


Exogenous Nitric Oxide-Induced Postharvest Gray Spot Disease Resistance in Loquat Fruit and Its Possible Mechanism of Action
Abstract
:1. Introduction
2. Results
2.1. Effects of SNP on Disease Incidence and Lesion Diameter of Loquat Fruit Inoculated with P. eriobotryfoli
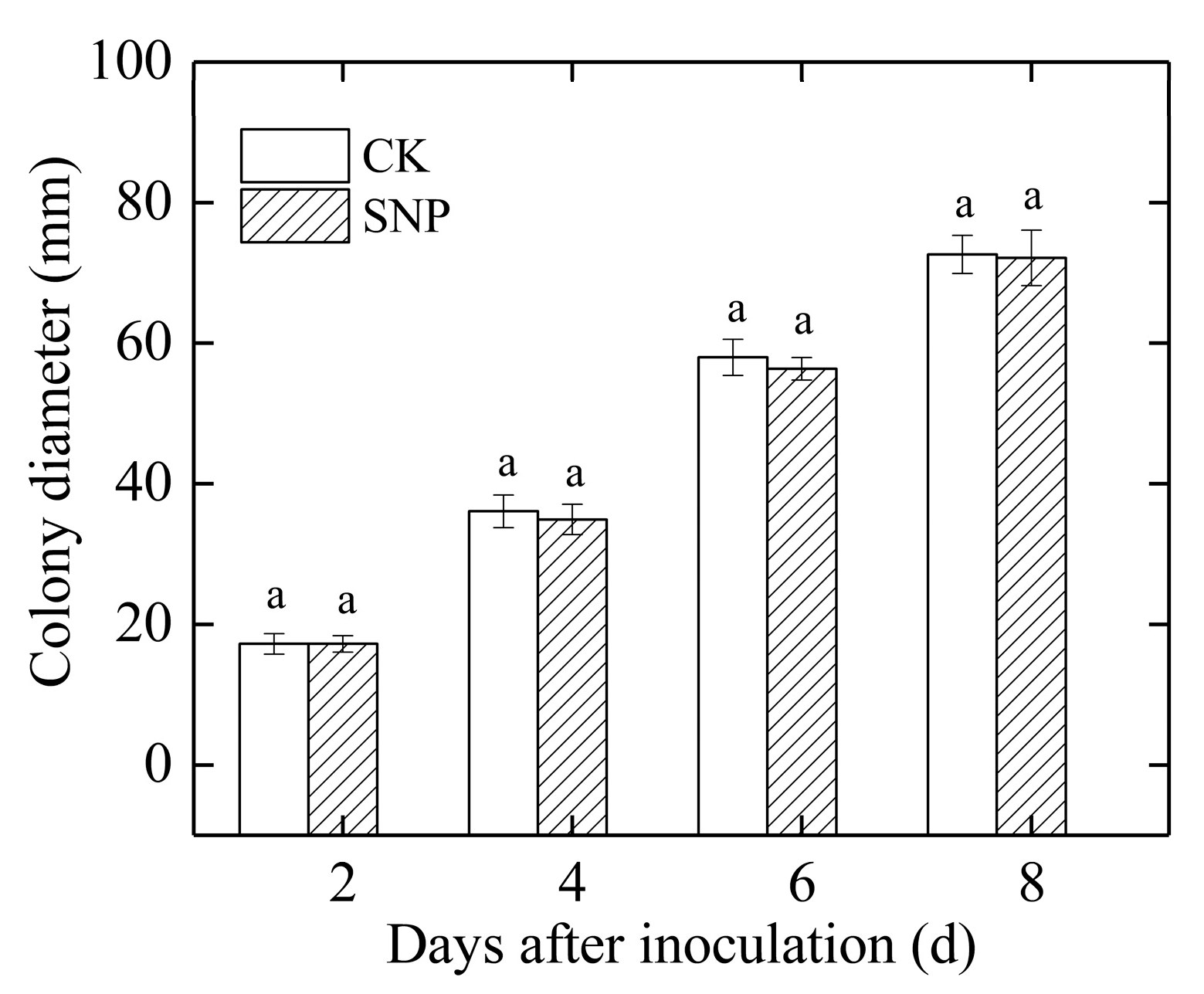
2.2. Effects of SNP on Mycelial Growth and Spore Germination of P. eriobotryfolia In Vitro
2.3. Effect of SNP on Activities of SOD, CAT and APX as Well as H2O2 Content in Loquat Fruit
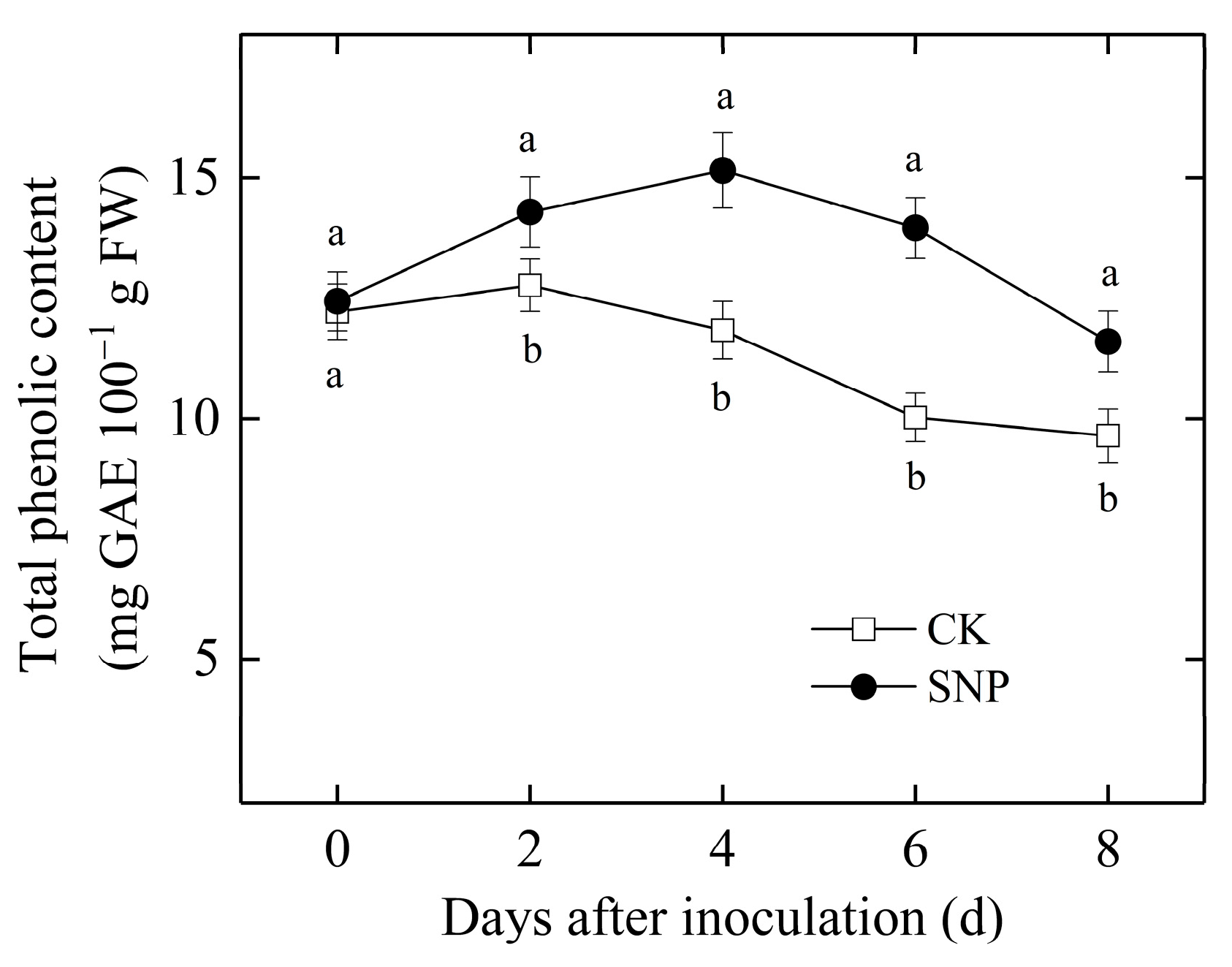
2.4. Effects of SNP on the Activities of GLU, CHI, PAL, and PPO and Total Phenolic Content of Loquat Fruit Inoculated with P. eriobotryfolia
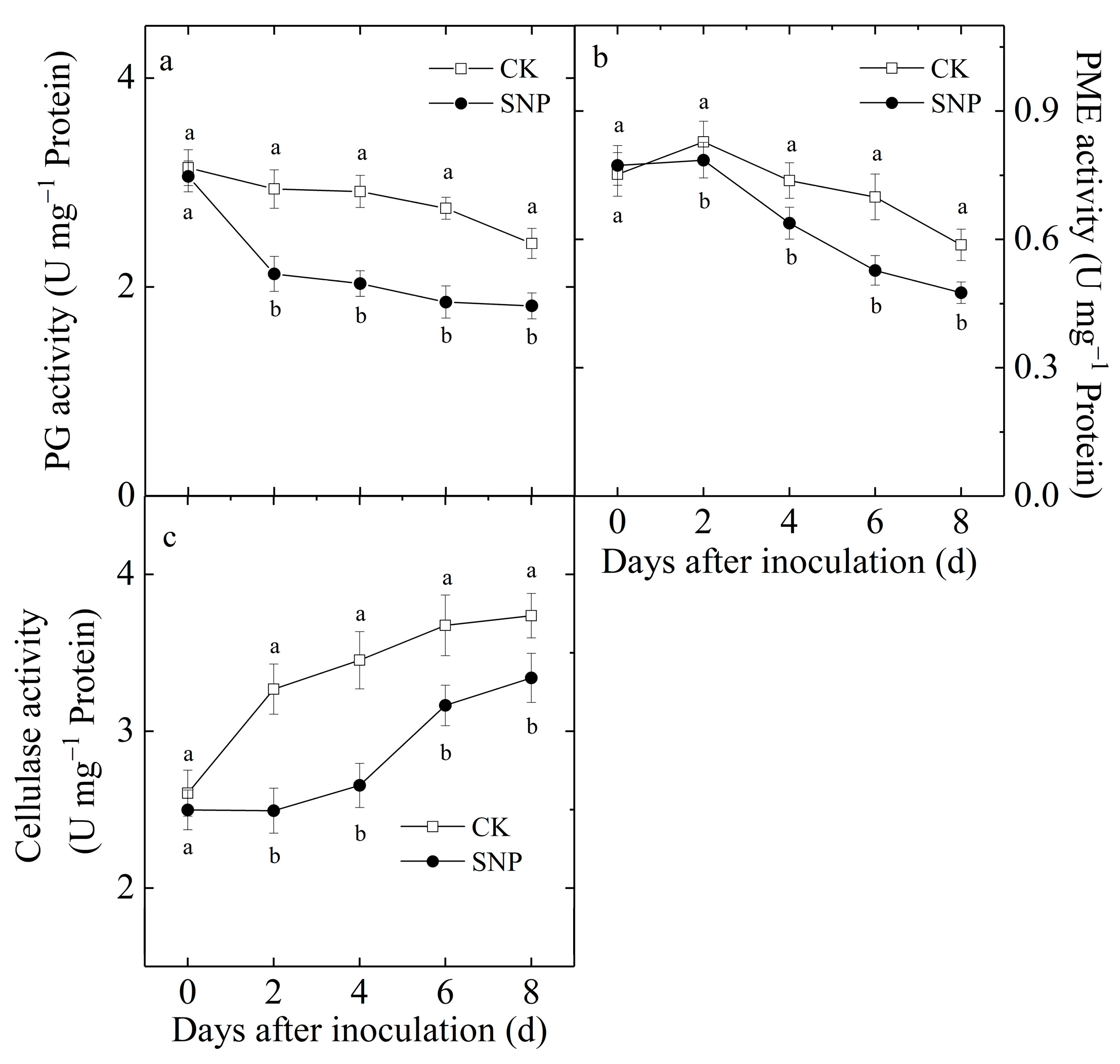
2.5. Effects of SNP on PG, PME and Cellulase Activities of Loquat Fruit Inoculated with P. eriobotryfolia
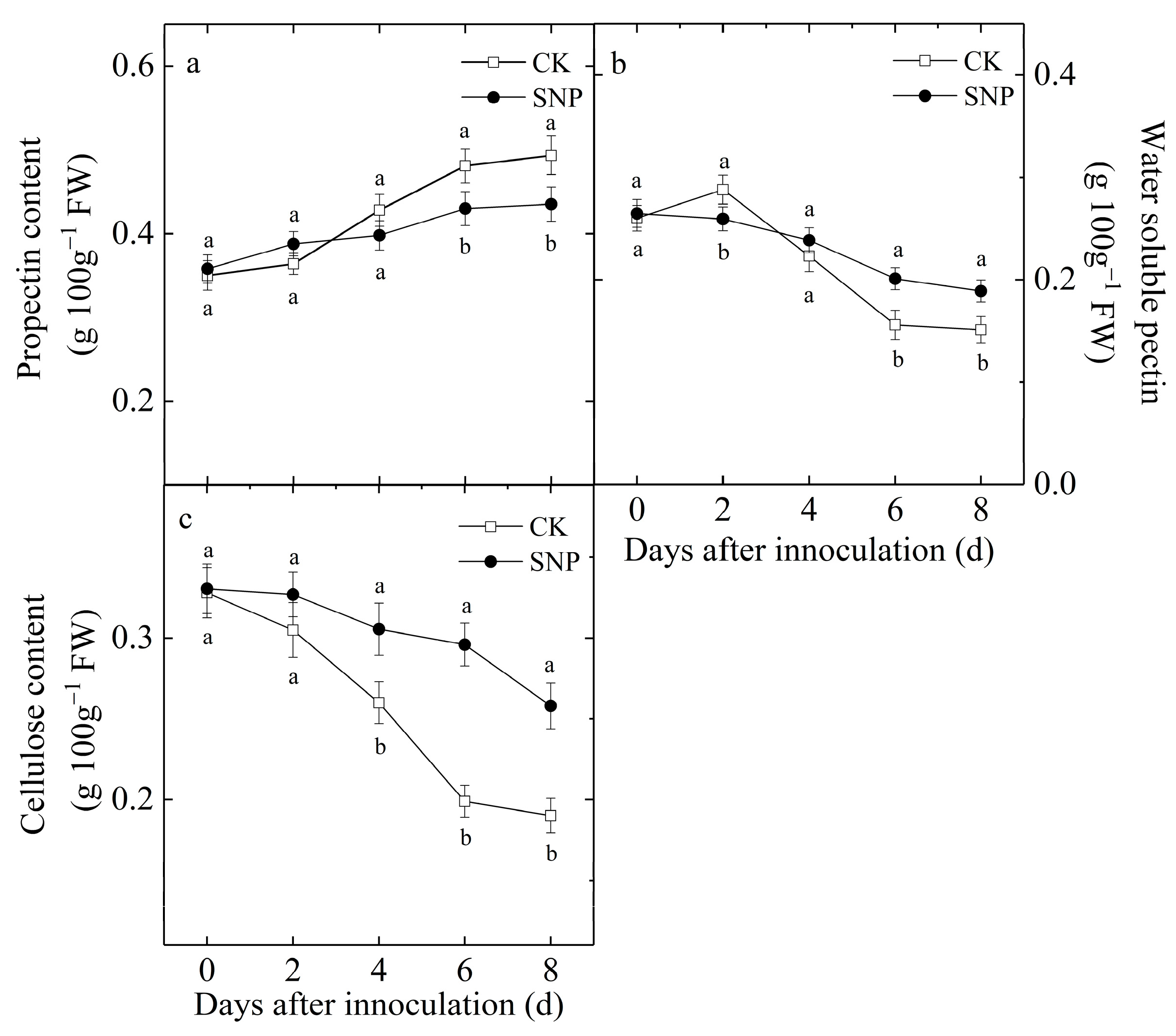
2.6. Effects of SNP on Propectin, WSP and Cellulose Contents of Loquat Fruit Inoculated with P. eriobotryfolia
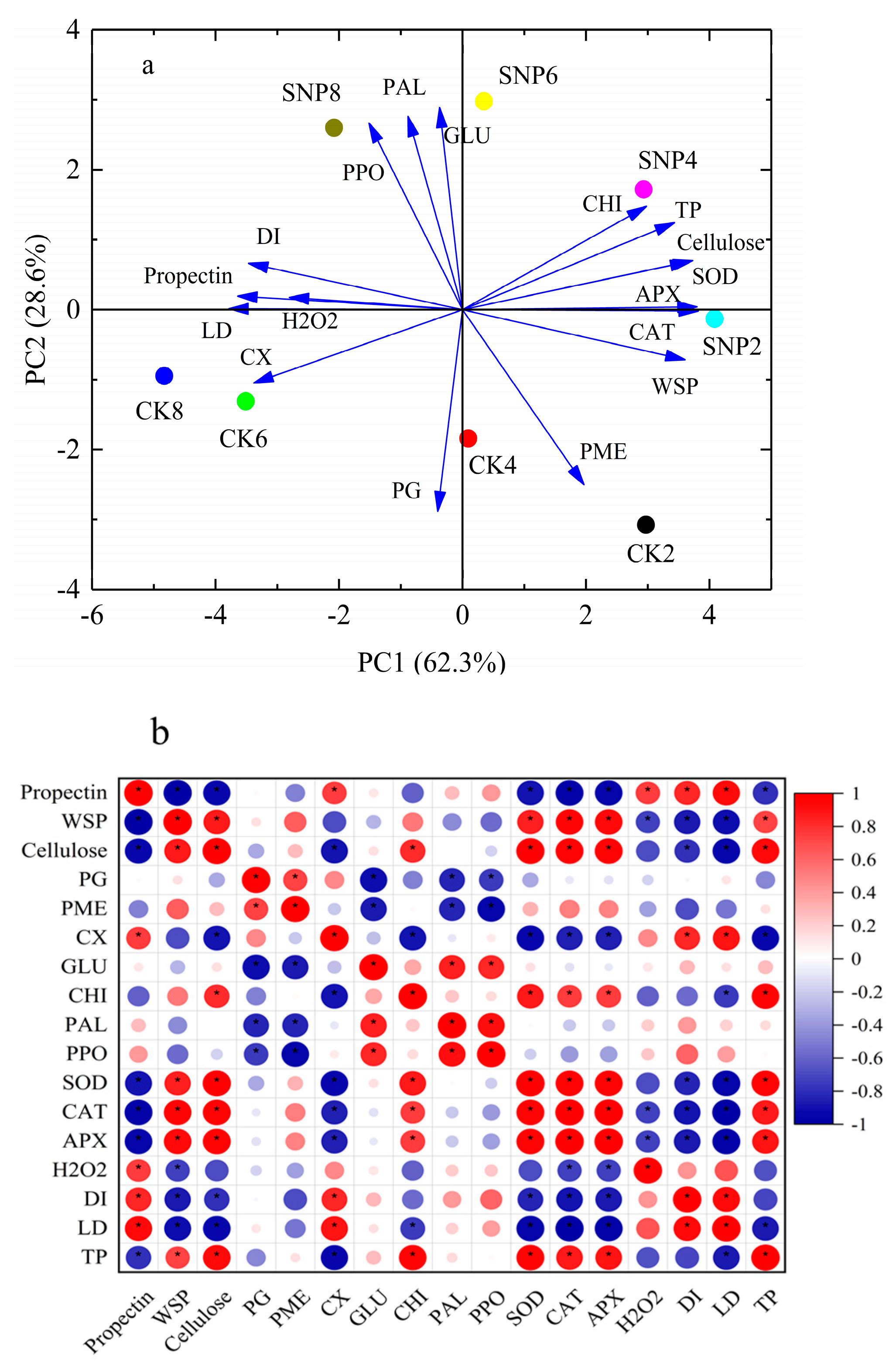
2.7. Principal Component and Pearson Correlation Analysis
3. Discussion
4. Materials and Methods
4.1. Fruit Materials
4.2. Pathogen
4.3. SNP Treatment and Inoculation
4.4. SOD, CAT and APX Activities and H2O2 Content
4.5. Total Phenolics Content
4.6. Pectin and Cellulose Content
4.7. Cell Wall-Related Enzymes
4.8. Disease Resistance-Related Enzyme Activities
4.9. Effect of SNP on Mycelial Growth and Spore Germination of P. eriobotryfolia In Vitro
4.9.1. Mycelial Growth
4.9.2. Spore Germination and Germ Tube Elongation
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alos, E.; Martinez-Fuentes, A.; Reig, C.; Mesejo, C.; Zacarías, L.; Agustí, M.; Rodrigo, M.J. Involvement of ethylene in color changes and carotenoid biosynthesis in loquat fruit (Eriobotrya japonica Lindl. cv. Algerie). Postharvest Biol. Technol. 2019, 149, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.Y.; Xu, F.; Shao, X.F. Changes in soluble sugar metabolism in loquat fruit during different cold storage. J. Food Sci. Tech. 2017, 54, 1043–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pareek, S.; Benkeblia, N.; Janick, J.; Cao, S.F.; Yahia, E.M. Postharvest physiology and technology of loquat (Eriobotrya japonica Lindl.) fruit. J. Sci. Food Agric. 2014, 94, 1495–1504. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.T.; Cao, S.F.; Di, Y.Q.; Liao, Y.X.; Zheng, Y.H. Effect of ethanol treatment on disease resistance against anthracnose rot in postharvest loquat fruit. Sci. Hortic. 2015, 188, 115–121. [Google Scholar] [CrossRef]
- Wang, B.; He, X.; Bi, Y.; Jiang, H.; Wang, Y.; Zheng, X.; Prusky, D. Preharvest sprays with sodium nitroprusside induce resistance in harvested muskmelon against the pink rot disease. J. Food Process. Preserv. 2021, 45, e15339. [Google Scholar] [CrossRef]
- Ling, C.; Xu, J.; Shao, S.; Wang, L.; Jin, P.; Zheng, Y.H. Effect of ultrasonic treatment combined with peracetic acid treatment reduces decay and maintains quality in loquat fruit. J. Food Qual. 2018, 2018, 7564056. [Google Scholar] [CrossRef] [Green Version]
- Pols, S.; van de Poel, B.; Hertog, M.L.A.T.M.; Nicolai, B.M. The regulatory role of nitric oxide and its significance for future postharvest applications. Postharvest Biol. Technol. 2022, 188, 111869. [Google Scholar] [CrossRef]
- He, J.Y.; Ren, Y.F.; Chen, C.; Liu, J.P.; Liu, H.Y.; Pei, Y. Defense responses of salicylic acid in mango fruit against postharvest anthracnose, caused by Colletotrichum gloeosporioides and its possible mechanism. J. Food Saf. 2017, 37, e12294. [Google Scholar] [CrossRef]
- He, J.; Ren, Y.; Chen, X.; Chen, H. Protective roles of nitric oxide on seed germination and seedling growth of rice (Oryza sativa L.) under cadmium stress. Ecotoxicol. Environ. Saf. 2014, 108, 114–119. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, W.; He, J.; Zhang, L.; Wei, Y.; Yang, M. Nitric oxide alleviates salt stress in seed germination and early seedling growth of pakchoi (Brassica chinensis L.) by enhancing physiological and biochemical parameters. Ecotoxicol. Environ. Saf. 2020, 187, 109785. [Google Scholar] [CrossRef]
- Zheng, X.L.; Hu, B.; Song, L.J.; Pan, J.; Liu, M.M. Changes in quality and defense resistance of kiwifruit in response to nitric oxide treatment during storage at room temperature. Sci. Hortic. 2017, 222, 187–192. [Google Scholar] [CrossRef]
- Khaliq, G.; Ullah, M.; Memon, S.A.; Ali, A.; Rashid, M. Exogenous nitric oxide reduces postharvest anthracnose disease and maintains quality of custard apple (Annona squamosa L.) fruit during ripening. J. Food Meas. Charact. 2021, 15, 707–716. [Google Scholar] [CrossRef]
- Lata, D.; Aftab, M.A.; Homa, F.; Ahmad, M.S.; Siddiqui, M.W. Effect of eco-safe compounds on postharvest quality preservation of papaya (Carica papaya L.). Acta Physiol. Plant. 2018, 40, 8. [Google Scholar] [CrossRef]
- Zhou, Y.H.; Li, S.M.; Zeng, K.F. Exogenous nitric oxide-induced postharvest disease resistance in citrus fruit to Colletotrichum gloeosporioides. J. Sci. Food Agric. 2016, 96, 505–512. [Google Scholar] [CrossRef]
- Lai, T.F.; Wang, Y.Y.; Li, B.Q.; Qin, G.Z.; Tian, S.P. Defense responses of tomato fruit to exogenous nitric oxide during postharvest storage. Postharvest Biol. Technol. 2011, 62, 127–132. [Google Scholar] [CrossRef]
- Shi, J.Y.; Liu, N.; Gu, R.X.; Zhu, L.Q.; Zhang, C.; Wang, Q.; Lei, Z.H.; Liu, Y.Y.; Ren, J.Y. Signals induced by exogenous nitric oxide and their role in controlling brown rot disease caused by Monilinia fructicola in postharvest peach fruit. J. Gen. Plant Pathol. 2015, 81, 68–76. [Google Scholar] [CrossRef]
- Hu, L.G.; Li, Y.C.; Bi, Y.; Li, J.P.; Bao, G.H.; Liu, J.J.; Yu, X.Y. Effects of nitric oxide on growth of Fusarium sulphureum and its virulence to potato tubers. Eur. Food Res. Technol. 2014, 238, 1007–1014. [Google Scholar] [CrossRef]
- Ren, Y.F.; He, J.Y.; Liu, H.Y.; Liu, G.Q.; Ren, X.L. Nitric oxide alleviates deterioration and preserves antioxidant properties in ‘Tainong’ mango fruit during ripening. Hortic. Environ. Biotechnol. 2017, 58, 27–37. [Google Scholar] [CrossRef]
- Hu, M.J.; Yang, D.P.; Huber, D.J.; Jiang, Y.M.; Li, M.; Gao, Z.Y.; Zhang, Z.K. Reduction of postharvest anthracnose and enhancement of disease resistance in ripening mango fruit by nitric oxide treatment. Postharvest Biol. Technol. 2014, 97, 115–122. [Google Scholar] [CrossRef]
- Gu, R.X.; Zhu, S.H.; Zhou, J.; Liu, N.; Shi, J.Y. Inhibition on brown rot disease and induction of defence response in harvested peach fruit by nitric oxide solution. Eur. J. Plant Pathol. 2014, 139, 369–378. [Google Scholar] [CrossRef]
- Wang, J.; Higgins, V.J. Nitric oxide has a regulatory effect in the germination of conidia of Colletotrichum coccodes. Fungal Genet. Biol. 2005, 42, 284–292. [Google Scholar] [CrossRef]
- Lazar, E.E.; Wills, R.B.; Ho, B.T.; Harris, A.M.; Spohr, L.J. Antifungal effect of gaseous nitric oxide on mycelium growth, sporulation and spore germination of the postharvest horticulture pathogens, Aspergillus niger, Monilinia fructicola and Penicillium italicum. Lett. Appl. Microbiol. 2008, 46, 688–692. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Liu, A.Y.; Chen, W.X.; Feng, S.J.; Shi, J.Y. Development and control of postharvest diseases of loquat fruit. Acta Hortic. 2007, 750, 437–443. [Google Scholar] [CrossRef]
- Zheng, Y.Y.; Hong, H.; Chen, L.; Li, J.Y.; Sheng, J.P.; Shen, L. LeMAPK1, LeMAPK2, and LeMAPK3 are associated with nitric oxide-induced defense response against Botrytis cinerea in the Lycopersicon esculentum fruit. J. Agric. Food Chem. 2014, 62, 1390–1396. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Xue, Y.; Tian, D.; Zhang, L.; Xiao, G.; He, J. Improvement of postharvest anthracnose resistance in mango fruit by nitric oxide and the possible mechanisms involved. J. Agric. Food Chem. 2020, 68, 15460–15467. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.F.; Zheng, Y.H.; Yang, Z.F.; Tang, S.S.; Jin, P.; Wang, K.T.; Wang, X.M. Effect of methyl jasmonate on the inhibition of Colletotrichum acutatum infection in loquat fruit and the possible mechanisms. Postharvest Biol. Technol. 2008, 49, 301–307. [Google Scholar] [CrossRef]
- Song, Y.P.; Hu, C.M.; Xue, Y.H.; Gu, J.Y.; He, J.Y.; Ren, Y.F. 24-Epibrassinolide enhances mango resistance to Colletotrichum gloeosporioides via activating multiple defense response. Sci. Hortic. 2022, 303, 111249. [Google Scholar] [CrossRef]
- Liu, K.; Wang, X.L.; Young, M. Effect of bentonite/potassium sorbate coatings on the quality of mangos in storage at ambient temperature. J. Food Eng. 2014, 137, 16–22. [Google Scholar] [CrossRef]
- Yuan, L.; Bi, Y.; Ge, Y.H.; Wang, Y.; Liu, Y.Y.; Li, G.L. Postharvest hot water dipping reduces decay by inducing disease resistance and maintaining firmness in muskmelon (Cucumis melo L.) fruit. Sci. Hortic. 2013, 161, 101–110. [Google Scholar] [CrossRef]
- Sinha, A.; Gill, P.P.S.; Jawandha, S.K.; Kaur, P.; Grewal, S.K. Salicylic acid enriched beeswax coatings suppress fruit softening in pears by modulation of cell wall degrading enzymes under different storage conditions. Food Packag. Shelf Life 2022, 32, 100821. [Google Scholar] [CrossRef]
- Guo, Q.; Wu, B.; Chen, W.X.; Zhang, Y.L.; Wang, J.D.; Li, X.P. Effects of nitric oxide treatment on the cell wall softening related enzymes and several hormones of papaya fruit during storage. Food Sci. Technol. Int. 2013, 20, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Aghdama, M.S.; Kakavand, F.; Rabiei, V.; Zaare-Nahandi, F. γ-Aminobutyric acid and nitric oxide treatments preserve sensory and nutritional quality of cornelian cherry fruits during postharvest cold storage by delaying softening and enhancing phenols accumulation. Sci. Hortic. 2019, 246, 812–817. [Google Scholar] [CrossRef]
- Gu, J.; Hu, C.; Jia, X.; Ren, Y.; Su, D.; He, J. Physiological and biochemical bases of spermidine-induced alleviation of cadmium and lead combined stress in rice. Plant Physiol. Biochem. 2022, 189, 104–114. [Google Scholar] [CrossRef]
- Ren, Y.F.; He, J.Y.; Liu, D.; Liu, J.P. Effect of nitric oxide on fruit quality of “Dawuxin” loquat during storage at room temperature. Sci. Technol. Food Ind. 2016, 37, 329–333. (In Chinese) [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNP (μM) | 6 h | 12 h | ||
|---|---|---|---|---|
| Spore Germination (%) | Germ Tube Length (μm) | Spore Germination (%) | Germ Tube Length (μm) | |
| 0 (CK) | 23.9 ± 2.9 a | 48.6 ± 7.6 a | 90.3 ± 4.1 a | 73.9 ± 17.2 a |
| 50 | 20.8 ± 2.7 a | 44.0 ± 9.3 a | 86.1 ± 3.9 a | 65.8 ± 14.5 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, Y.; Yan, T.; Hu, C.; Liu, D.; He, J. Exogenous Nitric Oxide-Induced Postharvest Gray Spot Disease Resistance in Loquat Fruit and Its Possible Mechanism of Action. Int. J. Mol. Sci. 2023, 24, 4369. https://doi.org/10.3390/ijms24054369
Ren Y, Yan T, Hu C, Liu D, He J. Exogenous Nitric Oxide-Induced Postharvest Gray Spot Disease Resistance in Loquat Fruit and Its Possible Mechanism of Action. International Journal of Molecular Sciences. 2023; 24(5):4369. https://doi.org/10.3390/ijms24054369
Chicago/Turabian StyleRen, Yanfang, Tengyu Yan, Chunmei Hu, Dong Liu, and Junyu He. 2023. "Exogenous Nitric Oxide-Induced Postharvest Gray Spot Disease Resistance in Loquat Fruit and Its Possible Mechanism of Action" International Journal of Molecular Sciences 24, no. 5: 4369. https://doi.org/10.3390/ijms24054369