3.1. Six Fish Species Examined were Positive with 10 Gill Monogenoidean Species (7 New) from the Cayo Arcas Reef, Gulf of Mexico
The sergeant-major
Abudefduf saxatilis (Linnaeus, 1758) harbored the highest number of monogenoidean species (5 spp.) followed by the beaugregory
Stegastes xanthurus (Poey, 1860) (with 3 monogenoidean species) and bicolor damselfish,
Stegastes partitus (Poey, 1868), beaugregory
Stegastes leucostictus (Müller & Troschel, 1848), brown chromis,
Chromis multilineata (Guichenot, 1853) (all Pomacentridae), and the spotfin butterflyfish
Chaetodon ocellatus Bloch 1787 (Chaetodontidae) with 1 monogenoidean species each (
Table 1).
3.1.1. Diagnosis of Paraeuryhaliotrema n. gen.
Class Monogenoidea Bychowsky, 1937
Subclass Polyonchoinea Bychowsky, 1937
Order Dactylogyridea Bychowsky, 1937
Dactylogyridae Bychowsky, 1933
The body comprising a body proper (cephalic region, trunk, short peduncle), haptor. Tegument smooth. Cephalic margin rounded. Head organs, cephalic lobes present. Eyespots present. Mouth subterminal, midventral, prepharyngeal; pharynx made up of muscular, glandular bulb; esophagus, intestinal ceca 2, confluent posterior to gonads, lacking diverticula. Common genital pore midventral, immediately posterior to intestinal bifurcation. Gonads intercaecal, overlapping; testis dorsal to germarium. Seminal vesicle a simple dilation of vas deferens; prostatic reservoir present. Copulatory complex composed of a male copulatory organ (MCO), accessory piece. MCO tubular, sclerotized, comprising a bulbous base from which arises coiled shaft in clockwise direction. Seminal receptacle not observed; vaginal aperture dextroventral. Vitellaria in trunk, peduncle, absent in regions of other reproductive organs. Haptor bilobed, armed with dorsal, ventral anchor-bar complexes, seven pairs of similar hooks, each with upright thumb, shank comprising one subunit. Type species: Paraeuryhaliotrema pomacentris n. gen., n. sp. from the gills of Stegastes xanthurus (Pomacentridae).
Taxonomic summary:—Type species: Paraeuryhaliotrema pomacentris n. sp. from the gills of Stegastes xanthurus (Poey, 1860), (Perciformes: Pomacentridae).
Etymology:—The genus name refers to the resemblance of the new taxon to Euryhaliotrema Kritsky and Boeger 2002, whose members are described and/or reported on the gills of snappers (Lutjanidae), drums and croakers (Sciaenidae), porgies (Sparidae) and grunts (Haemulidae) from the Gulf of Mexico and the eastern Pacific, Indowest Pacific and eastern Atlantic Ocean.
Remarks:—Previous investigations of monogenoids parasitizing pomacentrids from the Gulf of Mexico and Cuban waters have revealed the occurrence of species of
Neohaliotrema (Yamaguti 1965) Lim and Gibson 2010,
Paraneohaliotrema Zhukov 1976 and
Microcotyle Beneden and Hesse, 1863 (see [
1,
16,
17]). Species of these three genera are differentiated from
Paraeuryhaliotrema n. gen., n. sp. in having a coiled MCO (curved tube in
Neohaliotrema and
Paraneohaliotrema) and haptor with ventral anchor-bar complexes (haptor armed with clamps in
Microcotyle).
Paraeuryhaliotrema n. gen., n. sp. most resembles species of
Euryhaliotrema (Kritsky and Boeger 2002) Kritsky 2012, from fish hosts worldwide (including Gulf of Mexico) of the Ambassidae, Chaetodontidae, Caesionidae, Haemulidae, Lutjanidae, Sciaenidae, and Sparidae. Species of both genera share the following morphologic features: a coiled MCO with bulbous-shaped base, an accessory piece without articulation to the base of the MCO (see this latter character in the species-group 3 proposed by Sun and Yang [
18] for
Euryhaliotrema), vaginal aperture dextroventral and hooks, each with upright acute thumb, slender shank comprised of one subunit.
Paraeuryhaliotrema pomacentris n. gen., differs from species of
Euryhaliotrema by having a coiled MCO with rings in clockwise orientation (counterclockwise rings in
Euryhaliotrema), simple accessory piece (multibranched or poorly developed in
Euryhaliotrema), overlapping gonads (tandem or slightly overlapping in
Euryhaliotrema) and by general shape and size of hooks, i.e., 10–12 vs. 14–17 in species of
Euryhaliotrema (see [
6,
18,
19,
20,
21]).
Paraeuryhaliotrema pomacentris n. gen., n. sp. also differs of species of
Haliotrematoides Kritsky, Yang and Sun 2009 (which include features in common with those of
Euryhaliotrema), which have been reported on haemulids, lutjanids and sparids fishes from the Gulf of Mexico, Caribbean and Pacific ocean for Mexican and Cuban waters in lacking hook pairs 1, 2 and 3 on the ventral surface of peduncle, absence of an accessory piece and gonads tandem (present in
Haliotrematoides spp.) (see [
22,
23]).
Paraeuryhaliotrema pomacentris n. sp. is the first described dactylogyrid species on the pomacentrid
S. xanthurus from the Gulf of Mexico.
3.1.2. Description of Paraeuryhaliotrema pomacentris n. sp.
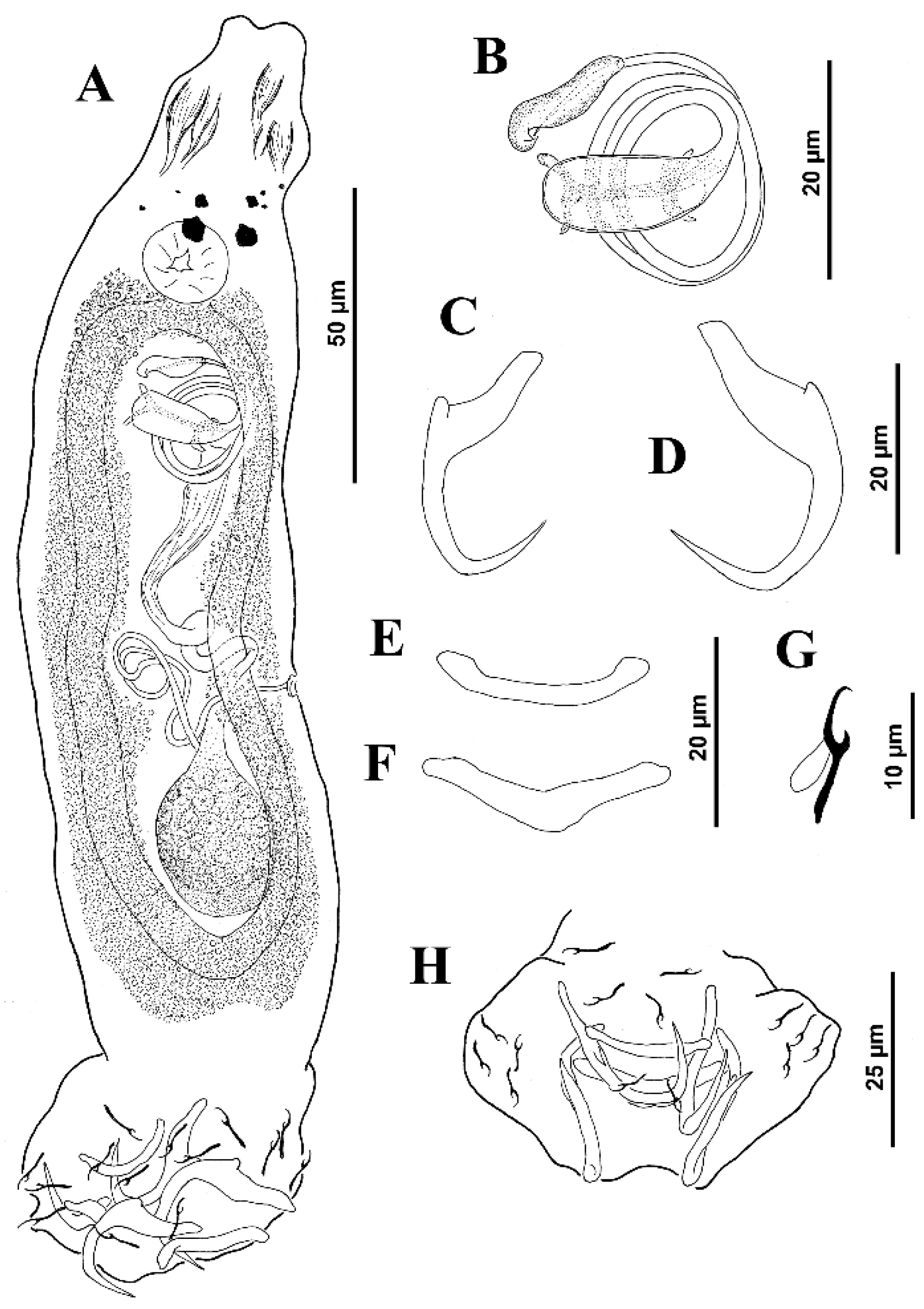
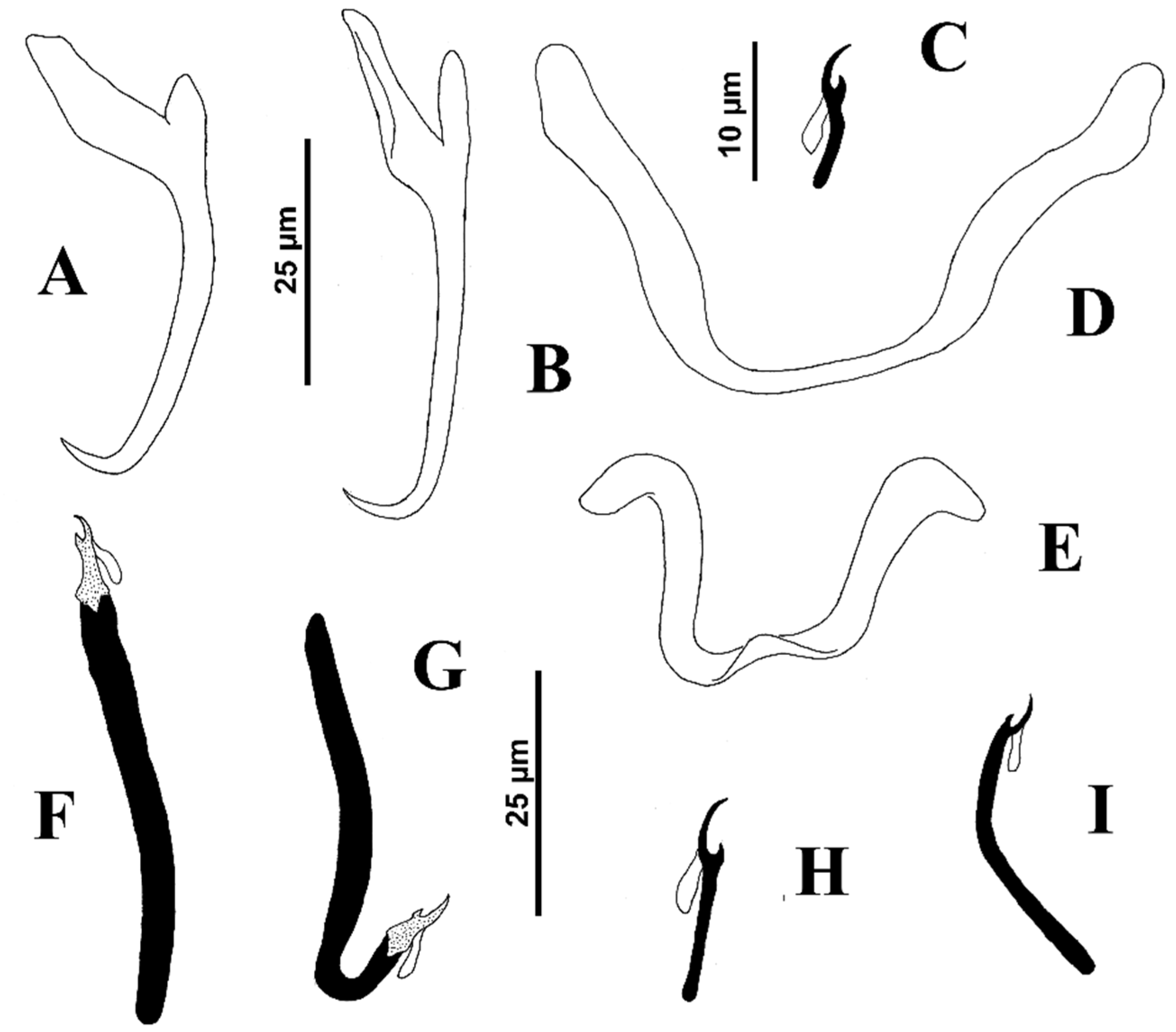
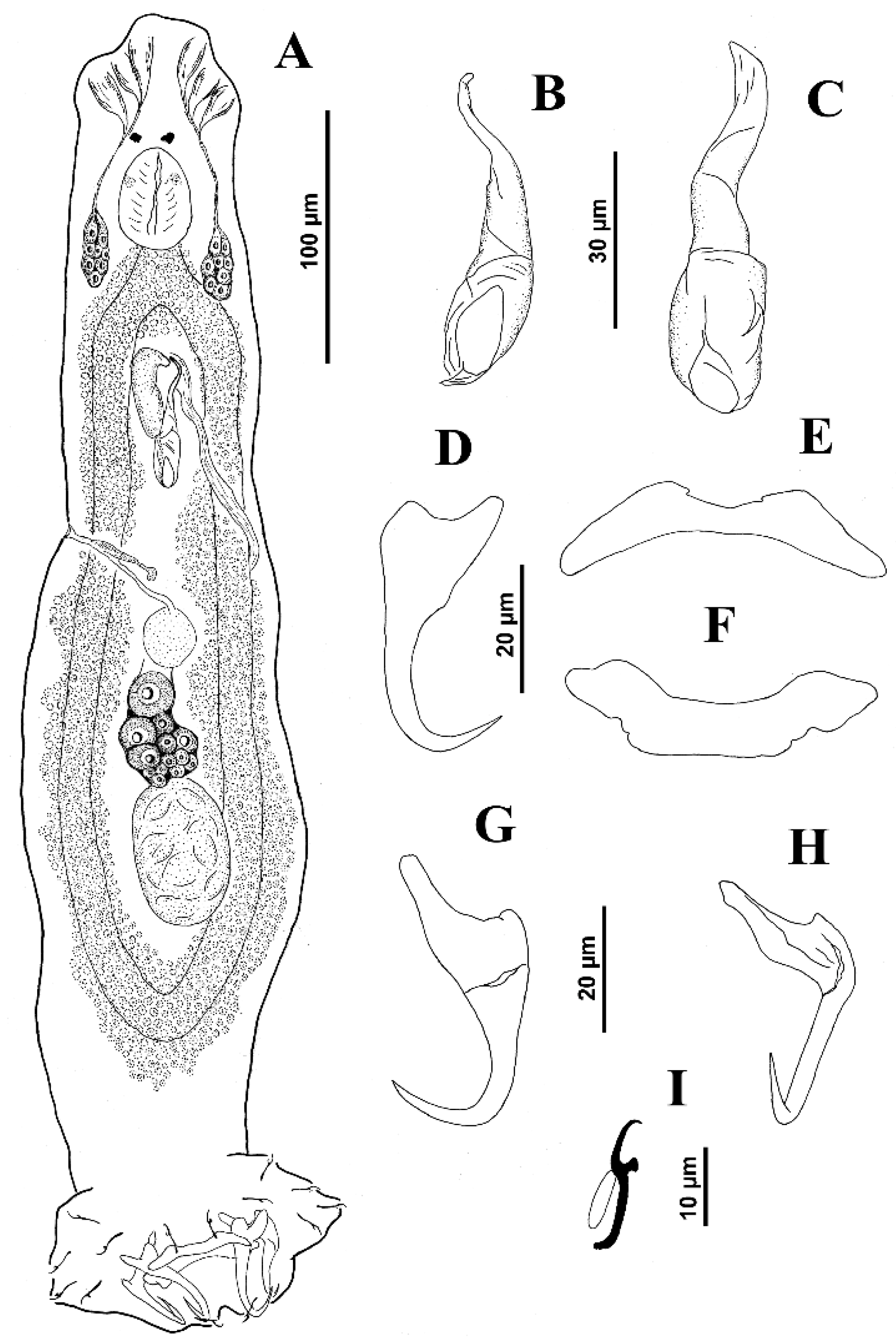
Body proper elongate, fusiform, 253 (217–285;
n = 6) long, with greatest width 48 (38–65;
n = 8) at level of gonads. Cephalic region moderately broad; cephalic lobes poorly differentiated. Three bilateral pairs of head organs poorly defined; cephalic glands not observed. Tegument smooth. Two pairs of eyespots, posterior pair larger than members of anterior pair; accessory chromatic granules small, few in cephalic region. Mouth subterminal, midventral, prepharyngeal. Pharynx subspherical, 15 (13–17;
n = 5) wide; esophagus short to nonexistent; intestinal ceca two, lacking diverticula, confluent posterior to testis. Peduncle broad and short; haptor sub-hexagonal, 55 (50–62;
n = 3) wide. Ventral anchor 24 (23–26;
n = 8) long, with rounded deep root, tapered superficial root, evenly elongate curved shaft and elongate point; base 12 (
n = 5) wide. Dorsal anchor 26 (26–27;
n = 8) long, with poorly developed superficial root, widened superficial root, distally tapered, arcing shaft with elongate point extending to near level of tip of superficial root; base 13 (12–13;
n = 5) wide. Ventral bar 20 (18–20;
n = 6) long, rod-shaped, with ends slightly expanded and directed anteriorly; dorsal bar 24 (21–27;
n = 7) long, V-shaped, with ends directed laterally. Hooks similar; each 11 (10–12;
n = 6) long, with uniform shank, tapered proximally, upright acute thumb, curved shaft and evenly curved point; filamentous hook (FH) loop about 60% shank length. Common genital pore ventral at level of intestinal bifurcation. Copulatory complex 26 (23–28;
n = 8) long; male copulatory organ (MCO) a coiled tube with about two and a half clockwise rings, elongate bulbous base with distal bilateral flange; proximal ring diameter, 16–20. Accessory piece 13 (11–14;
n = 3) long, flattened with distal claw enclosing terminal portion of MCO. Gonads overlapping, germarium pre-testicular (germarium ventral to testis); margins of testis obscured by the germarium. Testis, 22–25 long, 15–17 wide; proximal vas deferens slightly visible; seminal vesicle a simple dilation of distal vas deferens, lying on body midline posterior to MCO; prostatic reservoir not observed. Germarium pyriform, 34–35 long, 22 wide; oviduct, oötype and uterus not observed (see
Figure 1).
Vagina pore dextromarginal; vagina distally funnel shaped, proximal portion an elongate, meandering sclerotized duct before discharging into seminal receptacle situated anterior to germarium. Vitelline follicles scattered throughout trunk except in regions of reproductive organs. Egg not observed.
Taxonomic summary:—Type host: Beaugregory Stegastes xanthurus (Poey, 1860), (Perciformes: Pomacentridae). Type locality: Cayo Arcas Reef, Gulf of Mexico. Site in host: Gill lamellae.
Prevalence and intensity of infection:—A total of 2 fish infected of 2 examined (100%); intensity on infection, 4.5.
Specimens examined:—Holotype, eight paratypes in CNHE (11,766 and 11,767, respectively).
Etymology:—This species is derived from the family name of its host.
Remarks:—Paraeuryhaliotrema pomacentris n. sp. is the type species of the genus.
3.1.3. Description of Neohaliotrema variabilis n. sp.
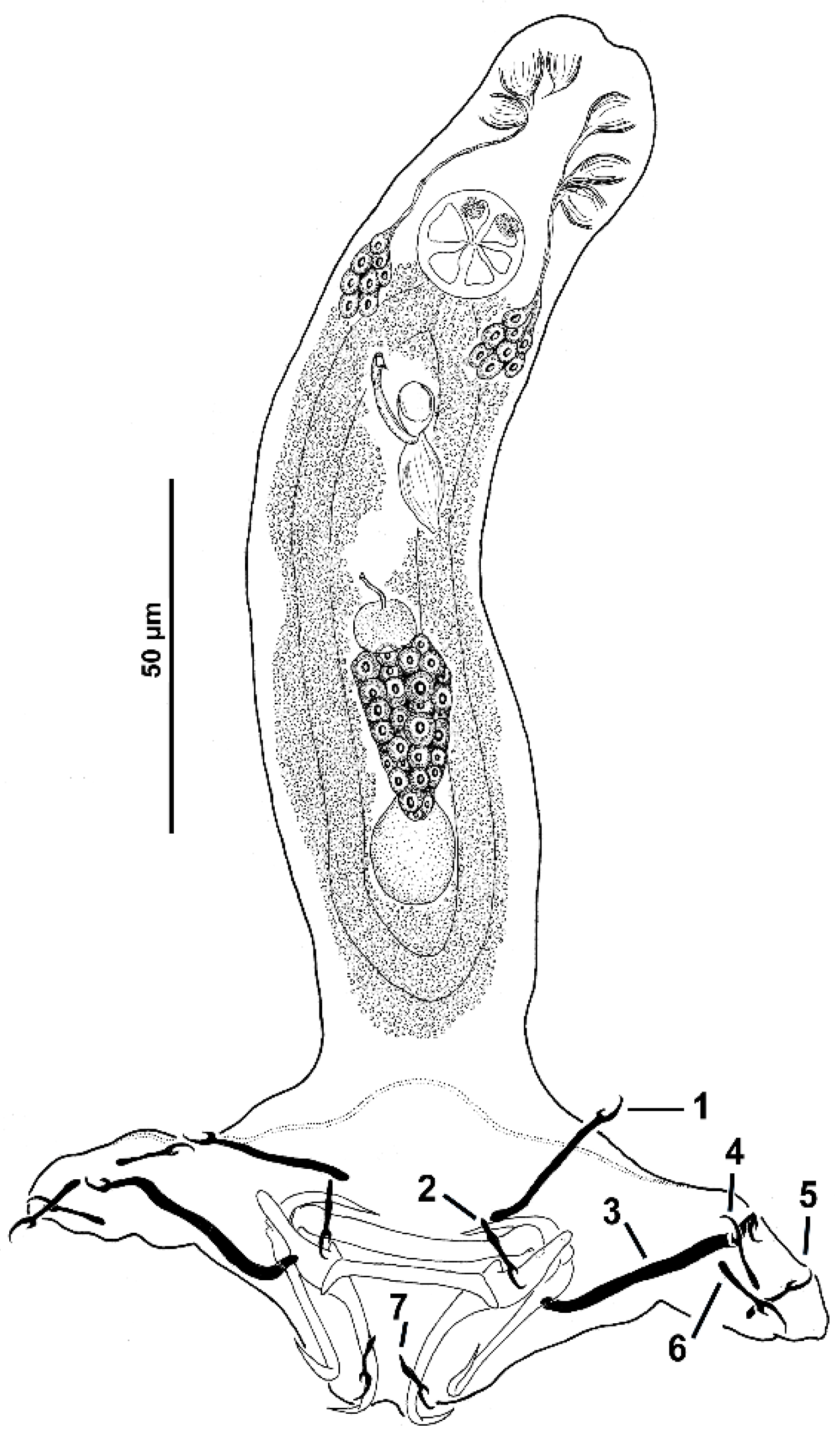
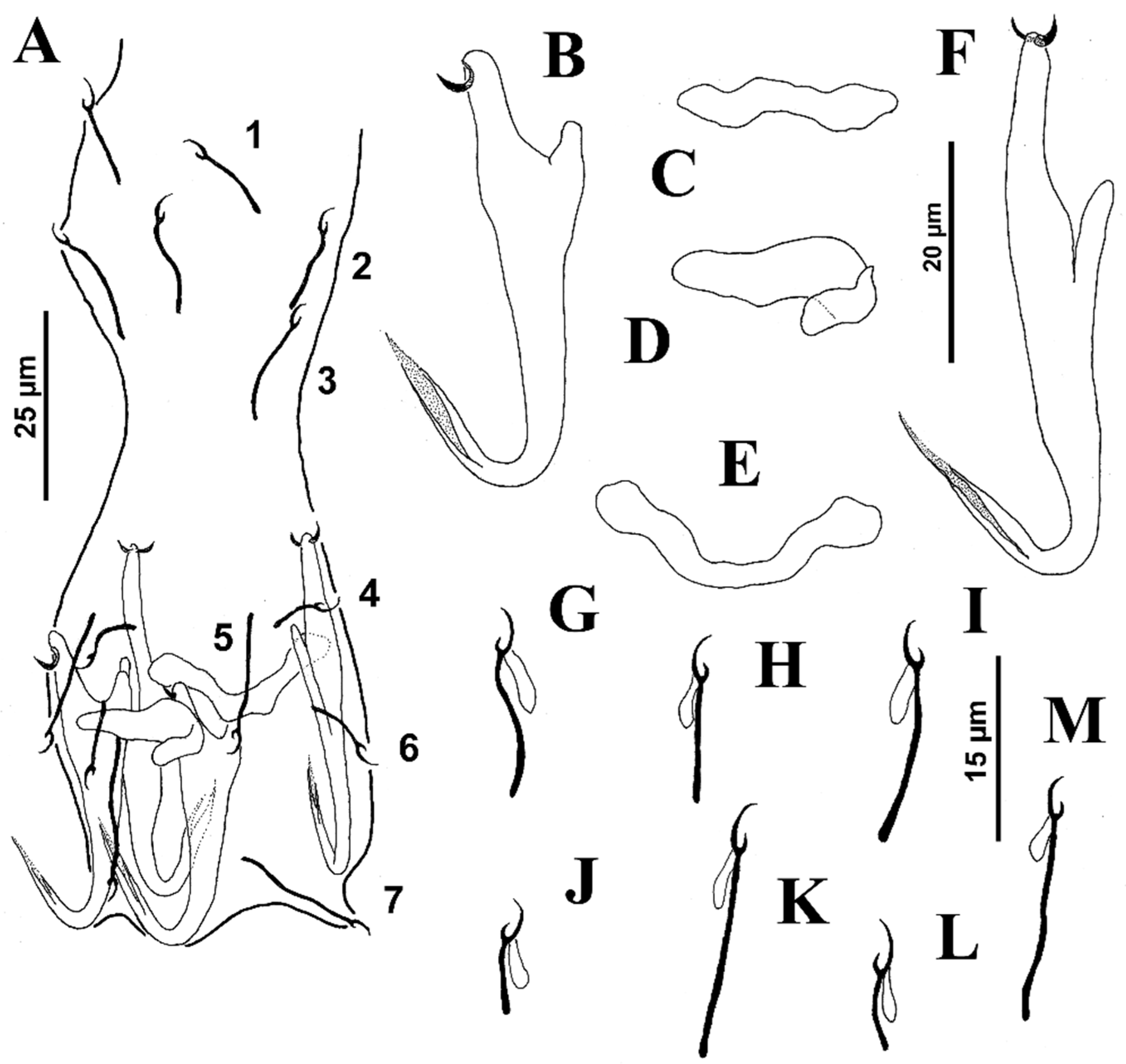
Body T-inverted shaped; greatest width at level of gonads. Cephalic region broad; cephalic lobes poorly differentiated. Three bilateral pairs of head organs well developed; cephalic glands comprising small cephalic cells posterolateral to pharynx. Tegument smooth. Two prominent eyespots (some specimens with lenses); free chromatic granules or scarce in cephalic region. Mouth subterminal, midventral, prepharyngeal. Pharynx subspherical; esophagus short to nonexistent; intestinal ceca two, lacking diverticula, confluent posterior to testis. Peduncle broad and short; haptor with bilateral elongated ends containing hook pairs 4–6. Ventral anchor, variable, with tapered deep root, moderately elongated rounded or tapered superficial root, straight shaft, short point. Dorsal anchor, variable, with poorly developed deep root, elongated acute superficial root, distally curved, straight shaft and short point. Ventral bar, variable, rod-shaped with tapered ends directed posteriorly; dorsal bar, rod-shaped, from straight to slightly arched with laterally rounded ends directed posteriorly. Hook pairs 1 and 7 smaller than other hook pairs; hooks 2 and 3 (morphometrically modified); hook pairs 1 and 7 with uniform shank, tapered proximally, upright acute thumb, curved shaft and evenly curved point; filamentous hook (FH) loop about 60% shank length; hook pairs 4–6 with slender shank, protruding thumb, curved shaft and evenly curved point; filamentous hook (FH) loop about 60% shank length; hook pairs 1 and 3 with uniform shank, protruding thumb, filamentous hook loop (FH) loop imperceptible; pair three robust with twisted shaft and point (usually). Common genital pore ventral at level of intestinal bifurcation. Copulatory complex comprising MCO and accessory piece; MCO a simple sclerotized, straight or C-shaped tube and delicate sheath-like accessory piece enclosing it, posteroventrally bent distally end; a conspicuous spherical MCO base. Gonads slightly overlapping, germarium pre-testicular (germarium ventral to testis) (see
Figure 2 and
Figure 3).
Testis subspherical; proximal vas deferens not observed; seminal vesicle elongated, a simple dilation of distal vas deferens, lying on body midline posterior to MCO; prostatic reservoir not observed. Germarium pyriform; oviduct, oötype and uterus not observed. Vagina pore dextromarginal, comprising a poorly sclerotized tube connected to subspherical seminal receptacle on midventral position, anterior to germarium. Vitelline follicles scattered throughout trunk except in regions of reproductive organs. Egg not observed. Measurements from different host species are provided in
Table 2.
Taxonomic summary:—Type Host: Bicolor damselfish Stegastes partitus (Poey, 1868), (Perciformes: Pomacentridae). Other hosts: Beaugregory Stegastes xanthurus (Poey, 1860) and Stegastes leucostictus (Müller & Troschel, 1848). Type locality: Cayo Arcas Reef, Gulf of Mexico. Site in host: Gill lamellae.
Prevalence and intensity of infection:—A total of 4 fish infected of 5 examined (80%); intensity on infection, 6.2.
Specimens examined:—Holotype, 24 paratypes in CNHE (11,768); 4 (from S. xanthurus) and 3 (from S. leucostictus) vouchers in CNHE (11,769 and 11,770, respectively).
Etymology:—This species is derived from Latin (variabilis= variable) and refers to the variable ventral bar and anchors.
Remarks:—This new species is allocated in
Neohaliotrema Yamaguti 1965 based on agreement with the amended generic diagnosis provided by Lim and Gibson [
14], i.e., species with a single pair of eyespots, hooks morphometrically modified (especially pairs 1, 2 and 3), gonads overlapping (testis overlaps postero-dorsal region of ovary) and MCO comprising a simple tube with accessory piece.
To date, there are 13 described and/or reported species of
Neohaliotrema from pomacentrid species of
Abudefduf and
Microspathodon from the Hawaiian coast, Gulf of Mexico off Cuba, China, Malaysia and Palmyra Atoll, Central Pacific (see [
14,
16,
24]). The new species differs from other species of the genus in lacking anchors with inner root sclerites and bars straight to broad-V- or U-shaped (present in all and some species from Malaysia, respectively) and vaginal aperture sinistral (dextrally oriented in the new species). Based on comparative haptoral morphology,
N. variabilis n. sp. most closely resembles
Neohaliotrema nagibinae Zhukov 1976 originally described on yellowtail
Microspathodon chrysurus (Cuvier, 1830) from the Gulf of Mexico.
Both monogenoidean species possess a haptor with bilateral elongated ends and morphometrically similar anchors (see metrical differences in
Table 2), especially dorsal anchors with elongate acute superficial root.
Neohaliotrema variabilis n. sp. differs from
N. nagibinae by having a rod-shaped ventral bar with tapered ends directed posteriorly (slightly curved in
N. nagibinae) and slightly arched dorsal bar with laterally rounded ends directed posteriorly (dorsal bar with comparatively curved and tapered ends in
N. nagibinae, see
Figure 1 on page 364 in Zhukov [
16]). Additionally, it differs in the size of its hooks and MCO; hook pair 1 (length 9–10 vs. 11–14 in
N. nagibinae), hook pair 2 (25–30 vs. 17–25) and hook pair 3 (39–42 vs. 54–66); MCO (length 12–20 vs. 30–31 in
N. nagibinae) (see
Table 1). Other structures, such as reproductive organs (i.e., gonads, vagina and accessory piece (all these herein described)) are not described and/or illustrated in the original description of
N. nagibinae (see [
16]). Only two specimens of
N. variabilis n. sp. exhibited an inconspicuous central hole on the haptor (not illustrated), it might be associated with the term of “fenestrated haptor” of Lim and Gibson [
14]. Furthermore, present specimens showed that vas deferens (not illustrated because it is densely covered by vitellaria) is looping in the left intestinal caecum (see
Figure 2) (vas deferens does not cross intestinal caecum in the emended generic diagnosis of Lim and Gibson [
14]). Although the shape of the ventral bar and anchors of
N. variabilis n. sp. are relatively constant, the size of these structures varied among specimens. These variants are considered conspecific since specimens with bars/anchors of intermediate size were found in the collection. While the ventral bar is slightly deformed as a result of coverslip pressure, the indistinct ends directed posteriorly of the ventral bar (see
Figure 3O), in one of the three specimens of
N. variabilis n. sp. found on
S. leucostictus, the basic morphology of the ventral bar shows their conspecificity with this species; the morphology of the hook pair two the with shank slightly showing a junction (see
Figure 3P) also is considered as a variation of
N. variabilis n. sp. Lastly, measurements of the ventral and dorsal bars for
N. nagibinae were imported from Zhukov 1976 and incorrectly added in
Table 2 presented by Lim and Gibson [
14], in that the ventral bar (as that connecting the ventral anchors, as illustrated in the Zhukov’s original drawings) is longer and the dorsal bar is shorter (see correction in
Table 1). Present finding of
N. variabilis n. sp. represents the first dactylogyrid described and reported on
Stegastes spp. from the Gulf of Mexico.
3.1.4. Description of Neohaliotrema manubrium n. sp.
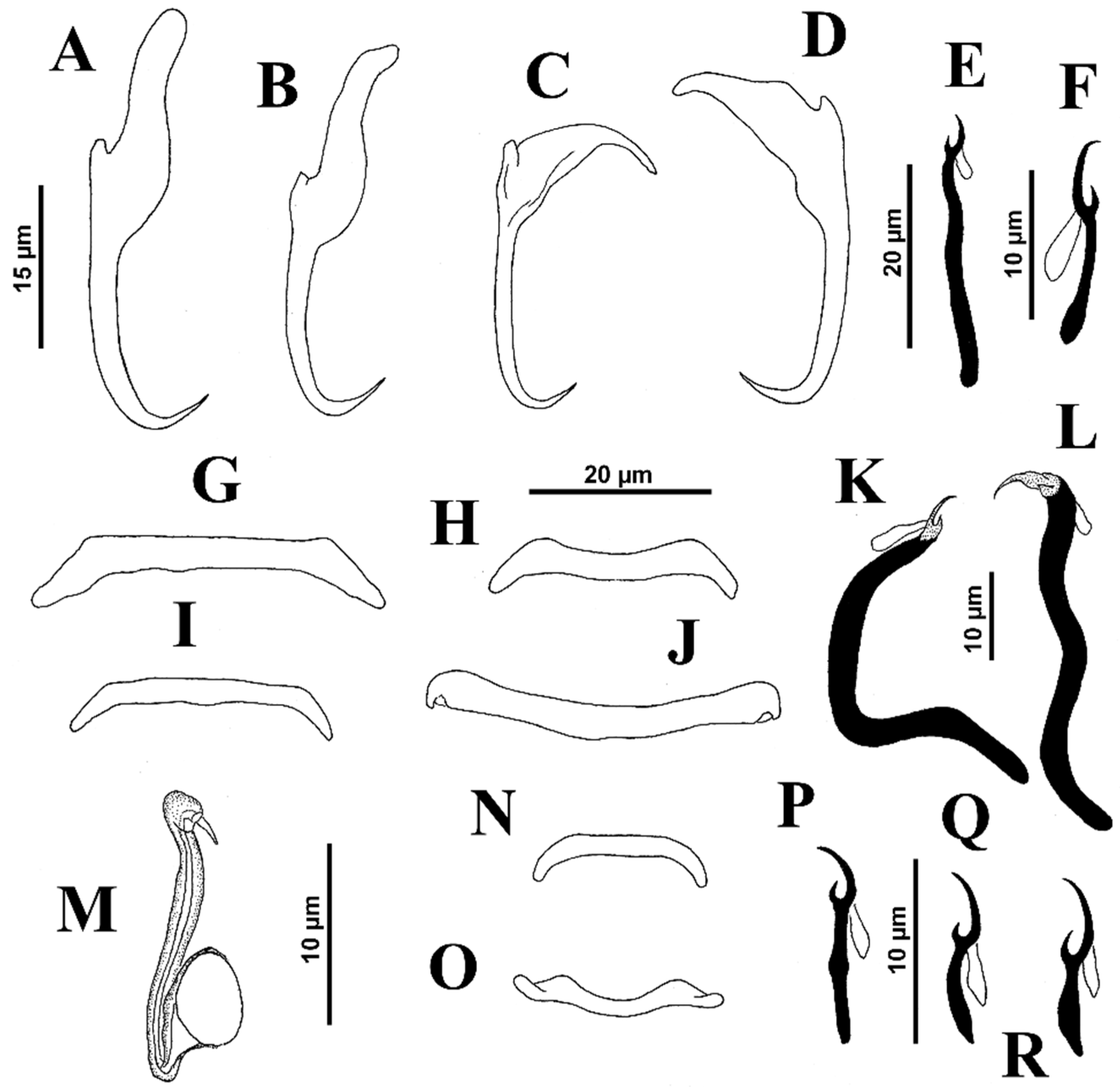
Body proper elongated with lateral margins usually parallel, 495 (390–570;
n = 20) long, with greatest width 71 (50–90;
n = 15) at level of gonads. The cephalic region narrowed; cephalic lobes poorly differentiated. Three bilateral pairs of head organs well developed; cephalic glands comprising small cephalic cells posterolateral to pharynx. Tegument smooth. Two eyespots, each with small lenses (in some specimens); accessory chromatic granules scarce in cephalic region. Mouth subterminal, midventral, prepharyngeal. Pharynx subspherical, 26 (21–30;
n = 17) wide; esophagus short to nonexistent; intestinal ceca two, lacking diverticula, confluent posterior to testis. Peduncle broad, moderately elongated; haptor 100 (82–110;
n = 25) wide, subhexagonal, with well-developed lateral lobes with gland cell accumulations (usually). Ventral anchor 39 (38–41;
n = 10) long, with rounded deep root, tapered superficial root, curved shaft and moderately elongated point; base 15 (15–16;
n = 9) wide. Dorsal anchor 43 (41–46;
n = 6) long, with well-developed tapered deep root, elongate tapered superficial root (approximately three times the length of deep root), arching shaft, moderately elongate point extending just past level of tip of superficial root; base 17 (16–18;
n = 3) wide. Ventral bar 46 (42–54;
n = 9) long (distance between ends), handle grip-shaped, straight on its midportion forming a quadrangular area with slightly expanded ends directed anterolaterally; dorsal bar 45 (41–57;
n = 17) long, U-shaped with enlarged ends directed laterally. Hook pairs 1–3 (morphometrically modified) bigger than hook pairs 4–7; hook pair 1- 41 (37–47;
n = 23) long; hook pair 2–31 (27–34;
n = 16) long; hook pair 3- 37 (34–42;
n = 18) long; hook pairs 4–7- 13 (12–14;
n = 16) long; hook pairs 1–3 (pair 1 being more robust than others, with hooklet connected to the shank by a conspicuous junction; J-shaped hook pair 2, with hooklet at distal end of curve of J, directed posteriorly and positioned back-to-back in X-shaped formation, dorsal to hook pair 1) with uniform shank, tapered proximally, upright acute thumb, curved shaft and delicate curved point; filamentous hook loop (FH) loop about 10% shank length; hook pairs 4–7 with slender shank, upright acute thumb, curved shaft and evenly curved point; filamentous hook loop (FH) loop about 50% shank length. Common genital pore ventral at level of intestinal bifurcation (see
Figure 4).
Copulatory complex 28 (24–35; n = 16) long; MCO a simple sclerotized arched tube covered by a delicate sheath-like accessory piece; a conspicuous subspherical MCO base. Gonads slightly overlapping, germarium pre-testicular (germarium ventral to testis). Testis, 63 (40–90; n = 7) long, 15 (15–40; n = 11) wide; proximal vas deferens not observed; seminal vesicle undifferentiated from distal vas deferens, lying on the body midline posterior to MCO; prostatic reservoir not observed. Germarium pyriform, 54 (40–65; n = 6) long, 23 (18–28; n = 6) wide; oviduct not observed; uterus conspicuous, with oötype surrounded by Mehlis’ gland. Vagina pore inconspicuous, comprising an unsclerotized tube (difficult to see) connected to subspherical seminal receptacle on midventral position, anterior to germarium. Vitelline follicles scattered throughout trunk except in regions of reproductive organs. Egg subovate, 24 long (not including filament), 18 wide, with long posterior filament.
Taxonomic summary:—Host: Sergeant-major Abudefduf saxatilis (Linnaeus, 1758) (Perciformes: Pomacentridae). Locality: Cayo Arcas Reef, Gulf of Mexico. Site in host: Gill lamellae.
Prevalence and intensity of infection:—A total of 10 fish infected of 14 examined (71.4%); intensity on infection, 2.7.
Specimens examined:—Holotype, 26 paratypes in CNHE (11,771 and 11,772, respectively).
Etymology:—The specific name is from Latin (manubrium= handle grip-shaped) and refers to the shape of the ventral bar.
Remarks:—This new species most resembles
Neohaliotrema bychowskii, originally described on the same host species by Zhukov [
16] in the general morphology of the haptoral sclerites, i.e., posteriorly directed J-shaped hook pair 2.
Neohaliotrema manubrium n. sp. differs from this latter species in having anchors with curved shaft (anchors with straight and elongated shafts in
N. bychowskii) and ventral bar with differentiated straight midportion (slender and elongated ventral bar in
N. bychowskii, see
Figure 1 in Zhukov [
16] and Figure 8C,D,G,H in the present study). Additionally, it differs in the length of the ventral/dorsal anchors and bars (ventral anchors: 38–41 vs. 45–50 in
N. bychowskii; dorsal anchors: 41–46 vs. 50–55; ventral bar: 42–54 vs. 52–77; dorsal bar: 41–57 vs. 52–77). Although abundance estimation was not determined,
N. manubrium n. sp. appears to be the most common species, based on number of specimens encountered on
A. saxatilis compared to other species of
Neohaliotrema (i.e.,
N. aliamanubrium n. sp.,
N. bifidum n. sp. and
N. bychowskii in the present study) on this host species from Cayo Arcas reef.
3.1.5. Description of Neohaliotrema aliamanubrium n. sp.
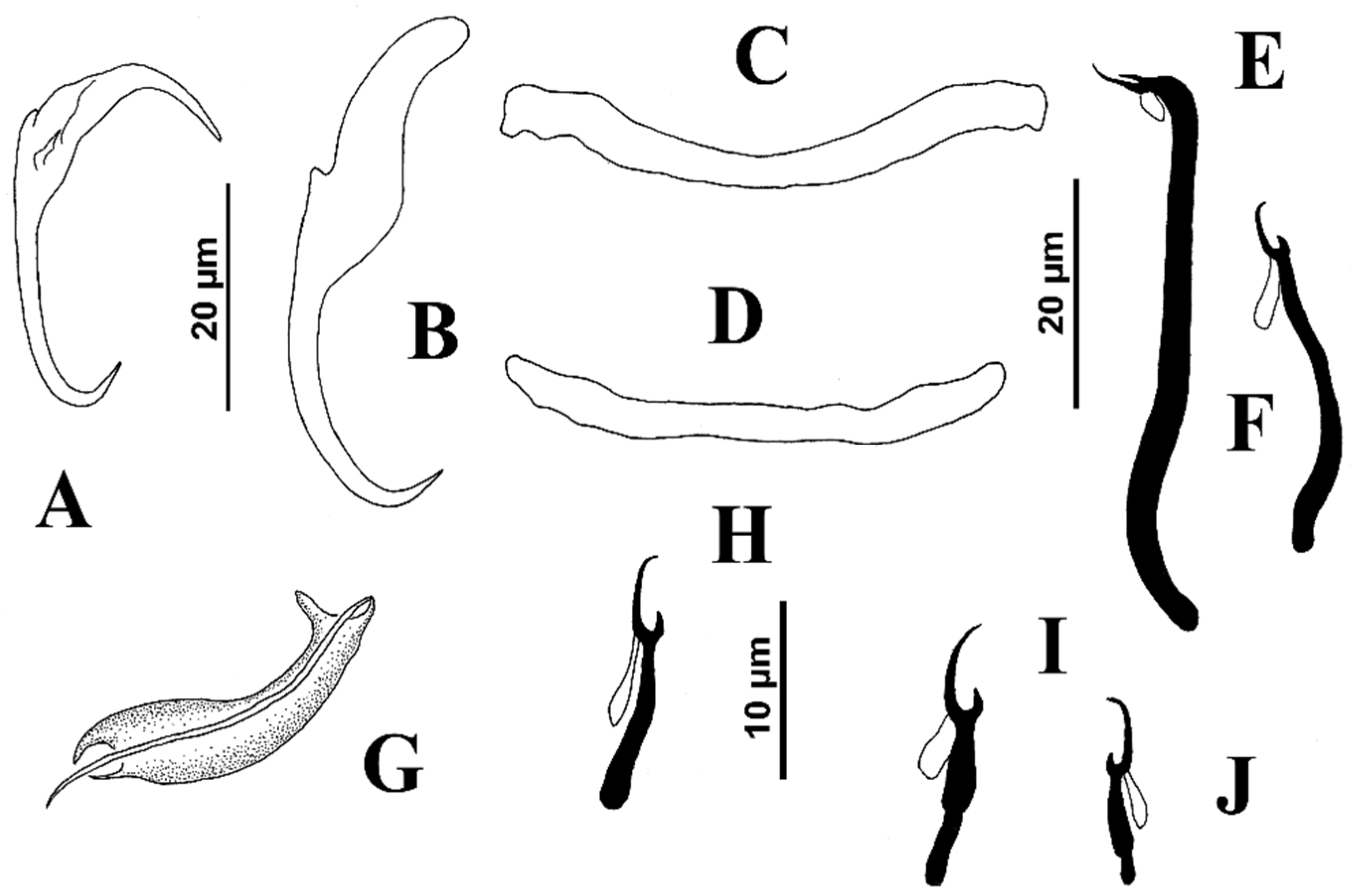
Reduced to short description and measurements of sclerotized parts of the haptor: Ventral anchor 44 (42–46;
n = 3) long, with tapered deep root, tapered superficial root, elongate and slightly curved shaft, short point; base 16–20 wide. Dorsal anchor 50 (49–51;
n = 3) long, prominent tapered deep root, moderately elongated tapered superficial root, elongate straight shaft, curved short point. Ventral bar 52–60 long (distance between ends), handle grip-shaped, with elongate expanded ends directed anteriorly; dorsal bar 40–52 long, U-shaped with curved ends directed posteriorly. Hook pairs 1–3 (morphometrically modified) bigger than hook pairs 4–7; hook pair 1,47 (45–51;
n = 4) long; hook pair 2–31 (30–32;
n = 3) long; hook pair 3 33–36 long; hook pairs 4–7- 11 (10–13;
n = 8) long; hook pairs 1–3 (pairs 1 and 2 being more robust, each with hooklet connected to the shank by a conspicuous junction; J-shaped hook pair 2, with hooklet at distal end of curve of J, directed posteriorly and positioned back-to-back in X-shaped formation, dorsal to hook pair 1) with uniform shank, tapered proximally, upright acute thumb, curved shaft and delicate curved point; filamentous hook loop (FH) loop about 0.5% shank length; hook pairs 4–7 with slender uniform shank, upright acute thumb, curved shaft and evenly curved point; filamentous hook loop (FH) loop about 40% shank length (see
Figure 5).
Taxonomic summary:—Type host: Sergeant-major Abudefduf saxatilis (Linnaeus, 1758) (Perciformes: Pomacentridae). Type locality: Cayo Arcas Reef, Gulf of Mexico. Site in host: Gill lamellae.
Prevalence and intensity of infection:—A total of 1 fish infected of 14 examined (7.1%); intensity on infection, 2.
Specimens examined:—Holotype, one paratype in CNHE (11,773 and 11,774, respectively).
Etymology:—The specific name is from Latin (alia= another + manubrium= handle grip-shaped) and refers to the shape of the ventral bar which most resembles that found in Neohaliotrema manubrium n. sp.
Remarks:—This species is described on the basis of the haptoral armament morphology because the only 2 available specimens were unstained and mounted in picrate which precluded observing details of the soft internal anatomy. Nonetheless, these specimens represent a new species, which is morphologically close to but distinct from
N. manubrium n. sp.
Neohaliotrema aliamanubrium n. sp. resembles
N. manubrium n. sp. by sharing a handle grip-shaped ventral bar and posteriorly directed J-shaped hook pair 2. The new species is easily separated from
N. manubrium n. sp. by having anchors dissimilar in shape and size, i.e., ventral anchor (length 42–46) with elongate and slightly curved shaft and short point vs. 38–41 long, with curved shaft and moderately elongate point in
N. manubrium n. sp.; dorsal anchor (length 49–51) with prominent tapered deep root, and straight elongate shaft with curved short point vs. 41–46 long, with moderately long rounded deep root, arching shaft with moderately elongate point extending just past level of tip of superficial root in
N. manubrium n. sp. (see
Figure 4I,J and
Figure 5A,B, respectively). Additionally, the ventral bar in
N. aliamanubrium n. sp. is noticeably longer than that from
N. manubrium n. sp., i.e., 52–60 vs. 42–54, respectively. The copulatory complex and vagina were not observed in
N. aliamanubrium n. sp., apparently these structures are unsclerotized.
3.1.6. Description of Neohaliotrema bifidum n. sp.
Reduced to short description and measurements of body and sclerotized parts of haptor: Body, 257 (240–285;
n = 3) long, greatest width 60, near body midlength. Two eyespots, each with small lenses; accessory chromatic granules, ovate, lightly scattered in cephalic region, ovate. Pharynx subspherical, 19–21 wide; haptor longer than wide, 55–60 wide. Ventral anchor 39 (38–40;
n = 4) long, with tapered deep root, rounded superficial root with a sclerite on its distal margin, elongated straight shaft with elongated and recurved bifid point; base 14 (13–14;
n = 3) wide. Dorsal anchor 49 (45–50;
n = 6) long, prominent tapered deep root, elongated tapered superficial root with sclerites on its distal margin, elongated straight shaft with elongated and curved bifid point; base 10 wide, not clearly differentiated with shaft. Ventral bar 22–25 long, rod-shaped; dorsal bar 25–29 long, U-shaped with dilated ends directed laterally. Hooks dissimilar in size with filamentous hook loop (FH) loop about 1–1.5 hooklet length, uniform shank; hook pair 1 17 (
n = 2) long; hook pair 2 15 long; hook pair 3 19 long; hook pair 4 12 long; hook pair 5 19 (18–20;
n = 4) long; hook pair 6 12 (10–15;
n = 4) long; hook pair 7 20 (20–21;
n = 3) long; hook pairs 1–3 located on peduncle (see
Figure 6).
Taxonomic summary:—Type host: Sergeant-major Abudefduf saxatilis (Linnaeus, 1758) (Perciformes: Pomacentridae). Type locality: Cayo Arcas Reef, Gulf of Mexico. Site in host: Gill lamellae.
Prevalence and intensity of infection:—A total of 1 fish infected of 14 examined (7.1%); intensity on infection, 3.
Specimens examined:—Holotype, 2 paratypes in CNHE (11,775 and 11,776, respectively).
Etymology:—The specific name is from Latin (bifidus= bifid) and refers to the bifid points of anchors of this parasite species.
Remarks:—
Neohaliotrema bifidum n. sp. on
A. saxatilis from Cayo Arcas, Gulf of Mexico is closest morphologically to
Neohaliotrema bombini Lim and Gibson [
14] from the indo Pacific sergeant
Abudefduf vaigensis (Quoy and Gaimard, 1825) from Malaysia, and
Abudeduf septemfasciatus (Cuvier) and
Abudefduf sordidus (Forsskål) from Palmyra Atoll Central Pacific [
24].
The characteristics shared by both monogenoidean species are anchors with bifid points and sclerites at the distal tip of their superficial roots (as “spatulate points with grooves and inner root sclerites” in Lim and Gibson [
14], hook pairs 1–3 on ventral surface of peduncle and smallest hook pair 6, i.e., 10–15 long (11–14 in
N. bombini) (see Lim and Gibson [
14]).
Neohaliotrema bifidum n. sp. differs from
N. bombini in the body length (i.e., 240–285 vs. 315–518 in
N. bombini), size of hook pairs 1–3 (i.e., 15–19 long vs. from 18 to 34 in
N. bombini), bars without anterior processes (present in
N. bombini), and ventral bar rod-shaped (V-shaped in
N. bombini). In other features,
N. bifidum n. sp. resembles species of
Haliotrematoides (Dactylogyridae) on haemulids, lutjanids and sparids fishes from the Gulf of Mexico and neighboring areas in having hook pairs 1–3 on peduncle.
Neohaliotrema bifidum n. sp. is differentiated from species of this latter genus in lacking four eyespots, dorsal anchor with perforated base and not well-developed deep roots, ventral bar usually with two submedial pockets, and hooks similar in size (all these characters are in species of
Haliotrematoides). Information on some internal features (i.e., copulatory complex and vagina) could not be determined in
N. bifidum n. sp. based on three unstained specimens mounted in picrate and glycerine jelly. Nonetheless, the species is easily differentiated from its closest congeneric species,
N. bombini, by the characters mentioned above.
3.1.7. Description of Neohaliotrema similium n. sp.
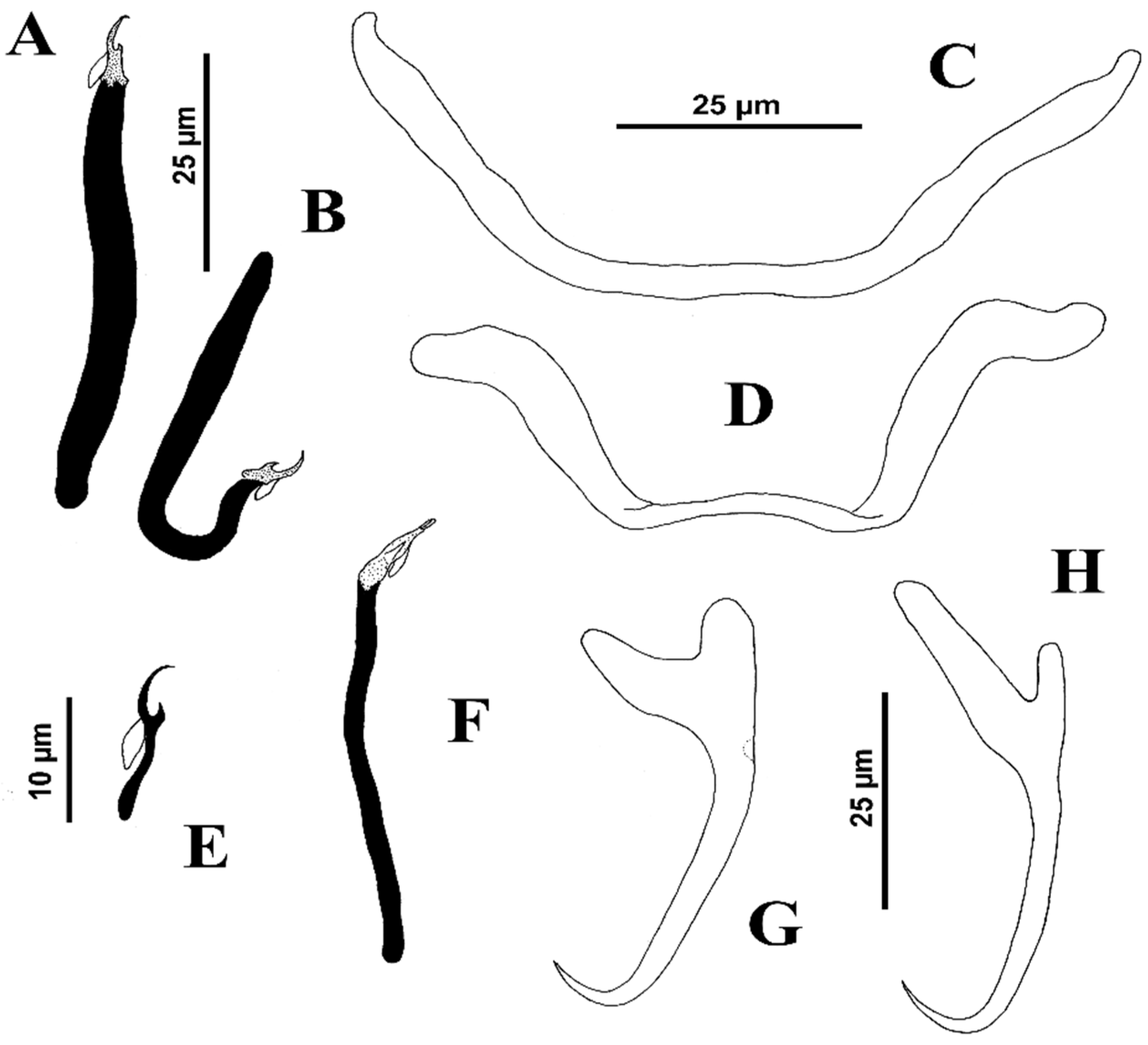
Reduced to short description and measurements of sclerotized parts of haptor: Body, 230–285 long, greatest width 82, near body mid-length. Two eyespots; accessory chromatic granules, ovate, lightly scattered in cephalic region. Pharynx subspherical, 19 wide. Haptor 90, wide. Ventral anchor 41 (39–43; n = 4) long, with poorly developed deep root, rounded superficial root, elongate and curved shaft, short point; base 12 (11–13;
n = 4) wide. Dorsal anchor 29 (27–30;
n = 3) long, inconspicuous tapered deep root, acute superficial root with curved tip directed posteriorly, slightly straight shaft, short point; base 18 (17–19;
n = 3) wide. Ventral bar 33–42 long (distance between ends), rod-shaped, with tapered ends; dorsal bar 50 (
n = 2) long, rod-shaped, slightly arched posteriorly with rounded ends. Hook pairs 2 and 3 (morphometrically modified) bigger than hook pairs 1, 4–7; hook pairs 1 and 4–6 14 (13–14;
n = 14) long; hook pair 2 27 (25–30;
n = 3) long; hook pair 3 50 (47–53;
n = 4) long; hook pair 7 10 (
n = 2) long; hook pairs 1 and 4–6, with variable shank, from slightly expanded to narrowed proximally, upright acute thumb, curved shaft and delicate curved point; filamentous hook loop (FH) loop about 50% shank length; hook pairs 2 and 3 (robust), with uniform shank, upright acute thumb, curved shaft and evenly curved point; filamentous hook loop (FH) loop about 20% and 0.5% shank length, respectively. Copulatory complex 22 long; MCO a delicate sclerotized tube covered by a grooved, Y-inverted shaped accessory piece; inconspicuous MCO base (see
Figure 7).
Taxonomic summary:—Type host: Stegastes xanthurus (Poey, 1860), (Perciformes: Pomacentridae). Type locality: Cayo Arcas Reef, Gulf of Mexico. Site of infection: Gill lamellae.
Prevalence and intensity of infection:—A total of 1 fish infected of 2 examined (50%); intensity on infection, 2.
Specimens deposited:—Holotype, 1 paratype in CNHE (11,777 and 11,778, respectively).
Etymology:—The specific name is from Latin (similis = resembling) and reflects the similarity of the species with its congeners.
Remarks:—This species most resembles N. variabilis n. sp. and N nagibinae in having similar anchors, i.e., ventral anchor with elongated rounded superficial root and dorsal anchor with acute superficial root, distally curved. Neohaliotrema similium n. sp. differs from these latter species in having a Y-inverted shaped accessory piece (rod-shaped with a conspicuous spherical MCO base in N. variabilis n. sp. and N nagibinae).
3.1.8. Description of Neohaliotrema bychowskii Zhukov, 1976
Supplemental observations (measurements from the original description follow those of the present study in parentheses, respectively): Body, 335 long, greatest width 70, near peduncle. Two eyespots with conspicuous lens; accessory chromatic granules scarce in cephalic region. Haptor, 145 wide, with well-developed lobes. Ventral anchor 47–48 (45–50) long, 18 wide (
n = 2), with well-differentiated roots, elongated straight shaft and short point. Dorsal anchor 52–53 long (50–55), well-developed rounded deep root, tapered superficial root, elongate and slightly curved shaft and point; base 18–19 wide. Ventral bar 92 long (distance between ends) (52–77), rod-shaped with enlarged ends directed anteriorly; dorsal bar 71 long (52–77), straight on its midportion with enlarged and expanded ends directed anterolaterally. Hook pairs 1–3 (morphometrically modified) bigger than hook pairs 4–7; hook pair 1–56–55 (49–65) long; hook pair 2–45–47 (49–65) long; hook pair 3–49–50 (49–65) long; hook pairs 4–7–13 (12–13;
n = 3) (10–16); hook pairs 1–3 with hooklet connected to the shank by a conspicuous junction, uniform shank, rounded proximally, upright acute thumb, curved shaft and delicate curved point; filamentous hook loop (FH) loop about 0.5% shank length; J-shaped hook pair 2, with hooklet at distal end of curve of J, directed posteriorly and positioned back-to-back in X-shaped formation, dorsal to hook pair 1); hook pairs 4–7 with slender shank, upright acute thumb, curved shaft and evenly curved point; filamentous hook loop (FH) loop about 50% shank length (see
Figure 8).
Taxonomic summary:—Host: Sergeant-major Abudefduf saxatilis (Linnaeus, 1758) (Perciformes: Pomacentridae). Locality: Cayo Arcas Reef from the Gulf of Mexico. Site in host: Gill lamellae.
Prevalence and intensity of infection:—A total of 1 fish infected of 14 examined (7.1%); intensity on infection, 1.
Specimens examined:—One reference specimen in CNHE (11,779).
Remarks:—This species was originally described from the same host species from the Gulf of Mexico by Zhukov [
16] providing illustrations of the sclerotized structures of the haptor and copulatory complex. Herein, the identification of a single specimen as
N. bychowskii is based on the comparative morphology of the haptoral structures which is basically equivalent morphometrically with that originally described. The single specimen examined is unstained, which precludes determination of internal anatomy, i.e., the morphology of the copulatory complex and vagina.
3.1.9. Description of Neohaliotrema macracanthum Zhukov, 1976
Supplemental observations (measurements from Zhukov [
16] follow those of the present study in parentheses, respectively): Body, 309 (275–340;
n = 4) (120–330) long, with greatest width 47 (45–50;
n = 4) (66–110) at level of body midlength. Two eyespots, each with lenses (present in 3 specimens); accessory chromatic granules in cephalic region, ovate. Haptor 70–95 wide. Ventral anchor 41 (40–46;
n = 7) (34–39) long, with well-differentiated roots, elongate straight shaft with distal inner blade, straight recurved point; base 13 (12–14;
n = 4) wide. Dorsal anchor 53 (50–55;
n = 6) (43–52) long, with robust rounded deep root, tapered superficial root, elongate and straight shaft with prominent distal inner blade, straight recurved point; base 16 (15–17;
n = 4) wide. Ventral bar 42 (35–48;
n = 3) (32–43), long (distance between ends), rod-shaped, slightly arched posteriorly; dorsal bar 40 long (34–42), slightly V-shaped, expanded posteriorly on its midportion with ends directed laterally. Hook pairs 1–3 (morphometrically modified) longer than hook pairs 4–7; hook pair 1 34 (32–35;
n = 4) (26–33) long; hook pair 2 26 (25–28;
n = 3) (24–28) long (proximal subunit); hook pair 3 38 (36–39;
n = 3) (35–45) long; hook pairs 4–7 11 (10–14;
n = 7) (10–16) long; hook pair 2, J-shaped with robust proximal subunit, with hooklet at distal end of curve of J (slender distal subunit), directed posteriorly and positioned back-to-back in X-shaped formation, dorsal to hook pair 1, each with uniform shank, tapered proximally, upright acute thumb, curved shaft and delicate curved point; filamentous hook (FH) loop about 0.5% shank length; hook pairs 4–7 with slender shank, upright acute thumb, curved shaft and a delicate point; filamentous hook (FH) loop about 50% shank length. MCO, U-shaped, 34 long (43–47 length for the copulatory organ tube in Zhukov, 1976).
Taxonomic summary:—Host: Sergeant-major Abudefduf saxatilis (Linnaeus, 1758)
Locality: Cayo Arcas Reef, Gulf of Mexico. Site in host: Gill lamellae.
Prevalence and intensity of infection:—A total of 1 fish infected of 14 examined (7.1%); intensity on infection, 4.
Specimens examined:—4 reference specimens in CNHE (11,780).
Remarks:—The only previous descriptive information on
N. macracanthum is that of the original description based on seven specimens from the same host species in the Gulf of Mexico off Cuba. This species is clearly differentiated from its congeners in having blades on the inner surfaces of the anchor shafts (see [
16]). Present specimens did not significantly differ in morphology from those in the original account; the differences in measurements of the MCO length (i.e., 34 vs. 43–47) are apparently attributable to intraspecific variation and/or effects of different methods of specimen collection/preparation and measuring. In addition of
N. macracanthum, which is also reported on night sergeant
Abudefduf taurus (Müller & Troschel) from the Gulf of Mexico off Cuba, there is another species of
Neohaliotrema with blades on the inner surfaces of their respective anchor shafts,
Neohaliotrema sp. on Indo-Pacific sergeant
Abudefduf vaigiensis (Quoy & Gaimard) from Malaysia [
14,
16]. These superficial blade-like projections on the distal shaft of the anchors have also been described in some species of
Haliotrematoides on haemulids, lutjanids and sparids fishes from the Gulf of Mexico. The resemblance of this latter characteristic between species of both monogenoidean genera likely represents an example of convergence in shape rather than a phylogenetic relationship.
3.1.10. Redescription of Haliotrema brevicirrus Zhukov, 1980
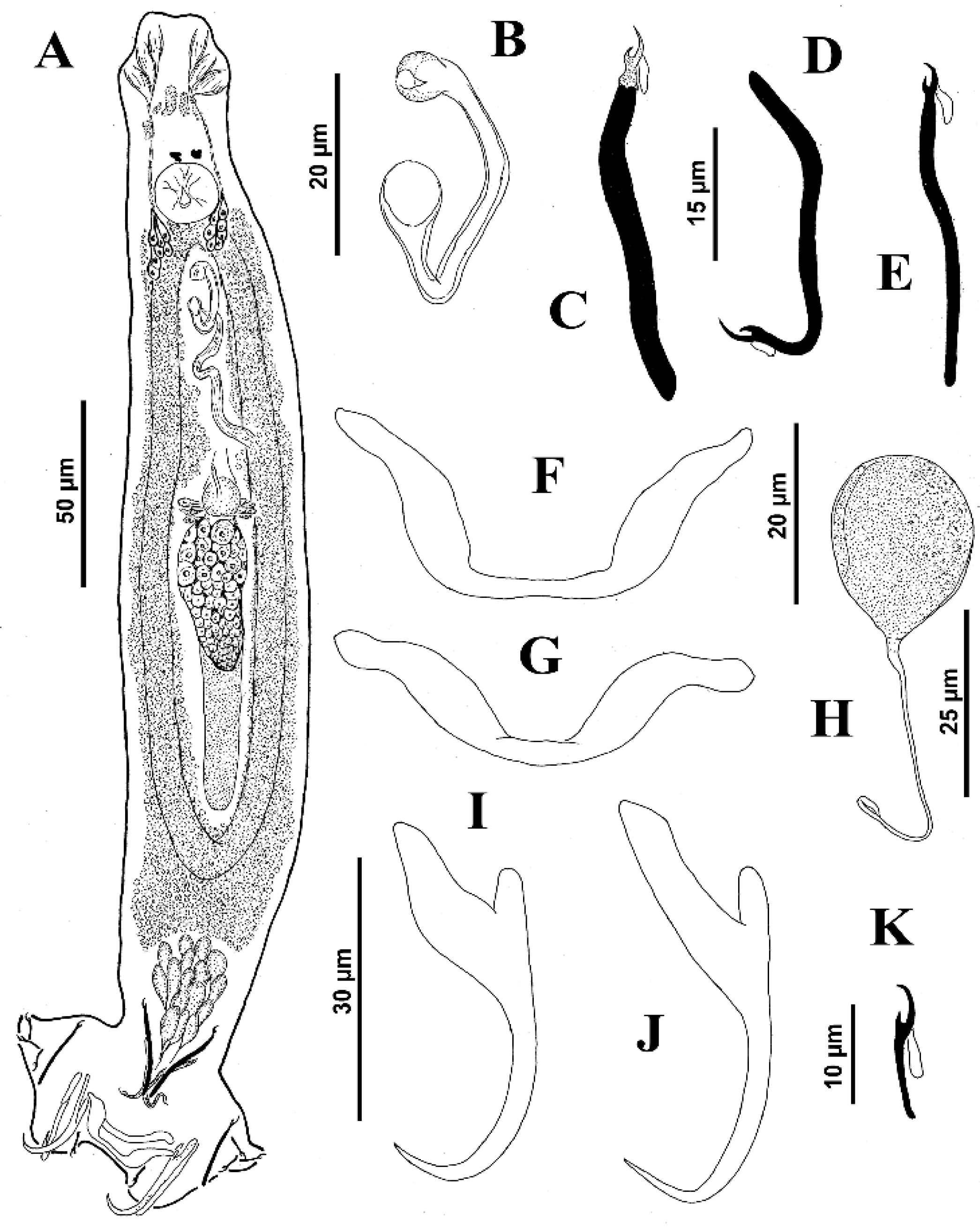
Measurements from the original description follow those of the present study in parentheses, respectively: Body fusiform, 575 (495–730;
n = 16) (210–370) long, with greatest width 101 (85–130;
n = 10) (80–140) at level of gonads. Cephalic region broad; cephalic lobes moderately developed. Three bilateral pairs of head organs connected to cephalic glands located posteriorly to pharynx. Two pairs of eyespots; members of anterior pair slightly closer together, accessory chromatic granules small, few in cephalic region. Mouth subterminal, midventral, prepharyngeal. Pharynx subovate, 33 (28–38;
n = 17) wide; oesophagus short; intestinal ceca confluent posterior to gonads, lacking diverticula. Peduncle elongated; haptor 112 (97–127;
n = 11) wide, sub-hexagonal. Ventral anchor 44 (41–47;
n = 8) (45–60) long, broad base with poorly differentiated roots, slightly straight shaft, moderately elongated point; base 20 (19–23;
n = 4) wide; dorsal anchor 42 (41–42;
n = 3) (51–68) long, base-shaft junction hinged, inconspicuous deep root, elongate superficial root, slightly curved shaft, elongated point. Ventral bar 49 (43–57;
n = 11) (50–67) long, opened M-shaped, with rounded ends; dorsal bar 50 (47–58;
n = 14) (51–68) long, slightly arched anteriorly, with sublateral anterior expansions. Hooks 12 (12–13;
n = 13) (12–14) long, all similar, with rounded thumb, delicate point, slender shank, uniform, slightly curved proximally; filamentous hooklet (FH) loop about 90% shank length. MCO, 55 (47–60;
n = 9) long, tapered, tubular, slightly twisted tip, cylinder-shaped base, terminating in diagonal opening; accessory piece, membranous, shaped like a vane, encircling MCO to form two complete spirals from anterior margin of base to terminal portion of MCO. Gonads in tandem, germarium pre-testicular; testis 75 (47–98;
n = 12) long, 37 (15–48;
n = 12) wide, subovate, intercecal, lying slightly posterior to body midlength; proximal vas deferens and seminal vesicle not observed. Prostatic reservoir large, pyriform, tapered distally; prostatic gland cells not observed. Germarium, 48 (32–65;
n = 14) long, 29 (24–40;
n = 13) wide, irregular; oviduct, oötype and uterus not observed. Vagina pore dextromarginal; vaginal canal narrowed, extending diagonally to join subspherical seminal receptacle, anterior to germarium. Vitelline follicles scattered throughout trunk except in regions of reproductive organs. Eggs not observed (see
Figure 9).
Taxonomic summary:—Host: Spotfin butterflyfish Chaetodon ocellatus Bloch 1787 (Acanthuriformes: Chaetodontidae). Locality: Cayo Arcas Reef, Gulf of Mexico. Site in host: Gill lamellae.
Prevalence and intensity of infection:—A total of 4 fish infected of 5 examined (80%); intensity on infection, 12.2.
Specimens examined:—A total of 19 reference specimens plus 30 additional specimens (not measured) in CNHE (11,781).
Remarks:—The original description of this species by Zhukov [
25] was based on four specimens from the gills of
Chaetodon striatus Linnaeus 1758 (type host) in association with two other butterflyfishes,
Chaetodon capistratus Linnaeus 1758 and
C. ocellatus, all from the Gulf of Mexico. Since then,
H. brevicirrus has not been reported on these latter or other filial hosts. The morphology of the haptoral sclerites and copulatory complex of the newly collected specimens of
H. brevicirrus from
C. ocellatus revealed that it is consistent with that of the original description of this species (see
Figure 1 in Zhukov [
25]).
However, while these latter structures are satisfactory for specific identification, other organs are not clearly identified in the original description. Therefore, we provided a needed redescription of
H. brevicirrus including an illustration of a whole specimen to clarify the morphology of this species, i.e., position of the vaginal aperture and gonads (both characters not described and/or illustrated in Zhukov [
25]). Additionally, the presence of an accessory piece on MCO found in the present specimens was not described in the original description of this species. Consistently, Cruces et al. [
26] also described an accessory piece in their new species,
Haliotrema sanchezae on
Scarus perrico Jordan & Gilbert (Scaridae) from Peru and listed 11other species of
Haliotrema possessing it of which only
H. pacificum, (Mizelle & Kritsky, 1969) Vala, Maillard & Overstreet, 1982, is found on a chaetodontid,
Johnrandallia nigrirostris (Gill, 1862) in the Mexican Pacific. Specimens from
C. ocellatus metrically differ from those specimens from
C. striatus by the body length (495–730 vs. 210–370 in the original description) and dorsal anchor length (41–42 vs. 51–68). These metric differences of the body and dorsal anchors in specimens from different host species, are considered as intraspecific variation (i.e., effect of the host size). Additionally, it is worth noting that the length of the MCO of
H. brevicirrus provided by Zhukov [
25] was measured in two parts, i.e., copulatory organ tube plus MCO base length. Therefore, we deduce that the MCO length of
H. brevicirrus from
C. ocellatus (present study) is nearly identical metrically to that of
H. brevicirrus from
C. striatus, i.e., 47–60 vs. 30–38 plus 19–25 (copulatory organ tube and MCO base of Zhukov [
27], respectively). In addition of those species of
Haliotrema from chaetodontids mentioned above, there is another species reported from the Mexican Pacific, i.e.,
H. cirrhitusi Mendoza-Franco & Violante-Gonzalez, 2011 from
Cirrhitus rivulatus Valenciennes (Cirrhitidae) [
28], and other 10 species described from Gulf of Mexico by Zhukov [
24,
26] (i.e.,
H. conspecta,
H. auribaculum,
H. curvicirrus,
H. longirectocirrus,
H. myripristisi,
H. tenuihamus,
H. papillibaculum,
H. ampliomacrohamus,
H. ampliomicrohamus and
H. teuthis) from fishes of the Scaridae, Holocentridae and Acanthuridae. However, a revision of these latter
Haliotrema spp. Is probably necessary to recognize their generic status. Only
Haliotrema lactophrys (MacCallum, 1915), Vala, Maillard & Overstreet, 1982, have been described and/or reported on scrawled cowfish
Acanthostracion quadricornis (type-host) (Linnaeus, 1758) and honeycomb cowfish
A. polygonius Poey, 1876 (Tetraodontiformes: Ostraciidae) from the Sainte Anne Guadeloupe, West Indies, New York aquarium and Campeche coast in the Gulf of Mexico, being this latter considered as undetermined in Mendoza-Garfias et al. [
29,
30]. So, it remains unclear whether host or locality are correctly registered for
H. lactophrys in the Gulf of Mexico.
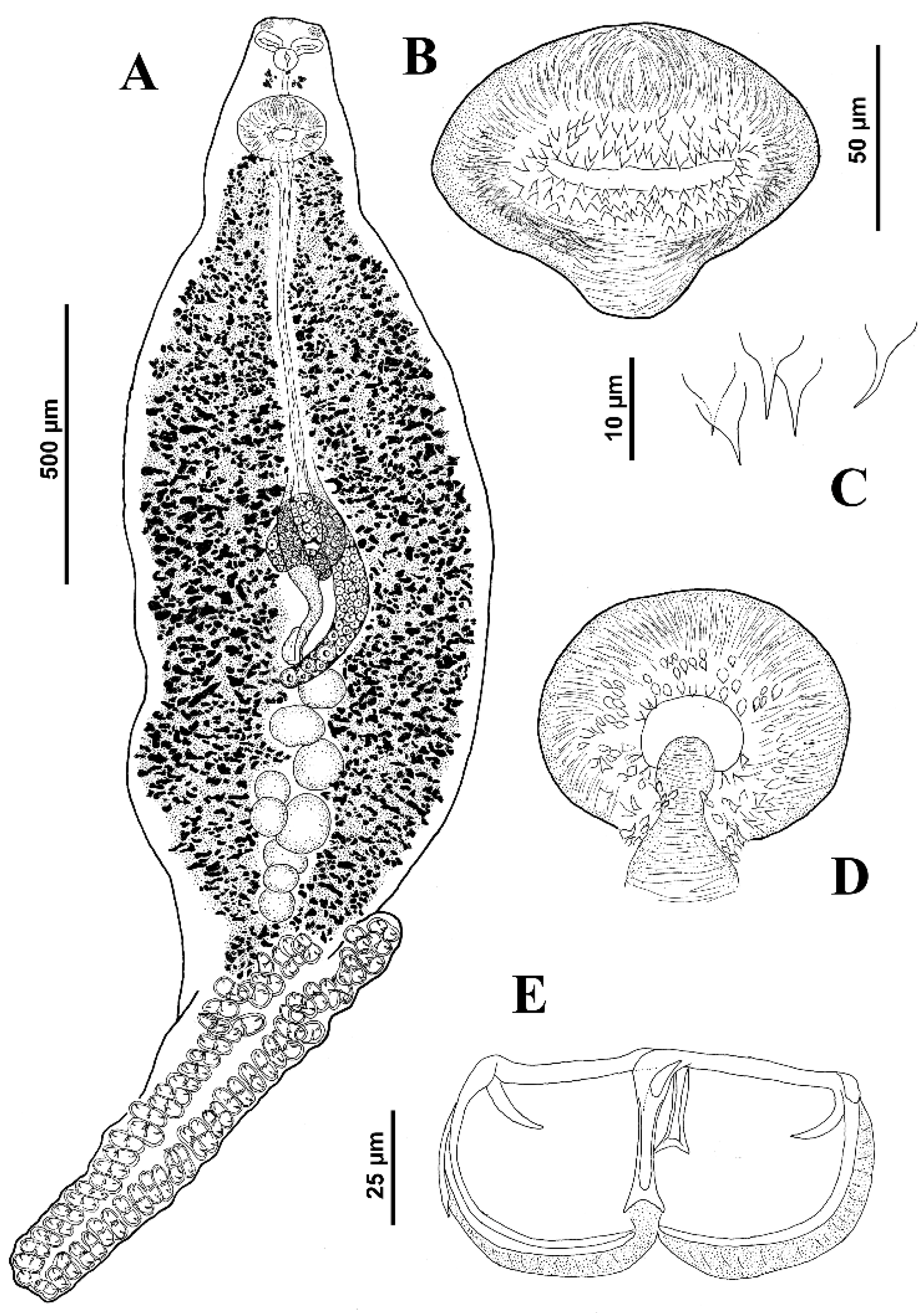
3.1.11. Description of Microcotyle multilineatus n. sp.
Body fusiform, anterior end narrow, total length of body 1837 (987–2350;
n = 12); body width, 550 (325–650;
n = 8) at level of ovary. Haptor subsymmetrical or symmetrical, 742 (600–900;
n = 3) long, armed with 70–72 clamps, arranged in two equal or sub-equal rows, joining at anterior and posterior extremities of haptor; 30 (25–40;
n = 18) long, 58 (45–72;
n = 19) wide. Mouth subventral, within funnel-shaped vestibule with pair of septate buccal suckers, elongate-oval, 30 (26–33;
n = 10) long, 51 (50–58;
n = 11) wide. Pharynx subspherical, 37 (32–40;
n = 5) long, 38 (35–40;
n = 6) wide; oesophagus short, connected to intestinal bifurcation at posterior margin of genital atrium. Caeca extends up to haptor peduncle, with inner and external lateral ramifications (external more profuse). Testes 11–14 in number, sub-elliptical to irregular, 65 (62–87;
n = 6) long, 74 (50–100;
n = 6) wide, grouped in testicular fields, post-germarial and pre-haptoral (partially extending into haptor peduncle); testes arranged in clusters of 1 or 2 rows generally, with some testes dorsoventrally overlapped. Vas deferens slender, straight, runs in midline to genital atrium; copulatory organ muscular 27 (23–30;
n = 8) wide, located in posterior margin of the genital atrium, 82 (70–90;
n = 6) long, 99 (95–100;
n = 6) wide. Genital atrium at 141 (125–162;
n = 6) from anterior extremity of body, atrium proper as inverted pear, comprising a medial muscular chamber, armed with small conical spines arranged in a tight circle on distal half of MCO bulb, with concentrically tips directed into genital atrium. Variation in focus shows surrounded subspherical dorsal spines at the chamber opening. Germarium 211 (187–250;
n = 3) long, 165 (145–185;
n = 4) wide, convoluted, begins at level of anteriormost testes, with proximal arch directed sinistro-dextrally, continues anteriorly in midline, connected to wider distal arch directed sinistro-dextrally at level of confluence of vitelline ducts, reflexes a last time posteriorly connecting to slender arch directed dextro-sinistrally. Oviduct and oötype with posterior Mehlis gland not observed. Precise junctions between oviduct, oötype, vitelline duct and genitointestinal canal not elucidated. Seminal receptacle well visible, dextral to germarium. Canal from vagina not observed. Vaginal pore mid-dorsal, often inconspicuous in most specimens, posterior to genital atrium; distance from vagina to anterior body margin, 275–302. Vitellarium densely scattered in most part of body from genital atrium to haptor. Vitelline ducts pair, united anteriorly and posteriorly; anterior junction not visible; posterior junction conspicuous as a Y in most specimens, at level of anterior margin of germarium and dorsal to it. Egg not observed (see
Figure 10).
Taxonomic summary:—Type host: Brown chromis, Chromis multilineata (Guichenot, 1853) (Perciformes: Pomacentridae). Type locality: Cayo Arcas Reef, Gulf of Mexico (0.21° N, −91.98° W). Site in host: Gill lamellae.
Prevalence and intensity of infection:—A total of 3 fish infected of 14 examined (21.4%); intensity on infection, 4.6.
Specimens examined:—Holotype and 13 paratypes in CNHE (11,782 and 11,783, respectively).
Etymology:—This species is derived from the specific name of its host.
Remarks:—This new species was allocated to
Microcotyle on the basis of the diagnosis provided by Mamaev [
31] and on examination of original descriptions of other species of this genus in Villora-Montero et al. [
32] and Muñoz and George-Nascimento [
33]: species mainly characterized by a conspicuous genital atrium with well-developed radial muscles, armed with conical spines (variable in size) communicated with small chambers, a convoluted ovary and an unarmed, long, single vaginal duct. Fernández and Corrada [
17] described their new species,
Microcotyle chromidis from the gills of the blue chromis,
Chromis cyanea (Poey, 1860) from Cuba from which it differs from
Microcotyle multilineatus n. sp. in having greater body length, i.e., 1850–2350 vs. 416–1035 long in
M. chromidis, smaller anterior clamps 25–36 long, 46–63 wide, posterior clamps 25–40 long, 52–72 wide vs. 74–122 long, 42–88 wide; number of testes, 11–14 vs. 24–29 and by the general morphology of the genital atrium and organization of their spines (single chamber surrounded with spines vs. more than one chamber with spines laterally distributed in
M. chromidis-see
Figure 1D in Fernández and Corrada [
17] and present study). The fact is that these latter authors indicated that
M. chromidis was also posteriorly reported on
Cr. multilineata in Cuba. However, these authors did not provide any comparative morphometric data of their
M. chromidis on
Cr. multilineata nor accession number for their parasite specimens deposited to any collection as reference. Based on present description,
M. multilineatus n. sp. is well differentiated from
M. chromidis on
Cr. cyanea. However, it is unclear if these specimens reported as
M. chromidis on
Cr. multilineata from Cuba are conspecific with
M. multilineatus n. sp. on the same host species from Cayo Arcas reef in the Gulf of Mexico. What is noteworthy is that four individuals of
Cr. cyanea (type host for
M. chromidis) were examined and resulted negative for monogenoids in the present study. So, it is possible that
Cr. cyanea and/or
Cr. multilineata were misidentified as hosts of
M. chromidis within same geographical area of Cuba. Microcotylids have been described and/or reported in the Mexican waters (including the Gulf Coast of USA, i.e., state of Florida) on a wide variety of members of the Centropomidae, Cirrhitidae, Chaetodontidae, Gerreidae, Haemulidae, Labridae, Lutjanidae, Mugilidae, Pomacanthidae, Pomatomidae, Sciaenidae, Serranidae, and Sparidae (see [
34,
35,
36]). However, a revision and accurately counting of some characters (i.e., presence or absence of spines in the chambers of the genital atrium) by studying morphologically optimal specimens (i.e., types or fresh collection containing uncontracted, relaxed and unfolded specimens) from confirmed hosts are necessary to validate the generic status of most of them (see [
32]). At present, only
Microcotyle multilineatus n. sp. has been described on a pomacentrid host from Mexico.


 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}