
Geographical Patterns in Functional Diversity of Chinese Terrestrial Vertebrates
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Diversity Metric Calculation and Hotspots Identification
3. Results
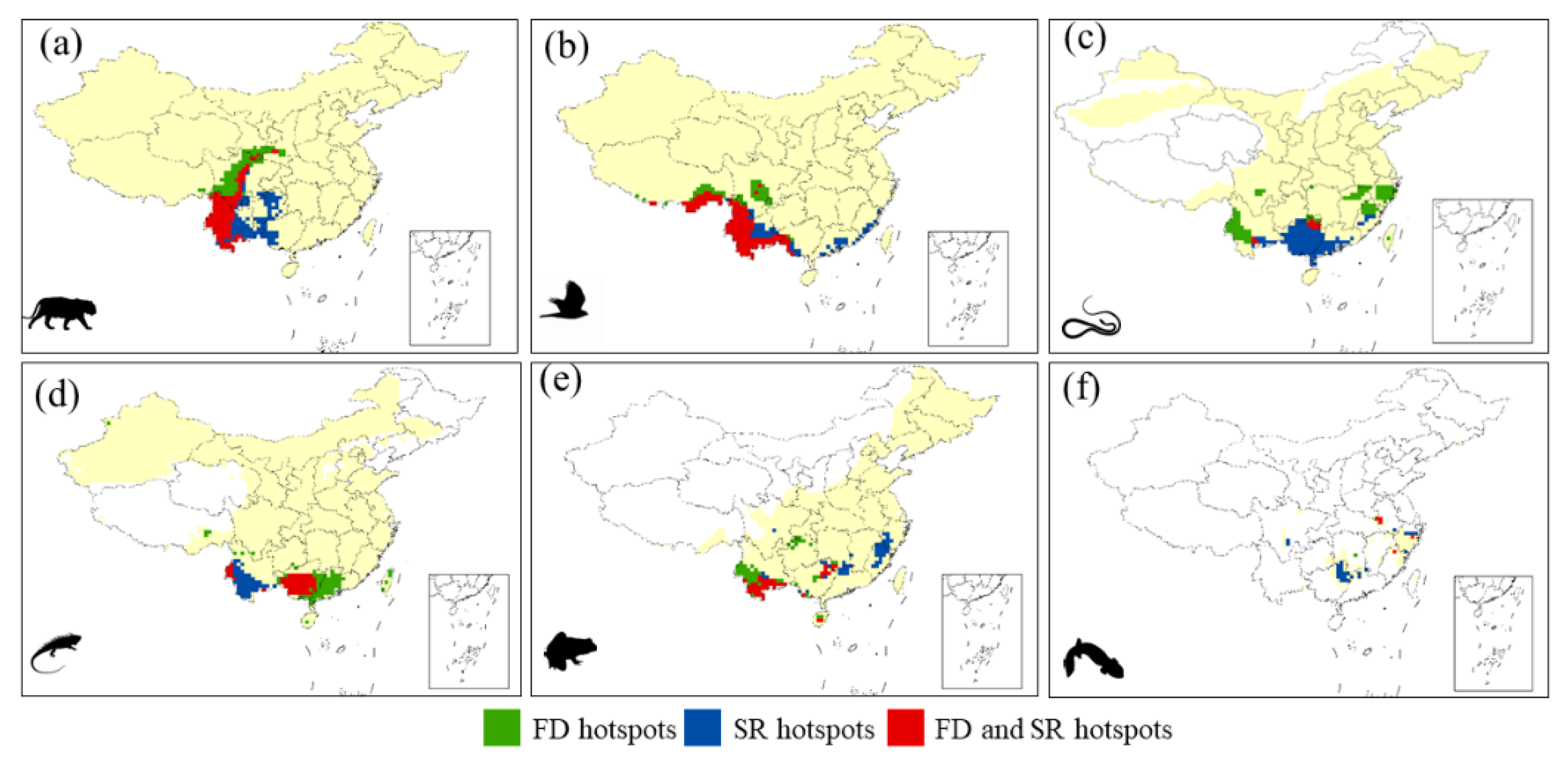
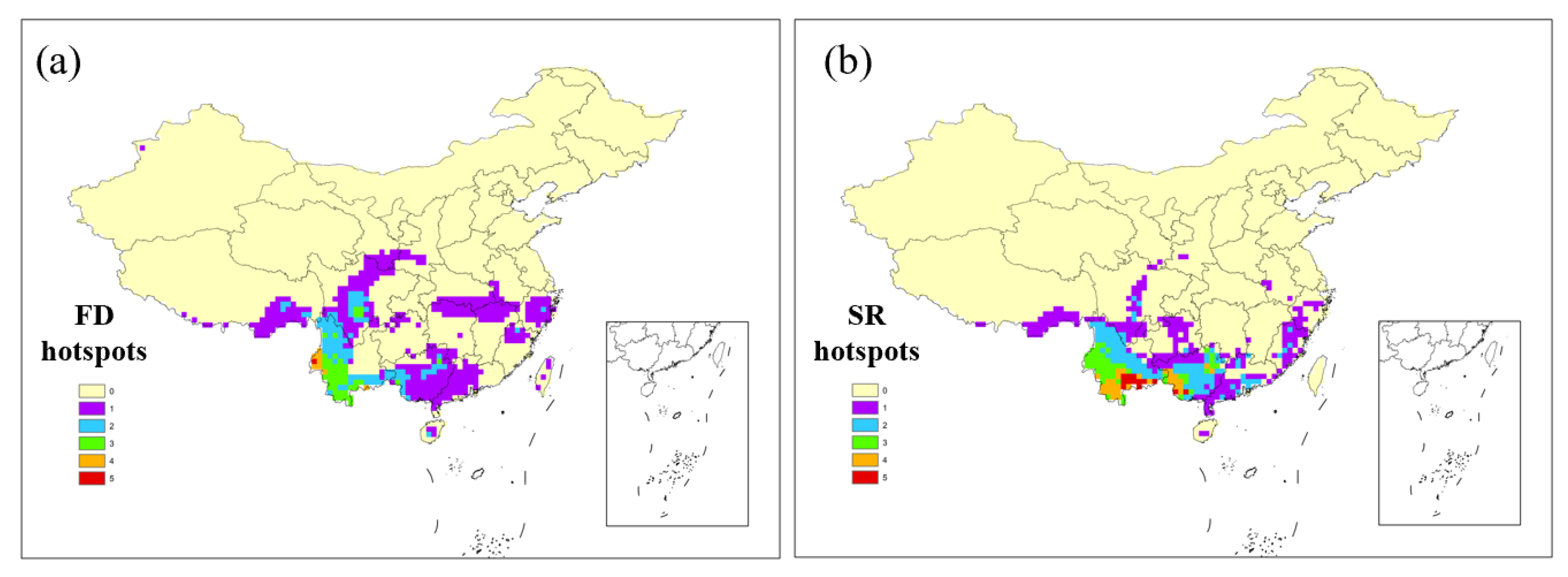
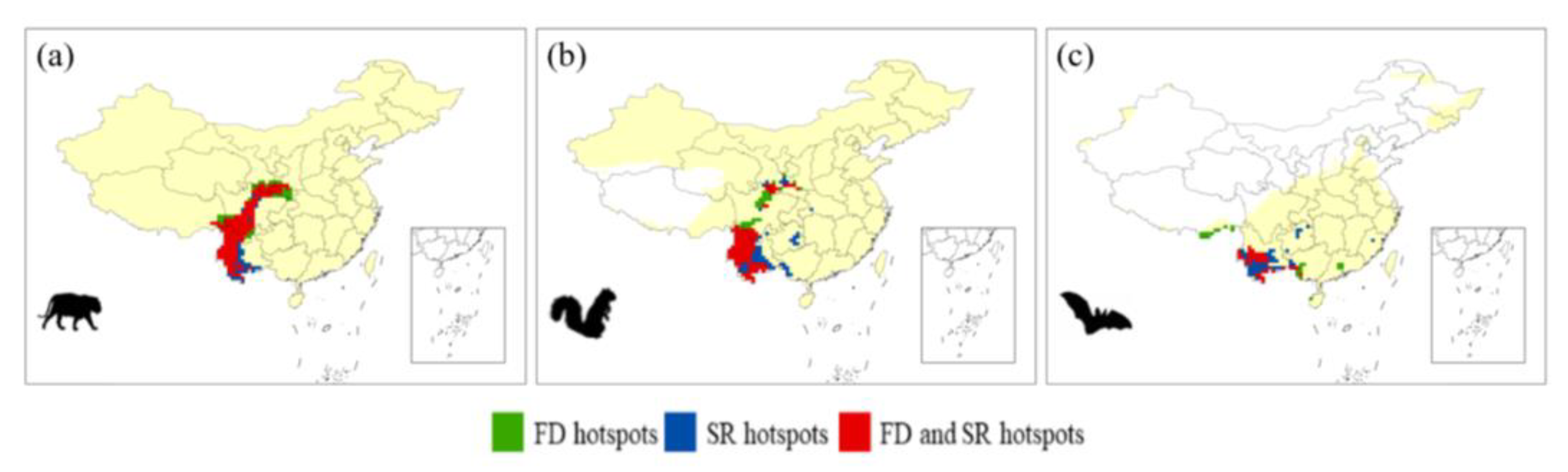
3.1. Distribution Patterns of the Hotspots
3.2. Differences between FD and SR Hotspots
4. Discussion
4.1. The Spatial Pattern of Functional Hotspots and Potential Causes
4.2. The Differences between FD Hotspots and SR Hotspots
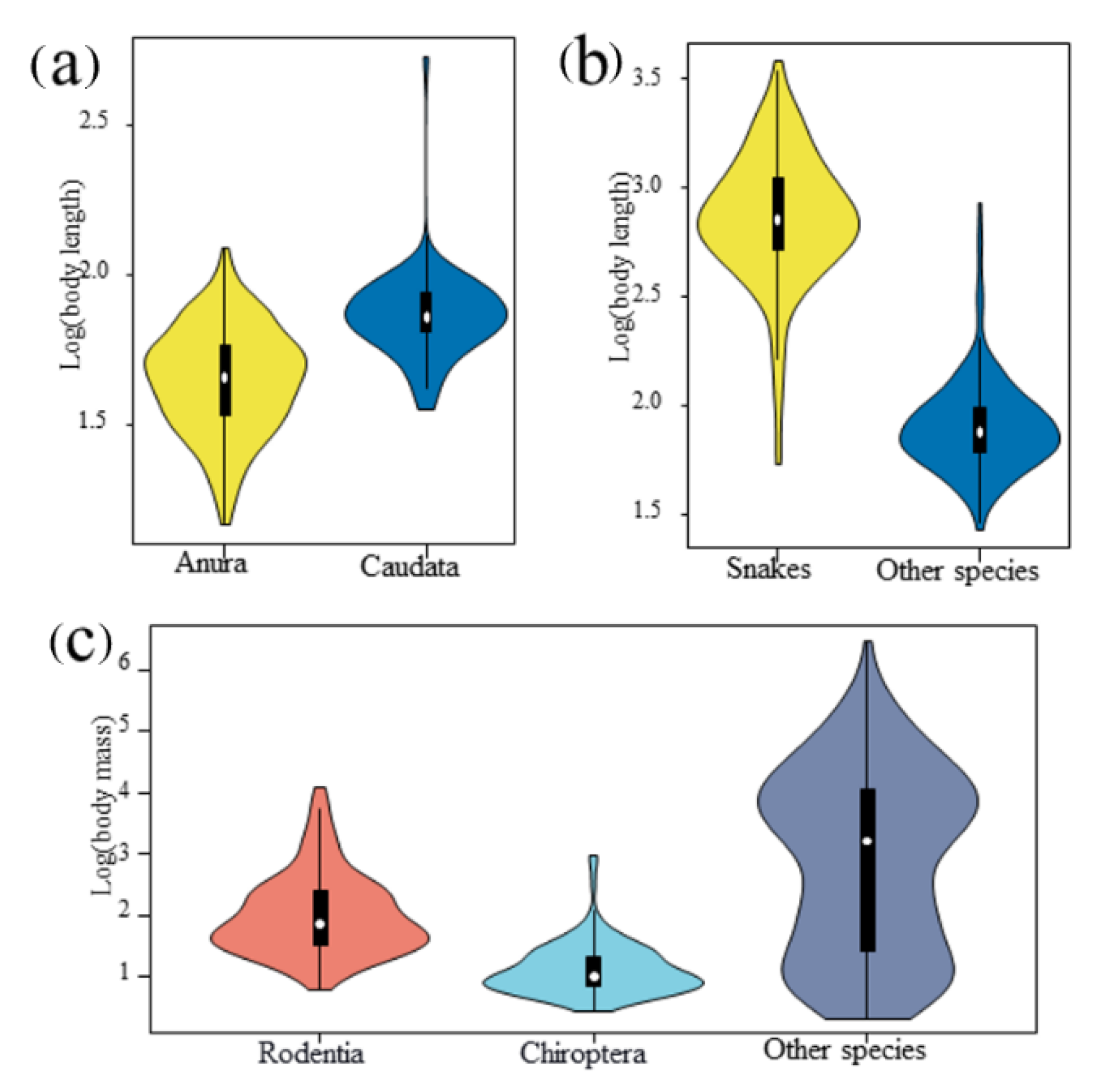
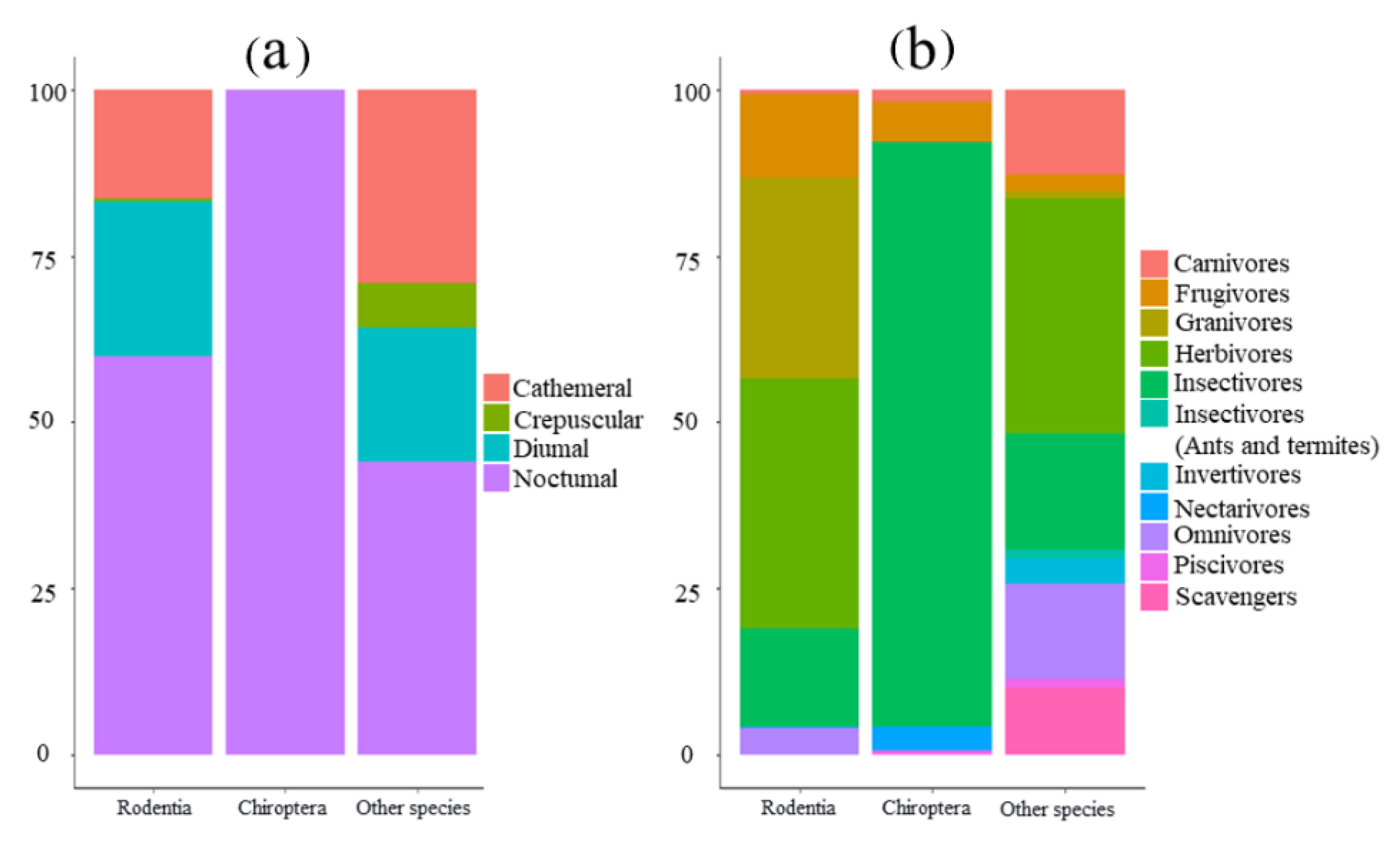
4.3. Distinct Patterns within a Taxonomic Unit
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Balmford, A.; Green, R.E.; Jenkins, M. Measuring the changing state of nature. Trends Ecol. Evol. 2003, 18, 326–330. [Google Scholar] [CrossRef]
- Cowie, R.H.; Bouchet, P.; Fontaine, B. The Sixth Mass Extinction: Fact, fiction or speculation? Biol. Rev. 2022, 97, 640–663. [Google Scholar] [CrossRef] [PubMed]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 1246752. [Google Scholar] [CrossRef] [PubMed]
- Steffen, W.; Richardson, K.; Rockström, J.; Cornell, S.E.; Fetzer, I.; Bennett, E.M.; Biggs, R.; Carpenter, S.R.; De Vries, W.; De Wit, C.A. Planetary boundaries: Guiding human development on a changing planet. Science 2015, 347, 1259855. [Google Scholar] [CrossRef] [Green Version]
- Mittermeier, R.A.; Gil, P.R.; Mittermeier, C.G. Megadiversity: Earth’s Biologically Wealthiest Nations; Agrupación Sierra Madre: Mexico City, Mexico, 1997. [Google Scholar]
- Ji, L. China Checklist of Animals; the Biodiversity Committee of Chinese Academy of Sciences, Ed.; Catalogue of Life China: 2022 Annual Checklist; Beijing, China, 2022. Available online: http://www.sp2000.org.cn/info (accessed on 1 May 2022).
- Jiang, Z.; Jiang, J.; Wang, Y.; Zhang, E.; Zhang, Y.; Li, L.; Xie, F.; Cai, B.; Cao, L.; Zheng, G. Red List of China’s Vertebrates. Biodivers. Sci. 2016, 24, 500–551. [Google Scholar]
- Jiang, J.; Xie, F.; Zang, C.; Cai, L.; Li, C.; Wang, B.; Li, J.; Wang, J.; Hu, J.; Wang, Y. Assessing the threat status of amphibians in China. Biodivers. Sci. 2016, 24, 588–597. [Google Scholar] [CrossRef]
- Xie, F.; Lau, M.W.N.; Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Fischman, D.L. Conservation needs of amphibians in China: A review. Sci. China Ser. C Life Sci. 2007, 50, 265–276. [Google Scholar] [CrossRef]
- Cao, L.; Zhang, E.; Zang, C.; Cao, W. Evaluating the status of China’s continental fish and analyzing their causes of endangerment through the red list assessment. Biodivers. Sci. 2016, 24, 598–609. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Huang, J.; Liu, C.; Zhang, J.; Lu, X.; Ma, K. Diversity hotspots and conservation gaps for the Chinese endemic seed flora. Biol. Conserv. 2016, 198, 104–112. [Google Scholar] [CrossRef]
- Marchese, C. Biodiversity hotspots: A shortcut for a more complicated concept. Glob. Ecol. Conserv. 2015, 3, 297–309. [Google Scholar] [CrossRef] [Green Version]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, C.N.; Pimm, S.L.; Joppa, L.N. Global patterns of terrestrial vertebrate diversity and conservation. Proc. Natl. Acad. Sci. USA 2013, 110, E2602–E2610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, H.; Lyu, L.; Shrestha, N.; Tang, Z.; Su, X.; Xu, X.; Dimitrov, D.; Wang, Z. Geographical patterns in phylogenetic diversity of Chinese woody plants and its application for conservation planning. Divers. Distrib. 2021, 27, 179–194. [Google Scholar] [CrossRef]
- Tang, Z.; Wang, Z.; Zheng, C.; Fang, J. Biodiversity in China’s mountains. Front. Ecol. Environ. 2006, 4, 347–352. [Google Scholar] [CrossRef]
- Chen, Y.; Bi, J. Biogeography and hotspots of amphibian species of China: Implications to reserve selection and conservation. Curr. Sci. 2007, 92, 480–489. [Google Scholar]
- Li, G.; Xiao, N.; Luo, Z.; Liu, D.; Zhao, Z.; Guan, X.; Zang, C.; Li, J.; Shen, Z. Identifying conservation priority areas for gymnosperm species under climate changes in China. Biol. Conserv. 2021, 253, 108914. [Google Scholar] [CrossRef]
- Xu, Y.; Huang, J.; Lu, X.; Ding, Y.; Zang, R. Priorities and conservation gaps across three biodiversity dimensions of rare and endangered plant species in China. Biol. Conserv. 2019, 229, 30–37. [Google Scholar] [CrossRef]
- Shrestha, N.; Wang, Z. Selecting priority areas for systematic conservation of Chinese Rhododendron: Hotspot versus complementarity approaches. Biodivers. Conserv. 2018, 27, 3759–3775. [Google Scholar] [CrossRef]
- Zhao, L.; Li, J.; Liu, H.; Qin, H. Distribution, congruence and hotspots of higher plants in China. Sci. Rep. 2016, 6, 19080. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Wu, Y.; Cao, Y.; Cao, M.; Tong, W.; Le, Z.; Lu, X.; Li, J.; Ma, F.; Liu, L. Low overlaps between hotspots and complementary sets of vertebrate and plant species in China. Biodivers. Conserv. 2018, 27, 2713–2727. [Google Scholar] [CrossRef]
- Wang, K.; Ren, J.; Chen, H.; Lyu, Z.-T.; Guo, X.; Jiang, K.; Chen, J.; Li, J.; Guo, P.; Wang, Y.-Y.; et al. The updated checklists of amphibians and reptiles of China. Biodivers. Sci. 2020, 28, 189–218. [Google Scholar]
- Chao, A.; Colwell, R.K. Biodiversity: Concepts, Dimensions, and Measures. In The Ecological and Societal Consequences of Biodiversity Loss; Wiley: Hoboken, NJ, USA, 2022; pp. 25–46. [Google Scholar]
- Cadotte, M.W.; Carscadden, K.; Mirotchnick, N. Beyond species: Functional diversity and the maintenance of ecological processes and services. J. Appl. Ecol. 2011, 48, 1079–1087. [Google Scholar] [CrossRef]
- Díaz, S.; Cabido, M. Vive la différence: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Owen, N.R.; Gumbs, R.; Gray, C.L.; Faith, D.P. Global conservation of phylogenetic diversity captures more than just functional diversity. Nat. Commun. 2019, 10, 859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devictor, V.; Mouillot, D.; Meynard, C.; Jiguet, F.; Thuiller, W.; Mouquet, N. Spatial mismatch and congruence between taxonomic, phylogenetic and functional diversity: The need for integrative conservation strategies in a changing world. Ecol. Lett. 2010, 13, 1030–1040. [Google Scholar] [CrossRef]
- Kuczynski, L.; Côte, J.; Toussaint, A.; Brosse, S.; Buisson, L.; Grenouillet, G. Spatial mismatch in morphological, ecological and phylogenetic diversity, in historical and contemporary European freshwater fish faunas. Ecography 2018, 41, 1665–1674. [Google Scholar] [CrossRef] [Green Version]
- Mazel, F.; Pennell, M.W.; Cadotte, M.W.; Diaz, S.; Dalla Riva, G.V.; Grenyer, R.; Leprieur, F.; Mooers, A.O.; Mouillot, D.; Tucker, C.M. Prioritizing phylogenetic diversity captures functional diversity unreliably. Nat. Commun. 2018, 9, 2888. [Google Scholar] [CrossRef] [Green Version]
- Mazel, F.; Mooers, A.O.; Riva, G.V.D.; Pennell, M.W. Conserving phylogenetic diversity can be a poor strategy for conserving functional diversity. Syst. Biol. 2017, 66, 1019–1027. [Google Scholar] [CrossRef] [Green Version]
- IUCN. IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2019. [Google Scholar]
- BirdLife International. Handbook of the Birds of the World, 2018. Bird Species Distribution Maps of the World. Version 2018.1. 2018. Available online: http://datazone.birdlife.org/species/requestdis (accessed on 1 January 2018).
- Roll, U.; Feldman, A.; Novosolov, M.; Allison, A.; Bauer, A.M.; Bernard, R.; Böhm, M.; Castro-Herrera, F.; Chirio, L.; Collen, B. The global distribution of tetrapods reveals a need for targeted reptile conservation. Nat. Ecol. Evol. 2017, 1, 1677–1682. [Google Scholar] [CrossRef]
- Vilela, B.; Villalobos, F. letsR: A new R package for data handling and analysis in macroecology. Methods Ecol. Evol. 2015, 6, 1229–1234. [Google Scholar] [CrossRef]
- Ding, C.; Liang, D.; Xin, W.; Li, C.; Ameca, E.; Jiang, Z. A dataset on the morphological, life-history and ecological traits of the mammals in China. Biodivers. Sci. 2022, 20, 1–10. [Google Scholar] [CrossRef]
- Wang, Y.; Song, Y.; Zhong, Y.; Chen, C.; Zhao, Y.; Zeng, D.; Wu, Y.; Ding, P. A dataset on the life-history and ecological traits of Chinese birds. Biodivers. Sci. 2021, 29, 1149–1153. [Google Scholar] [CrossRef]
- Pyron, R.A.; Burbrink, F.T. Early origin of viviparity and multiple reversions to oviparity in squamate reptiles. Ecol. Lett. 2014, 17, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Zhao, E. Snakes of China; Anhui Science and Technology Press: Anhui, China, 2006. [Google Scholar]
- Scharf, I.; Feldman, A.; Novosolov, M.; Pincheira-Donoso, D.; Das, I.; Böhm, M.; Uetz, P.; Torres-Carvajal, O.; Bauer, A.; Roll, U.; et al. Late bloomers and baby boomers: Ecological drivers of longevity in squamates and the tuatara. Glob. Ecol. Biogeogr. 2015, 24, 396–405. [Google Scholar] [CrossRef]
- Feldman, A.; Sabath, N.; Pyron, R.; Mayrose, I.; Meiri, S. Body sizes and diversification rates of lizards, snakes, amphisbaenians and the tuatara. Glob. Ecol. Biogeogr. 2016, 25, 187–197. [Google Scholar] [CrossRef]
- Zhong, Y.; Chen, C.; Wang, Y. A dataset on the life-history and ecological traits of Chinese lizards. Biodivers. Sci. 2022, 30, 22071. [Google Scholar] [CrossRef]
- Song, Y.; Chen, C.; Wang, Y. A dataset on the life-history and ecological traits of Chinese amphibians. Biodivers. Sci. 2022, 30, 22053. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Schwilk, D.W.; Ackerly, D.D. A trait-based test for habitat filtering: Convex hull volume. Ecology 2006, 87, 1465–1471. [Google Scholar] [CrossRef]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [Green Version]
- Mouchet, M.A.; Villéger, S.; Mason, N.W.; Mouillot, D. Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules. Funct. Ecol. 2010, 24, 867–876. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B.; Laliberté, M. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology; R package version 1.0-12. Available online: https://rdrr.io/cran/FD/man/FD-package.html (accessed on 19 August 2014).
- Shrestha, N.; Xu, X.; Meng, J.; Wang, Z. Vulnerabilities of protected lands in the face of climate and human footprint changes. Nat. Commun. 2021, 12, 1632. [Google Scholar] [CrossRef] [PubMed]
- Fei, L.; Ye, C.; Huang, Y. Key to Chinese Amphibians; Publishing House for Scientific and Technological Literature: Chongqing, China, 1990. [Google Scholar]
- Tejero-Cicuéndez, H.; Tarroso, P.; Carranza, S.; Rabosky, D. Desert lizard diversity worldwide: Effects of environment, time, and evolutionary rate. Glob. Ecol. Biogeogr. 2022, 31, 776–790. [Google Scholar] [CrossRef]
- Schoener, T.W. The Anolis lizards of Bimini: Resource partitioning in a complex fauna. Ecology 1968, 49, 704–726. [Google Scholar] [CrossRef]
- Van der Vaart, A.W. Asymptotic Statistics; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Warren, D.; Glor, R.; Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evolution 2008, 62, 2868–2883. [Google Scholar] [CrossRef]
- Polato, N.R.; Gill, B.A.; Shah, A.A.; Gray, M.M.; Casner, K.L.; Barthelet, A.; Messer, P.W.; Simmons, M.P.; Guayasamin, J.M.; Encalada, A.C.; et al. Narrow thermal tolerance and low dispersal drive higher speciation in tropical mountains. Proc. Natl. Acad. Sci. USA 2018, 115, 12471–12476. [Google Scholar] [CrossRef] [Green Version]
- He, K.; Jiang, X. Sky islands of southwest China. I: An overview of phylogeographic patterns. Chin. Sci. Bull. 2014, 59, 585–597. [Google Scholar] [CrossRef]
- Li, H.; Li, R. Plant Resources and Geography of the Gaoligong Mountains in Southeast Tibet; Hubei Science and Technology Press: Wuhan, China, 2021. [Google Scholar]
- Fei, X.; Jin, Y.; Zhang, Y.; Sha, L.; Liu, Y.; Song, Q.; Zhou, W.; Liang, N.; Yu, G.; Zhang, L. Eddy covariance and biometric measurements show that a savanna ecosystem in Southwest China is a carbon sink. Sci. Rep. 2017, 7, 41025. [Google Scholar] [CrossRef] [Green Version]
- Rahbek, C.; Borregaard, M.; Antonelli, A.; Colwell, R.; Holt, B.; Nogués-Bravo, D.; Rasmussen, C.; Richardson, K.; Rosing, M.; Whittaker, R.; et al. Building mountain biodiversity: Geological and evolutionary processes. Science 2019, 365, 1114–1119. [Google Scholar] [CrossRef]
- Zhang, D.R.; Hui, H.; Yu, G.H.; Song, X.Q.; Liu, S.; Yuan, S.Q.; Xiao, H.; Rao, D.Q. Shared response to changes in drainage basin: Phylogeography of the Yunnan small narrow-mouthed frog, Glyphoglossus yunnanensis (Anura: Microhylidae). Ecol. Evol. 2020, 10, 1567–1580. [Google Scholar] [CrossRef] [Green Version]
- Holt, B.G.; Lessard, J.-P.; Borregaard, M.K.; Fritz, S.A.; Araújo, M.B.; Dimitrov, D.; Fabre, P.-H.; Graham, C.H.; Graves, G.R.; Jønsson, K.A. An update of Wallace’s Zoogeographic regions of the world. Science 2013, 339, 74–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filardi, C.E.; Moyle, R.G. Single origin of a pan-Pacific bird group and upstream colonization of Australasia. Nature 2005, 438, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Jønsson, K.A.; Fabre, P.-H.; Ricklefs, R.E.; Fjeldså, J. Major global radiation of corvoid birds originated in the proto-Papuan archipelago. Proc. Natl. Acad. Sci. USA 2011, 108, 2328–12333. [Google Scholar] [CrossRef]
- Jiang, Z. China’s Mammal Diversity and Geographic Distribution; Science Press: Beijing, China, 2015. [Google Scholar]
- Pollock, L.J.; Thuiller, W.; Jetz, W. Large conservation gains possible for global biodiversity facets. Nature 2017, 546, 141–144. [Google Scholar] [CrossRef]
- Pough, F.; Andrews, R.; Crump, M.; Savitzky, A.; Wells, K.; Brandley, M. Herpetology; Sinauer Associates: Sunderland, MA, USA, 2016. [Google Scholar]
- Amori, G.; Spartaco, G.; Luiselli, L. Do biodiversity hotspots match with rodent conservation hotspots? Biodivers. Conserv. 2011, 20, 3693–3700. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Schoener’s D | Hellinger’s Distance |
|---|---|---|
| Anura | 0.535 | 0.692 |
| Birds | 0.348 | 0.813 |
| Caudata | 0.606 | 0.46 |
| Lizards | 0.714 | 0.479 |
| Snakes | 0.731 | 0.425 |
| Mammals | 0.343 | 0.907 |
| Rodentia | 0.649 | 0.516 |
| Chiroptera | 0.804 | 0.274 |
| Mammalian speceis besides Rodentia and Chiroptera | 0.752 | 0.307 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Huang, N.; Zhou, W. Geographical Patterns in Functional Diversity of Chinese Terrestrial Vertebrates. Diversity 2022, 14, 987. https://doi.org/10.3390/d14110987
Sun X, Huang N, Zhou W. Geographical Patterns in Functional Diversity of Chinese Terrestrial Vertebrates. Diversity. 2022; 14(11):987. https://doi.org/10.3390/d14110987
Chicago/Turabian StyleSun, Xinyuan, Na Huang, and Weiwei Zhou. 2022. "Geographical Patterns in Functional Diversity of Chinese Terrestrial Vertebrates" Diversity 14, no. 11: 987. https://doi.org/10.3390/d14110987