
GmGSTU23 Encoding a Tau Class Glutathione S-Transferase Protein Enhances the Salt Tolerance of Soybean (Glycine max L.)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
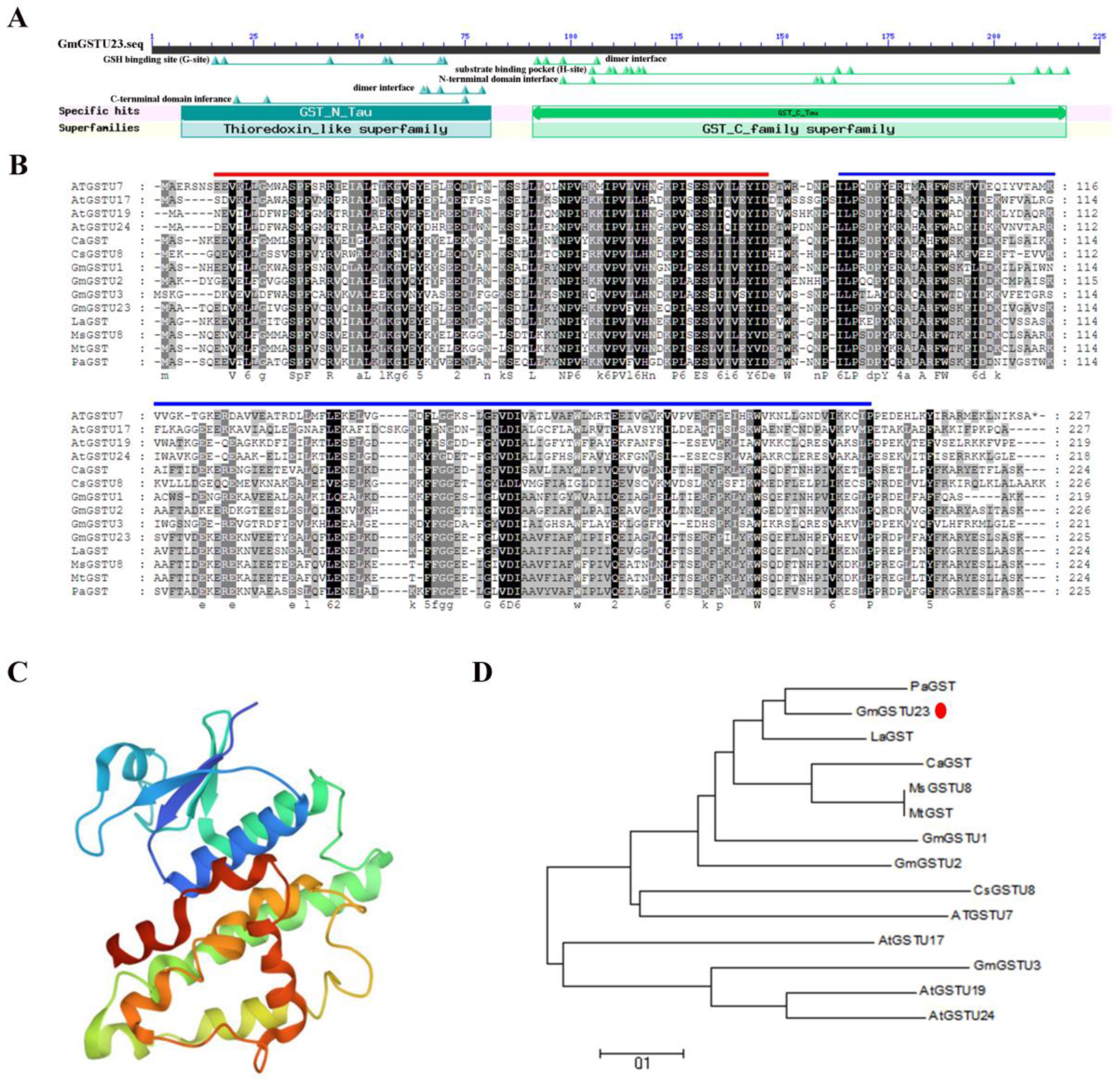
2.1. Isolation and Bioinformatics Analysis of the GmGSTU23
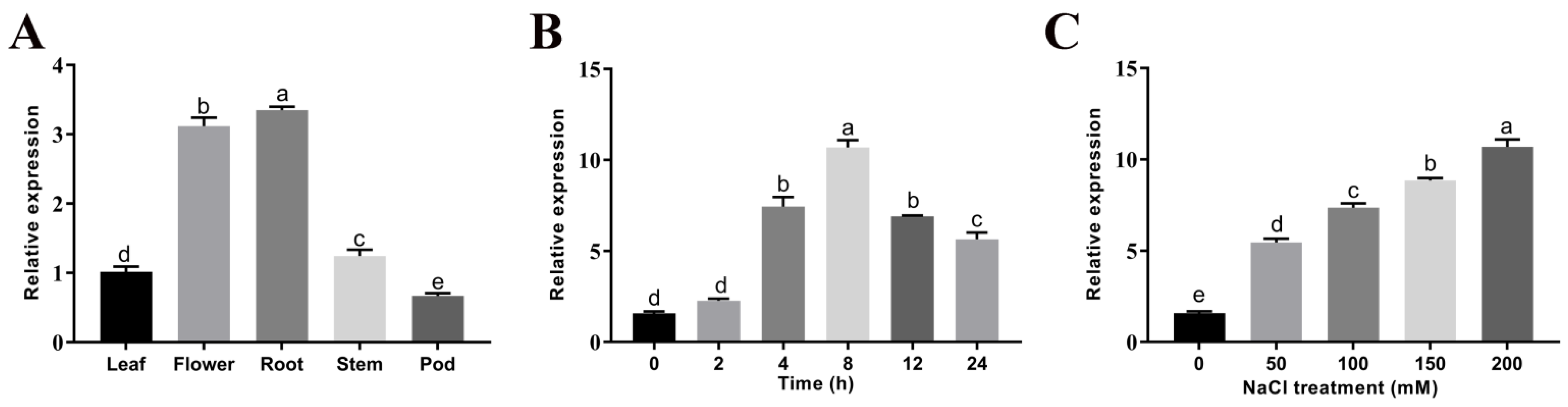
2.2. Expression Pattern Analysis of the GmGSTU23
2.3. Generation and Detection of the GmGSTU23 Overexpression Lines
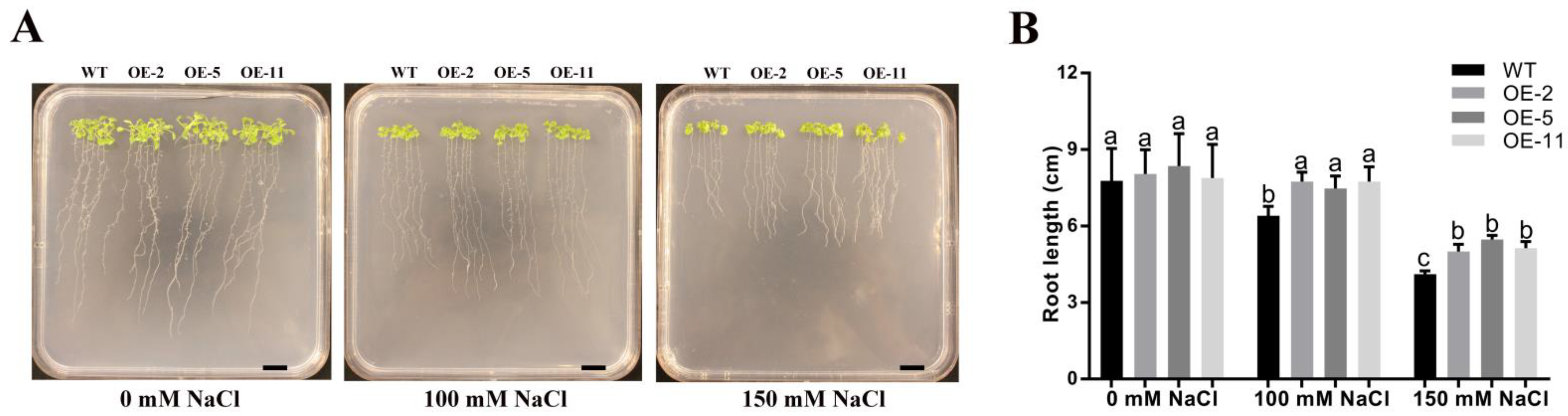
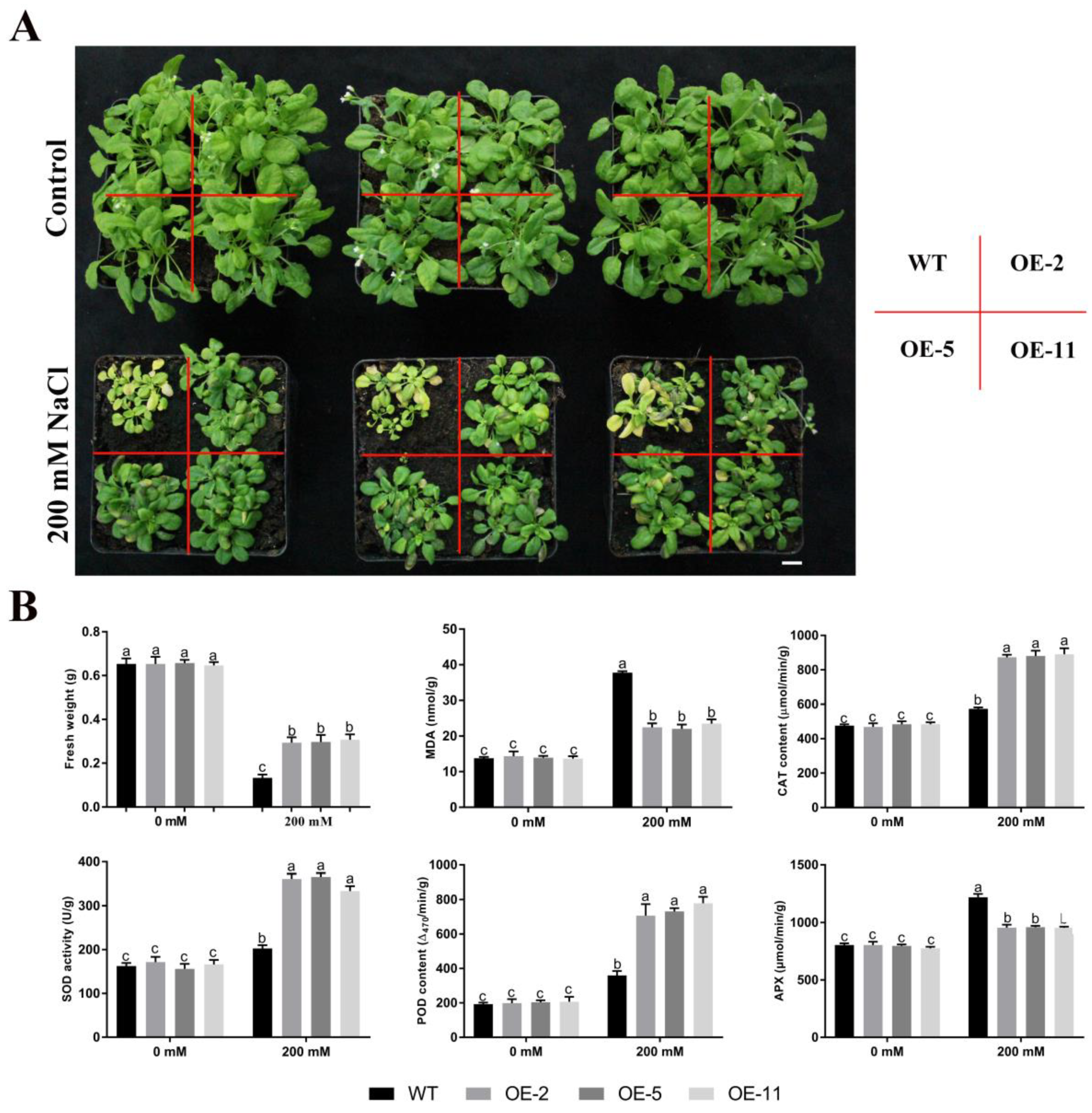
2.4. Overexpression of GmGSTU23 Confers Phenotypic Tolerance to Salt Stress
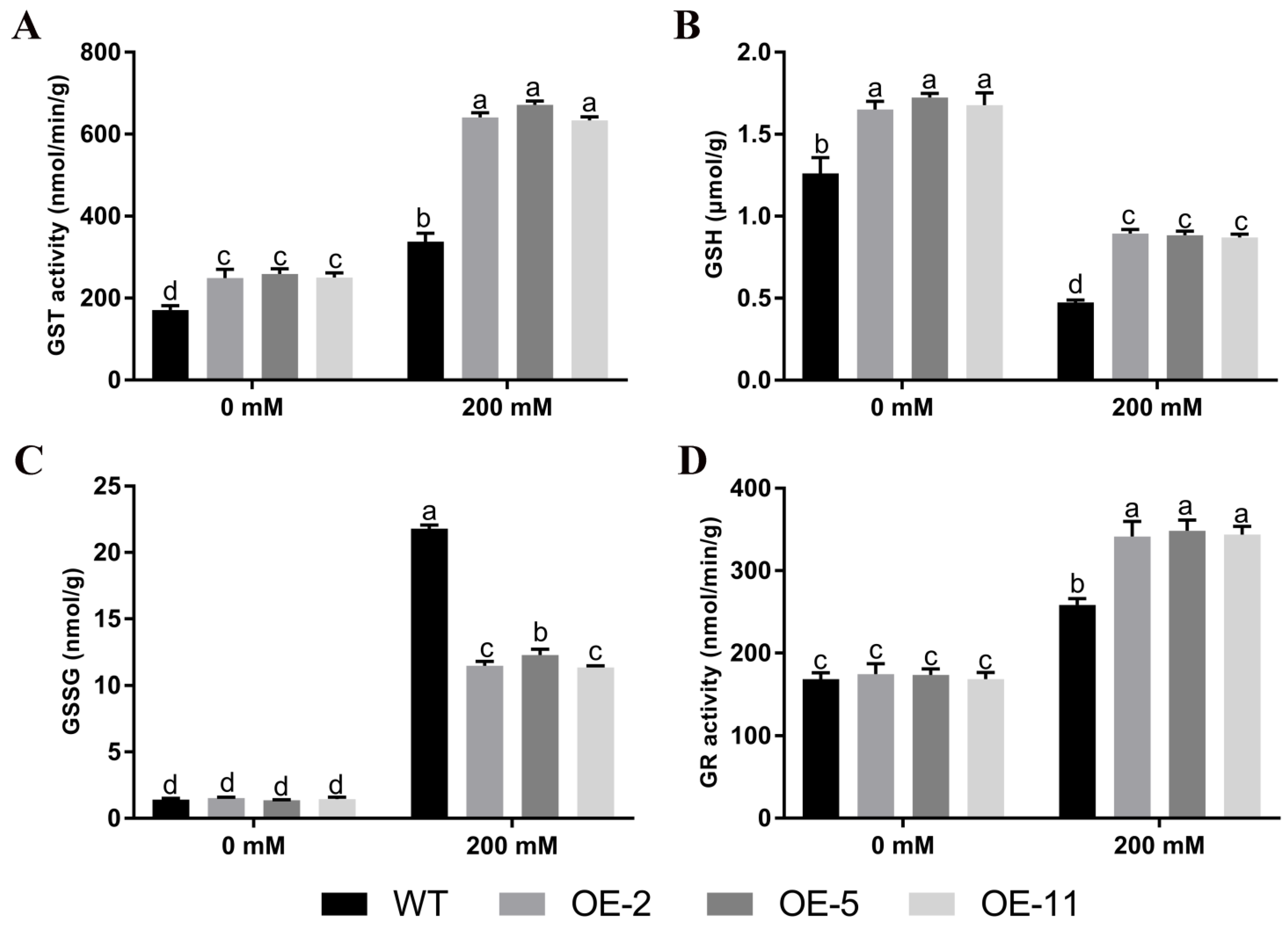
2.5. Changes of Glutathione Pool Composition and Related Enzymes under Salt Stress
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Analysis of Gene Expression Pattern
4.3. Cloning and Bioinformatics Analysis of GmGSTU23
4.4. Vector Construction and Transgenic Lines Generation
4.5. Phenotypes Analysis of GmGSTU23 Transgenic Lines
4.6. Physiological Analysis of GmGSTU23 Transgenic Lines
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, A. Soil salinization and waterlogging: A threat to environment and agricultural sustainability. Ecol. Indic. 2015, 57, 128–130. [Google Scholar] [CrossRef]
- Litalien, A.; Zeeb, B. Curing the earth: A review of anthropogenic soil salinization and plant-based strategies for sustainable mitigation. Sci. Total. Environ. 2020, 698, 134235. [Google Scholar] [CrossRef] [PubMed]
- Teh, S.Y.; Koh, H.L. Climate change and soil salinization: Impact on agriculture, water and food security. Int. J. Agric. For. Plant. 2016, 2, 1–9. [Google Scholar]
- Li, J.; Pu, L.; Han, M.; Zhu, M.; Zhang, R.; Xiang, Y. Soil salinization research in China: Advances and prospects. J. Geogr. Sci. 2014, 24, 943–960. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Shani, U.; Dudley, L.M. Field studies of crop response to water and salt stress. Soil Sci. Soc. Am. J. 2001, 65, 1522–1528. [Google Scholar] [CrossRef]
- van Zelm, E.; Zhang, Y.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Gorham, J.; Wyn Jones, R.G.; McDonnell, E. Some mechanisms of salt tolerance in crop plants. Biosalin. Action Bioprod. Saline Water 1985, 89, 15–40. [Google Scholar]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef]
- Muchate, N.S.; Nikalje, G.C.; Rajurkar, N.S.; Suprasanna, P.; Nikam, T.D. Plant salt stress: Adaptive responses, tolerance mechanism and bioengineering for salt tolerance. Bot. Rev. 2016, 82, 371–406. [Google Scholar] [CrossRef]
- Edwards, R.; Dixon, D.P.; Walbot, V. Plant glutathione S-transferases: Enzymes with multiple functions in sickness and in health. Trends Plant Sci. 2000, 5, 193–198. [Google Scholar] [CrossRef]
- Prasannath, K. Plant defense-related enzymes against pathogens: A review. AGRIEAST J. Agric. Sci. 2017, 11, 38–48. [Google Scholar] [CrossRef]
- Cunningham, F.X., Jr.; Gantt, E. Genes and enzymes of carotenoid biosynthesis in plants. Annu. Rev. Plant Biol. 1998, 49, 557. [Google Scholar] [CrossRef]
- Gullner, G.; Komives, T.; Király, L.; Schröder, P. Glutathione S-transferase enzymes in plant-pathogen interactions. Front. Plant Sci. 2018, 9, 1836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, S.; Rahman, I.A.; Islam, T.; Ghosh, A. Genome-wide identification and expression analysis of glutathione S-transferase gene family in tomato: Gaining an insight to their physiological and stress-specific roles. PLoS ONE 2017, 12, e0187504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, S.; Sajib, S.D.; Jui, Z.S.; Arabia, S.; Islam, T.; Ghosh, A. Genome-wide identification of glutathione S-transferase gene family in pepper, its classification, and expression profiling under different anatomical and environmental conditions. Sci. Rep. 2019, 9, 9101. [Google Scholar] [CrossRef] [Green Version]
- McGonigle, B.; Keeler, S.J.; Lau, S.M.C.; Koeppe, M.K.; O’Keefe, D.P. A genomics approach to the comprehensive analysis of the glutathione S-transferase gene family in soybean and maize. Plant Physiol. 2000, 124, 1105–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, U.; Edwards, R.; Dixon, D.P.; Mauch, F. Probing the diversity of the Arabidopsis glutathione S-transferase gene family. Plant Mol. Biol. 2002, 49, 515–532. [Google Scholar] [CrossRef]
- Soranzo, N.; Sari Gorla, M.; Mizzi, L.; Mauch, F. Organisation and structural evolution of the rice glutathione S-transferase gene family. Mol. Genet. Genom. 2004, 271, 511–521. [Google Scholar] [CrossRef]
- Thomas, P.D.; Ebert, D.; Muruganujan, A.; Mushayahama, T.; Albou, L.P.; Mi, H. PANTHER: Making genome-scale phylogenetics accessible to all. Protein Sci. 2022, 31, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Pickett, C.B.; Lu, A.Y.H. Glutathione S-transferases: Gene structure, regulation, and biological function. Annu. Rev. Biochem. 1989, 58, 743–764. [Google Scholar] [CrossRef] [PubMed]
- Droog, F. Plant glutathione S-transferases, a tale of theta and tau. J. Plant Growth Regul. 1997, 16, 95–107. [Google Scholar] [CrossRef]
- Marrs, K.A. The functions and regulation of glutathione S-transferases in plants. Annu. Rev. Plant Biol. 1996, 47, 127–158. [Google Scholar] [CrossRef]
- Vuilleumier, S. Bacterial glutathione S-transferases: What are they good for? J. Bacteriol. 1997, 179, 1431–1441. [Google Scholar] [CrossRef] [Green Version]
- Daniel, V. Glutathione S-transferases: Gene structure and regulation of expression. Crit. Rev. Biochem. Mol. Biol. 1993, 28, 173–207. [Google Scholar] [CrossRef] [PubMed]
- Wilce, M.C.J.; Parker, M.W. Structure and function of glutathione S-transferases. Biochim. Biophys. Acta BBA-Protein Struct. Mol. Enzymol. 1994, 1205, 1–18. [Google Scholar] [CrossRef]
- Dalton, D.A.; Boniface, C.; Turner, Z.; Lindahl, A.; Kim, H.J.; Jelinek, L.; Govindarajulu, M.; Finger, R.E.; Taylor, C.G. Physiological roles of glutathione s-transferases in soybean root nodules. Plant Physiol. 2009, 150, 521–530. [Google Scholar] [CrossRef] [Green Version]
- Becana, M.; Matamoros, M.A.; Udvardi, M.; Dalton, D.A. Recent insights into antioxidant defenses of legume root nodules. New Phytol. 2010, 188, 960–976. [Google Scholar] [CrossRef] [Green Version]
- Dalton, D.A.; Scott, J.; Jelinek, L. Characterization of Glutathione S-Transferases from Legume Root Nodules. In Biological Nitrogen Fixation: Towards Poverty Alleviation through Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2008; pp. 203–204. [Google Scholar]
- Wen, K.; Li, X.; Huang, R.; Nian, H. Application of exogenous glutathione decreases chromium translocation and alleviates its toxicity in soybean (Glycine max L.). Ecotoxicol. Environ. Saf. 2022, 234, 113405. [Google Scholar] [CrossRef]
- García-Garijo, A.; Tejera, N.A.; Lluch, C.; Palma, F. Metabolic responses in root nodules of Phaseolus vulgaris and Vicia sativa exposed to the imazamox herbicide. Pestic Biochem Physiol. 2014, 111, 19–23. [Google Scholar] [CrossRef]
- Neuefeind, T.; Reinemer, P.; Bieseler, B. Plant glutathione S-transferases and herbicide detoxification. Biol. Chem. 1997, 378, 199–205. [Google Scholar]
- DeRidder, B.P.; Dixon, D.P.; Beussman, D.J.; Edwards, R.; Goldsbrough, P.B. Induction of glutathione S-transferases in Arabidopsis by herbicide safeners. Plant Physiol. 2002, 130, 1497–1505. [Google Scholar] [CrossRef] [Green Version]
- Estévez, I.H.; Hernández, M.R. Plant glutathione S-transferases: An overview. Plant Gene 2020, 23, 100233. [Google Scholar] [CrossRef]
- Cao, Q.; Lv, W.; Jiang, H.; Chen, X.; Wang, X.; Wang, Y. Genome-wide identification of glutathione S-transferase gene family members in tea plant (Camellia sinensis) and their response to environmental stress. Int. J. Biol. Macromol. 2022, 205, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhang, D.; Chen, L.; Wang, J.; Zhang, W.H. Genome-wide analysis of the glutathione S-Transferase family in wild Medicago ruthenica and drought-tolerant breeding application of MruGSTU39 gene in cultivated alfalfa. Theor. Appl. Genet. 2022, 135, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Liu, Y.; Rao, J.; Wang, G.; Li, H.; Ge, F.; Chen, C. Overexpression of the glutathione S-transferase gene from Pyrus pyrifolia fruit improves tolerance to abiotic stress in transgenic tobacco plants. Mol. Biol. 2013, 47, 515–523. [Google Scholar] [CrossRef]
- Kumar, S.; Asif, M.H.; Chakrabarty, D.; Tripathi, R.D.; Dubey, R.S.; Trivedi, P.K. Expression of a rice Lambda class of glutathione S-transferase, OsGSTL2, in Arabidopsis provides tolerance to heavy metal and other abiotic stresses. J. Hazard. Mater. 2013, 248–249, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.I.; Noh, E.W.; Kim, H.J.; Shim, D. Overexpression of poplar GSTU51 confers selective tolerance to both mercury and methyl viologen but not to CDNB or cadmium in transgenic poplars. Plant Biotechnol. Rep. 2013, 7, 175–184. [Google Scholar] [CrossRef]
- Qi, Y.C.; Liu, W.Q.; Qiu, L.Y.; Zhang, S.M.; Ma, L.; Zhang, H. Overexpression of glutathione S-transferase gene increases salt tolerance of Arabidopsis. Russ. J. Plant Physiol. 2010, 57, 233–240. [Google Scholar] [CrossRef]
- Xu, J.; Xing, X.J.; Tian, Y.S.; Peng, R.H.; Xue, Y.; Zhao, W.; Yao, Q.H. Transgenic Arabidopsis Plants Expressing Tomato Glutathione S-Transferase Showed Enhanced Resistance to Salt and Drought Stress. PLoS ONE 2015, 10, e0136960. [Google Scholar] [CrossRef] [PubMed]
- Ji, W.; Zhu, Y.; Li, Y.; Yang, L.; Zhao, X.; Cai, H.; Bai, X. Over-expression of a glutathione S-transferase gene, GsGST, from wild soybean (Glycine soja) enhances drought and salt tolerance in transgenic tobacco. Biotechnol. Lett. 2010, 32, 1173–1179. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Yu, C.; Liu, R.; Li, X.; Huang, H.; Wang, X.; Zhang, C.; Jiang, N.; Li, X.; Cheng, S.; et al. The Glutathione S-Transferase PtGSTF1 Improves Biomass Production and Salt Tolerance through Regulating Xylem Cell Proliferation, Ion Homeostasis and Reactive Oxygen Species Scavenging in Poplar. Int. J. Mol. Sci. 2022, 23, 11288. [Google Scholar] [CrossRef]
- Zhang, Y.; He, J.; Xiao, Y.; Zhang, Y.; Liu, Y.; Wan, S.; Liu, L.; Dong, Y.; Liu, H.; Yu, Y. CsGSTU8, a Glutathione S-Transferase From Camellia sinensis, Is Regulated by CsWRKY48 and Plays a Positive Role in Drought Tolerance. Front. Plant Sci. 2021, 12, 795919. [Google Scholar] [CrossRef]
- Zhao, F.; Zhang, H. Salt and paraquat stress tolerance results from co-expression of the Suaeda salsa glutathione S-transferase and catalase in transgenic rice. Plant Cell Tissue Organ Cult. 2006, 86, 349–358. [Google Scholar] [CrossRef]
- Specht, J.E.; Diers, B.W.; Nelson, R.L.; de Toledo, J.F.F.; Torrion, J.A.; Grassini, P. Soybean. Yield Gains Major US Field Crop. 2014, 33, 311–355. [Google Scholar]
- List, G.R. Soybean lecithin: Food, industrial uses, and other applications. Polar Lipids 2015, 1–33. [Google Scholar] [CrossRef]
- Chen, K.I.; Erh, M.H.; Su, N.W.; Liu, W.H.; Chou, C.C.; Cheng, K.C. Soyfoods and soybean products: From traditional use to modern applications. Appl. Microbiol. Biotechnol. 2012, 96, 9–22. [Google Scholar] [CrossRef]
- Erdman, J.W., Jr.; Fordyce, E.J. Soy products and the human diet. Am. J. Clin. Nutr. 1989, 49, 725–737. [Google Scholar] [CrossRef]
- Essa, T.A. Effect of salinity stress on growth and nutrient composition of three soybean (Glycine max L. Merrill) cultivars. J. Agron. Crop Sci. 2002, 188, 86–93. [Google Scholar] [CrossRef]
- Phang, T.; Shao, G.; Lam, H. Salt tolerance in soybean. J. Integr. Plant Biol. 2008, 50, 1196–1212. [Google Scholar] [CrossRef] [PubMed]
- Ghassemi-Golezani, K.; Taifeh-Noori, M.; Oustan, S.; Moghaddam, M.; Seyyed-Rahmani, S. Oil and protein accumulation in soybean grains under salinity stress. Not. Sci. Biol. 2010, 2, 64–67. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Shen, Y.; Mu, K.; Cai, W.; Zhao, Y.; Shen, H.; Wang, X.; Ma, H. Phenylalanine Ammonia Lyase GmPAL1.1 Promotes Seed Vigor under High-Temperature and -Humidity Stress and Enhances Seed Germination under Salt and Drought Stress in Transgenic Arabidopsis. Plants 2022, 11, 3239. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Li, M.; Xie, M.; Liu, X.; Ni, M.; Shao, G.; Song, C.; Yim, A.K.-Y.; Tao, Y.; Wong, F.-L.; et al. Identification of a novel salt tolerance gene in wild soybean by whole-genome sequencing. Nat. Commun. 2014, 5, 4340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, R.; Qu, Y.; Guo, Y.; Yu, L.; Liu, Y.; Jiang, J.; Chen, J.; Ren, Y.; Liu, G.; Tian, L.; et al. Salinity tolerance in soybean is modulated by natural variation in GmSALT3. Plant J. 2014, 80, 937–950. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Chen, X.; Gu, H.; Wu, B.; Zhang, H.; Yuan, X.; Cui, X. GmHKT1; 4, a novel soybean gene regulating Na+/K+ ratio in roots enhances salt tolerance in transgenic plants. Plant Growth Regul. 2014, 73, 299–308. [Google Scholar] [CrossRef]
- Liao, Y.; Zou, H.F.; Wei, W.; Hao, Y.-J.; Tian, A.-G.; Huang, J.; Liu, Y.-F.; Zhang, J.-S.; Chen, S.-Y. Soybean GmbZIP44, GmbZIP62 and GmbZIP78 genes function as negative regulator of ABA signaling and confer salt and freezing tolerance in transgenic Arabidopsis. Planta 2008, 228, 225–240. [Google Scholar] [CrossRef]
- Chronopoulou, E.G.; Papageorgiou, A.C.; Ataya, F.; Nianiou-Obeidat, I.; Madesis, P.; Labrou, N.E. Expanding the Plant GSTome Through Directed Evolution: DNA Shuffling for the Generation of New Synthetic Enzymes with Engineered Catalytic and Binding Properties. Front. Plant Sci. 2018, 9, 1737. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.K.; Shabala, S. Mechanisms of Plant Responses and Adaptation to Soil Salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef]
- Berthomieu, P.; Conejero, G.; Nublat, A.; Brackenbury, W.J.; Lambert, C.; Savio, C.; Uozumi, N.; Oiki, S.; Yamada, K.; Cellier, F.; et al. Functional analysis of AtHKT1 in Arabidopsis shows that Na+ recirculation by the phloem is crucial for salt tolerance. EMBOJ 2003, 22, 2004–2014. [Google Scholar] [CrossRef] [Green Version]
- Alemán, F.; Caballero, F.; Ródenas, R.; Rivero, R.M.; Martínez, V.; Rubio, F. The F130S point mutation in the Arabidopsis high-affinity K+ transporter AtHAK5 increases K+ over Na+ and Cs+ selectivity and confers Na+ and Cs+ tolerance to yeast under heterologous expression. Front. Plant Sci. 2014, 5, 430. [Google Scholar] [CrossRef] [PubMed]
- Chai, C.; Subudhi, P.K. Comprehensive analysis and expression profiling of the OsLAX and OsABCB auxin transporter gene families in rice (Oryza sativa) under phytohormone stimuli and abiotic stresses. Front. Plant Sci. 2016, 7, 593. [Google Scholar] [CrossRef] [Green Version]
- Halfter, U.; Ishitani, M.; Zhu, J.K. The Arabidopsis SOS2 protein kinase physically interacts with and is activated by the calcium-binding protein SOS3. Proc. Natl. Acad. Sci. USA 2000, 97, 3735–3740. [Google Scholar] [CrossRef] [PubMed]
- Soma, F.; Mogami, J.; Yoshida, T.; Abekura, M.; Takahashi, F.; Kidokoro, S.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. ABA-unresponsive SnRK2 protein kinases regulate mRNA decay under osmotic stress in plants. Nat. Plants 2017, 3, 16204. [Google Scholar] [CrossRef]
- Wei, J.T.; Zhao, S.P.; Zhang, H.Y.; Jin, L.G.; Yu, T.F.; Zheng, L.; Ma, J.; Chen, J.; Zhou, Y.B.; Chen, M.; et al. GmDof41 regulated by the DREB1-type protein improves drought and salt tolerance by regulating the DREB2-type protein in soybean. Int. J. Biol. Macromol. 2023, 230, 123255. [Google Scholar] [CrossRef]
- Singh, D.; Debnath, P.; Sane, A.P.; Sane, V.A. Tomato (Solanum lycopersicum) WRKY23 enhances salt and osmotic stress tolerance by modulating the ethylene and auxin pathways in transgenic Arabidopsis. Plant Physiol. Biochem. 2023, 195, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Ugurlar, F.; Ashraf, M.; Alam, P.; Ahmad, P. Nitric oxide and hydrogen sulfide work together to improve tolerance to salinity stress in wheat plants by upraising the AsA-GSH cycle. Plant Physiol Biochem. 2023, 194, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Noctor, G. Oxidant and antioxidant signalling in plants: A re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ. 2010, 28, 1056–1071. [Google Scholar] [CrossRef]
- Hasan, M.K.; Liu, C.; Wang, F.; Ahammed, G.J.; Zhou, J.; Xu, M.X.; Yu, J.Q.; Xia, X.J. Glutathione-mediated regulation of nitric oxide, S-nitrosothiol and redox homeostasis confers cadmium tolerance by inducing transcription factors and stress response genes in tomato. Chemosphere 2016, 161, 536–545. [Google Scholar] [CrossRef]
- Vaish, S.; Parveen, R.; Gupta, D.; Basantani, M.K. Genome-wide identification and characterization of glutathione S-transferase gene family in Musa acuminata L. AAA group and gaining an insight to their role in banana fruit development. J. Appl. Genet. 2022, 63, 609–631. [Google Scholar] [CrossRef]
- He, G.; Guan, C.N.; Chen, Q.X.; Gou, X.J.; Liu, W.; Zeng, Q.Y.; Lan, T. Genome-Wide Analysis of the Glutathione S-Transferase Gene Family in Capsella rubella: Identification, Expression, and Biochemical Functions. Front. Plant Sci. 2016, 7, 1325. [Google Scholar] [CrossRef] [Green Version]
- Vaish, S.; Awasthi, P.; Tiwari, S.; Tiwari, S.K.; Gupta, D.; Basantani, M.K. In silico genome-wide identification and characterization of the glutathione S-transferase gene family in Vigna radiata. Genome 2018, 61, 311–322. [Google Scholar] [CrossRef] [Green Version]
- Jianan, Z.; Li, W.; Zhang, Y.; Song, W.; Jiang, H.; Zhao, J.; Zhan, Y.; Teng, W.; Qiu, L.; Zhao, X.; et al. Identification of glutathione transferase gene associated with partial resistance to Sclerotinia stem rot of soybean using genome-wide association and linkage mapping. Theor. Appl. Genet. 2021, 134, 2699–2709. [Google Scholar] [CrossRef] [PubMed]
- Horváth, E.; Bela, K.; Holinka, B.; Riyazuddin, R.; Gallé, Á.; Hajnal, Á.; Hurton, Á.; Fehér, A.; Csiszár, J. The Arabidopsis glutathione transferases, AtGSTF8 and AtGSTU19 are involved in the maintenance of root redox homeostasis affecting meristem size and salt stress sensitivity. Plant Sci. 2019, 283, 366–374. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.Y.; Kim, E.Y.; Kim, C.S.; Guh, J.O.; Kim, K.C.; Cho, B.H. Characterization of a glutathione S-transferase gene ATGST1 in Ara-bidopsis thaliana. Plant Cell Rep. 1998, 17, 700–704. [Google Scholar] [CrossRef]
- Sengupta, S.; Pehlivan, N.; Mangu, V.; Rajasekaran, K.; Baisakh, N. Characterization of a Stress-Enhanced Promoter from the Grass Halophyte, Spartina alterniflora L. Biology 2022, 11, 1828. [Google Scholar] [CrossRef]
- Zhang, X.; Xue, Y.; Wang, H.; Nisa, Z.U.; Jin, X.; Yu, L.; Liu, X.; Yu, Y.; Chen, C. Genome-wide identification and characterization of NHL gene family in response to alkaline stress, ABA and MEJA treatments in wild soybean (Glycine soja). PeerJ 2022, 10, e14451. [Google Scholar] [CrossRef] [PubMed]
- Czarnecka, E.; Nagao, R.T.; Key, J.L.; Gurley, W.B. Characterization of Gmhsp26-A, a stress gene encoding a divergent heat shock protein of soybean: Heavy-metal-induced inhibition of intron processing. Mol. Cell. Biol. 1988, 8, 1113–1122. [Google Scholar]
- Sarry, J.E.; Kuhn, L.; Ducruix, C.; Lafaye, A.; Junot, C.; Hugouvieux, V.; Jourdain, A.; Bastien, O.; Fievet, J.B.; Vailhen, D.; et al. The early responses of Arabidopsis thaliana cells to cadmium exposure explored by protein and metabolite profiling analyses. Proteomics 2006, 6, 2180–2198. [Google Scholar] [CrossRef] [PubMed]
- Libault, M.; Farmer, A.; Brechenmacher, L.; Drnevich, J.; Langley, R.J.; Bilgin, D.D.; Radwan, O.; Neece, D.J.; Clough, S.J.; May, G.D.; et al. Complete transcriptome of the soybean root hair cell, a single-cell model, and its alteration in response to Bradyrhizobium japonicum infection. Plant Physiol. 2010, 152, 541–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Severin, A.J.; Woody, J.L.; Bolon, Y.T.; Joseph, B.; Diers, B.W.; Farmer, A.D.; Muehlbauer, G.J.; Nelson, R.T.; Grant, D.; Specht, J.E.; et al. RNA-Seq Atlas of Glycine max: A guide to the soybean transcriptome. BMC Plant Biol. 2010, 10, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozfidan-Konakci, C.; Yildiztugay, E.; Arikan, B.; Alp-Turgut, F.N.; Turan, M.; Cavusoglu, H.; Sakalak, H. Responses of individual and combined polystyrene and polymethyl methacrylate nanoplastics on hormonal content, fluorescence/photochemistry of chlorophylls and ROS scavenging capacity in Lemna minor under arsenic-induced oxidative stress. Free Radic. Biol. Med. 2023, 196, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Zia-Ur-Rehman, M.; Anayatullah, S.; Irfan, E.; Hussain, S.M.; Rizwan, M.; Sohail, M.I.; Jafir, M.; Ahmad, T.; Usman, M.; Alharby, H.F. Nanoparticles assisted regulation of oxidative stress and antioxidant enzyme system in plants under salt stress: A review. Chemosphere 2023, 314, 137649. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Xian, P.; Wang, H.; Lin, R.; Lian, T.; Cheng, Y.; Ma, Q.; Nian, H. Transcription Factor GmWRKY142 Confers Cadmium Resistance by Up-Regulating the Cadmium Tolerance 1-Like Genes. Front. Plant Sci. 2020, 11, 724. [Google Scholar] [CrossRef] [PubMed]
- Hanway, J.J.; Thompson, H.E. How a soybean plant develops. Iowa State Univ. Sci. Technol. Coop. Ext. Serv. 1967. Available online: https://dr.lib.iastate.edu/handle/20.500.12876/90212 (accessed on 4 February 2023).
- Fehr, W.R.; Caviness, C.E. Stages of Soybean Development; Iowa State University of Science and Technology, Cooperative Extension Service: Ames, IA, USA, 1977. [Google Scholar]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Pang, Y.; Zhong, Y.; Cai, Z.; Ma, Q.; Wen, K.; Nian, H. GmGSTU23 Encoding a Tau Class Glutathione S-Transferase Protein Enhances the Salt Tolerance of Soybean (Glycine max L.). Int. J. Mol. Sci. 2023, 24, 5547. https://doi.org/10.3390/ijms24065547
Li X, Pang Y, Zhong Y, Cai Z, Ma Q, Wen K, Nian H. GmGSTU23 Encoding a Tau Class Glutathione S-Transferase Protein Enhances the Salt Tolerance of Soybean (Glycine max L.). International Journal of Molecular Sciences. 2023; 24(6):5547. https://doi.org/10.3390/ijms24065547
Chicago/Turabian StyleLi, Xingang, Yuanting Pang, Yiwang Zhong, Zhandong Cai, Qibin Ma, Ke Wen, and Hai Nian. 2023. "GmGSTU23 Encoding a Tau Class Glutathione S-Transferase Protein Enhances the Salt Tolerance of Soybean (Glycine max L.)" International Journal of Molecular Sciences 24, no. 6: 5547. https://doi.org/10.3390/ijms24065547