
Co-Transplantation of Barcoded Lymphoid-Primed Multipotent (LMPP) and Common Lymphocyte (CLP) Progenitors Reveals a Major Contribution of LMPP to the Lymphoid Lineage
 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
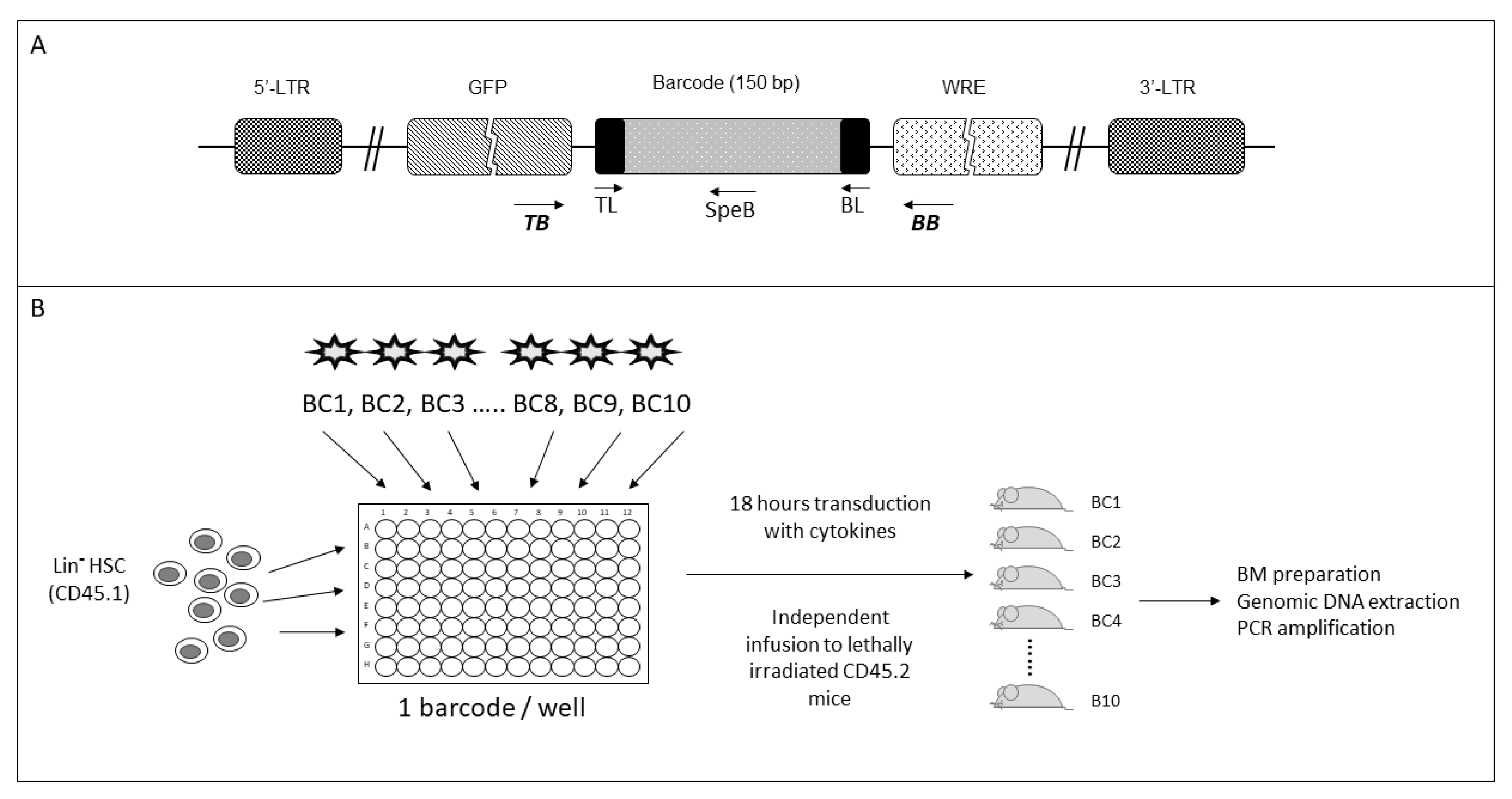
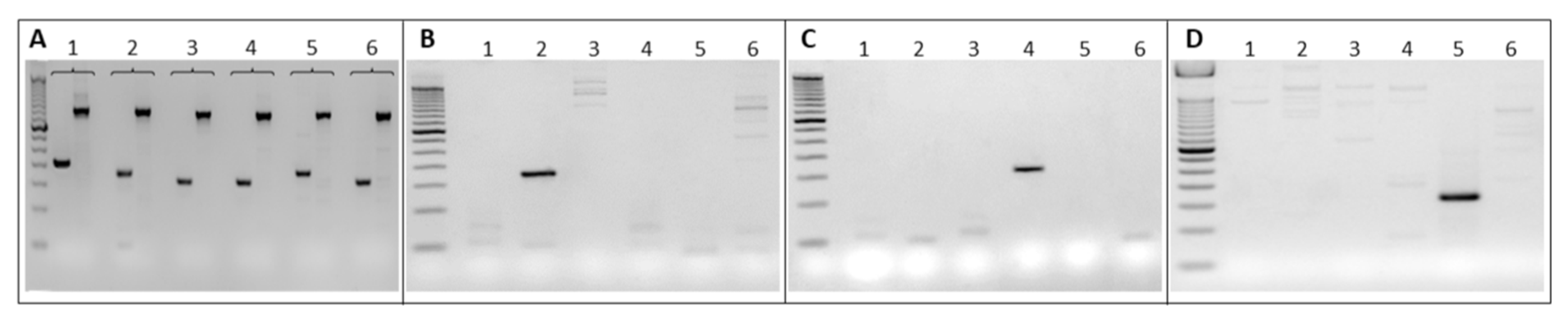
2.1. Selection of Barcodes for Hematopoietic Progenitor Labeling
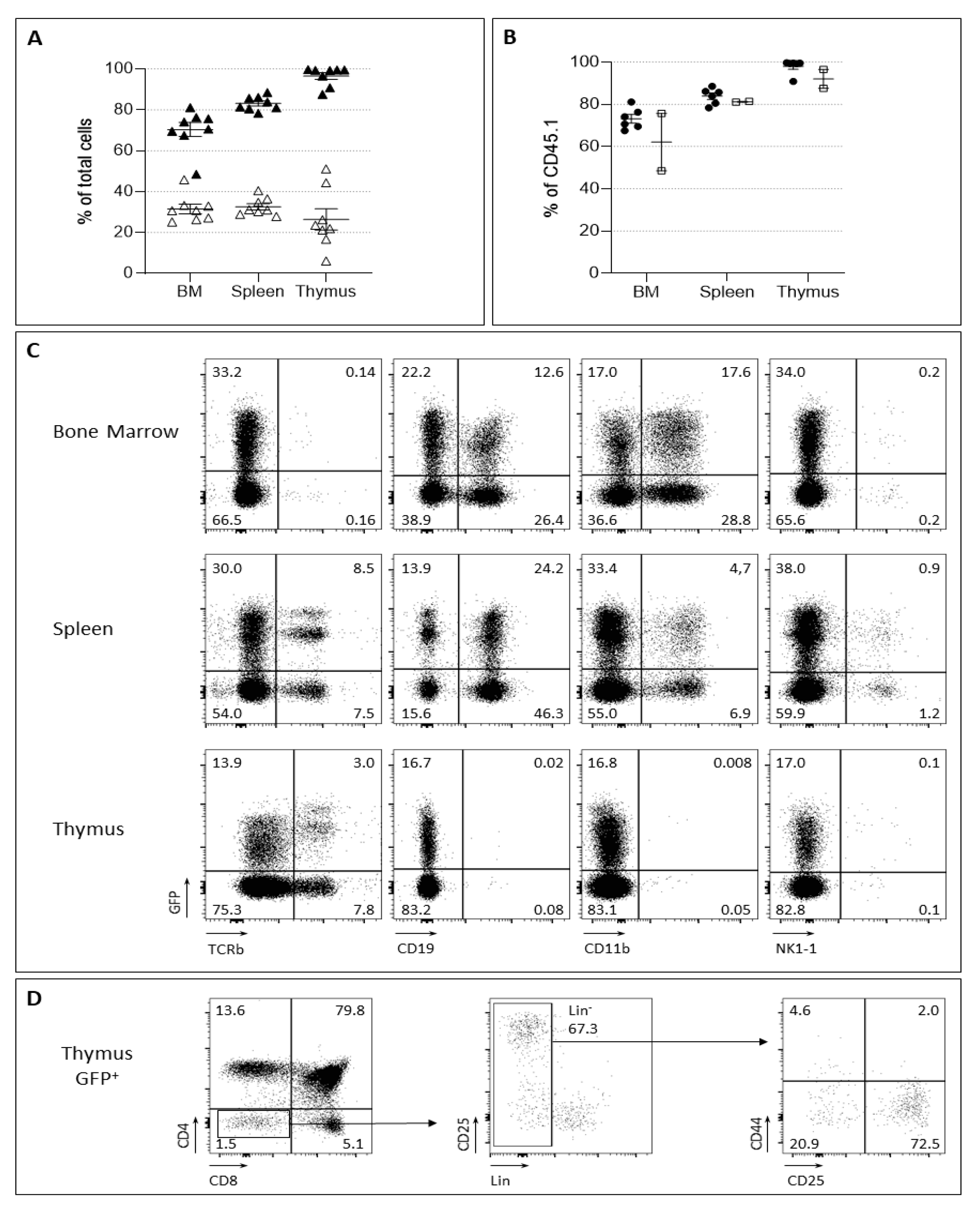
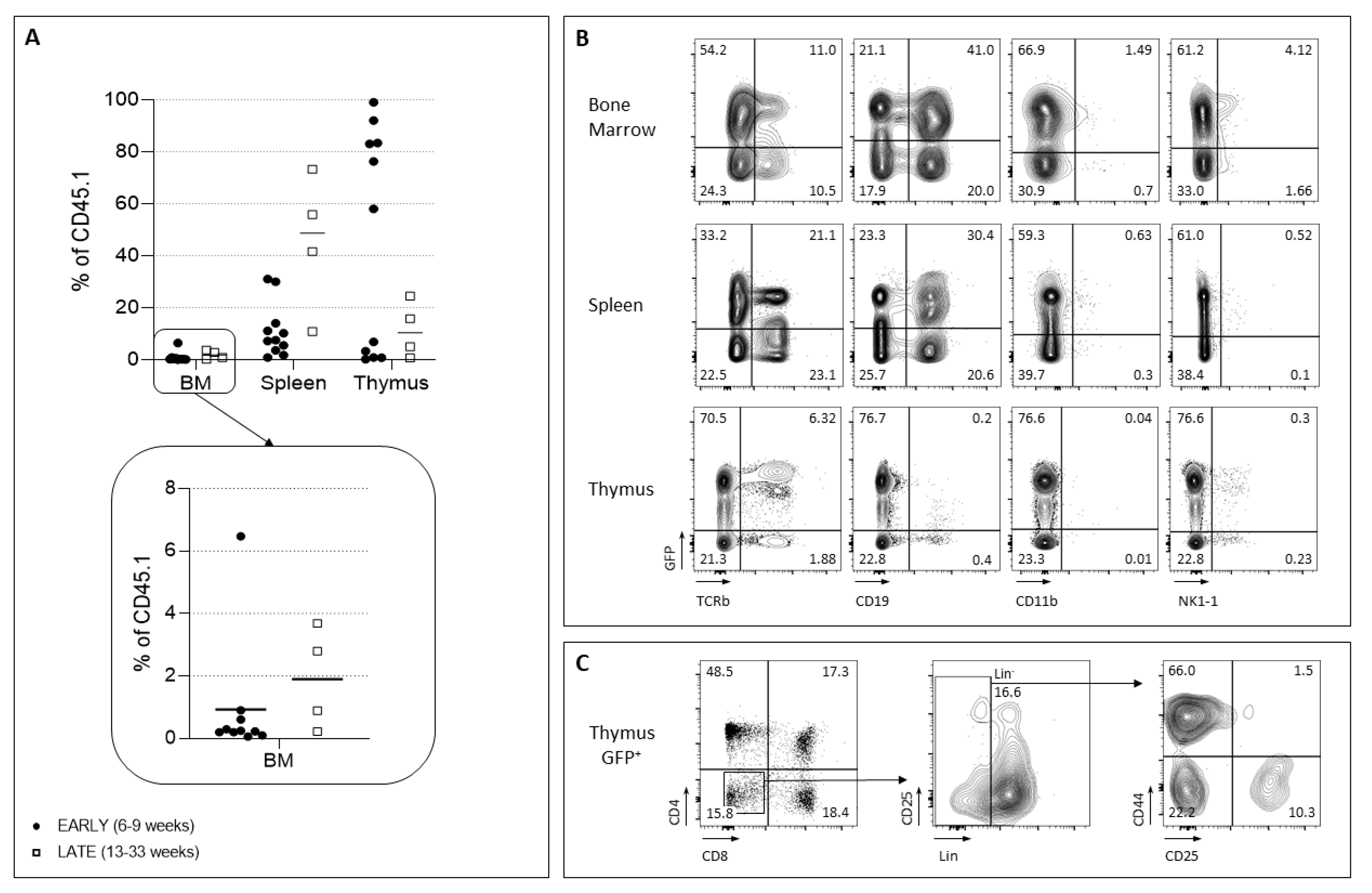
2.2. Barcoded HSC Transplantation for In Vivo LMPP and CLP Production
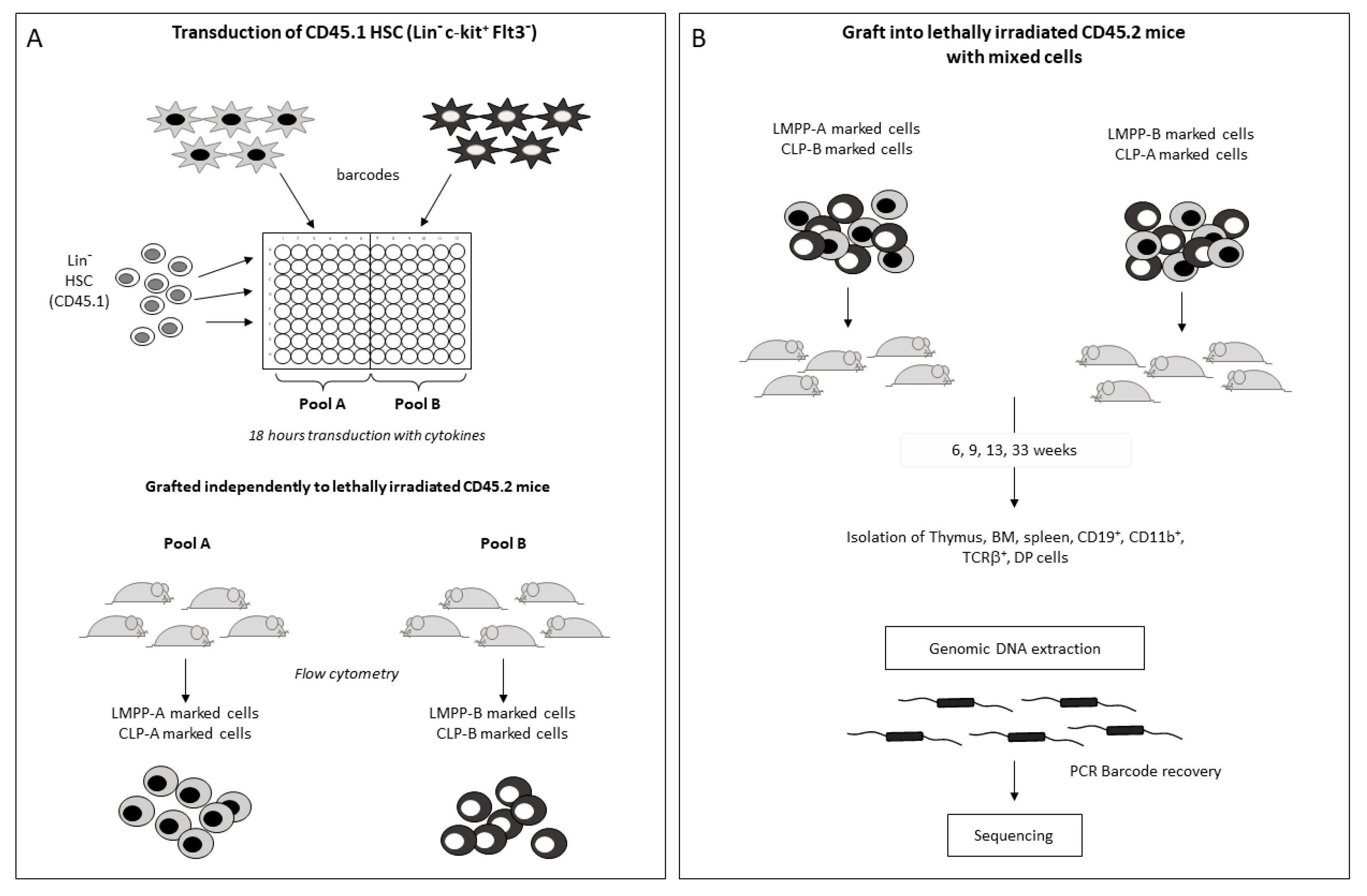
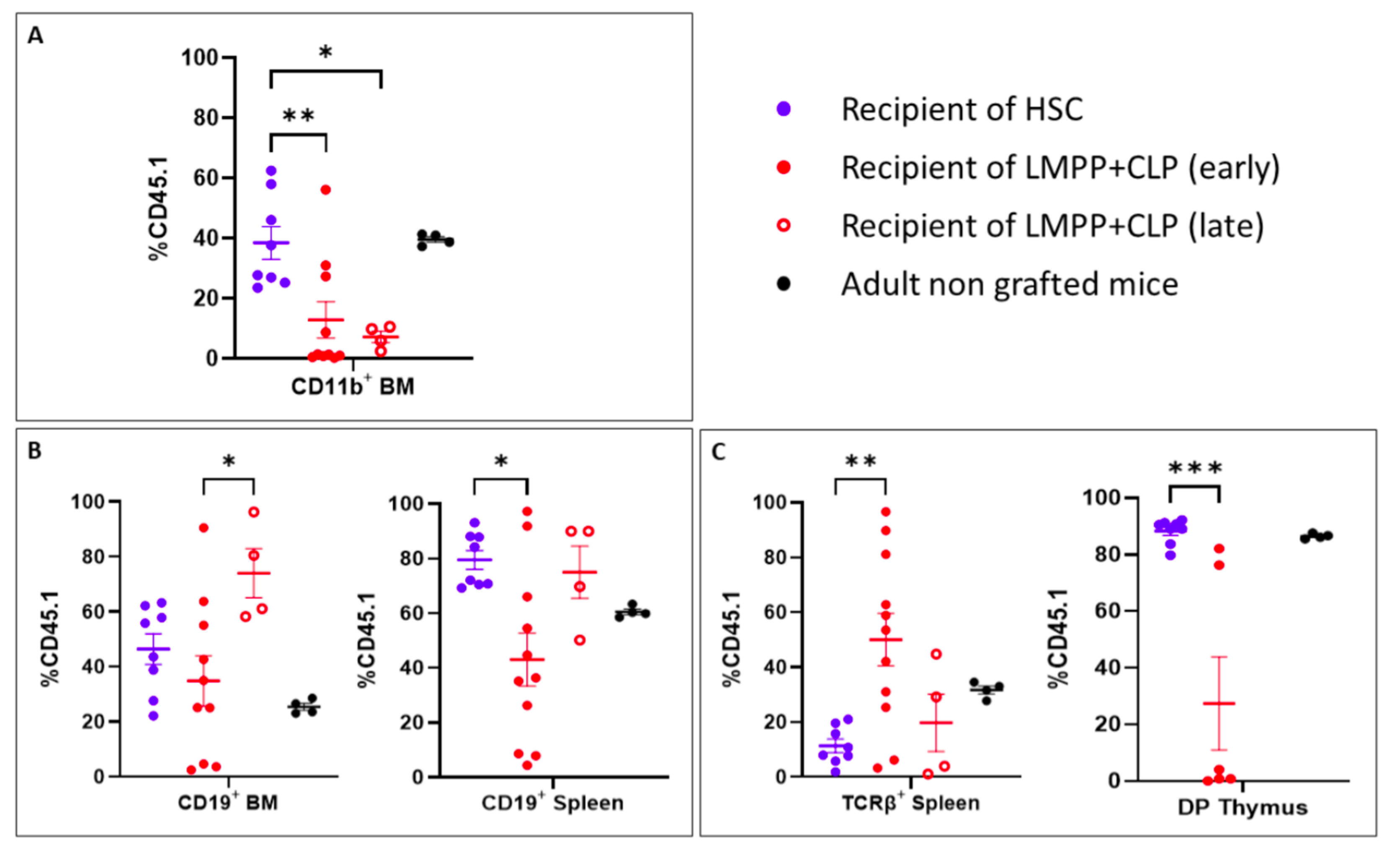
2.3. LMPP- and CLP-Barcoded Progenitor Production and Fate Determination
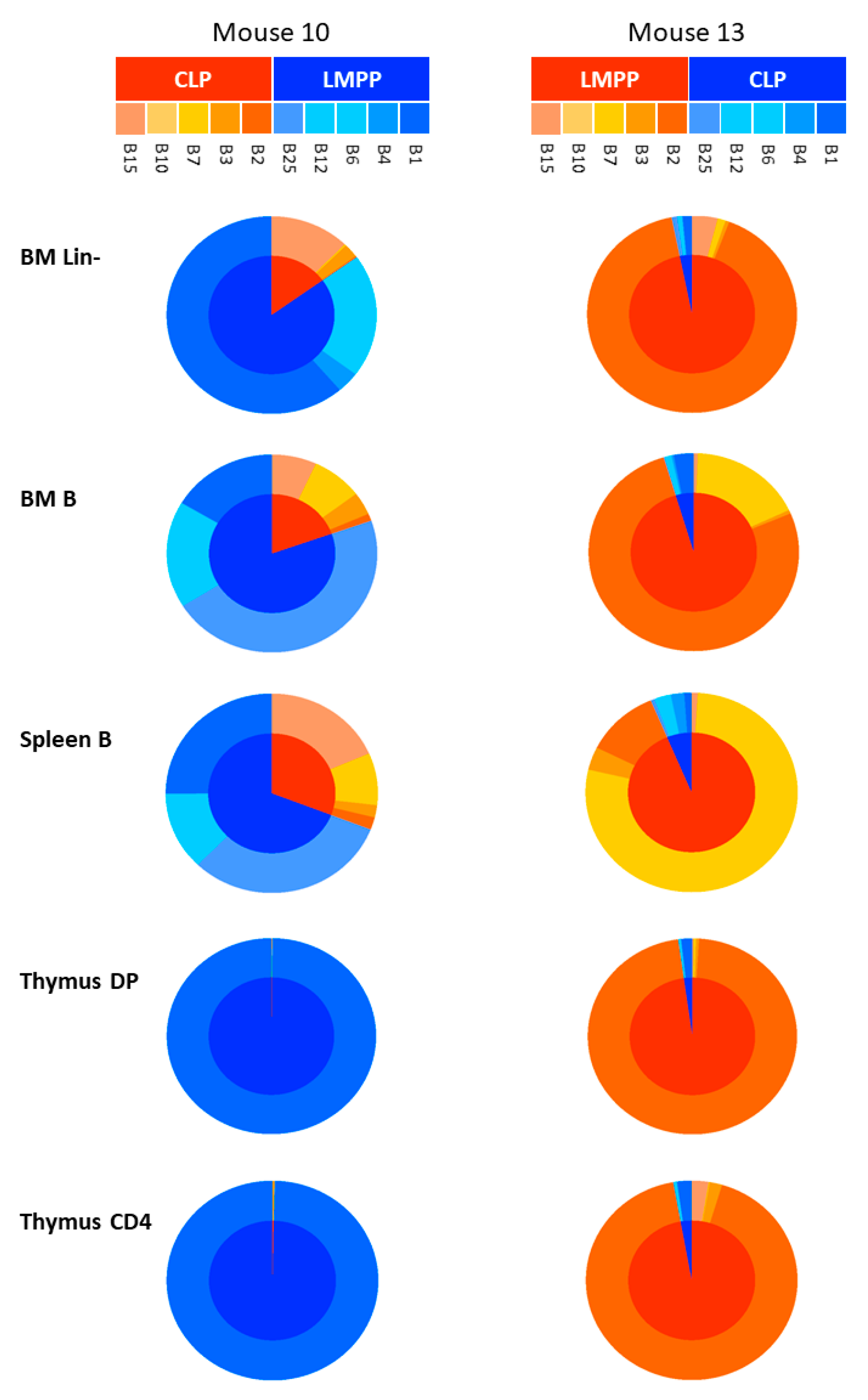
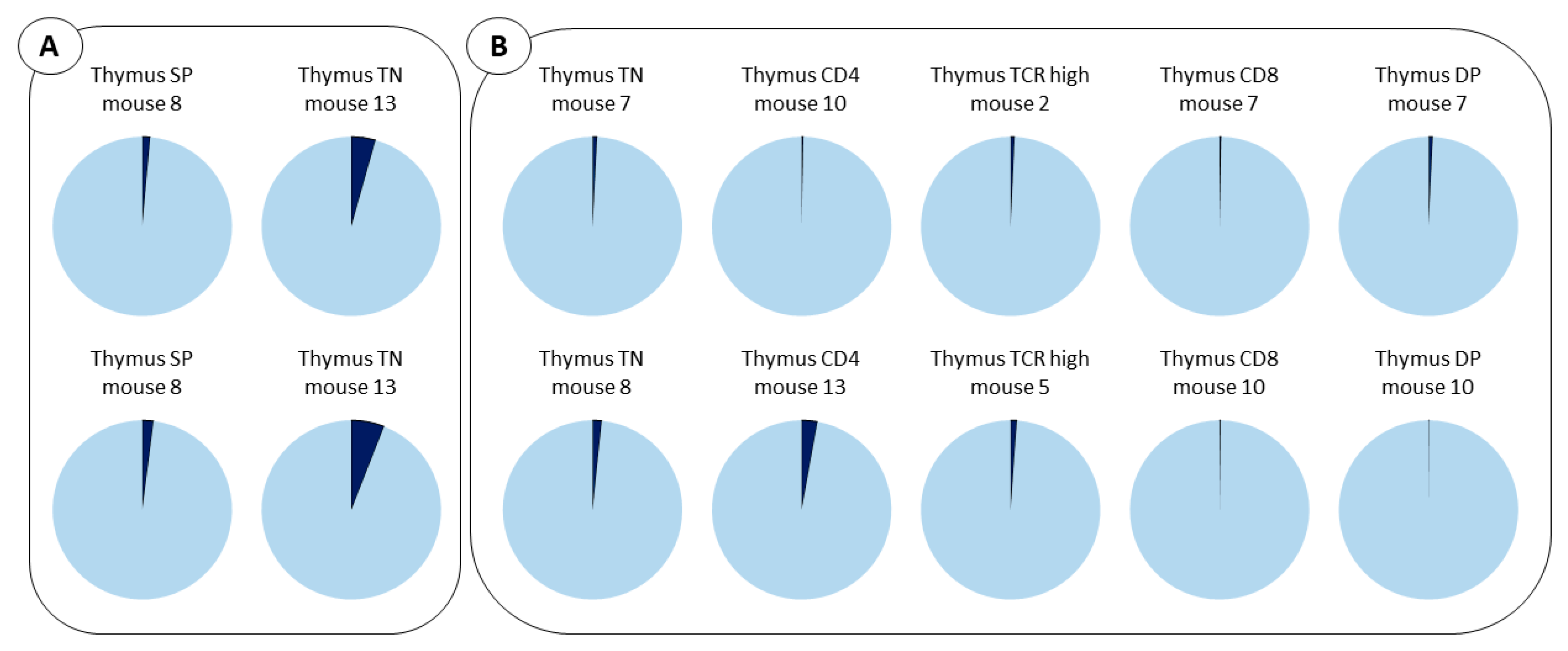
2.4. Cell Distribution of Barcodes Highlights the Major Role of LMPP Progenitors
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Antibodies and Cytometer
4.3. Isolation of Bone Marrow Progenitors
4.4. Cells Transduction
4.5. Grafting Strategy
4.6. Genomic DNA Extraction for Barcode Analysis
4.7. Analysis of Barcoded Genomic DNA
4.8. Sequencing of Tissue Extracted Barcodes
4.9. Bioinformatics Processing of BC Reads
4.10. Normalization of BC Read Counts
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luc, S.; Buza-Vidas, N.; Jacobsen, S.E.W. Delineating the Cellular Pathways of Hematopoietic Lineage Commitment. Semin. Immunol. 2008, 20, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Pietras, E.M.; Reynaud, D.; Kang, Y.-A.; Carlin, D.; Calero-Nieto, F.J.; Leavitt, A.D.; Stuart, J.M.; Göttgens, B.; Passegué, E. Functionally Distinct Subsets of Lineage-Biased Multipotent Progenitors Control Blood Production in Normal and Regenerative Conditions. Cell Stem Cell 2015, 17, 35–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, M.; Weissman, I.L.; Akashi, K. Identification of Clonogenic Common Lymphoid Progenitors in Mouse Bone Marrow. Cell 1997, 91, 661–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosselin, J.; Sii-Felice, K.; Payen, E.; Chretien, S.; Tronik-Le Roux, D.; Leboulch, P. Arrayed Lentiviral Barcoding for Quantification Analysis of Hematopoietic Dynamics. Stem Cells 2013, 31, 2162–2171. [Google Scholar] [CrossRef] [PubMed]
- Verovskaya, E.; Broekhuis, M.J.C.; Zwart, E.; Ritsema, M.; van Os, R.; de Haan, G.; Bystrykh, L.V. Heterogeneity of Young and Aged Murine Hematopoietic Stem Cells Revealed by Quantitative Clonal Analysis Using Cellular Barcoding. Blood 2013, 122, 523–532. [Google Scholar] [CrossRef]
- Lu, R.; Neff, N.F.; Quake, S.R.; Weissman, I.L. Tracking Single Hematopoietic Stem Cells in Vivo Using High-Throughput Sequencing in Conjunction with Viral Genetic Barcoding. Nat. Biotechnol. 2011, 29, 928–933. [Google Scholar] [CrossRef] [Green Version]
- Naik, S.H.; Perié, L.; Swart, E.; Gerlach, C.; van Rooij, N.; de Boer, R.J.; Schumacher, T.N. Diverse and Heritable Lineage Imprinting of Early Haematopoietic Progenitors. Nature 2013, 496, 229–232. [Google Scholar] [CrossRef]
- Cordeiro Gomes, A.; Hara, T.; Lim, V.Y.; Herndler-Brandstetter, D.; Nevius, E.; Sugiyama, T.; Tani-Ichi, S.; Schlenner, S.; Richie, E.; Rodewald, H.-R.; et al. Hematopoietic Stem Cell Niches Produce Lineage-Instructive Signals to Control Multipotent Progenitor Differentiation. Immunity 2016, 45, 1219–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.; Morrison, S.J. Haematopoietic Stem Cells and Early Lymphoid Progenitors Occupy Distinct Bone Marrow Niches. Nature 2013, 495, 231–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visnjic, D.; Kalajzic, Z.; Rowe, D.W.; Katavic, V.; Lorenzo, J.; Aguila, H.L. Hematopoiesis Is Severely Altered in Mice with an Induced Osteoblast Deficiency. Blood 2004, 103, 3258–3264. [Google Scholar] [CrossRef] [PubMed]
- Terashima, A.; Okamoto, K.; Nakashima, T.; Akira, S.; Ikuta, K.; Takayanagi, H. Sepsis-Induced Osteoblast Ablation Causes Immunodeficiency. Immunity 2016, 44, 1434–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaedi, M.; Steer, C.A.; Martinez-Gonzalez, I.; Halim, T.Y.F.; Abraham, N.; Takei, F. Common-Lymphoid-Progenitor-Independent Pathways of Innate and T Lymphocyte Development. Cell Rep. 2016, 15, 471–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brugman, M.H.; Wiekmeijer, A.-S.; van Eggermond, M.; Wolvers-Tettero, I.; Langerak, A.W.; de Haas, E.F.E.; Bystrykh, L.V.; van Rood, J.J.; de Haan, G.; Fibbe, W.E.; et al. Development of a Diverse Human T-Cell Repertoire despite Stringent Restriction of Hematopoietic Clonality in the Thymus. Proc. Natl. Acad. Sci. USA 2015, 112, E6020–E6027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Website Available online: Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 26 October 2016).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michaels, V.; Chalabi, S.; Legrand, A.; Renard, J.; Tejerina, E.; Daouya, M.; Fabrega, S.; Megret, J.; Olaso, R.; Boland, A.; et al. Co-Transplantation of Barcoded Lymphoid-Primed Multipotent (LMPP) and Common Lymphocyte (CLP) Progenitors Reveals a Major Contribution of LMPP to the Lymphoid Lineage. Int. J. Mol. Sci. 2023, 24, 4368. https://doi.org/10.3390/ijms24054368
Michaels V, Chalabi S, Legrand A, Renard J, Tejerina E, Daouya M, Fabrega S, Megret J, Olaso R, Boland A, et al. Co-Transplantation of Barcoded Lymphoid-Primed Multipotent (LMPP) and Common Lymphocyte (CLP) Progenitors Reveals a Major Contribution of LMPP to the Lymphoid Lineage. International Journal of Molecular Sciences. 2023; 24(5):4368. https://doi.org/10.3390/ijms24054368
Chicago/Turabian StyleMichaels, Victoria, Smahane Chalabi, Agnes Legrand, Julie Renard, Emmanuel Tejerina, Marina Daouya, Sylvie Fabrega, Jérôme Megret, Robert Olaso, Anne Boland, and et al. 2023. "Co-Transplantation of Barcoded Lymphoid-Primed Multipotent (LMPP) and Common Lymphocyte (CLP) Progenitors Reveals a Major Contribution of LMPP to the Lymphoid Lineage" International Journal of Molecular Sciences 24, no. 5: 4368. https://doi.org/10.3390/ijms24054368