
Diversity of Silica-Scaled Chrysophytes (Stramenopiles: Chrysophyceae) from Indonesian Papua

Abstract
:1. Introduction
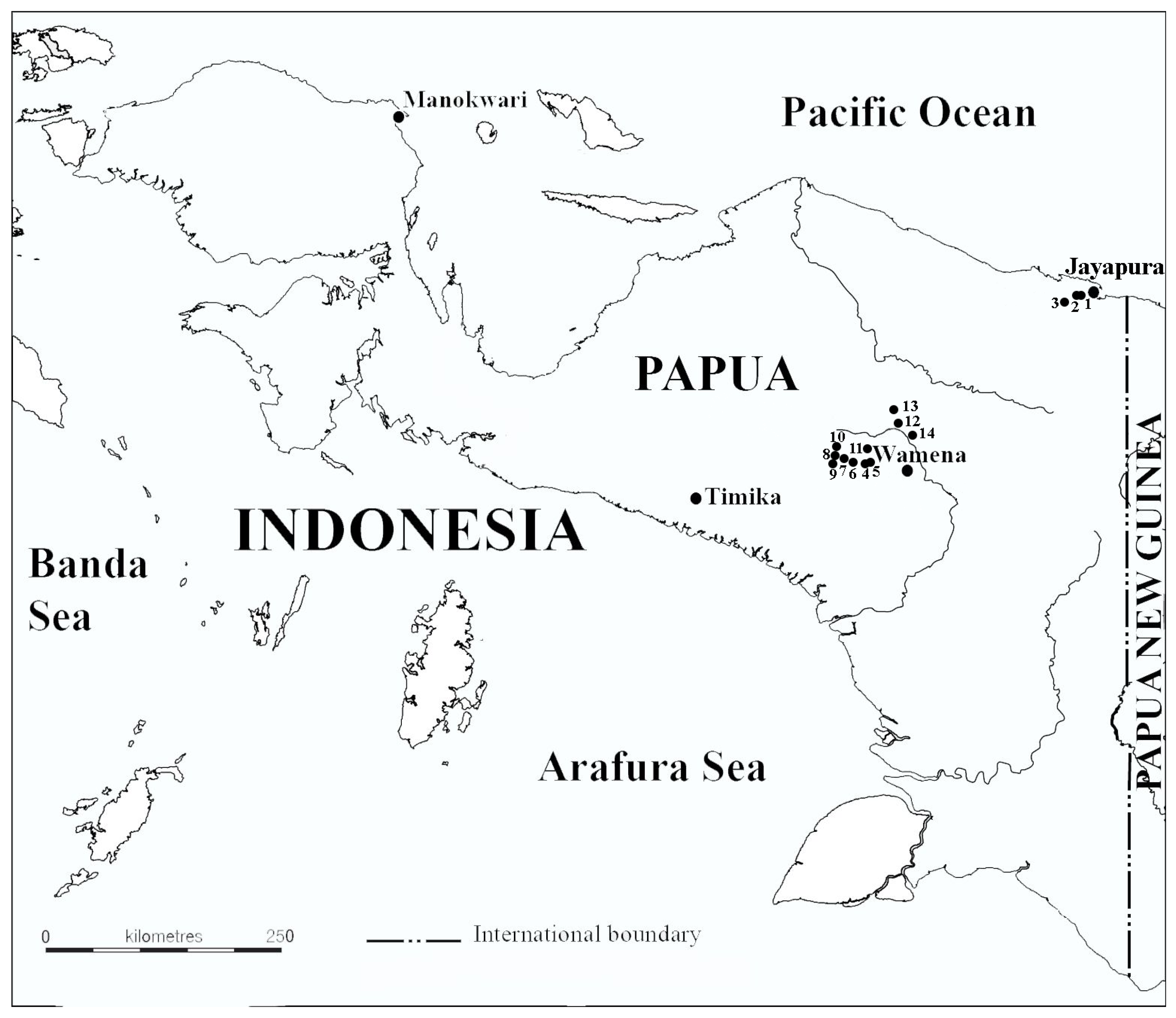
2. Materials and Methods
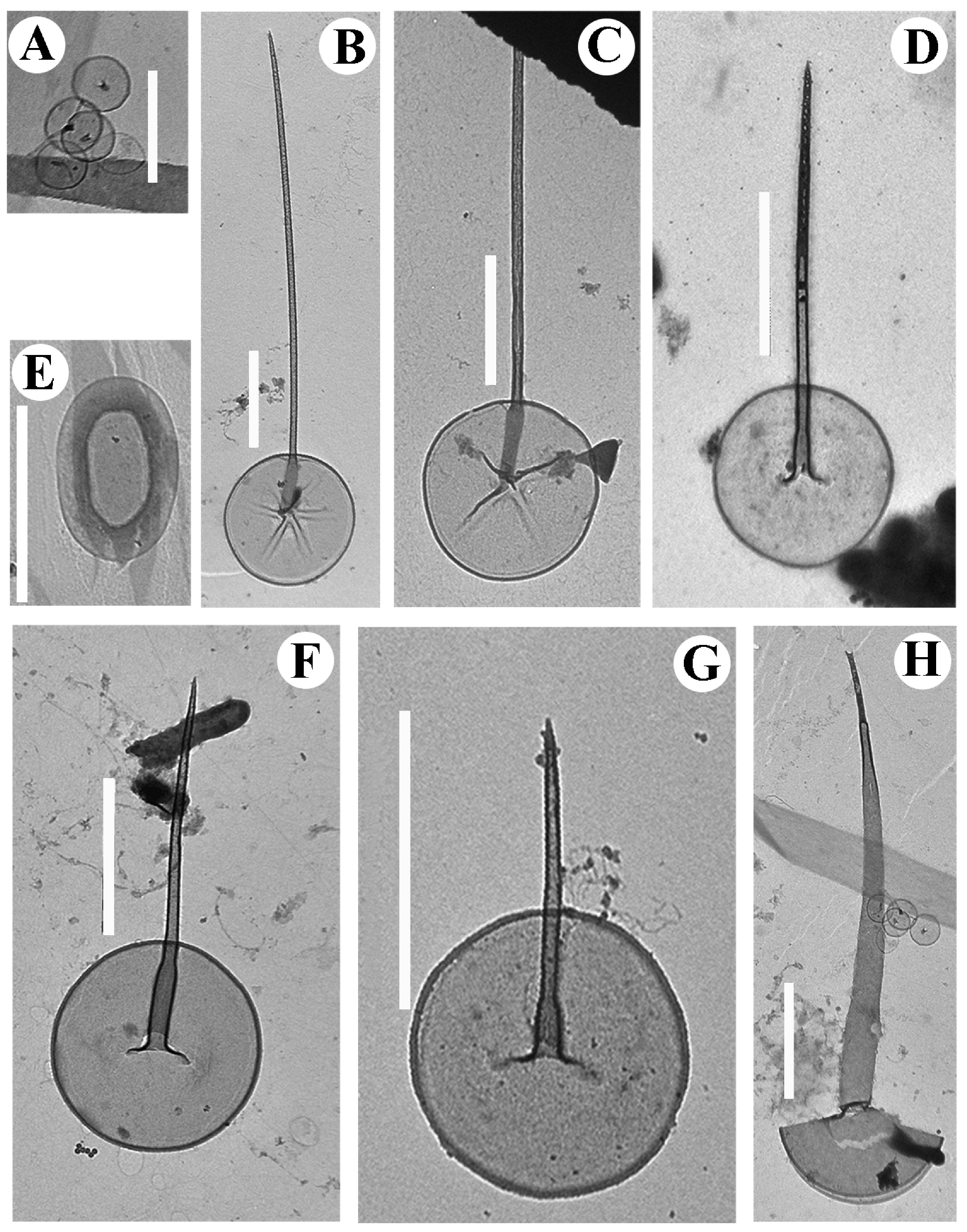
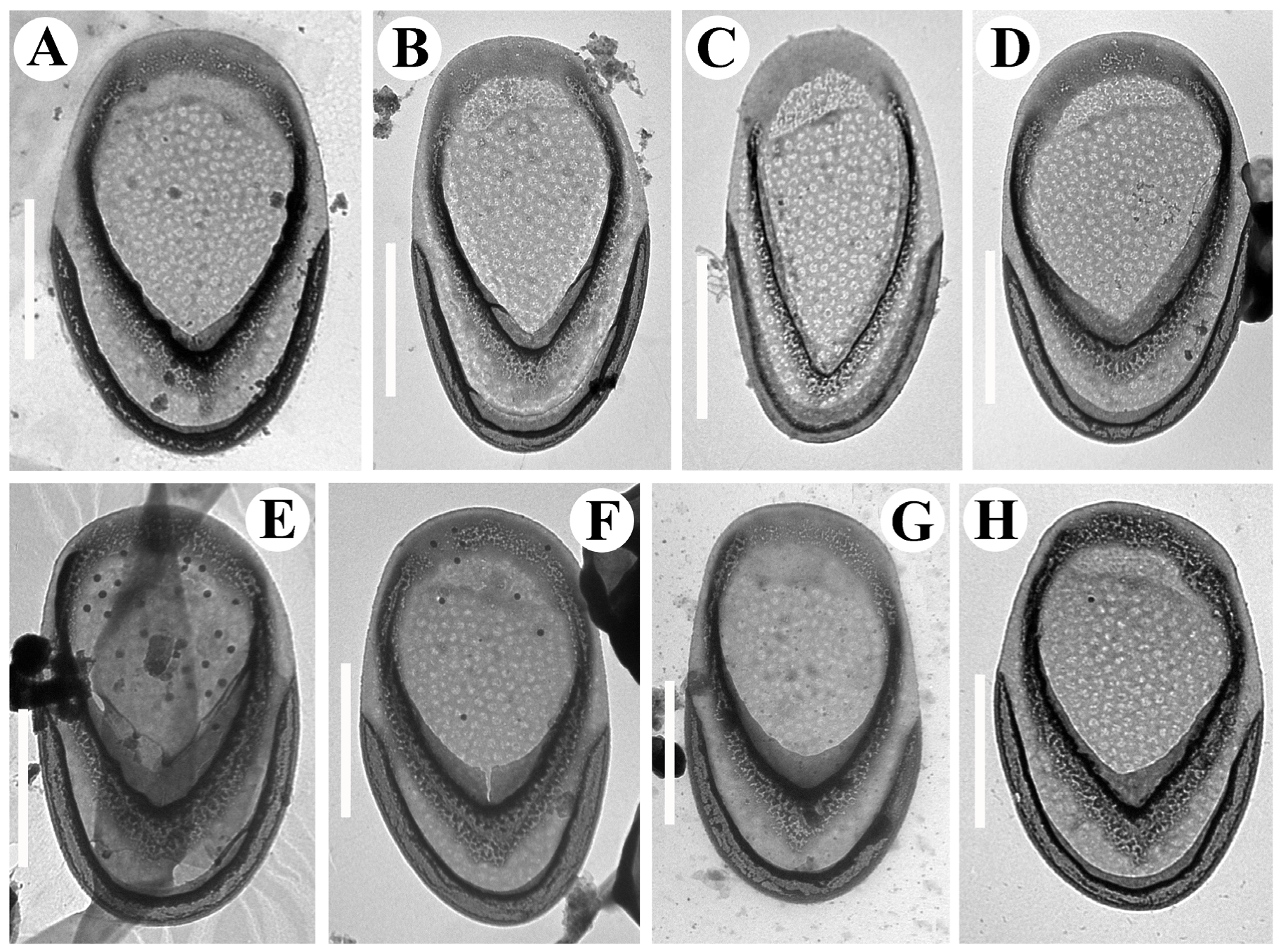
3. Results
4. Discussion
4.1. Comparison of Silica-Scaled Chrysophyte Floras of Indonesian Papua and Papua New Guinea
4.2. Endemism in Silica-Scaled Chrysophytes from Indonesian Papua
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siver, P.A.; Jo, B.Y.; Kim, J.I.; Shin, W.; Lott, A.M.; Wolfe, A.P. Assessing the evolutionary history of the class Synurophyceae (Heterokonta) using molecular, morphometric, and paleobiological approaches. Am. J. Bot. 2015, 102, 921–941. [Google Scholar] [CrossRef] [PubMed]
- Scoble, J.M.; Cavalier-Smith, T. Scale evolution in Paraphysomonadida (Chrysophyceae): Sequence phylogeny and revised taxonomy of Paraphysomonas, new genus Clathromonas, and 25 new species. Eur. J. Protistol. 2014, 50, 551–592. [Google Scholar] [CrossRef] [PubMed]
- Kristiansen, J. Biogeography of silica-scaled chrysophytes. Nova Hedw. Beih. 2001, 122, 23–39. [Google Scholar]
- Řezáčová, M.; Neustupa, J. Distribution of the genus Mallomonas (Synurophyceae)–ubiquitous dispersal in microorganisms evaluated. Protist 2007, 158, 29–37. [Google Scholar] [CrossRef]
- Siver, P.A.; Lott, A.M. Biogeographic patterns in scaled chrysophytes from the east coast of North America. Freshw. Biol. 2012, 57, 451–466. [Google Scholar] [CrossRef]
- Kristiansen, J. Dispersal and biogeography of silica-scaled chrysophytes. Biodivers. Conserv. 2008, 17, 419–426. [Google Scholar] [CrossRef]
- Compère, P. Mallomonas bronchartiana, Chrysophyceé nouvelle du lac Tchad. Bull. Jard. Bot. Nat. Belg. 1974, 44, 61–63. [Google Scholar] [CrossRef]
- Wei, Y.-X.; Yuan, X.-P. Studies on silica-scaled chrysophytes from the tropics and subtropics of China. Nova Hedw. Beih. 2001, 122, 169–187. [Google Scholar]
- Gusev, E.S.; Nguyen, T.H.T. Silica-scaled chrysophytes (Chrysophyceae and Synurophyceae) from Vietnam (Khanh Hoa and Quang Nam provinces). Nova Hedw. 2011, 93, 191–199. [Google Scholar] [CrossRef]
- Gusev, E.S. Studies on synurophycean algae from mangrove wetlands (Vietnam). Nova Hedw. Beih. 2013, 142, 87–98. [Google Scholar]
- Wei, Y.-X.; Yuan, X.-P.; Kristiansen, J. Silica-scaled chrysophytes from Hainan, Guangdong Provinces and Hong Kong Special Administrative Region, China. Nord. J. Bot. 2014, 32, 881–896. [Google Scholar] [CrossRef]
- Gusev, E.S.; Doan-Nhu, H.; Nguyen-Ngoc, L. Silica-scaled chrysophytes from Cat Tien National Park (Dong Nai Province, Viet Nam). Nova Hedw. 2017, 105, 347–364. [Google Scholar] [CrossRef]
- Gusev, E.S.; Doan-Nhu, H.; Nguyen-Ngoc, L.; Guseva, E.E.; Luom, P.T. Silica-scaled chrysophytes from Cam Ranh Peninsula (Khanh Hoa Province, Vietnam). Nova Hedw. Beih. 2019, 148, 63–76. [Google Scholar] [CrossRef]
- Gusev, E.; Martynenko, N.; Tran, H. Studies on Algae from the Order Synurales (Chrysophyceae) in Northern Vietnam. Diversity 2021, 13, 602. [Google Scholar] [CrossRef]
- Doan-Nhu, H.; Tinh, T.T.; Gusev, E.S.; Kulikovskiy, M.S.; Phan-Tan, L.; Nguyen-Ngoc, L. Taxonomic composition of silica-scaled chrysophytes in a tropical mountain reservoir. Inl. Wat. Biol. 2021, 14, 490–499. [Google Scholar] [CrossRef]
- Gusev, E.; Martynenko, N. Diversity of Silica-Scaled Chrysophytes in Central Vietnam. Water 2022, 14, 65. [Google Scholar] [CrossRef]
- Němcová, Y.; Bulant, P.; Kristiansen, J. Mallomonas solea-ferrea and Mallomonas siveri (Chrysophyceae/Synurophyceae): Two new taxa from the Western Cape (South Africa). Nova Hedw. 2011, 93, 375–384. [Google Scholar] [CrossRef]
- Němcová, Y.; Kreidlová, J. Two new species of Mallomonas (Chrysophyceae: Synurales): Mallomonas temonis and Mallomonas divida. Phytotaxa 2013, 87, 11–18. [Google Scholar] [CrossRef]
- Piątek, J. Mallomonas camerunensis sp. nov. (Chrysophyceae, Stramenopiles) from a shallow puddle in the Guineo-Congolian rainforest (Cameroon). Pol. Bot. J. 2015, 60, 119–126. [Google Scholar] [CrossRef]
- Gusev, E.S. A new species of the genus Mallomonas (Synurophyceae), Mallomonas spinosa sp. nov., from Vietnam. Phytotaxa 2012, 66, 1–5. [Google Scholar] [CrossRef]
- Gusev, E.S. A new species of the genus Mallomonas (Synurales, Chrysophyceae), Mallomonas fimbriata sp. nov. Phytotaxa 2015, 195, 291–296. [Google Scholar] [CrossRef]
- Gusev, E.; Siver, P.A. Mallomonas neoampla sp. nov. from Vietnam, a new species that bridges the gap between fossil and modern taxa. Nova Hedw. 2017, 104, 521–528. [Google Scholar] [CrossRef]
- Gusev, E.S.; Kulikovskiy, M.S. Two new species of genus Mallomonas from swamp localities in Vietnam. Phytotaxa 2020, 468, 121–129. [Google Scholar] [CrossRef]
- Gusev, E.S.; Siver, P.A.; Shin, W. Mallomonas bronchartiana Compère revisited: Two new species described from Asia. Cryptogam. Algol. 2017, 38, 3–16. [Google Scholar] [CrossRef]
- Gusev, E.S.; Kapustin, D.A.; Martynenko, N.A.; Guseva, E.E.; Kulikovskiy, M.S. Mallomonas gusakovii sp. nov. (Chrysophyceae, Synurales), a new species from Phu Quoc Island, Vietnam. Phytotaxa 2019, 406, 199–205. [Google Scholar] [CrossRef]
- Gusev, E.; Guseva, E.; Kezlya, E.; Kulikovskiy, M. Mallomonas minuscula sp. nov. (Synurales, Chrysophyceae), a new member in the section Torquatae from Vietnam. Fottea 2019, 19, 132–137. [Google Scholar] [CrossRef]
- Gusev, E.; Kezlya, E. Mallomonas lusca sp. nov.—A rare species from Southeast Asia. Phytotaxa 2021, 529, 105–112. [Google Scholar] [CrossRef]
- Gusev, E.S.; Kulikovskiy, M.S. Mallomonas siderea sp. nov. (Synurales, Chrysophyceae), a new tropical species from the section Torquatae. Nova Hedw. 2021, 113, 291–301. [Google Scholar] [CrossRef]
- Krieger, W. Die Desmidiaceen der Deutschen limnologischen Sunda-Expedition. Arch. Hydrobiol. Suppl. 1932, 11, 129–230. [Google Scholar]
- Behre, K. Die Süsswasseralgaen der Wallacea-Expedition (ohne die Diatomeen und Peridineen). Arch. Hydrobiol. Suppl. 1956, 23, 1–104. [Google Scholar]
- Scott, A.M.; Prescott, G.W. Indonesian desmids. Hydrobiologia 1961, 17, 1–132. [Google Scholar] [CrossRef]
- Conrad, W. Flagellates des Iles de la Sonde (Euglénacées). Bull. Mus. R. Hist. Nat. Belg. 1938, 14, 1–20. [Google Scholar]
- Hustedt, F. Die fossile Diatomeenfl ora in den Ablagerungen des Tobasees auf Sumatra. Arch. Hydrobiol. Suppl. 1935, 14, 143–192. [Google Scholar]
- Hustedt, F. Systematische und ökologische Untersuchungen über die Diatomeenflora von Java, Bali und Sumatra nach dem Material der Deutschen Limnologischen Sunda-Expedition. Teil, I. Systematischer Teil. Arch. Hydrobiol. Suppl. 1938, 15, 393–506. [Google Scholar]
- Hustedt, F. Süßwasser-Diatomeen des indomalayischen Archipels und der Hawaii-Inseln. Int. Rev. ges. Hydrobiol. Hydrogr. 1942, 42, 1–252. [Google Scholar] [CrossRef]
- Scott, A.M.; Prescott, G.W. Notes on Indonesian freshwater algae II. Ichthyodontum, a new desmid genus from Sumatra. Reinwardtia 1956, 3, 351–362. [Google Scholar]
- Hamsher, S.E.; Graeff, C.L.; Stepanek, J.G.; Kociolek, J.P. Frustular morphology and polyphyly in freshwater Denticula (Bacillariophyceae) species, and the description of Tetralunata gen. nov. (Epithemiaceae, Rhopalodiales). Pl. Ecol. Evol. 2014, 147, 346–365. [Google Scholar] [CrossRef]
- Kapustin, D.A.; Kulikovskiy, M.S.; Kociolek, J.P. Celebesia gen. nov., a new cymbelloid diatom genus from the ancient Lake Matano (Sulawesi Island, Indonesia). Nova Hedw. Beih. 2017, 146, 147–155. [Google Scholar] [CrossRef]
- Kapustin, D.A.; Kociolek, J.P.; Glushchenko, A.M.; Kulikovskiy, M.S. Four new species of Cymbella (Bacillariophyta) from the ancient Malili Lakes (Sulawesi Island, Indonesia). Bot. Zhurn. 2019, 104, 766–780. [Google Scholar] [CrossRef]
- Kapustin, D.A.; Kociolek, J.P.; Glushchenko, A.M.; Kulikovskiy, M.S. A rediscovery of Cymbella mirabilis Hustedt, a rare endemic diatom, and description of Alveocymba gen. nov. Diat. Res. 2020, 35, 281–287. [Google Scholar] [CrossRef]
- Kapustin, D.A.; Glushchenko, A.M.; Kociolek, J.P.; Kulikovskiy, M.S. Encyonopsis indonesica sp. nov., a new diatom from the ancient lake Matano (Sulawesi, Indonesia). PhytoKeys 2021, 175, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kapustin, D.A.; Glushchenko, A.M.; Kulikovskiy, M.S. Achnanthidium bratanense sp. nov. (Bacillariophyceae, Achnanthidiaceae), a new diatom from the Lake Bratan (Bali, Indonesia). PhytoKeys 2022, 188, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Gusev, E.S.; Gusakov, V.A.; Guseva, E.E.; Kulikovskiy, M.S.; Tsvetkov, A.I.; Định, C.N. Flora of Silica-Scaled Chrysophytes (Chrysophyceae: Synurales, Paraphysomonadales) of the Mekong Delta. Inl. Wat. Biol. 2020, 13, 349–357. [Google Scholar] [CrossRef]
- Němcová, Y.; Rott, E. Diversity of silica-scaled chrysophytes in high-altitude Alpine sites (North Tyrol, Austria) including a description of Mallomonas pechlaneri sp. nov. Cryptogam., Algol. 2018, 39, 63–83. [Google Scholar] [CrossRef]
- Gusev, E.S.; Čertnerová, D.; Škaloudová, M.; Škaloud, P. Exploring cryptic diversity and distribution patterns in the Mallomonas kalinae/rasilis species complex with a description of a new taxon — Mallomonas furtiva sp. nov. J. Euk. Microbiol. 2018, 65, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Hansen, P.; Kristiansen, J. Mallomonas madagascariensis, M. lemuriocellata and M. crocodilorum (Synurophyceae), three new species from Madagascar. Nord. J. Bot. 1995, 15, 215–223. [Google Scholar] [CrossRef]
- Cronberg, G. Scaled chrysophytes from the tropics. Nova Hedw. Beih. 1989, 95, 191–232. [Google Scholar]
- Neustupa, J.; Řezáčová, M. The genus Mallomonas (Mallomonadales, Synurophyceae) in several Southeast Asian urban water bodies — the biogeographic implications. Nova Hedw. 2007, 84, 249–259. [Google Scholar] [CrossRef]
- Němcová, Y.; Kreidlová, J.; Kosová, A.; Neustupa, J. Lakes and pools of Aquitaine region (France)—A biodiversity hotspot of Synurales in Europe. Nova Hedw. 2012, 95, 1–24. [Google Scholar] [CrossRef]
- Siver, P.A.; Lott, A.M. Fossil species of Mallomonas from an Eocene Maar Lake with recessed dome structures: Early attempts securing bristles to the cell covering? Nova Hedw. 2012, 95, 517–529. [Google Scholar] [CrossRef]
- Němcová, Y.; Kreidlová, J.; Pusztai, M.; Neustupa, J. Mallomonas pumilio group (Chrysophyceae/Stramenopiles) – a revision based on the scale/scale-case morphology and analysis of scale shape. Nova Hedw. Beih. 2013, 142, 27–49. [Google Scholar]
- Jo, B.Y.; Kim, J.I.; Škaloud, P.; Siver, P.A.; Shin, W. Multigene phylogeny of Synura (Synurophyceae) and descriptions of four new species based on morphological and DNA evidence. Eur. J. Phycol. 2016, 51, 413–430. [Google Scholar] [CrossRef]
- Kristiansen, J.; Preisig, H.R. Chrysophyte and haptophyte algae. Part 2: Synurophyceae. In Süsswasserflora von Mitteleuropa, 2nd ed.; Büdel, B., Gärtner, G., Krienitz, L., Preisig, H.R., Schagerl, M., Eds.; Spektrum Akademischer Verlag: Berlin–Heidelberg, Germany, 2007; Volume 1, pp. 1–252. [Google Scholar]
- Kapustin, D.A.; Gusev, E.S.; Kulikovskiy, M.S. Mallomonas papuensis sp. nov. (Chrysophyceae, Synurales), a new species from the high mountain bog pool in Papua province, Indonesia. Phytotaxa 2019, 402, 281–287. [Google Scholar] [CrossRef]
- Vyverman, W. A systematic account of the algal flora of the seasonal swamps in the southern part of Irian Jaya (Indonesia). In Recent Trends in Algal Taxonomy; Vidyavati, Mahato, A.K., Eds.; Associated Publishing Co.: New Delhi, India, 2006; Volume 2, pp. 3–19. [Google Scholar]
- Croome, R.L.; Tyler, P.A. Distribution of silica-scaled Chrysophyceae (Paraphysomonadaceae and Mallomonadaceae) in Australian inland waters. Aust. J. Mar. Freshw. Res. 1985, 36, 839–853. [Google Scholar] [CrossRef]
- Vyverman, W.; Cronberg, G. Scale bearing chrysophytes from Papua New Guinea. Nord. J. Bot. 1993, 13, 111–120. [Google Scholar] [CrossRef]
- Kapustin, D.A.; Gusev, E.S.; Lilitskaya, G.G.; Kulikovskiy, M.S. Silica-scaled chrysophytes from the Ukrainian Polissia. Cryptogam., Algol. 2020, 41, 121–135. [Google Scholar] [CrossRef]
- Kristiansen, J.; Lind, J.F. Endemicity in silica-scaled chrysophytes. Nova Hedw. Beih. 2005, 128, 65–83. [Google Scholar]
- Kapustin, D.A.; Gusev, E.S. Silica-scaled chrysophytes from West Java (Indonesia) including description of a new Chrysosphaerella species. Nova Hedw. Beih. 2019, 148, 11–20. [Google Scholar] [CrossRef]
- Siver, P.A.; Lott, A.M. The scaled chrysophyte flora in freshwater ponds and lakes from Newfoundland, Canada, and their relationship to environmental variables. Cryptogam. Algol. 2017, 38, 325–347. [Google Scholar] [CrossRef]
- Zachos, J.C.; Dickens, G.R.; Zeebe, R.E. An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature 2008, 451, 279–283. [Google Scholar] [CrossRef]
- Wolfe, A.P.; Reyes, A.V.; Royer, D.L.; Greenwood, D.R.; Doria, G.; Gagen, M.H.; Siver, P.A.; Westgate, J.A. Middle Eocene CO2 and climate reconstructed from the sediment fill of a subarctic kimberlite maar. Geology 2017, 45, 619–622. [Google Scholar] [CrossRef]
- Siver, P.A.; Wolfe, A.P. Tropical ochrophyte algae from the Eocene of Northern Canada: A biogeographic response to past global warming. Palaios 2009, 24, 192–198. [Google Scholar] [CrossRef] [Green Version]
- Gusev, E.; Kezlya, E.; Tran, H.; Kulikovskiy, M. Mallomonas vietnamica sp. nov. (Synurales, Chrysophyceae), a new species, that shares some features with fossil taxa. Cryptogam. Algol. 2021, 42, 39–46. [Google Scholar] [CrossRef]
- Olefeld, J.L.; Bock, C.; Jensen, M.; Vogt, J.C.; Sieber, G.; Albach, D.; Boenigk, J. Centers of endemism of freshwater protists deviate from pattern of taxon richness on a continental scale. Sci. Rep. 2020, 10, 14431. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality Name | Sample Type | Date | Coordinates | Elevation, m a.s.l. | T, °C | pH | SC, µS·cm−1 | |
|---|---|---|---|---|---|---|---|---|
| 1 | Lake Sentani | Plankton | 14 September 2015 | 2°38′23.7″ S 140°34′04.3″ E | 81 | 29.7 | 7.6 | 230 |
| 2 | Lake Sentani | Periphyton | 14 September 2015 | 2°38′23.7″ S 140°34′04.3″ E | 81 | 30 | 7.6 | 240 |
| 3 | Water body along the road 1 | Plankton | 14 September 2015 | 2°39′54.72″ S 140°32′51.47″ E | 119 | n/a | n/a | n/a |
| 4 | Lake Habbema | Plankton | 15 September 2015 | 4°07′49.5″ S 138°40′22.9″ E | 3331 | 17 | 7.33 | 22 |
| 5 | Lake Habbema | Squeeze from moss | 15 September 2015 | 4°07′49.5″ S 138°40′22.9″ E | 3331 | 17 | 7.33 | n/a |
| 6 | Peat-bog pool | Plankton | 15 September 2015 | 4°7′40.02″ S 138°40′3.90″ E | 3333 | 16.7 | 4.75 | 8 |
| 7 | Stream in a bog | Plankton | 15 September 2015 | 4°7′36.30″ S 138°39′55.56″ E | 3327 | n/a | 4.78 | n/a |
| 8 | Puddle near the stream | Plankton | 15 September 2015 | 4°7′34.68″ S 138°39′53.34″ E | 3330 | n/a | n/a | n/a |
| 9 | Peat-bog pool | Benthos | 15 September 2015 | 4°7′36.60″ S 138°39′53.58″ E | 3328 | n/a | n/a | n/a |
| 10 | Small puddle in a bog | Plankton | 15 September 2015 | 4°7′30.96″ S 138°39′53.88″ E | 3337 | n/a | n/a | n/a |
| 11 | Lake Anagera | Plankton | 16 September 2015 | 3°57′07.4″ S 138°52′27.6″ E | 1656 | 25 | 7.3 | 229 |
| 12 | Water body along the road 2 | Plankton | 16 September 2015 | 3°54′33.4″ S 138°50′15.1″ E | 1661 | 25 | 7.06 | 234 |
| 13 | Small ephemeral pond | Plankton | 16 September 2015 | 3°54′09.9″ S 138°50′17.1″ E | 1730 | 27 | 6.99 | 162 |
| 14 | Water body along the road 3 | Plankton | 16 September 2015 | 3°54′49.6″ S 138°51′27.4″ E | 1654 | 30 | 7.05 | 47 |
| Taxon | Site Number | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |
| Paraphysomonadales | ||||||||||||||
| Paraphysomonas cf. segmenta Scoble and Cavalier-Smith | × | |||||||||||||
| Paraphysomonas sp. | × | |||||||||||||
| Paraphysomonas uniformis Scoble and Cavalier-Smith | × | |||||||||||||
| Paraphysomonas vulgaris subsp. vulgaris Scoble and Cavalier-Smith | × | |||||||||||||
| Paraphysomonas vulgaris subsp. brevispina Scoble and Cavalier-Smith | × | × | ||||||||||||
| Chromulinales | ||||||||||||||
| Chrysosphaerella annulata Kristiansen and Tong | × | × | ||||||||||||
| Spiniferomonas sp. | × | × | ||||||||||||
| Synurales | ||||||||||||||
| Mallomonas cratis K. Harris and D.E. Bradley | × | |||||||||||||
| Mallomonas cyathellata Wujek and Asmund | × | × | ||||||||||||
| Mallomonas cf. elongata Reverdin | × | |||||||||||||
| Mallomonas furtiva Gusev, Čertnerová, Škaloudová and Škaloud | × | |||||||||||||
| Mallomonas mangofera var. foveata (Dürrschmidt) Kristiansen | × | × | ||||||||||||
| Mallomonas cf. mangofera var. reticulata (Cronberg) Kristiansen | × | × | × | |||||||||||
| Mallomonas multisetigera Dürrschmidt | × | × | × | |||||||||||
| Mallomonas papillosa K. Harris and D.E. Bradley emend. K. Harris | × | |||||||||||||
| Mallomonas papuensis Kapustin, Gusev and Kulikovskiy | × | × | ||||||||||||
| Mallomonas cf. portae-ferreae L.S. Péterfi and Asmund | × | × | ||||||||||||
| Mallomonas preisigii Siver | × | × | ||||||||||||
| Mallomonas solea-ferrea var. irregularis Němcová, Kreidlova, Pusztai and Neustupa | × | |||||||||||||
| Mallomonas striata var. serrata K. Harris and D.E. Bradley | × | × | × | |||||||||||
| Mallomonas sp. 1 | × | × | ||||||||||||
| Mallomonas sp. 2 | × | × | × | |||||||||||
| Synura cf. longitubularis Jo, Shin, Kim and Siver | × | × | × | |||||||||||
| Synura mammillosa E. Takahashi | × | × | × | × | × | × | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gusev, E.; Kapustin, D.; Martynenko, N.; Kulikovskiy, M. Diversity of Silica-Scaled Chrysophytes (Stramenopiles: Chrysophyceae) from Indonesian Papua. Diversity 2022, 14, 726. https://doi.org/10.3390/d14090726
Gusev E, Kapustin D, Martynenko N, Kulikovskiy M. Diversity of Silica-Scaled Chrysophytes (Stramenopiles: Chrysophyceae) from Indonesian Papua. Diversity. 2022; 14(9):726. https://doi.org/10.3390/d14090726
Chicago/Turabian StyleGusev, Evgeniy, Dmitry Kapustin, Nikita Martynenko, and Maxim Kulikovskiy. 2022. "Diversity of Silica-Scaled Chrysophytes (Stramenopiles: Chrysophyceae) from Indonesian Papua" Diversity 14, no. 9: 726. https://doi.org/10.3390/d14090726