
SiMYBS3, Encoding a Setaria italica Heterosis-Related MYB Transcription Factor, Confers Drought Tolerance in Arabidopsis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
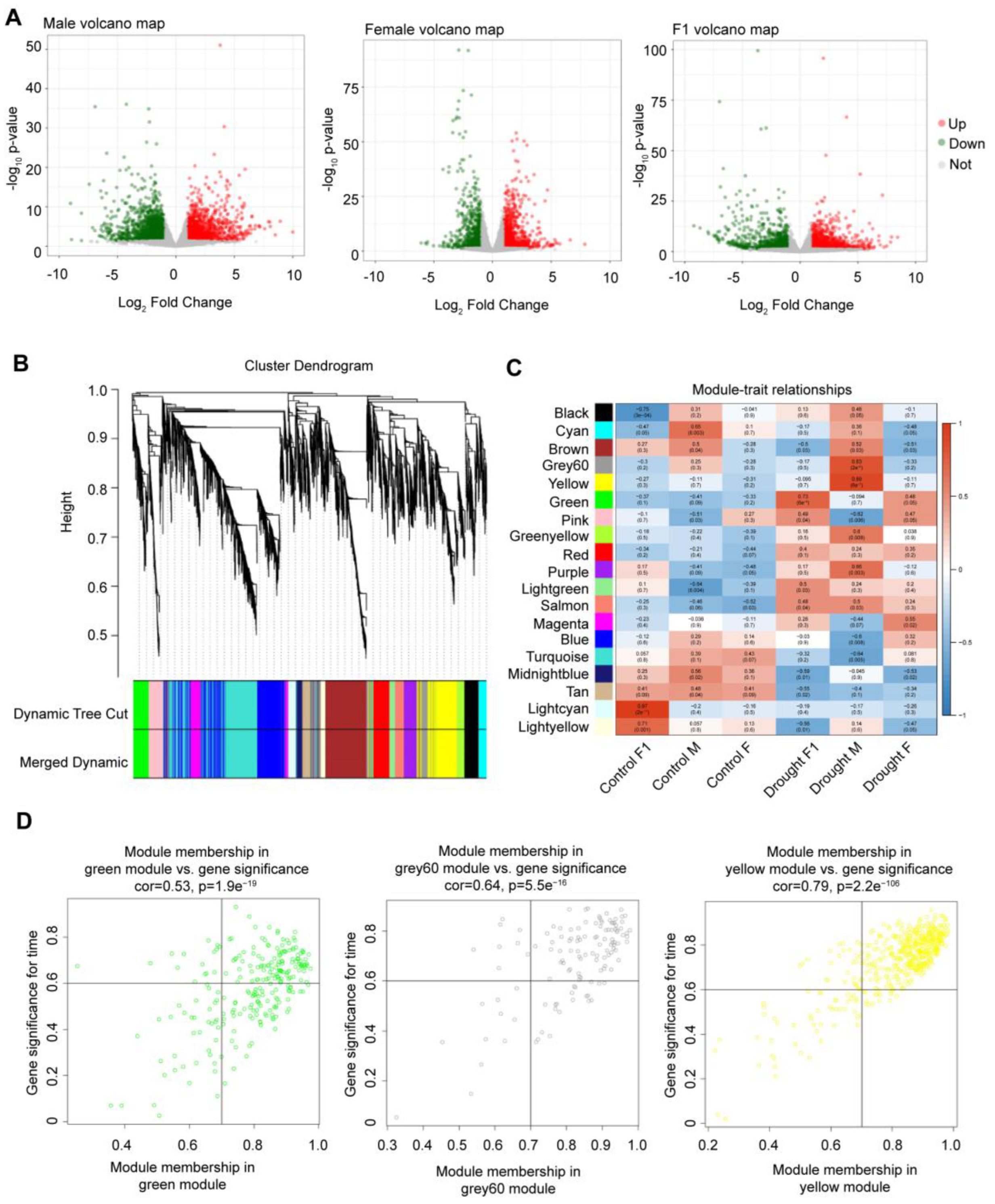
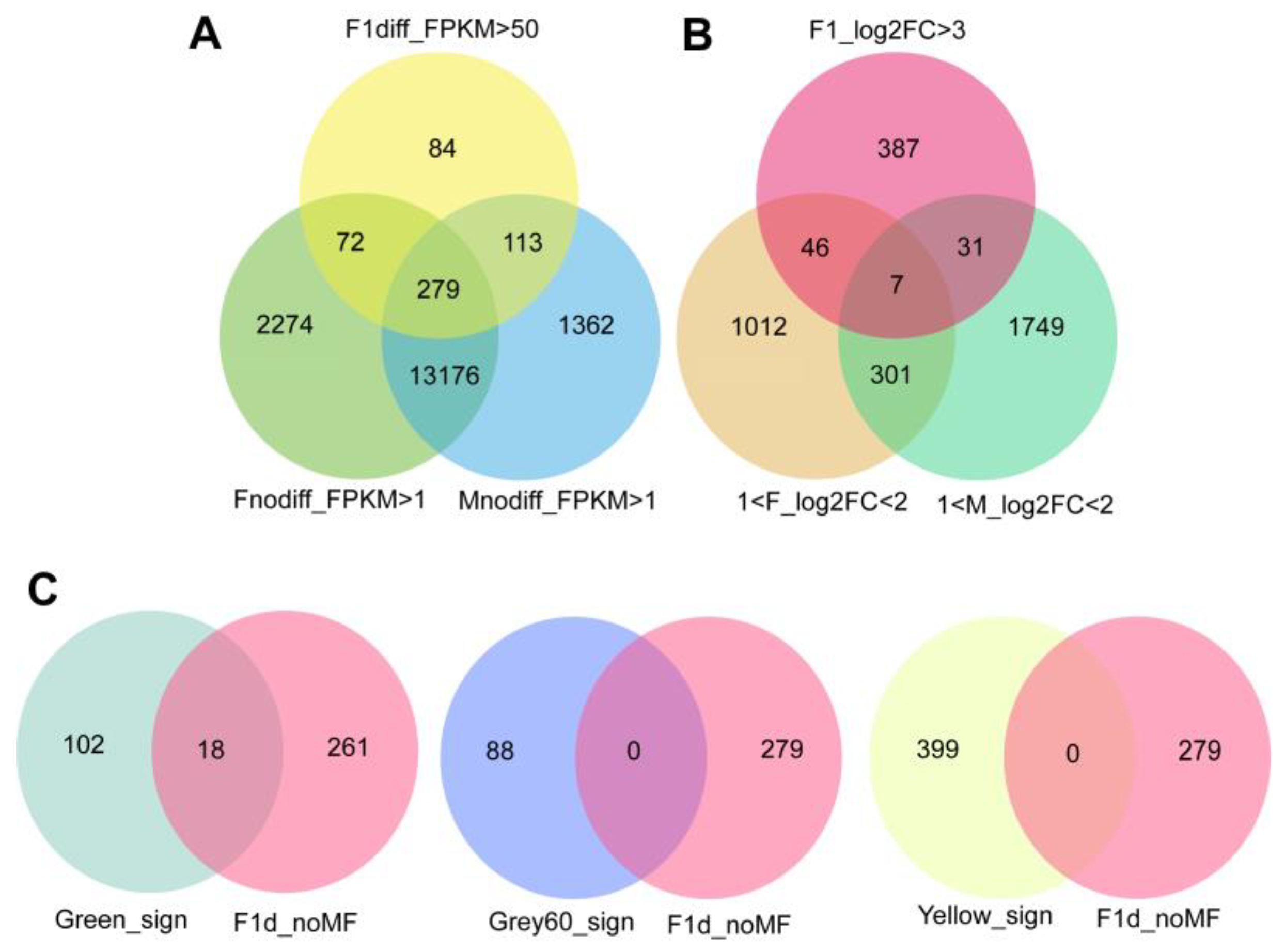
2.1. Identification of Drought-Resistant and Heterotic Genes
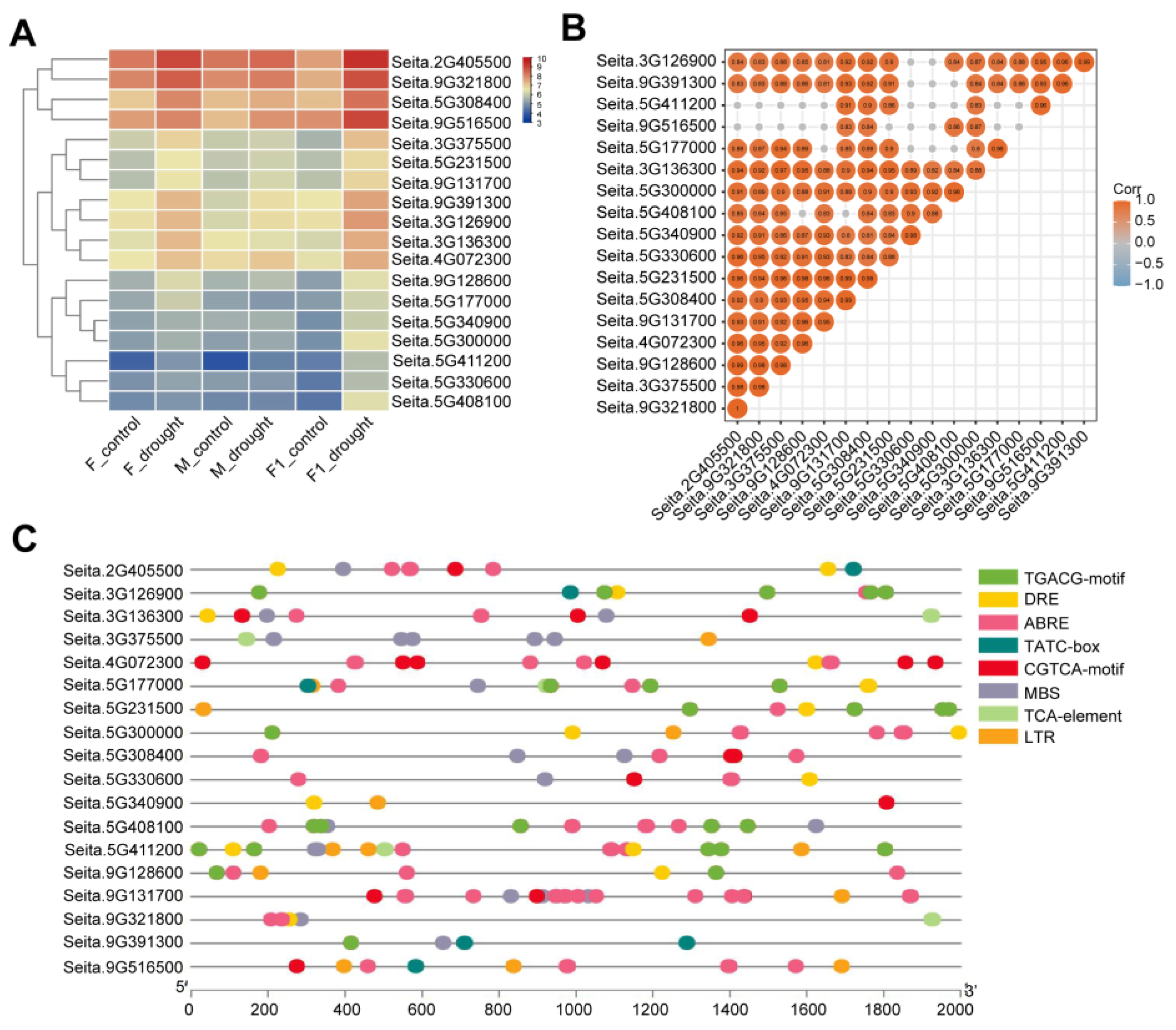
2.2. Analysis of Drought-Resistant and Heterotic Genes
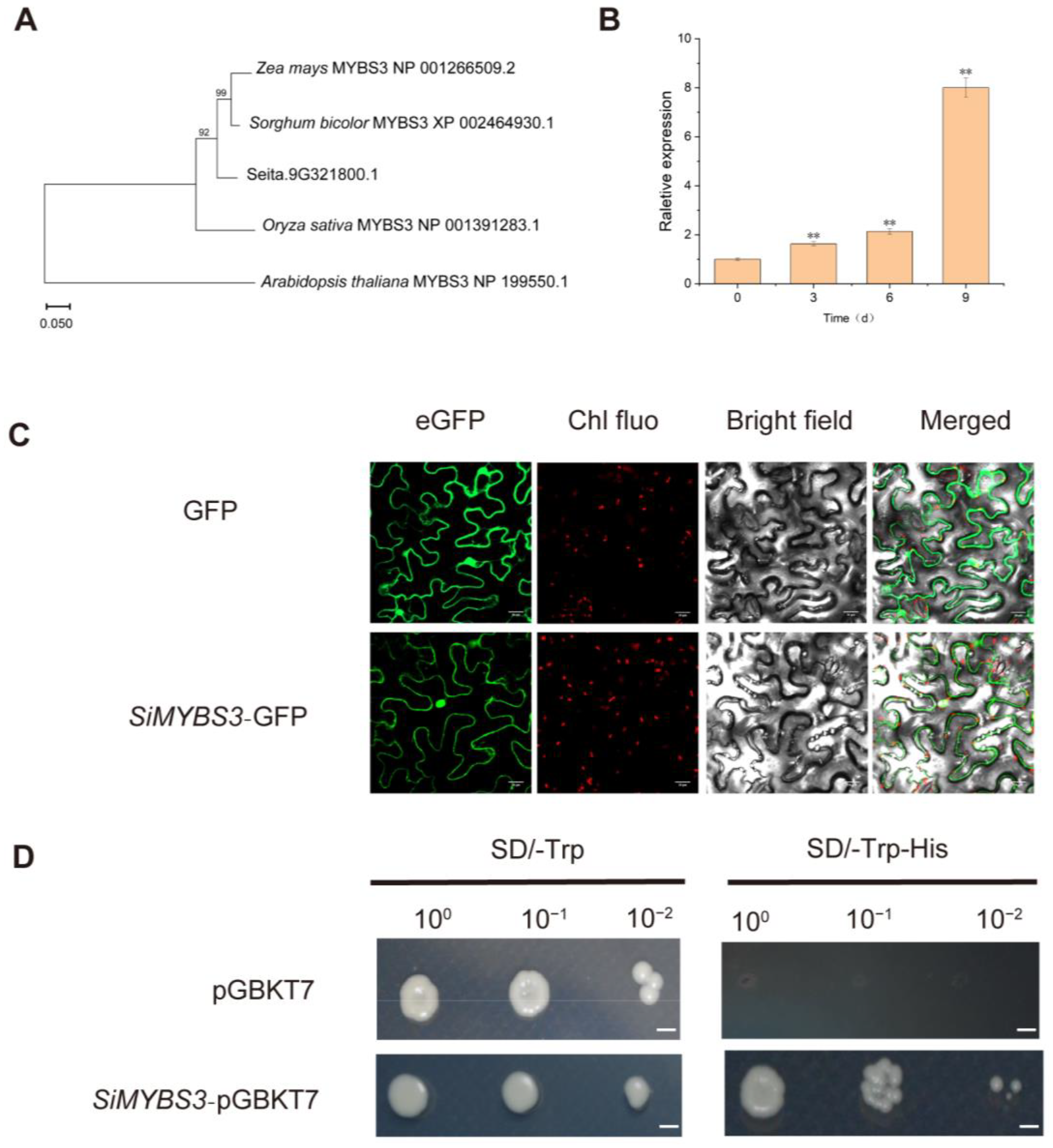
2.3. Phylogenetic Relationship, Expression Patterns, Subcellular Localization, and Transactivation Activity Analysis of SiMYBS3
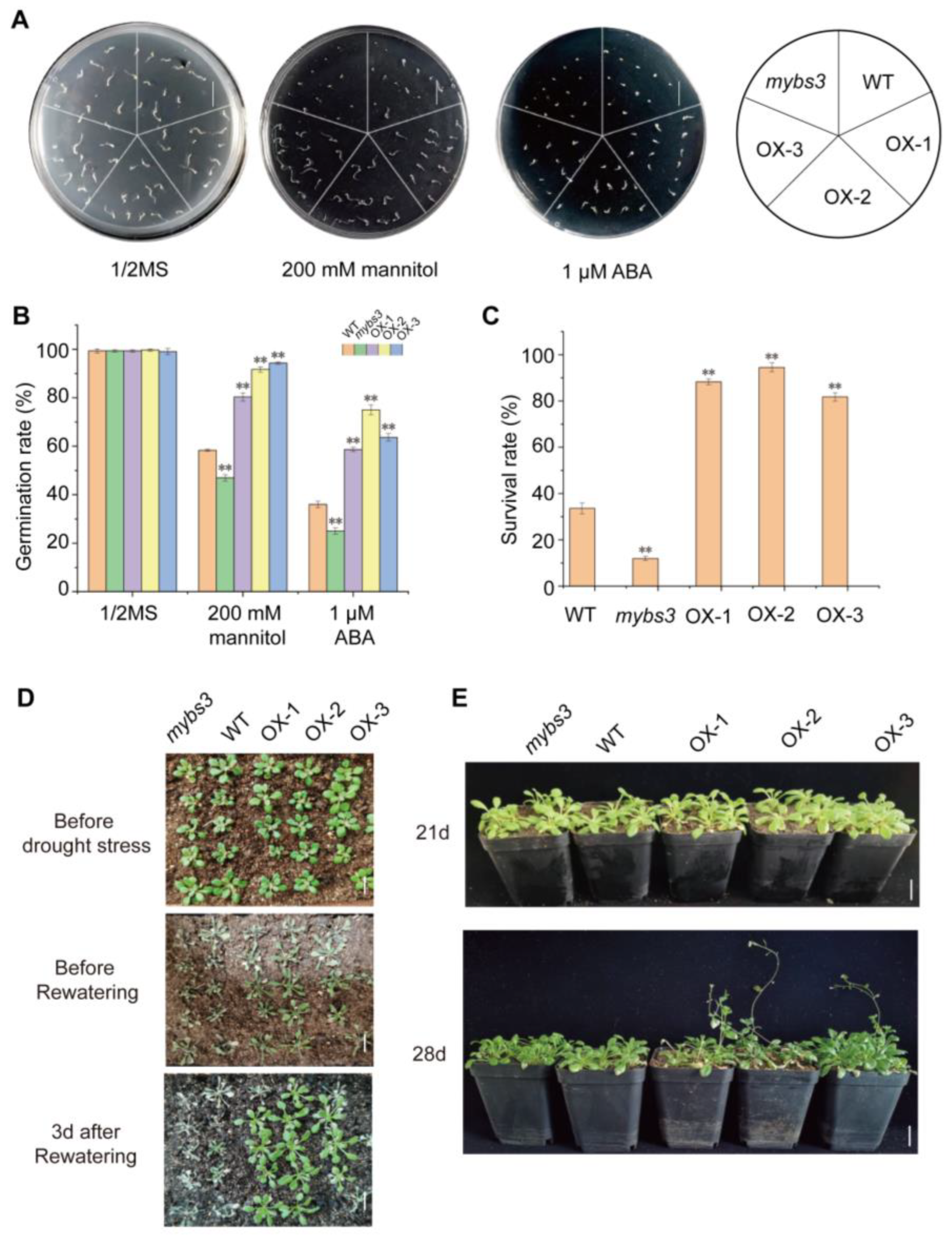
2.4. Overexpression of SiMYBS3 in Arabidopsis Led to Improved Drought Tolerance
3. Discussion
3.1. RNA-Seq Can Be Used to Mine Drought Resistance and Heterotic Genes
3.2. Functions of SiMYBS3
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. RNA Extraction, cDNA Synthesis, and RNA-Seq
4.3. WGCNA Analysis and Identification of DEGs
4.4. Cis-Acting Elements and Phylogenetic Tree Analysis
4.5. Real-Time Quantitative PCR
4.6. Subcellular Localization and Transcriptional Activity Analysis
4.7. Phenotypic Observations in Arabidopsis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Birchler, J.A.; Yao, H.; Chudalayandi, S.; Vaiman, D.; Veitia, R.A. Heterosis. Plant Cell 2010, 22, 2105–2112. [Google Scholar] [CrossRef] [Green Version]
- Govindaraju, D.R. An elucidation of over a century old enigma in genetics-Heterosis. PLoS Biol. 2019, 17, e3000215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lippman, Z.B.; Zamir, D. Heterosis: Revisiting the magic. Trends Genet 2007, 23, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, L.L.; Xing, F.; Kudrna, D.A.; Yao, W.; Copetti, D.; Mu, T.; Li, W.; Song, J.M.; Xie, W.; et al. Extensive sequence divergence between the reference genomes of two elite indica rice varieties Zhenshan 97 and Minghui 63. Proc. Natl. Acad. Sci. USA 2016, 113, 5163–5171. [Google Scholar] [CrossRef] [Green Version]
- Marcon, C.; Paschold, A.; Malik, W.A.; Lithio, A.; Baldauf, J.A.; Altrogge, L.; Opitz, N.; Lanz, C.; Schoof, H.; Nettleton, D.; et al. Stability of single-parent gene expression complementation in maize hybrids upon water deficit stress. Plant Physiol. 2017, 173, 1247–1257. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ma, C.; Wang, X.; Wu, M.; Shao, J.; Huang, L.; Yuan, L.; Fu, Z.; Li, W.; Zhang, X.; et al. Global transcriptional profiling between inbred parents and hybrids provides comprehensive insights into ear-length heterosis of maize (Zea mays). BMC Plant Biol. 2021, 21, 118. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Yao, X.; Mi, D.; Li, Z.; Yang, B.; Zheng, Y.; Qi, Y.; Guo, J. Comparative transcriptome analysis reveals the biocontrol mechanism of Bacillus velezensis F21 against Fusarium wilt on watermelon. Front. Microbiol. 2019, 10, 652. [Google Scholar] [CrossRef]
- Kong, X.; Chen, L.; Wei, T.; Zhou, H.; Bai, C.; Yan, X.; Miao, Z.; Xie, J.; Zhang, L. Transcriptome analysis of biological pathways associated with heterosis in Chinese cabbage. Genomics 2020, 112, 4732–4741. [Google Scholar] [CrossRef]
- Shalby, N.; Mohamed, I.A.A.; Xiong, J.; Hu, K.; Yang, Y.; Nishawy, E.; Yi, B.; Wen, J.; Ma, C.; Shen, J.; et al. Overdominance at the gene expression level plays a critical role in the hybrid root growth of Brassica napus. Int. J. Mol. Sci. 2021, 22, 9246. [Google Scholar] [CrossRef]
- Shu, H.Y.; Zhou, H.; Mu, H.L.; Wu, S.H.; Jiang, Y.L.; Yang, Z.; Hao, Y.Y.; Zhu, J.; Bao, W.L.; Cheng, S.H.; et al. Integrated analysis of mRNA and non-coding RNA transcriptome in pepper (Capsicum chinense) hybrid at seedling and flowering stages. Front. Genet. 2021, 12, 685788. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, C.; He, Y.; Zong, J.; Yang, X.; Si, H.; Sun, Z.; Hu, J.; Liang, W.; Zhang, D. Mutation in CSA creates a new photoperiod-sensitive genic male sterile line applicable for hybrid rice seed production. Proc. Natl. Acad. Sci. USA 2013, 110, 76–81. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Liu, J.; Ma, Q.; Qin, Y.; Wang, H.; Chen, P.; Ma, L.; Fu, X.; Zhu, L.; Wei, H.; et al. Deficiencies in the formation and regulation of anther cuticle and tryphine contribute to male sterility in cotton PGMS line. BMC Genom. 2020, 21, 825. [Google Scholar] [CrossRef]
- Hashimoto, S.; Wake, T.; Nakamura, H.; Minamiyama, M.; Araki-Nakamura, S.; Ohmae-Shinohara, K.; Koketsu, E.; Okamura, S.; Miura, K.; Kawaguchi, H.; et al. The dominance model for heterosis explains culm length genetics in a hybrid sorghum variety. Sci. Rep. 2021, 11, 4532. [Google Scholar] [CrossRef]
- Ju, C.; Zhang, F.; Gao, Y.; Zhang, W.; Yan, J.; Dai, J.; Li, J. Cloning, chromosome mapping and expression analysis of an R2R3-MYB gene under-expressed in maize hybrid. Mol. Biol. Rep. 2006, 33, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Quan, R.; Chen, G.; Yu, G.; Li, X.; Han, Z.; Wu, W.W.; Li, G.R.; Shi, J.L.; Li, B. An R2R3-MYB transcription factor VyMYB24, isolated from wild grape Vitis yanshanesis J. X. Chen., regulates the plant development and confers the tolerance to drought. Front. Plant Sci. 2022, 13, 966641. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Chen, T.; Kan, J.; Yao, Y.; Guo, D.; Yang, Y.; Ling, X.; Wang, J.; Zhang, B. The GhMYB36 transcription factor confers resistance to biotic and abiotic stress by enhancing PR1 gene expression in plants. Plant Biotechnol. J. 2022, 20, 722–735. [Google Scholar] [CrossRef]
- Song, Q.; Kong, L.; Yang, X.; Jiao, B.; Hu, J.; Zhang, Z.; Xu, C.; Luo, K. PtoMYB142, a poplar R2R3-MYB transcription factor, contributes to drought tolerance by regulating wax biosynthesis. Tree Physiol. 2022, 42, 2133–2147. [Google Scholar] [CrossRef]
- Yao, C.; Li, W.; Liang, X.; Ren, C.; Liu, W.; Yang, G.; Zhao, M.; Yang, T.; Li, X.; Han, D. Molecular cloning and characterization of MbMYB108, a Malus baccata MYB transcription factor gene, with functions in tolerance to cold and drought stress in transgenic Arabidopsis thaliana. Int. J. Mol. Sci. 2022, 23, 4846. [Google Scholar] [CrossRef]
- Su, C.F.; Wang, Y.C.; Hsieh, T.H.; Lu, C.A.; Tseng, T.H.; Yu, S.M. A novel MYBS3-dependent pathway confers cold tolerance in rice. Plant Physiol. 2010, 153, 145–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, L.; Newsome, S.D.; Chen, F.H.; Wang, H.; Guilderson, T.P.; Bettinger, R.L. Agricultural origins and the isotopic identity of domestication in northern China. Proc. Natl. Acad. Sci. USA 2009, 106, 5523–5528. [Google Scholar] [CrossRef] [Green Version]
- Pant, S.R.; Irigoyen, S.; Doust, A.N.; Scholthof, K.B.; Mandadi, K.K. Setaria: A food crop and translational research model for C4 grasses. Front. Plant Sci. 2016, 7, 1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muthamilarasan, M.; Prasad, M. Advances in Setaria genomics for genetic improvement of cereals and bioenergy grasses. Theor. Appl. Genet. 2015, 128, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.P.; Cheng, J.Y.; Wen, X.J.; Wang, J.X.; Shi, G.Y.; Yao, J.Y.; Hou, L.Y.; Sun, Q.; Xiang, P.; Yuan, X.Y.; et al. Transcriptomic studies reveal a key metabolic pathway contributing to a well-maintained photosynthetic system under drought stress in foxtail millet (Setaria italica L.). PeerJ 2018, 6, 4752. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Zhang, H.; Li, X.; Shen, H.; Gao, J.; Hou, S.; Zhang, B.; Mayes, S.; Bennett, M.; Ma, J.; et al. A mini foxtail millet with an Arabidopsis-like life cycle as a C4 model system. Nat. Plants 2020, 6, 1167–1178. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.L.; Shi, G.L.; Zhang, X.L.; Zhao, Z.H.; Wang, X.M. Effect of plant pattern and density on yield of hybrid millet ‘ZZG 19’ in ordos region. J. Agric. 2020, 10, 5. [Google Scholar]
- Chen, L.; Bian, J.; Shi, S.; Yu, J.; Khanzada, H.; Wassan, G.M.; Zhu, C.; Luo, X.; Tong, S.; Yang, X.; et al. Genetic analysis for the grain number heterosis of a super-hybrid rice WFYT025 combination using RNA-Seq. Rice 2018, 11, 37. [Google Scholar] [CrossRef] [Green Version]
- Swanson-Wagner, R.A.; Jia, Y.; DeCook, R.; Borsuk, L.A.; Nettleton, D.; Schnable, P.S. All possible modes of gene action are observed in a global comparison of gene expression in a maize F1 hybrid and its inbred parents. Proc. Natl. Acad. Sci. USA 2006, 103, 6805–6810. [Google Scholar] [CrossRef] [Green Version]
- Paschold, A.; Jia, Y.; Marcon, C.; Lund, S.; Larson, N.B.; Yeh, C.T.; Ossowski, S.; Lanz, C.; Nettleton, D.; Schnable, P.S.; et al. Complementation contributes to transcriptome complexity in maize (Zea mays L.) hybrids relative to their inbred parents. Genome Res. 2012, 22, 2445–2454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehraj, H.; Kawanabe, T.; Shimizu, M.; Miyaji, N.; Akter, A.; Dennis, E.S.; Fujimoto, R. In Arabidopsis thaliana heterosis level varies among individuals in an F1 hybrid population. Plants 2020, 9, 414. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.L.; Sun, X.M.; Zhang, S.G. Mechanism on differential gene expression and heterosis formation. Hereditas 2013, 35, 714–726. [Google Scholar] [CrossRef]
- Kwon, Y.; Kim, J.H.; Nguyen, H.N.; Jikumaru, Y.; Kamiya, Y.; Hong, S.W.; Lee, H. A novel Arabidopsis MYB-like transcription factor, MYBH, regulates hypocotyl elongation by enhancing auxin accumulation. J. Exp. Bot. 2013, 64, 3911–3922. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Yang, X.; He, K.; Liu, M.; Li, J.; Gao, Z.; Lin, Z.Q.; Zhang, Y.F.; Wang, X.X.; Qiu, X.M.; et al. The MYB transcription factor superfamily of Arabidopsis: Expression analysis and phylogenetic comparison with the rice MYB family. Plant Mol. Biol. 2006, 60, 107–124. [Google Scholar]
- Zhang, Y.; Liang, W.; Shi, J.; Xu, J.; Zhang, D. MYB56 encoding a R2R3 MYB transcription factor regulates seed size in Arabidopsis thaliana. J. Integr. Plant Biol. 2013, 55, 1166–1178. [Google Scholar] [CrossRef]
- Wan, J.; Wang, R.; Zhang, P.; Sun, L.; Ju, Q.; Huang, H.; Lü, S.; Tran, L.S.; Xu, J. MYB70 modulates seed germination and root system development in Arabidopsis. iScience 2021, 24, 103228. [Google Scholar] [CrossRef]
- Wang, X.M.; Wu, R.; Shen, T.S.; Li, Z.A.; Li, C.; Wu, B.K.; Jiang, H.Y.; Zhao, G.W. An R2R3-MYB transcription factor OsMYBAS1 promotes seed germination under different sowing depths in transgenic rice. Plants 2022, 11, 139. [Google Scholar] [CrossRef]
- Ren, D.; Rao, Y.; Yu, H.; Xu, Q.; Cui, Y.; Xia, S.; Yu, X.; Liu, H.; Hu, H.; Xue, D.; et al. MORE FLORET1 encodes a MYB transcription factor that regulates spikelet development in rice. Plant Physiol. 2020, 184, 251–265. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Dou, Y.; Li, M.; Qu, P.; He, Z.; Liu, Y.; Xu, Z.; Chen, J.; Chen, M.; Ma, Y. SiMYB3 in foxtail millet (Setaria italica) confers tolerance to low-nitrogen stress by regulating root growth in transgenic plants. Int. J. Mol. Sci. 2019, 20, 5741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Luo, M.; Sun, X.; Yan, J.; Shi, H.; Yan, H.; Yan, R.; Wang, S.; Tang, W.; Zhou, Y.; et al. SiMYB19 from foxtail millet (Setaria italica) confers transgenic rice tolerance to high salt stress in the field. Int. J. Mol. Sci. 2022, 23, 756. [Google Scholar] [CrossRef]
- Xu, W.; Tang, W.; Wang, C.; Ge, L.; Sun, J.; Qi, X.; He, Z.; Zhou, Y.; Chen, J.; Xu, Z.; et al. SiMYB56 confers drought stress tolerance in transgenic rice by regulating lignin biosynthesis and ABA signaling pathway. Front. Plant Sci. 2020, 11, 785. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhou, X.T.; Dai, K.L.; Yuan, X.Y.; Guo, P.Y.; Shi, W.P.; Zhou, M.X. Comprehensive analysis of YABBY gene family in foxtail millet (Setaria italica) and functional characterization of SiDL. J. Integr. Agric. 2022, 21, 2876–2887. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Li, H.; Handsakerm, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. The Subread aligner: Fast, accurate and scalable read mapping by seed-and-vote. Nucleic Acids Res. 2013, 41, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Rubio, V. ggplot2—Elegant graphics for data analysis (2nd Edition). J. Stat. Softw. 2017, 77, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Zhang, S.; Sun, M.; Guo, Y.; Zhao, S.; Zhou, X.; Bai, X.; Dai, K.; Li, H.; Yuan, X.; et al. SiMYBS3, Encoding a Setaria italica Heterosis-Related MYB Transcription Factor, Confers Drought Tolerance in Arabidopsis. Int. J. Mol. Sci. 2023, 24, 5418. https://doi.org/10.3390/ijms24065418
Liu X, Zhang S, Sun M, Guo Y, Zhao S, Zhou X, Bai X, Dai K, Li H, Yuan X, et al. SiMYBS3, Encoding a Setaria italica Heterosis-Related MYB Transcription Factor, Confers Drought Tolerance in Arabidopsis. International Journal of Molecular Sciences. 2023; 24(6):5418. https://doi.org/10.3390/ijms24065418
Chicago/Turabian StyleLiu, Xin, Shuai Zhang, Mengbo Sun, Yurong Guo, Shaoxing Zhao, Xutao Zhou, Xionghui Bai, Keli Dai, Huixia Li, Xiangyang Yuan, and et al. 2023. "SiMYBS3, Encoding a Setaria italica Heterosis-Related MYB Transcription Factor, Confers Drought Tolerance in Arabidopsis" International Journal of Molecular Sciences 24, no. 6: 5418. https://doi.org/10.3390/ijms24065418