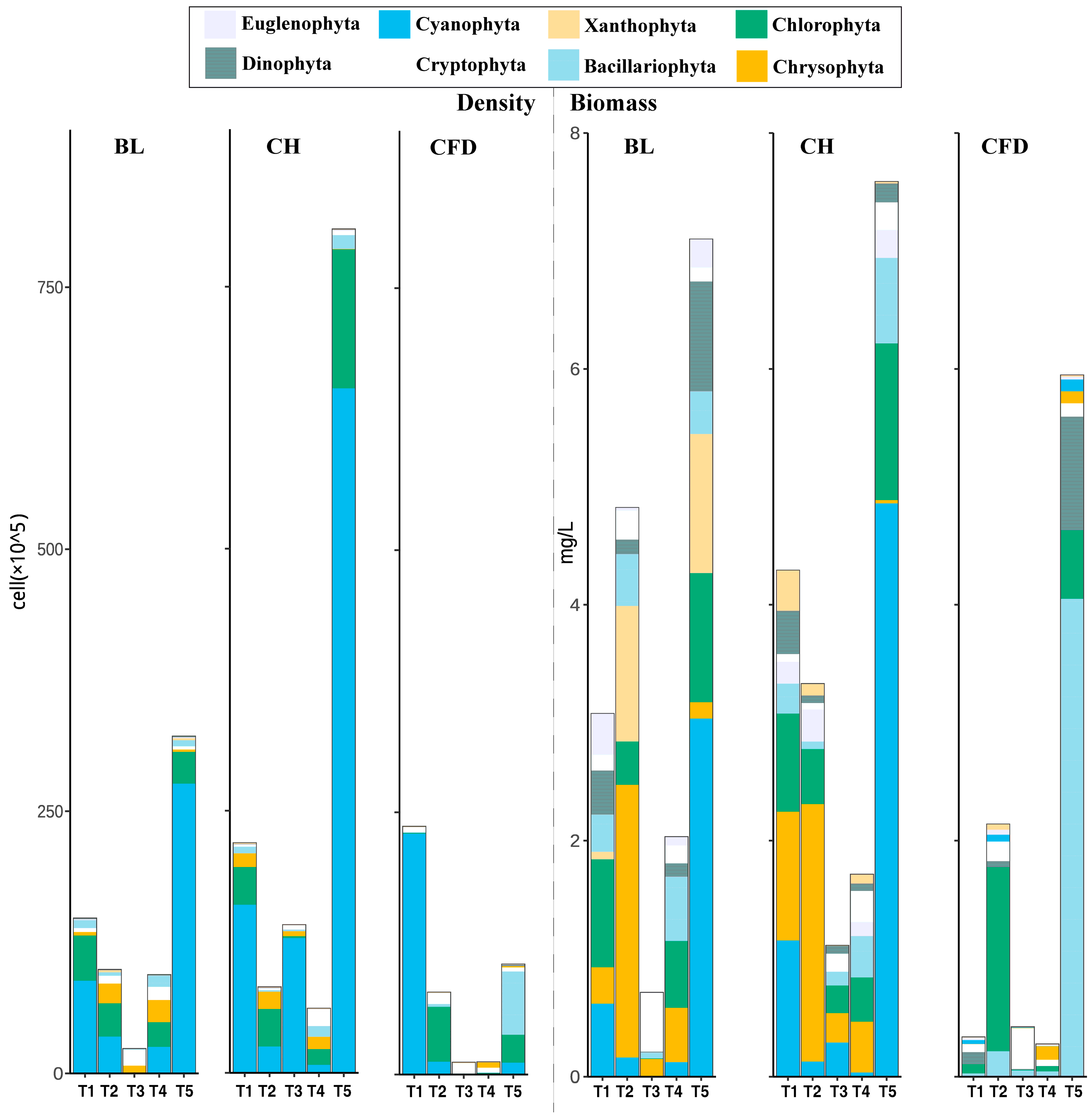
3.2. Phytoplankton Density and Biomass
3.3. Phytoplankton Community Diversity
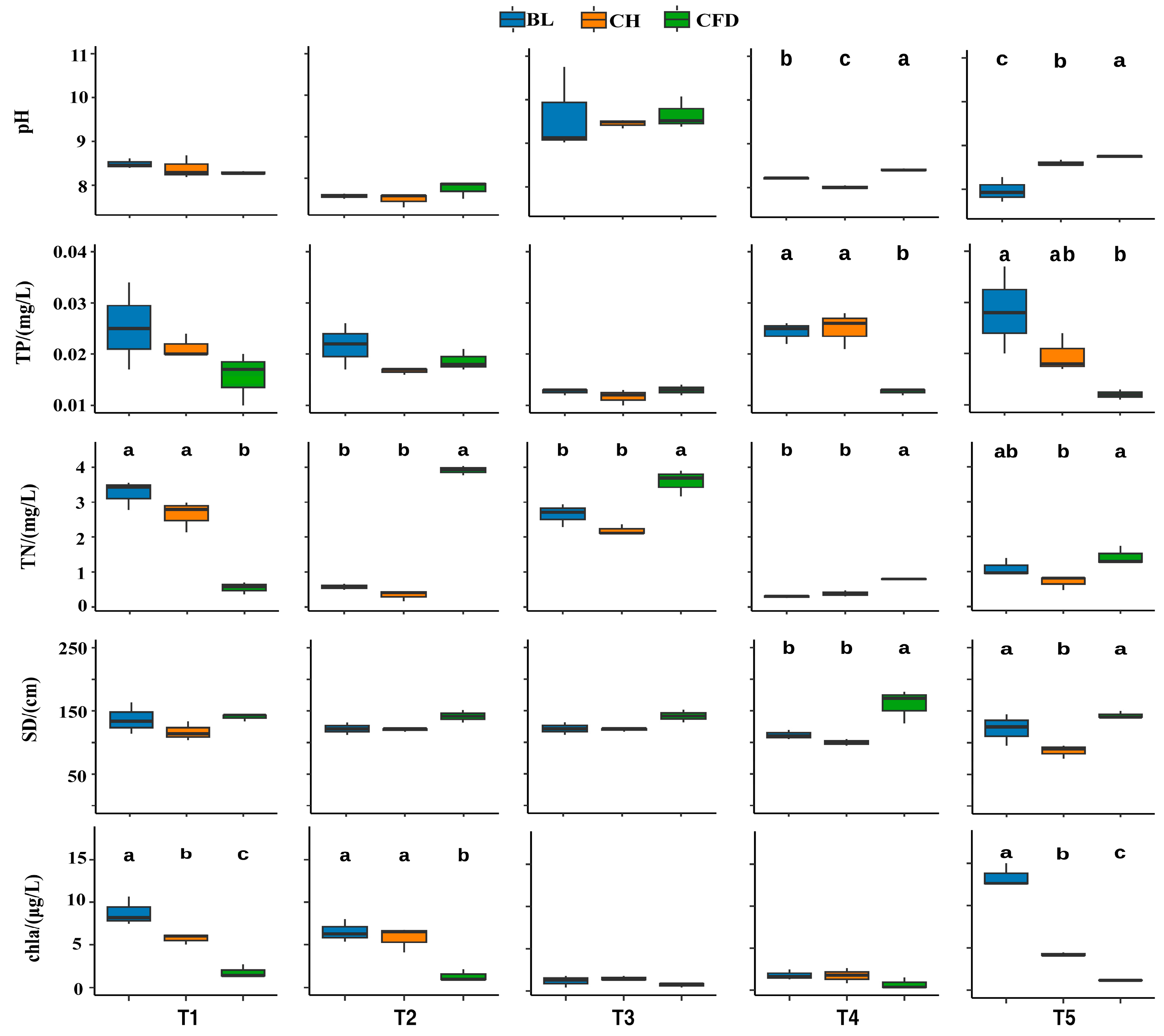
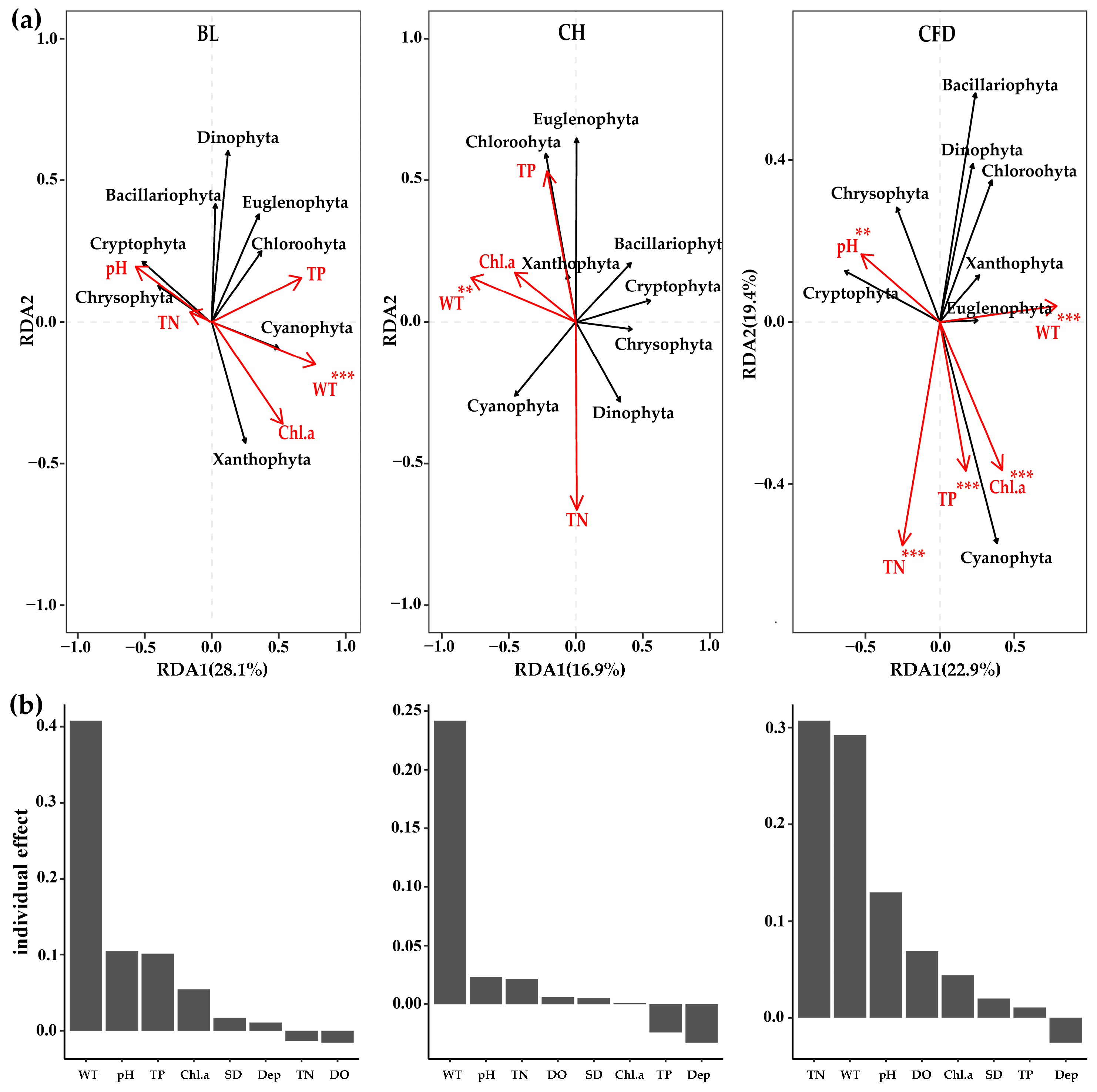
3.4. Relationship between Phytoplankton and Environmental Factors
4. Discussion
4.1. Dynamics of Phytoplankton Communities under Different Pond Conditions
4.2. Variations of Phytoplankton Community Diversity and Stability
4.3. Assessment of Water Trophic Status and the Potential Role of Biomanipulation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pond | Seasonal Comparison | Difference% | Species 1 | Contribution% | Species 2 | Contribution% | Species 3 | Contribution% |
|---|---|---|---|---|---|---|---|---|
| BL Pond | T1–T2 | 60.46 | Scenedesmus abundans | 3.50 | Scenedesmus quadricauda | 3.02 | ||
| T1–T3 | 93.89 | Microcystis aeruginosa | 9.47 | Raphidocelis subcapitata | 5.48 | Scenedesmus bijuga | 4.84 | |
| T1–T4 | 69.44 | Dactylococcopsis rhaphidioides | 6.84 | Chrysococcus diaphanus | 5.90 | Kephyrion ovale | 3.84 | |
| T1–T5 | 54.57 | |||||||
| T2–T3 | 83.16 | Scenedesmus bijuga | 7.67 | Raphidocelis subcapitata | 6.88 | Dinobryon sertularia | 6.23 | |
| T2–T4 | 73.94 | Dactylococcopsis rhaphidioides | 7.32 | Chrysococcus diaphanus | 5.51 | Kephyrion ovale | 3.99 | |
| T2–T5 | 68.05 | Microcystis wesenbergii | 9.58 | |||||
| T3–T4 | 69.13 | Dactylococcopsis rhaphidioides | 12.32 | Chroomonas caudata | 7.46 | Scenedesmus bijuga | 6.84 | |
| T3–T5 | 89.69 | Microcystis aeruginosa | 12.43 | Microcystis wesenbergii | 10.82 | Pseudanabaena limnetica | 6.82 | |
| T4–T5 | 76.03 | Microcystis aeruginosa | 9.21 | Microcystis wesenbergii | 8.37 | Dactylococcopsis rhaphidioides | 6.45 | |
| CH Pond | T1–T2 | 63.41 | Dinobryon sertularia | 7.13 | Dinobryon cylindricum | 6.45 | Merismopedia tenuissima | 4.02 |
| T1–T3 | 85.03 | Microcystis aeruginosa | 11.36 | Microcystis wesenbergii | 6.81 | Dinobryon cylindricum | 6.17 | |
| T1–T4 | 71.69 | Dinobryon cylindricum | 5.56 | Dactylococcopsis rhaphidioides | 4.51 | Kephyrion ovale | 3.99 | |
| T1–T5 | 54.86 | Cyclotella ocellata | 4.86 | Merismopedia tenuissima | 3.79 | Scenedesmus aculeolatus | 3.78 | |
| T2–T3 | 85.92 | Dinobryon sertularia | 9.24 | Scenedesmus bijuga | 9.11 | Pseudanabaena limnetica | 7.71 | |
| T2–T4 | 81.33 | Dinobryon sertularia | 6.80 | Chroomonas acuta | 5.55 | Achnanthes exigua | 5.01 | |
| T2–T5 | 72.15 | Microcystis aeruginosa | 10.46 | Microcystis wesenbergii | 9.61 | Oscillatoria amphibia | 9.35 | |
| T3–T4 | 72.26 | Dactylococcopsis rhaphidioides | 7.72 | Achnanthes exigua | 7.59 | Chroomonas acuta | 6.72 | |
| T3–T5 | 88.98 | Microcystis aeruginosa | 12.64 | Oscillatoria amphibia | 10.47 | Microcystis wesenbergii | 9.78 | |
| T4–T5 | 77.71 | Microcystis aeruginosa | 11.09 | Oscillatoria amphibia | 9.32 | Microcystis wesenbergii | 8.71 | |
| CFD Pond | T1–T2 | 78.84 | Scenedesmus bijuga | 9.88 | Raphidocelis subcapitata | 5.77 | Coelastrum reticulatum | 4.75 |
| T1–T3 | 74.54 | Merismopedia minima | 47.77 | Chroomonas caudata | 16.55 | Microcystis aeruginosa | 7.66 | |
| T1–T4 | 77.34 | Merismopedia minima | 41.59 | Kephyrion ovale | 11.69 | Microcystis aeruginosa | 7.40 | |
| T1–T5 | 83.55 | Synedra acus | 11.92 | Cyclotella ocellata | 8.22 | Chlorella vulgaris | 6.14 | |
| T2–T3 | 81.82 | Scenedesmus bijuga | 11.00 | Raphidocelis subcapitata | 6.40 | Coelastrum reticulatum | 5.73 | |
| T2–T4 | 85.96 | Scenedesmus bijuga | 9.91 | Microcystis aeruginosa | 5.29 | Raphidocelis subcapitata | 5.21 | |
| T2–T5 | 65.87 | |||||||
| T3–T4 | 63.62 | Chroomonas caudata | 24.06 | Kephyrion ovale | 20.08 | Chrysococcus diaphanus | 8.04 | |
| T3–T5 | 85.96 | Synedra acus | 13.71 | Cyclotella ocellata | 9.27 | Chlorella vulgaris | 7.30 | |
| T4–T5 | 86.30 | Synedra acus | 12.96 | Cyclotella ocellata | 8.30 | Chlorella vulgaris | 6.63 |
References
- Schindler, D.W. Recent advances in the understanding and management of eutrophication. Limnol. Oceanogr. 2006, 51, 356–363. [Google Scholar] [CrossRef]
- Conley, D.J.; Paerl, H.W.; Howarth, R.W.; Boesch, D.F.; Seitzinger, S.P.; Havens, K.E.; Lancelot, C.; Likens, G.E. Controlling eutrophication: Nitrogen and phosphorus. Science 2009, 323, 1014–1015. [Google Scholar] [CrossRef]
- Søndergaard, M.; Jensen, J.P.; Jeppesen, E. Role of sediment and internal loading of phosphorus in shallow lakes. Hydrobiologia 2007, 581, 373–385. [Google Scholar] [CrossRef]
- Smith, V.H.; Schindler, D.W. Eutrophication science: Where do we go from here? Trends Ecol. Evol. 2009, 24, 201–207. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Kitchell, J.F.; Hodgson, J.R. Cascading trophic interactions and lake productivity. BioScience 1985, 35, 634–639. [Google Scholar] [CrossRef]
- McQueen, D.J. Manipulating lake community structure: Where do we go from here? Freshw. Biol. 1990, 23, 613–620. [Google Scholar] [CrossRef]
- Jeppesen, E.; Søndergaard, M.; Søndergaard, M.; Christoffersen, K. (Eds.) The Structuring Role of Submerged Macrophytes in Lakes; Springer: New York, NY, USA, 1997. [Google Scholar]
- Liu, J.K.; Xie, P. Unraveling the enigma of the disappearance of water bloom from the East Lake (Lake Donghu) of Wuhan. Resour. Environ. Yangtze Basin 1999, 3, 85–92. [Google Scholar]
- Dionisio Pires, L.M.; Ibelings, B.W.; Brehm, M.; Van Donk, E. Comparing Grazing on Lake Seston by Dreissena and Daphnia: Lessons for Biomanipulation. Microb. Ecol. 2005, 50, 242–252. [Google Scholar] [CrossRef]
- Søndergaard, M.; Jeppesen, E.; Lauridsen, T.L. Lake restoration: Successes, failures and long-term effects. J. Appl. Ecol. 2008, 45, 782–797. [Google Scholar] [CrossRef]
- Qin, B.Q.; Xu, P.Z.; Wu, Q.L.; Luo, L.C.; Zhang, Y.L. Environmental issues of lake Taihu, China. Hydrobiologia 2007, 581, 3–14. [Google Scholar] [CrossRef]
- Zhang, Y.; Qin, B.; Chen, W.; Zhu, G. A study of the diatom assemblages in the sediments of the northern part of Taihu Lake, China. J. Paleolimnol. 2010, 43, 415–426. [Google Scholar]
- Fu, Z.Y. Application of pre-reservoir in water resource allocation and water environment protection of Yuqiao Reservoi. Haihe Water Resour. 2019, 1, 44–45. [Google Scholar]
- Hu, H.J.; Wei, Y.X. The Freshwater Algae of China-Systematics, Taxonomy and Ecology, 1st ed.; Science Press: Beijing, China, 2006; pp. 1–1023. [Google Scholar]
- Han, M.S.; Shu, Y.F. Chinese Freshwater Organisms Atlas, 1st ed.; Ocean Press: Beijing, China, 1995; pp. 1–390. [Google Scholar]
- Huang, X.F. Survey, Observation and Analysis of Lake Ecology, 1st ed.; Standards Press of China: Beijing, China, 2000; pp. 72–77. [Google Scholar]
- The State Environmental Protection Administration. Water and Wastewater Monitoring and Analysis Method, 4th ed.; China Environmental Science Press: Beijing, China, 2002; pp. 1–836. [Google Scholar]
- Dufrêne, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Margalef, R. Information theory in ecology. Gen. Syst. 1958, 3, 36–71. [Google Scholar]
- Pielou, E.C. Species-diversity and pattern-diversity in the study of ecological succession. J. Theor. Biol. 1996, 10, 370–383. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.C.; Tu, Q.Y. Specification For Lake Eutrophication Survey, 2nd ed.; China Environmental Science Press: Beijing, China, 1990; pp. 286–291. [Google Scholar]
- Wang, M.C.; Liu, X.Q.; Zhang, J.H. Evaluate method and classification standard on lake eutrophication. Environ. Monit. China 2002, 18, 47–49. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing, Reference Index Version 4.2.2; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 20 November 2023).
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Nie, Y. Phytoplankton Community Structure and Eutrophication Status in Yuqiao Reservoir. Master’s Dissertation, Tianjin University of Science and Technology, Tianjin, China, 2016. [Google Scholar]
- Gao, K.; Li, Z.L.; Zhao, X.H.; Zhang, X.X.; Mei, P.W.; Zhang, Z. Spatiotemporal dynamics of and influencing factors on the phytoplankton community in the Yuqiao Reservoir. J. Agric. Resour. Environ. 2023, 1–19. Available online: https://link.cnki.net/urlid/12.1437.s.20230811.1642.002 (accessed on 20 November 2023).
- Liu, J.K.; Xie, P. Direct control of microcystis bloom through the use of Planktivorous Carp-closure Experiments and Lake Fishery. Ecol. Sci. 2003, 3, 193–198. [Google Scholar]
- Ghadouani, A.; Pinel-Alloul, B.; Prepas, E.E. Effects of experimentally induced cyanobacterial blooms on crustacean zooplankton communities: Cyanobacteria effects on herbivores. Freshw. Biol. 2003, 48, 363–381. [Google Scholar] [CrossRef]
- Mehner, T.; Kasprzak, P.; Wysujack, K.; Laude, U.; Koschel, R. Restoration of a stratified lake (Feldberger Haussee, Germany) by a combination of nutrient load reduction and long-term biomanipulation. Int. Rev. Hydrobiol. 2001, 86, 253–265. [Google Scholar] [CrossRef]
- Wu, Z.B.; Qiu, D.R.; He, F.; Fu, G.P.; Cheng, S.P.; Ma, J.M. Effects of rehabilitation of submerged macrophytes on nutrient level of a eutrophic lake. Chin. J. Appl. Ecol. 2003, 14, 1351–1353. [Google Scholar]
- Shi, X.L.; Yang, J.S.; Chen, K.N.; Zhang, M.; Yang, Z.; Yu, Y. Review on the control and mitigation strategies of lake cyanobacterial blooms. J. Lake Sci. 2022, 34, 349–375. [Google Scholar]
- Ma, J.R.; Deng, J.M.; Qin, B.Q.; Long, S.X. Progress and prospects on cyanobacteria bloom-forming mechanism in lakes. Acta Ecol. Sin. 2013, 33, 3020–3030. [Google Scholar]
- Kuang, Q.J.; Ma, P.M.; Hu, Z.Y.; Zhou, G.J. Study on the evaluation and treatment of lake eutrophication by means of algae biology. J. Saf. Environ. 2005, 2, 87–91. [Google Scholar]
- Jia, H.Y.; Xu, J.F.; Lei, J.S. Relationship of community structure of phytoplankton and environmental factors in Danjiangkou Reservoir bay. Yangtze River 2019, 50, 52–58. [Google Scholar]
- Bai, L.J.; Zhang, Z.Y.; Wang, L.; Zhou, B.B.; Li, Y.C.; Shao, X.D.; Su, S.; Liu, Q. Analysis of plankton community and fishery resources in Xiangshui Reservoir. J. Dalian Ocean. Univ. 2020, 35, 280–287. [Google Scholar]
- Drenner, R.W.; Hambright, R.K.D. Piscivores, Trophic Cascades, and Lake Management. Sci. World J. 2002, 2, 284–307. [Google Scholar] [CrossRef]
- Cui, F.Y.; Lin, T.; Ma, F.; Zhang, L.Q. Experimental Studies on Biomanipulation of Silver Carp and Bighead Carp in Water Resources Management. J. Nanjing Univ. Sci. Technol. 2004, 06, 668–672. [Google Scholar]
- Chen, S.L.; Liu, X.F.; Hua, L. The role of Silver carp and Bighead in the cycling of Nitrogen and Phosphorus in the East Lake ecosystem. Acta Hydrobiol. Sin. 1991, 1, 8–26. [Google Scholar]





| Fish Species | Size of Stocked Fish (cm) | Density of Stocked Fish (ind/hm2) | |
|---|---|---|---|
| CH Pond 1 | CFD Pond 2 | ||
| Topmouth culter (Culter alburnus) | 5 | 120 | 120 |
| Chinese perch (Siniperca chuatsi) | 10 | 60 | 60 |
| Grass carp (Ctenopharyngodon idellus) | 8 | 105 | |
| Bream (Megalobrama amblycephala) | 5 | 300 | |
| Silver carp (Hypophthalmichthys molitrix) | 20 | 225 | |
| Bighead carp (Aristichthys nobilis) | 20 | 75 | |
| Yellow tail (Xenocypris microlepis) | 9 | 225 | |
| Time Period | Number of Species in BL Pond 1 | Number of Species in CH Pond 2 | Number of Species in CFD Pond 3 | Number of Shared Species |
|---|---|---|---|---|
| T1 (Late June 2022) | 91 | 90 | 46 | 25 |
| T2 (Late September 2022) | 71 | 65 | 73 | 29 |
| T3 (Late November 2022) | 37 | 38 | 36 | 17 |
| T4 (Late March 2023) | 72 | 73 | 41 | 29 |
| T5 (Early July 2023) | 73 | 50 | 56 | 36 |
| Total | 162 | 159 | 131 | 99 |
| Dominant Species | BL Pond 1 | CH Pond 2 | CFD Pond 3 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T1 | T2 | T3 | T4 | T5 | T1 | T2 | T3 | T4 | T5 | T1 | T2 | T3 | T4 | T5 | |
| Cyanophyta | |||||||||||||||
| Microcystis aeruginosa | 0.43 | 0.14 | 0.31 | 0.59 | 0.04 | 0.55 | 0.05 | ||||||||
| Pseudanabaena limnetica | 0.03 | 0.04 | 0.02 | 0.04 | 0.89 | 0.02 | |||||||||
| Cylindrospermum majus | 0.03 | 0.14 | |||||||||||||
| Microcystis wesenbergii | 0.06 | 0.07 | 0.11 | ||||||||||||
| Microcystis marginata | 0.03 | ||||||||||||||
| Merismopedia minima | 0.04 | 0.03 | 0.04 | 0.96 | 0.52 | ||||||||||
| Merismopedia tenuissima | 0.04 | 0.04 | |||||||||||||
| Dactylococcopsis rhaphidioides | 0.25 | 0.03 | 0.11 | ||||||||||||
| Oscillatoria amphibia | |||||||||||||||
| Dolichospermum bergii | 0.02 | ||||||||||||||
| Bacillariophyta | |||||||||||||||
| Achnanthes exigua | 0.03 | 0.05 | 0.11 | ||||||||||||
| Synedra acus | 0.02 | 0.03 | 0.03 | 0.45 | |||||||||||
| Cyclotella ocellata | 0.12 | ||||||||||||||
| Chrysophyta | |||||||||||||||
| Dinobryon cylindricum | 0.05 | ||||||||||||||
| Dinobryon sertularia | 0.16 | 0.02 | 0.20 | ||||||||||||
| Kephyrion ovale | 0.03 | 0.05 | 0.08 | 0.29 | |||||||||||
| Chrysococcus diaphanus | 0.27 | 0.16 | 0.03 | 0.05 | 0.09 | ||||||||||
| Dinobryon divergens | 0.03 | 0.04 | |||||||||||||
| Dinobryon bavaricum | 0.03 | ||||||||||||||
| Chlorophyta | |||||||||||||||
| Raphidocelis subcapitata | 0.07 | 0.09 | 0.04 | 0.04 | 0.04 | 0.10 | 0.03 | 0.02 | 0.07 | ||||||
| Scenedesmus abundans | 0.03 | ||||||||||||||
| Scenedesmus bijuga | 0.05 | 0.12 | 0.07 | 0.04 | 0.23 | 0.06 | 0.06 | 0.25 | 0.05 | ||||||
| Crucigenia quadrata | 0.03 | 0.03 | 0.08 | 0.06 | 0.04 | ||||||||||
| Scenedesmus quadricauda | 0.02 | 0.02 | 0.03 | ||||||||||||
| Crucigenia tetrapedia | 0.03 | 0.02 | 0.03 | 0.04 | |||||||||||
| Coelastrum reticulatum | 0.05 | ||||||||||||||
| Coelastrum microporum | 0.04 | ||||||||||||||
| Planctonema lauterbornii | 0.04 | ||||||||||||||
| Chlorella vulgaris | 0.06 | 0.07 | |||||||||||||
| Schroederia setigera | 0.05 | ||||||||||||||
| Cryptophyta | |||||||||||||||
| Chroomonas acuta | 0.02 | 0.31 | 0.13 | 0.04 | 0.22 | 0.11 | 0.42 | 0.35 | 0.03 | ||||||
| Chroomonas caudata | 0.03 | 0.33 | 0.02 | 0.46 | |||||||||||
| Pond | Time Period | Shannon–Wiener Diversity Index (H′) | Simpson’s Diversity Index (D) | Margalef’s Richness Index (Dm) | Pielou’s Evenness Index (J) |
|---|---|---|---|---|---|
| BL Pond | T1 | 2.54 | 0.797 | 6.34 | 0.56 |
| T2 | 2.99 | 0.920 | 5.07 | 0.70 | |
| T3 | 1.54 | 0.724 | 2.91 | 0.43 | |
| T4 | 2.70 | 0.880 | 5.16 | 0.63 | |
| T5 | 1.71 | 0.688 | 4.81 | 0.40 | |
| CH Pond | T1 | 1.90 | 0.637 | 6.10 | 0.42 |
| T2 | 2.52 | 0.868 | 4.70 | 0.60 | |
| T3 | 0.63 | 0.202 | 2.61 | 0.17 | |
| T4 | 2.92 | 0.908 | 5.40 | 0.68 | |
| T5 | 1.53 | 0.652 | 3.65 | 0.37 | |
| CFD Pond | T1 | 0.23 | 0.075 | 3.07 | 0.06 |
| T2 | 2.89 | 0.903 | 5.31 | 0.67 | |
| T3 | 1.32 | 0.610 | 3.01 | 0.37 | |
| T4 | 2.09 | 0.780 | 3.43 | 0.56 | |
| T5 | 2.27 | 0.770 | 3.97 | 0.56 |
| Time Period | BL Pond | CH Pond | CFD Pond |
|---|---|---|---|
| T1 (Late June 2022) | 50.4 | 52.1 | 39.9 |
| T5 (Early July 2023) | 52.3 | 46.4 | 40.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Yang, J.; Lin, X.; Tian, B.; Zhang, T.; Ye, S. Phytoplankton Community Dynamics in Ponds with Diverse Biomanipulation Approaches. Diversity 2024, 16, 75. https://doi.org/10.3390/d16020075
Zhang Y, Yang J, Lin X, Tian B, Zhang T, Ye S. Phytoplankton Community Dynamics in Ponds with Diverse Biomanipulation Approaches. Diversity. 2024; 16(2):75. https://doi.org/10.3390/d16020075
Chicago/Turabian StyleZhang, Yantao, Jie Yang, Xiaoman Lin, Biao Tian, Tanglin Zhang, and Shaowen Ye. 2024. "Phytoplankton Community Dynamics in Ponds with Diverse Biomanipulation Approaches" Diversity 16, no. 2: 75. https://doi.org/10.3390/d16020075