
Molecular Mechanisms of Hyperoxia-Induced Neonatal Intestinal Injury

Abstract
:1. Introduction
2. Embryonic Intestine Development
3. Intestinal Histological Changes after Hyperoxia
4. Mechanisms Associated with Hyperoxia-Induced Intestinal Injury (Table 1)
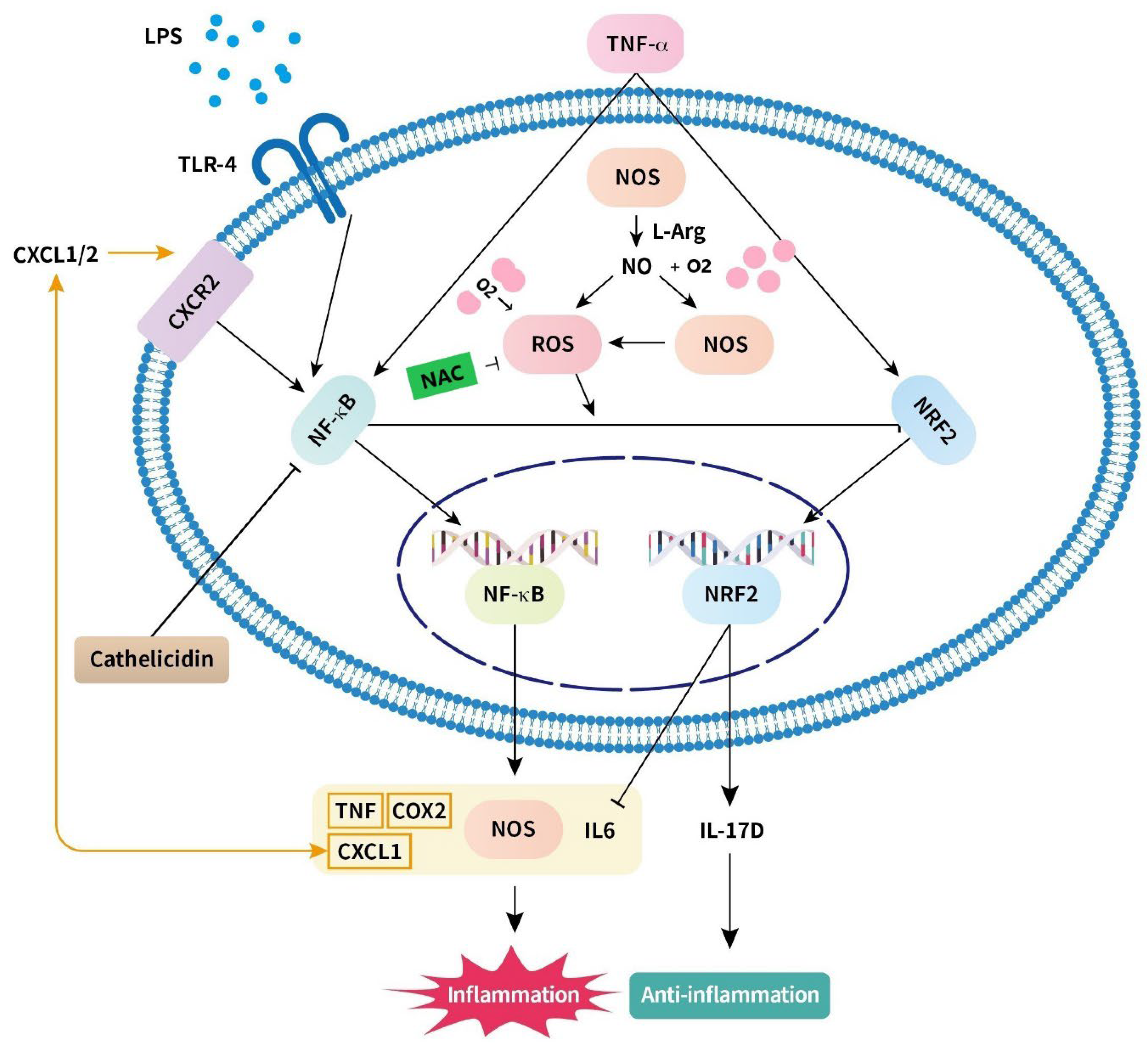
4.1. Nitric Oxide (Figure 1)
4.2. Nuclear Factor-κB (Figure 1)
4.3. Reactive Oxygen Species (Figure 1)
4.4. Toll-like Receptor (Figure 1)
4.5. Chemokine (CXC Motif) Ligand (Figure 1)
4.6. IL-6 (Figure 1)

{kind=link}
| Injury Mechanism | ||||
| Candidate | Mechanism | Model | Outcome | Reference |
| Excessive Nitric oxide (NO) |
| Rat; Piglet |
| [23,45,47] |
| Nuclear factor-κB (NF-κB) |
| Human HT-29 cell; Rat |
| [30,52] |
| Reactive oxygen species (ROS) |
| Rat; Human Caco-2 cell; Human NCM460 cell |
| [24,32,36,56] |
| Toll-like receptor-4 (TLR4) |
| Rat; Human cell |
| [40,52,59] |
| CXC motif ligand-1 (CXCL1) |
| Rat |
| [24] |
| Interleukin-6 (IL-6) |
| Rat; Piglet |
| [18,24,47,64] |
| Protective Mechanism | ||||
| Candidate | Mechanism | Model | Outcome | Reference |
| Nuclear factor erythroid 2-related factor 2 (Nrf2) |
| Rat; Piglet; Human NCM460 cell; Human F244 cell; Murine embryonic fibroblasts cell |
| [54,65,66] |
| Interleukin-17D (IL-17D) |
| Rat; Human F244 cell; Murine embryonic fibroblasts cell |
| [65,66] |
| N-acetylcysteine (NAC) |
| Rat |
| [36] |
| Arginyl-glutamine Arg-Gln |
| Rat; preterm neonate |
| [67,68] |
| Deoxyribonucleic acid (DHA) |
| Rat |
| [67,69] |
| Cathelicidin |
| Rat; Piglet; Porcine epithelial cell line J2 |
| [22,70,71] |
| Health gut microbiome |
| Rat; Human infant |
| [24,38,72,73,74,75,76] |

5. Protective Mechanism under Hyperoxic Conditions (Table 1)
5.1. Nuclear Factor Erythroid 2-Related Factor 2 and IL-17D (Figure 1)
5.2. N-Acetylcysteine (Figure 1)
5.3. Arginyl–Glutamine (Figure 1)
5.4. Docosahexaenoic Acid (Figure 1)
5.5. Cathelicidin (Figure 1)
5.6. Health Gut Microbiome
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cheng, H.L. Effect of hyperoxia and hypercapnia on tissue oxygen and perfusion response in the normal liver and kidney. PLoS ONE 2012, 7, e40485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathias, M.; Chang, J.; Perez, M.; Saugstad, O. Supplemental Oxygen in the Newborn: Historical Perspective and Current Trends. Antioxidants 2021, 10, 1879. [Google Scholar] [CrossRef] [PubMed]
- Torbati, D.; Tan, G.H.; Smith, S.; Frazier, K.S.; Gelvez, J.; Fakioglu, H.; Totapally, B.R. Multiple-organ effect of normobaric hyperoxia in neonatal rats. J. Crit. Care 2006, 21, 85–93, discussion 93–84. [Google Scholar] [CrossRef]
- Giusto, K.; Wanczyk, H.; Jensen, T.; Finck, C. Hyperoxia-induced bronchopulmonary dysplasia: Better models for better therapies. Dis. Model. Mech. 2021, 14, dmm047753. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Lei, X.; Wang, J.; Dong, W. Protective effects of resveratrol on hyperoxia-induced lung injury in neonatal rats by alleviating apoptosis and ROS production. J. Matern. Fetal Neonatal Med. 2020, 33, 4150–4158. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, V. Hyperoxia-derived lung damage in preterm infants. Semin. Fetal. Neonatal. Med. 2010, 15, 223–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Indrio, F.; Neu, J.; Pettoello-Mantovani, M.; Marchese, F.; Martini, S.; Salatto, A.; Aceti, A. Development of the Gastrointestinal Tract in Newborns as a Challenge for an Appropriate Nutrition: A Narrative Review. Nutrients 2022, 14, 1405. [Google Scholar] [CrossRef]
- Sanidad, K.Z.; Zeng, M.Y. Neonatal gut microbiome and immunity. Curr. Opin. Microbiol. 2020, 56, 30–37. [Google Scholar] [CrossRef]
- Yu, J.C.; Khodadadi, H.; Malik, A.; Davidson, B.; Salles, É.D.S.L.; Bhatia, J.; Hale, V.L.; Baban, B. Innate Immunity of Neonates and Infants. Front. Immunol. 2018, 9, 1759. [Google Scholar] [CrossRef] [Green Version]
- Jena, A.; Montoya, C.A.; Mullaney, J.A.; Dilger, R.N.; Young, W.; McNabb, W.C.; Roy, N.C. Gut-Brain Axis in the Early Postnatal Years of Life: A Developmental Perspective. Front. Integr. Neurosci. 2020, 14, 44. [Google Scholar] [CrossRef]
- Esrefoglu, M.; Cetin, A. Development of Small and Large Intestine. Bezmialem. Sci. 2016, 5, 36–40. [Google Scholar] [CrossRef]
- Chin, A.M.; Hill, D.R.; Aurora, M.; Spence, J.R. Morphogenesis and maturation of the embryonic and postnatal intestine. Semin. Cell Dev. Biol. 2017, 66, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Clayburgh, D.R.; Shen, L.; Turner, J.R. A porous defense: The leaky epithelial barrier in intestinal disease. Lab. Investig. A J. Tech. Methods Pathol. 2004, 84, 282–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Zwart, L.L.; Haenen, H.E.; Versantvoort, C.H.; Wolterink, G.; van Engelen, J.G.; Sips, A.J. Role of biokinetics in risk assessment of drugs and chemicals in children. Regul. Toxicol. Pharmacol. 2004, 39, 282–309. [Google Scholar] [CrossRef] [PubMed]
- Pácha, J. Development of intestinal transport function in mammals. Physiol. Rev. 2000, 80, 1633–1667. [Google Scholar] [CrossRef]
- Henning, S.J. Postnatal development: Coordination of feeding, digestion, and metabolism. Am. J. Physiol. 1981, 241, G199–G214. [Google Scholar] [CrossRef]
- Dotinga, B.M.; Mintzer, J.P.; Moore, J.E.; Hulscher, J.B.F.; Bos, A.F.; Kooi, E.M.W. Maturation of Intestinal Oxygenation: A Review of Mechanisms and Clinical Implications for Preterm Neonates. Front. Pediatr. 2020, 8, 354. [Google Scholar] [CrossRef]
- Elgin, T.G.; Fricke, E.M.; Gong, H.; Reese, J.; Mills, D.A.; Kalantera, K.M.; Underwood, M.A.; McElroy, S.J. Fetal exposure to maternal inflammation interrupts murine intestinal development and increases susceptibility to neonatal intestinal injury. Dis. Model. Mech. 2019, 12, dmm040808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Q.; Walker, W.A. Innate immunity of the gut: Mucosal defense in health and disease. J. Pediatr. Gastroenterol. Nutr. 2004, 38, 463–473. [Google Scholar] [CrossRef] [Green Version]
- Montalban-Arques, A.; Chaparro, M.; Gisbert, J.P.; Bernardo, D. The Innate Immune System in the Gastrointestinal Tract: Role of Intraepithelial Lymphocytes and Lamina Propria Innate Lymphoid Cells in Intestinal Inflammation. Inflamm. Bowel Dis. 2018, 24, 1649–1659. [Google Scholar] [CrossRef] [Green Version]
- Lenfestey, M.W.; Neu, J. Gastrointestinal Development: Implications for Management of Preterm and Term Infants. Gastroenterol. Clin. N. Am. 2018, 47, 773–791. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.C.; Chen, C.M. Cathelicidin attenuates hyperoxia-induced intestinal injury through inhibition of NF-kappaB activity in newborn rats. Exp. Mol. Pathol. 2020, 113, 104269. [Google Scholar] [CrossRef] [PubMed]
- Giannone, P.J.; Bauer, J.A.; Schanbacher, B.L.; Reber, K.M. Effects of hyperoxia on postnatal intestinal development. Biotech. Histochem. 2007, 82, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tao, Y.; Xu, J.; He, Y.; Zhang, W.; Jiang, Z.; He, Y.; Liu, H.; Chen, M.; Zhang, W.; et al. Hyperoxia Provokes Time- and Dose-Dependent Gut Injury and Endotoxemia and Alters Gut Microbiome and Transcriptome in Mice. Front. Med. 2021, 8, 732039. [Google Scholar] [CrossRef]
- Sherman, M.P.; Bennett, S.H.; Hwang, F.F.; Sherman, J.; Bevins, C.L. Paneth cells and antibacterial host defense in neonatal small intestine. Infect. Immun. 2005, 73, 6143–6146. [Google Scholar] [CrossRef] [Green Version]
- Vaishnava, S.; Behrendt, C.L.; Ismail, A.S.; Eckmann, L.; Hooper, L.V. Paneth cells directly sense gut commensals and maintain homeostasis at the intestinal host-microbial interface. Proc. Natl. Acad. Sci. USA 2008, 105, 20858–20863. [Google Scholar] [CrossRef] [Green Version]
- Lueschow, S.R.; McElroy, S.J. The Paneth Cell: The Curator and Defender of the Immature Small Intestine. Front. Immunol. 2020, 11, 587. [Google Scholar] [CrossRef] [Green Version]
- Chou, H.C.; Chen, C.M. Neonatal hyperoxia disrupts the intestinal barrier and impairs intestinal function in rats. Exp. Mol. Pathol. 2017, 102, 415–421. [Google Scholar] [CrossRef]
- Liu, D.Y.; Li, J.J. Effect of hyperoxia on the intestinal IgA secretory component in neonatal rats and on intestinal epithelial cells in vitro. Braz. J. Med. Biol. Res. 2010, 43, 1034–1041. [Google Scholar] [CrossRef] [Green Version]
- Bruno, M.E.; Frantz, A.L.; Rogier, E.W.; Johansen, F.E.; Kaetzel, C.S. Regulation of the polymeric immunoglobulin receptor by the classical and alternative NF-κB pathways in intestinal epithelial cells. Mucosal. Immunol. 2011, 4, 468–478. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.Y.; Jiang, T.; Wang, S.; Cao, X. Effect of hyperoxia on pulmonary SIgA and its components, IgA and SC. J. Clin. Immunol. 2013, 33, 1009–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Tang, S.; Xin, J.; Liu, D. Influence of reactive oxygen species on secretory component in the intestinal epithelium during hyperoxia. Exp. Med. 2017, 14, 4033–4040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davids, B.J.; Palm, J.E.; Housley, M.P.; Smith, J.R.; Andersen, Y.S.; Martin, M.G.; Hendrickson, B.A.; Johansen, F.E.; Svärd, S.G.; Gillin, F.D.; et al. Polymeric immunoglobulin receptor in intestinal immune defense against the lumen-dwelling protozoan parasite Giardia. J. Immunol. 2006, 177, 6281–6290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.M.; Liu, D.Y. Mechanism of Neonatal Intestinal Injury Induced by Hyperoxia Therapy. J. Immunol. Res. 2022, 2022, 2316368. [Google Scholar] [CrossRef]
- Chen, C.M.; Chou, H.C. Hyperoxia disrupts the intestinal barrier in newborn rats. Exp. Mol. Pathol. 2016, 101, 44–49. [Google Scholar] [CrossRef]
- Liu, D.Y.; Lou, W.J.; Zhang, D.Y.; Sun, S.Y. ROS Plays a Role in the Neonatal Rat Intestinal Barrier Damages Induced by Hyperoxia. Biomed. Res. Int. 2020, 2020, 8819195. [Google Scholar] [CrossRef]
- Bölke, E.; Jehle, P.M.; Orth, K.; Steinbach, G.; Hannekum, A.; Storck, M. Changes of gut barrier function during anesthesia and cardiac surgery. Angiology 2001, 52, 477–482. [Google Scholar] [CrossRef]
- Lo, Y.C.; Chen, K.Y.; Chou, H.C.; Lin, I.H.; Chen, C.M. Neonatal hyperoxia induces gut dysbiosis and behavioral changes in adolescent mice. J. Chin. Med. Assoc. 2021, 84, 290–298. [Google Scholar] [CrossRef]
- Berg, R.D. Bacterial translocation from the gastrointestinal tract. Adv. Exp. Med. Biol. 1999, 473, 11–30. [Google Scholar] [CrossRef]
- Hackam, D.J.; Good, M.; Sodhi, C.P. Mechanisms of gut barrier failure in the pathogenesis of necrotizing enterocolitis: Toll-like receptors throw the switch. Semin. Pediatr. Surg. 2013, 22, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Bein, A.; Eventov-Friedman, S.; Arbell, D.; Schwartz, B. Intestinal tight junctions are severely altered in NEC preterm neonates. Pediatr. Neonatol. 2018, 59, 464–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Managlia, E.; Yan, X.; De Plaen, I.G. Intestinal Epithelial Barrier Function and Necrotizing Enterocolitis. Newborn 2022, 1, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Kubes, P.; Suzuki, M.; Granger, D.N. Nitric oxide: An endogenous modulator of leukocyte adhesion. Proc. Natl. Acad. Sci. USA 1991, 88, 4651–4655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaboury, J.; Woodman, R.C.; Granger, D.N.; Reinhardt, P.; Kubes, P. Nitric oxide prevents leukocyte adherence: Role of superoxide. Am. J. Physiol. 1993, 265, H862–H867. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.M.; Liu, C.H.; Chen, L.W. Nitric oxide synthase inhibitor ameliorates oral total parenteral nutrition-induced barrier dysfunction. Shock 2000, 13, 135–139. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.M.; Chou, H.C.; Yang, Y.S.H.; Su, E.C.; Liu, Y.R. Predicting Hyperoxia-Induced Lung Injury from Associated Intestinal and Lung Dysbiosis in Neonatal Mice. Neonatology 2021, 118, 163–173. [Google Scholar] [CrossRef]
- Di Lorenzo, M.; Krantis, A. Altered nitric oxide production in the premature gut may increase susceptibility to intestinal damage in necrotizing enterocolitis. J. Pediatr. Surg. 2001, 36, 700–705. [Google Scholar] [CrossRef]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [Green Version]
- Shea, L.M.; Beehler, C.; Schwartz, M.; Shenkar, R.; Tuder, R.; Abraham, E. Hyperoxia activates NF-kappaB and increases TNF-alpha and IFN-gamma gene expression in mouse pulmonary lymphocytes. J. Immunol. 1996, 157, 3902–3908. [Google Scholar] [CrossRef]
- Ye, Y.; Lin, P.; Zhang, W.; Tan, S.; Zhou, X.; Li, R.; Pu, Q.; Koff, J.L.; Dhasarathy, A.; Ma, F.; et al. DNA Repair Interacts with Autophagy To Regulate Inflammatory Responses to Pulmonary Hyperoxia. J. Immunol. 2017, 198, 2844–2853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Liu, F.; Li, Y.; Tang, R.; Wang, J. mRNA expression of TLR4, TLR9 and NF-κB in a neonatal murine model of necrotizing enterocolitis. Mol. Med. Rep. 2016, 14, 1953–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babu, D.; Lee, J.S.; Park, S.Y.; Thapa, D.; Choi, M.K.; Kim, A.R.; Park, Y.J.; Kim, J.A. Involvement of NF-kappaB in the inhibitory actions of Platycarya strobilacea on the TNF-alpha-induced monocyte adhesion to colon epithelial cells and chemokine expression. Arch. Pharmacal. Res. 2008, 31, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, T.; Liu, Y.; Sun, S.; Liu, D. Nuclear factor erythroid 2-related factor 2 potentiates the generation of inflammatory cytokines by intestinal epithelial cells during hyperoxia by inducing the expression of interleukin 17D. Toxicology 2021, 457, 152820. [Google Scholar] [CrossRef] [PubMed]
- Andreadou, I.; Schulz, R.; Papapetropoulos, A.; Turan, B.; Ytrehus, K.; Ferdinandy, P.; Daiber, A.; Di Lisa, F. The role of mitochondrial reactive oxygen species, NO and H2S in ischaemia/reperfusion injury and cardioprotection. J. Cell. Mol. Med. 2020, 24, 6510–6522. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Tang, S.; Xin, J.; Wei, Y.; Liu, D. Reactive oxygen species induce injury of the intestinal epithelium during hyperoxia. Int. J. Mol. Med. 2018, 41, 322–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozinsky, A.; Underhill, D.M.; Fontenot, J.D.; Hajjar, A.M.; Smith, K.D.; Wilson, C.B.; Schroeder, L.; Aderem, A. The repertoire for pattern recognition of pathogens by the innate immune system is defined by cooperation between toll-like receptors. Proc. Natl. Acad. Sci. USA 2000, 97, 13766–13771. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like Receptors and the Control of Immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef]
- Egan, C.E.; Sodhi, C.P.; Good, M.; Lin, J.; Jia, H.; Yamaguchi, Y.; Lu, P.; Ma, C.; Branca, M.F.; Weyandt, S.; et al. Toll-like receptor 4-mediated lymphocyte influx induces neonatal necrotizing enterocolitis. J. Clin. Investig. 2016, 126, 495–508. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Lu, X.; Zhang, Y. IRAK Inhibitor Protects the Intestinal Tract of Necrotizing Enterocolitis by Inhibiting the Toll-Like Receptor (TLR) Inflammatory Signaling Pathway in Rats. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 3366–3373. [Google Scholar] [CrossRef]
- Baggiolini, M. Chemotactic and inflammatory cytokines—CXC and CC proteins. Adv. Exp. Med. Biol. 1993, 351, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Fu, B.; Deng, B.; Zeng, Y.; Wan, X.; Qu, L. Overexpression of Chemokine (C-X-C) ligand 1 (CXCL1) associated with tumor progression and poor prognosis in hepatocellular carcinoma. Cancer Cell Int. 2014, 14, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyake, M.; Goodison, S.; Urquidi, V.; Gomes Giacoia, E.; Rosser, C.J. Expression of CXCL1 in human endothelial cells induces angiogenesis through the CXCR2 receptor and the ERK1/2 and EGF pathways. Lab. Investig. A J. Tech. Methods Pathol. 2013, 93, 768–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasier, A.R. The nuclear factor-kappaB-interleukin-6 signalling pathway mediating vascular inflammation. Cardiovasc. Res. 2010, 86, 211–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saddawi-Konefka, R.; Seelige, R.; Gross, E.T.; Levy, E.; Searles, S.C.; Washington, A., Jr.; Santosa, E.K.; Liu, B.; O’Sullivan, T.E.; Harismendy, O.; et al. Nrf2 Induces IL-17D to Mediate Tumor and Virus Surveillance. Cell Rep. 2016, 16, 2348–2358. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhang, D.; Cai, Q.; Liu, D.; Sun, S. Involvement of nuclear factor erythroid 2related factor 2 in neonatal intestinal interleukin17D expression in hyperoxia. Int. J. Mol. Med. 2020, 46, 1423–1432. [Google Scholar] [CrossRef]
- Li, N.; Ma, L.; Liu, X.; Shaw, L.; Li Calzi, S.; Grant, M.B.; Neu, J. Arginyl-glutamine dipeptide or docosahexaenoic acid attenuates hyperoxia-induced small intestinal injury in neonatal mice. J. Pediatr. Gastroenterol. Nutr. 2012, 54, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Polycarpou, E.; Zachaki, S.; Tsolia, M.; Papaevangelou, V.; Polycarpou, N.; Briana, D.D.; Gavrili, S.; Kostalos, C.; Kafetzis, D. Enteral L-arginine supplementation for prevention of necrotizing enterocolitis in very low birth weight neonates: A double-blind randomized pilot study of efficacy and safety. JPEN J. Parenter. Enter. Nutr. 2013, 37, 617–622. [Google Scholar] [CrossRef]
- Lu, J.; Jilling, T.; Li, D.; Caplan, M.S. Polyunsaturated fatty acid supplementation alters proinflammatory gene expression and reduces the incidence of necrotizing enterocolitis in a neonatal rat model. Pediatr. Res. 2007, 61, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Yi, H.; Yu, C.; Zhang, H.; Song, D.; Jiang, D.; Du, H.; Wang, Y. Cathelicidin-BF suppresses intestinal inflammation by inhibiting the nuclear factor-κB signaling pathway and enhancing the phagocytosis of immune cells via STAT-1 in weanling piglets. Int. Immunopharmacol. 2015, 28, 61–69. [Google Scholar] [CrossRef]
- Han, F.; Lu, Z.; Liu, Y.; Xia, X.; Zhang, H.; Wang, X.; Wang, Y. Cathelicidin-BF ameliorates lipopolysaccharide-induced intestinal epithelial barrier disruption in rat. Life Sci. 2016, 152, 199–209. [Google Scholar] [CrossRef]
- Morais, L.H.; Schreiber, H.L.t.; Mazmanian, S.K. The gut microbiota-brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Puri, K.; Taft, D.H.; Ambalavanan, N.; Schibler, K.R.; Morrow, A.L.; Kallapur, S.G. Association of Chorioamnionitis with Aberrant Neonatal Gut Colonization and Adverse Clinical Outcomes. PLoS ONE 2016, 11, e0162734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Q.; Lu, S.; Zhu, Y.; Wei, B.; Chen, Y.; Bai, F. Bacterial endotoxin-induced maternal inflammation leads to fetal intestinal injury and affects microbial colonization in the neonatal period. J. Matern. Fetal Neonatal Med. 2021, 35, 6917–6927. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Zhang, X.; Lv, A.; Fan, S.; Zhang, J. Saccharomyces boulardii modulates necrotizing enterocolitis in neonatal mice by regulating the sirtuin 1/NF-κB pathway and the intestinal microbiota. Mol. Med. Rep. 2020, 22, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.M.; Underwood, M.A. Probiotics and necrotizing enterocolitis. Semin. Pediatr. Surg. 2018, 27, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, W.; Su, Z.Y.; Kong, A.N. The complexity of the Nrf2 pathway: Beyond the antioxidant response. J. Nutr. Biochem. 2015, 26, 1401–1413. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Tu, H.; Liu, Y.; Chen, T.; Cao, J. Interleukin-17D Aggravates Sepsis by Inhibiting Macrophage Phagocytosis. Crit. Care Med. J. 2020, 48, e58–e65. [Google Scholar] [CrossRef]
- Lee, Y.; Clinton, J.; Yao, C.; Chang, S.H. Interleukin-17D Promotes Pathogenicity During Infection by Suppressing CD8 T Cell Activity. Front. Immunol. 2019, 10, 1172. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, E.H.; Suzuki, T.; Funayama, R.; Nagashima, T.; Hayashi, M.; Sekine, H.; Tanaka, N.; Moriguchi, T.; Motohashi, H.; Nakayama, K.; et al. Nrf2 suppresses macrophage inflammatory response by blocking proinflammatory cytokine transcription. Nat. Commun. 2016, 7, 11624. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Guo, Q.; Duan, X.; Xu, Z.; Wang, Q. l-arginine inhibited apoptosis of fish leukocytes via regulation of NF-κB-mediated inflammation, NO synthesis, and anti-oxidant capacity. Biochimie 2019, 158, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.S.; Shah, V.S.; Kelly, L.E. Arginine supplementation for prevention of necrotising enterocolitis in preterm infants. Cochrane Database Syst. Rev. 2017, 4, CD004339. [Google Scholar] [CrossRef] [PubMed]
- Lapillonne, A.; Jensen, C.L. Reevaluation of the DHA requirement for the premature infant. Prostaglandins Leukot. Essent. Fat. Acids 2009, 81, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Doss, M.; White, M.R.; Tecle, T.; Hartshorn, K.L. Human defensins and LL-37 in mucosal immunity. J. Leukoc. Biol. 2010, 87, 79–92. [Google Scholar] [CrossRef]
- Kåhrström, C.T.; Pariente, N.; Weiss, U. Intestinal microbiota in health and disease. Nature 2016, 535, 47. [Google Scholar] [CrossRef] [Green Version]
- Dickson, R.P. The microbiome and critical illness. Lancet. Respir. Med. 2016, 4, 59–72. [Google Scholar] [CrossRef] [Green Version]
- Humberg, A.; Fortmann, I.; Siller, B.; Kopp, M.V.; Herting, E.; Gopel, W.; Hartel, C.; German Neonatal Network, German Center for Lung Research and Priming Immunity at the beginning of life (PRIMAL) Consortium. Preterm birth and sustained inflammation: Consequences for the neonate. Semin. Immunopathol. 2020, 42, 451–468. [Google Scholar] [CrossRef]
- Ali, S.K.M.; Mohammed, N.; Qureshi, N.; Gupta, S. Oxygen therapy in preterm infants: Recommendations for practice. Paediatr. Child Health 2021, 31, 1–6. [Google Scholar] [CrossRef]
- Saugstad, O.D. Oxygenation of the Immature Infant: A Commentary and Recommendations for Oxygen Saturation Targets and Alarm Limits. Neonatology 2018, 114, 69–75. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.-C.; Chou, H.-C.; Chen, C.-M. Molecular Mechanisms of Hyperoxia-Induced Neonatal Intestinal Injury. Int. J. Mol. Sci. 2023, 24, 4366. https://doi.org/10.3390/ijms24054366
Wang H-C, Chou H-C, Chen C-M. Molecular Mechanisms of Hyperoxia-Induced Neonatal Intestinal Injury. International Journal of Molecular Sciences. 2023; 24(5):4366. https://doi.org/10.3390/ijms24054366
Chicago/Turabian StyleWang, Hsiao-Chin, Hsiu-Chu Chou, and Chung-Ming Chen. 2023. "Molecular Mechanisms of Hyperoxia-Induced Neonatal Intestinal Injury" International Journal of Molecular Sciences 24, no. 5: 4366. https://doi.org/10.3390/ijms24054366